
Evaluating Inflorescence Morphology in Two Species and Subspecies of the Genus Hierochloë R. Brown

 , , , , , ,
, , , , , , 
Abstract
1. Introduction
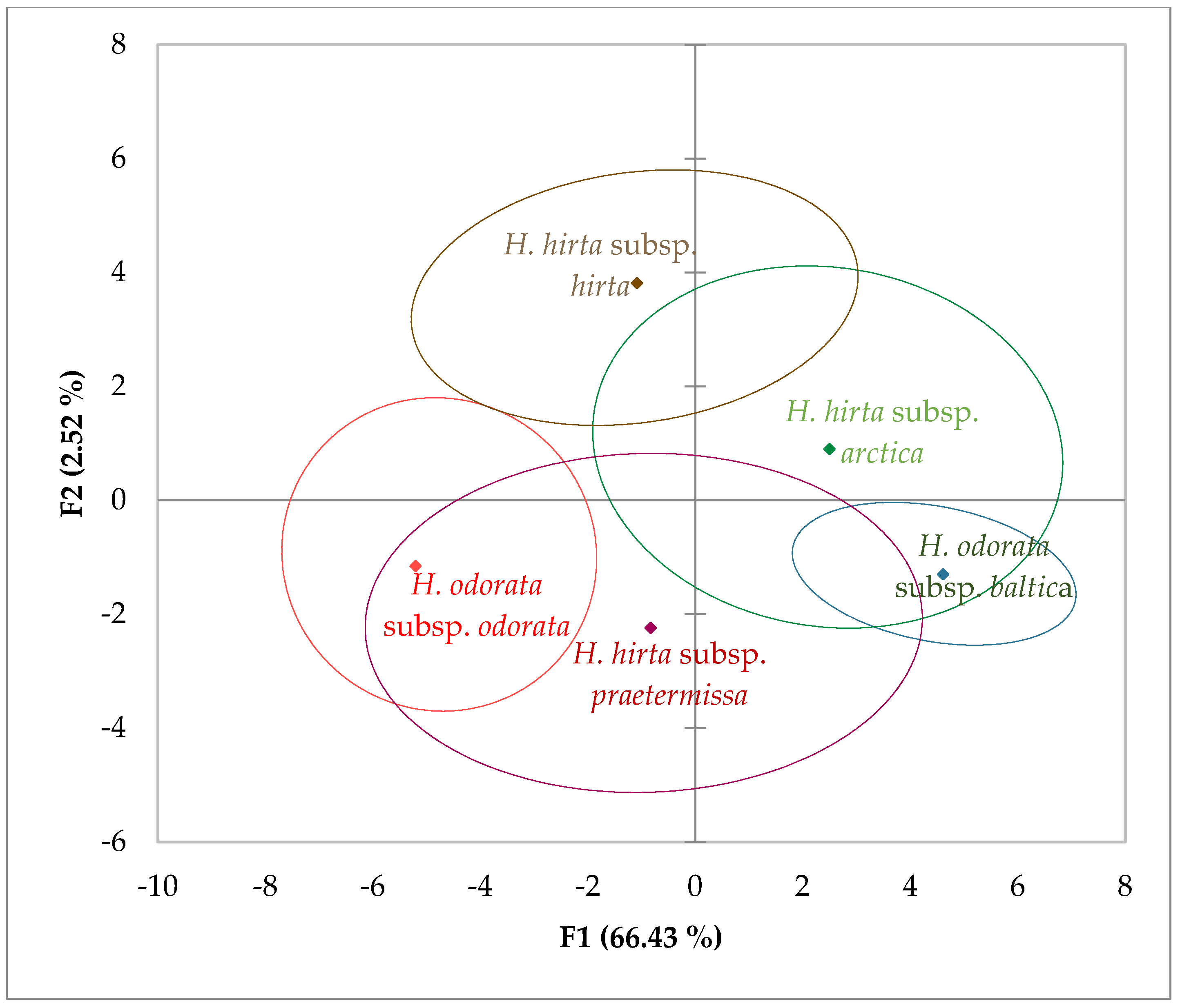
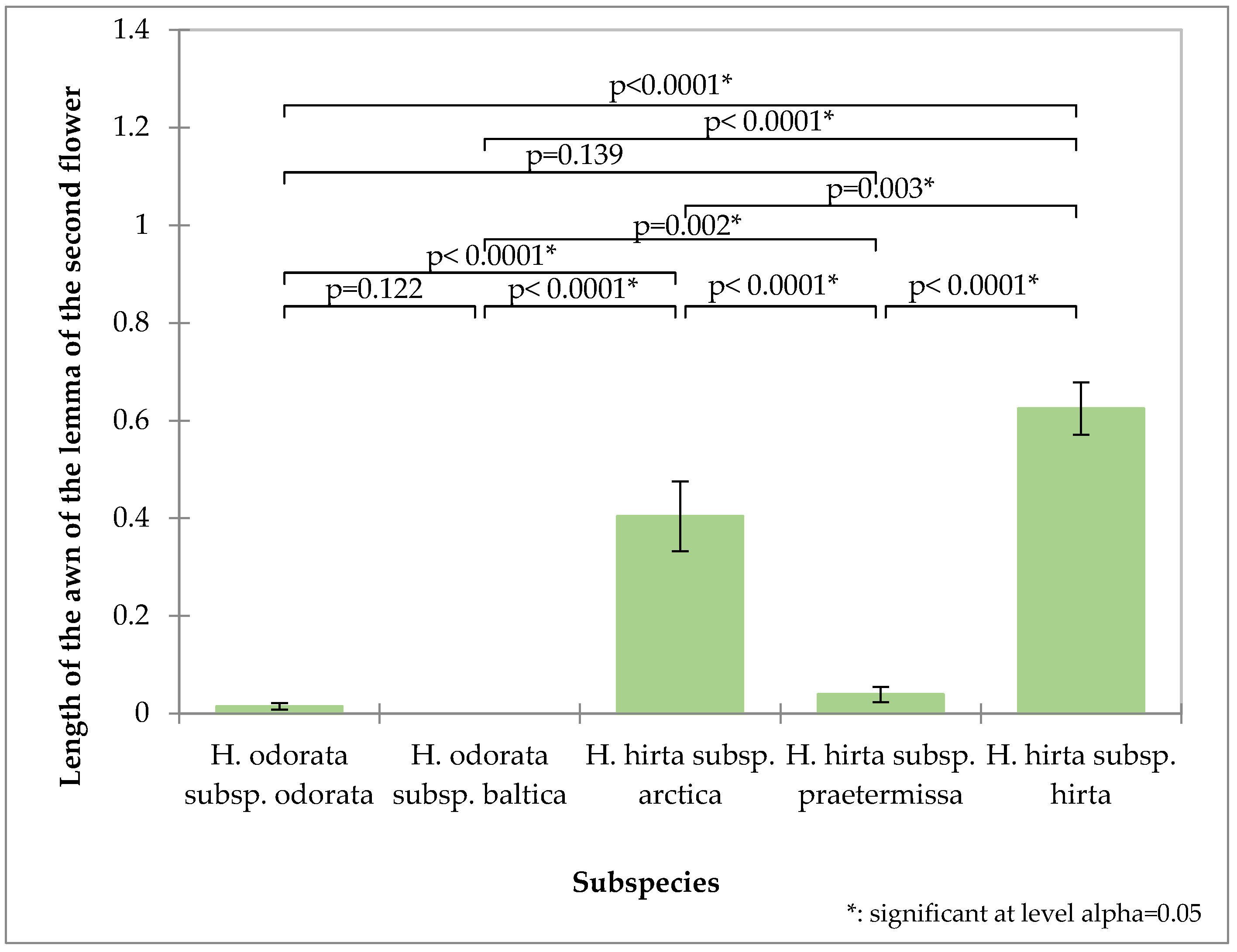
2. Results
3. Discussion
4. Materials and Methods
- Hierochloë odorata
- ○
- Hierochloë odorata (L.) Wahlenb. subsp. odorata G. Weim. (number of herbarium speciment: 9359, 9364 9395, 492192, 9365, 9363, 9361)
- ○
- Hierochloë odorata (L.) Wahlenb. subsp. baltica G. Weim. (number of herbarium speciment: 486885, 486885, 9386, 9369, 9303)
- Hierochloë hirta
- ○
- Hierochloë hirta (Schrank) Borbás subsp. praetermissa (G. Weim.) B. Walln. (number of herbarium speciment: 1221679, 1245158, 767323)
- ○
- Hierochloë hirta (Schrank) Borbás subsp. arctica (Presl) G. Weim. (number of herbarium speciment: 9393, 522684, 9383, 9384)
- ○
- Hierochloë hirta (Schrank) Borbás subsp. hirta G. Weim. (number of herbarium speciment: 9358, 570435, 505794, 487263, 487262)
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H. odorata subsp. odorata | |||||||||||||||||
| 9359 | 95 | 32 | 57 | 44 | 38 | 24 | 31 | 26 | 5.6 | 4.8 | 4.9 | 4 | 0.1 | 3.8 | 3.9 | 0.1 | 3.1 |
| 5.6 | 5.1 | 4.6 | 4.2 | 0.1 | 4.2 | 4.2 | 0.1 | 4.1 | |||||||||
| 5.1 | 4.5 | 5 | 4.1 | 0 | 3.9 | 4 | 0 | 3.6 | |||||||||
| 5.3 | 4.2 | 5.1 | 4.5 | 0.1 | 3.9 | 4.2 | 0 | 4.1 | |||||||||
| 4.8 | 4.2 | 4.4 | 4.1 | 0.1 | 3.9 | 4.1 | 0 | 3.9 | |||||||||
| 5.6 | 4.9 | 5.6 | 4.5 | 0 | 4.1 | 4.1 | 0 | 3.9 | |||||||||
| 9359 | 79 | 23 | 28 | 22 | 24 | 18 | 14 | 11 | 5.1 | 4.4 | 4.9 | 3.2 | 0.1 | 3.1 | 4.2 | 0 | 3.1 |
| 4.3 | 4.2 | 4.1 | 3.1 | 0 | 2.8 | 4.1 | 0 | 2.9 | |||||||||
| 5.1 | 4.5 | 4.7 | 3.6 | 0.1 | 3.2 | 4.3 | 0.1 | 3.3 | |||||||||
| 4.8 | 4.4 | 4.4 | 3.5 | 0 | 3.1 | 4.1 | 0 | 3.3 | |||||||||
| 5.2 | 4.2 | 5.1 | 4.3 | 0 | 3.5 | 4.1 | 0 | 3.2 | |||||||||
| 5.1 | 4.9 | 4.7 | 3.5 | 0.1 | 3.1 | 3.4 | 0.1 | 3.1 | |||||||||
| 9359 | 90 | 24 | 48 | 34 | 34 | 24 | 31 | 21 | 4.7 | 3.9 | 4.5 | 3.5 | 0 | 3.2 | 3.8 | 0 | 3.3 |
| 4.5 | 4.1 | 4.2 | 3.7 | 0.1 | 3.2 | 3.2 | 0 | 3.4 | |||||||||
| 4.1 | 3.6 | 3.9 | 3.2 | 0 | 2.6 | 3.1 | 0 | 3 | |||||||||
| 4.3 | 4 | 4.1 | 3.1 | 0.1 | 2.5 | 3 | 0 | 3 | |||||||||
| 4.5 | 4 | 4.2 | 3.6 | 0 | 3.1 | 3.1 | 0 | 3.5 | |||||||||
| 4.4 | 4.1 | 3.7 | 3.2 | 0 | 3.2 | 3.2 | 0 | 3.3 | |||||||||
| 9364 | 85 | 24 | 42 | 32 | 32 | 23 | 24 | 17 | 5.2 | 4.8 | 5.1 | 4.2 | 0 | 3.2 | 3.4 | 0 | 3.1 |
| 5.3 | 4.9 | 5.1 | 4.1 | 0 | 3.6 | 3.9 | 0 | 3.5 | |||||||||
| 5.2 | 4.9 | 5.1 | 3.7 | 0 | 3.2 | 3.6 | 0 | 3.1 | |||||||||
| 5.3 | 5 | 5 | 4 | 0.1 | 3.5 | 3.4 | 0 | 3.1 | |||||||||
| 5.2 | 4.6 | 5.1 | 4 | 0 | 3 | 3.2 | 0 | 3.1 | |||||||||
| 5.1 | 4.9 | 4.2 | 3.9 | 0.1 | 3.4 | 3.4 | 0 | 3.3 | |||||||||
| 9364 | 73 | 22 | 34 | 32 | 29 | 26 | 23 | 16 | 4.2 | 3.5 | 4 | 3.4 | 0 | 3.1 | 3.1 | 0.1 | 3.3 |
| 3.9 | 3.8 | 3.3 | 3.2 | 0 | 3.1 | 3.3 | 0 | 3 | |||||||||
| 3.9 | 3.3 | 3.6 | 3.4 | 0 | 3.3 | 3.2 | 0 | 3.2 | |||||||||
| 4.1 | 3.8 | 3.2 | 3.2 | 0.1 | 3.2 | 3.2 | 0 | 3.1 | |||||||||
| 4.1 | 3.1 | 3.9 | 3.2 | 0 | 3.1 | 3.1 | 0.1 | 3.1 | |||||||||
| 4.1 | 3.2 | 3.9 | 3.2 | 0.1 | 3.1 | 3.2 | 0 | 3.2 | |||||||||
| 9364 | 77 | 22 | 42 | 32 | 28 | 17 | 26 | 18 | 5.3 | 5 | 5.1 | 4.2 | 0.1 | 4 | 4.1 | 0 | 4 |
| 5.1 | 4.1 | 4.8 | 4 | 0.1 | 3.9 | 4 | 0 | 3.6 | |||||||||
| 5.2 | 4.4 | 5.1 | 4.2 | 0 | 3.8 | 3.9 | 0 | 3.7 | |||||||||
| 4.9 | 4.5 | 4.8 | 4.6 | 0.1 | 3.9 | 4 | 0 | 3.8 | |||||||||
| 5.2 | 5 | 4.8 | 4.1 | 0 | 3.6 | 3.8 | 0 | 3.8 | |||||||||
| 4.8 | 4.5 | 4.3 | 4 | 0.1 | 3.8 | 3.7 | 0 | 3.8 | |||||||||
| 9395 | 72 | 21 | 39 | 34 | 28 | 21 | 19 | 17 | 4 | 3.8 | 3.5 | 3.4 | 0 | 3.3 | 3.3 | 0 | 3.3 |
| 4.2 | 4 | 4.1 | 3.4 | 0.1 | 3.2 | 3.1 | 0 | 3.1 | |||||||||
| 4.2 | 3.8 | 3.9 | 3.4 | 0.1 | 3.2 | 3 | 0 | 3.1 | |||||||||
| 4.4 | 3.4 | 4.2 | 3.5 | 0 | 3.2 | 3.1 | 0 | 3.2 | |||||||||
| 4.2 | 3.4 | 4.1 | 3.3 | 0.2 | 3.4 | 3.2 | 0.1 | 3.4 | |||||||||
| 4.2 | 3.2 | 4.1 | 3.2 | 0 | 3.1 | 3.2 | 0 | 2.7 | |||||||||
| 9395 | 104 | 33 | 50 | - | 38 | 22 | 33 | 20 | 6.2 | 5.5 | 6.1 | 4.9 | 0 | 4.1 | 4.7 | 0 | 4.2 |
| 6.2 | 6 | 6.1 | 4.9 | 0 | 4.6 | 4.7 | 0 | 4.1 | |||||||||
| 6.1 | 5.9 | 6 | 4.8 | 0.2 | 4.2 | 4.7 | 0.1 | 4.4 | |||||||||
| 6.2 | 6.1 | 6.1 | 3.9 | 0 | 3.3 | 4.8 | 0 | 4.3 | |||||||||
| 6.2 | 6 | 6.1 | 5.1 | 0.1 | 4.3 | 4.8 | 0.1 | 4.3 | |||||||||
| 6.1 | 5.9 | 6.1 | 4.9 | 0 | 4.5 | 4.7 | 0 | 4.3 | |||||||||
| 492192 | 82 | 28 | 34 | - | 24 | 19 | 23 | 17 | 4.1 | 4 | 3.8 | 3.7 | 0 | 3.2 | 3.4 | 0 | 3.1 |
| 4.1 | 3.9 | 4 | 3.8 | 0.2 | 3.1 | 3.6 | 0.1 | 3.2 | |||||||||
| 4.2 | 4.1 | 4.1 | 3.8 | 0.1 | 3.2 | 3.6 | 0 | 3.1 | |||||||||
| 4.3 | 4.1 | 4.2 | 3.6 | 0 | 3.2 | 3.6 | 0 | 3.2 | |||||||||
| 4.1 | 3.2 | 4 | 3.7 | 0.2 | 3.1 | 3.4 | 0 | 3.1 | |||||||||
| 4.3 | 4.1 | 4.2 | 3.5 | 0 | 3.4 | 3.5 | 0.1 | 3.2 | |||||||||
| 492192 | 68 | 17 | 26 | 24 | 27 | 17 | 18 | 16 | 4.2 | 3.9 | 4.1 | 3.4 | 0.1 | 3.4 | 3.4 | 0 | 3.2 |
| 4.3 | 4 | 4.2 | 3.9 | 0.1 | 3.5 | 3.7 | 0 | 3.4 | |||||||||
| 4.6 | 4.2 | 4.5 | 4 | 0.2 | 3.3 | 3.5 | 0.1 | 3.6 | |||||||||
| 4.5 | 4.2 | 4.4 | 4 | 0 | 3.5 | 3.6 | 0 | 3.4 | |||||||||
| 4.8 | 4.3 | 4.7 | 3.8 | 0.1 | 3.4 | 3.7 | 0 | 3.5 | |||||||||
| 4.6 | 4.1 | 4.4 | 3.9 | 0.2 | 3.3 | 3.6 | 0.1 | 3.4 | |||||||||
| 9365 | 58 | 18 | 38 | 17 | 19 | 10 | 12 | 9 | 4.2 | 4.1 | 4.1 | 4 | 0.1 | 3.6 | 3.7 | 0 | 3.5 |
| 5.1 | 4.4 | 4.9 | 4 | 0.1 | 3.8 | 4 | 0.1 | 3.9 | |||||||||
| 4.6 | 4.5 | 4.3 | 3.9 | 0.2 | 3.8 | 4 | 0.1 | 3.6 | |||||||||
| 4.6 | 4.2 | 4.5 | 3.9 | 0 | 3.5 | 3.7 | 0 | 3.6 | |||||||||
| 4.3 | 4 | 4.2 | 4 | 0 | 3.6 | 3.5 | 0 | 3.4 | |||||||||
| 4.1 | 3.9 | 4 | 3.8 | 0.1 | 3.2 | 3.6 | 0 | 3.1 | |||||||||
| 9365 | 67 | 21 | 35 | 23 | 24 | 14 | 24 | 13 | 4.4 | 4 | 4.3 | 4 | 0 | 3.9 | 4 | 0 | 3.8 |
| 4.5 | 4.1 | 4.4 | 4.3 | 0 | 4.1 | 3.9 | 0 | 3.8 | |||||||||
| 4.5 | 4.1 | 4.3 | 4.1 | 0 | 4 | 3.8 | 0 | 3.8 | |||||||||
| 4.3 | 4.1 | 4.2 | 4.1 | 0.1 | 3.8 | 3.9 | 0 | 4 | |||||||||
| 4.2 | 4.1 | 4.1 | 3.8 | 0 | 3.5 | 3.7 | 0 | 3.8 | |||||||||
| 4.3 | 3.9 | 4.2 | 4 | 0 | 3.8 | 3.6 | 0 | 3.6 | |||||||||
| 9363 | 85 | 24 | 48 | 32 | 35 | 24 | 22 | 18 | 4.5 | 4.4 | 4.3 | 4 | 0.1 | 3.8 | 3.7 | 0.1 | 3.8 |
| 5.2 | 4.8 | 5.1 | 4 | 0.1 | 4.1 | 4 | 0.1 | 3.6 | |||||||||
| 5.1 | 4.8 | 4.9 | 4 | 0.1 | 3.6 | 4 | 0 | 3.3 | |||||||||
| 5.3 | 5.1 | 5.2 | 4 | 0 | 3.4 | 3.9 | 0 | 3.6 | |||||||||
| 5.3 | 3.9 | 5.2 | 4 | 0.2 | 3.5 | 3.6 | 0 | 3.4 | |||||||||
| 4.8 | 4.4 | 4.7 | 3.9 | 0.1 | 3.5 | 3.9 | 0 | 3.6 | |||||||||
| 9363 | 69 | 23 | 33 | 25 | 22 | 12 | 25 | 14 | 4.5 | 4.1 | 4.3 | 4 | 0 | 3.9 | 4 | 0 | 3.8 |
| 4.7 | 4.1 | 4.4 | 4.3 | 0 | 4.1 | 3.9 | 0 | 3.8 | |||||||||
| 4.5 | 4.1 | 4.3 | 4.1 | 0 | 4 | 3.8 | 0 | 3.9 | |||||||||
| 4.4 | 4.1 | 4.3 | 4.2 | 0 | 3.8 | 3.9 | 0 | 3.9 | |||||||||
| 4.2 | 4.1 | 4.1 | 3.8 | 0 | 3.5 | 3.7 | 0 | 3.8 | |||||||||
| 4.3 | 3.9 | 4.2 | 4 | 0 | 3.8 | 3.6 | 0 | 3.6 | |||||||||
| 9361 | 98 | 33 | 46 | 26 | 24 | 23 | 19 | 14 | 4.5 | 4.3 | 3.9 | 3.9 | 0.1 | 4 | 3.1 | 0 | 3.5 |
| 5.3 | 5.1 | 5.2 | 4 | 0.1 | 3.4 | 3.9 | 0 | 3.6 | |||||||||
| 4.8 | 4.3 | 4.7 | 3.8 | 0.1 | 3.4 | 3.7 | 0 | 3.5 | |||||||||
| 4.2 | 4.1 | 4.1 | 3.8 | 0 | 3.5 | 3.7 | 0 | 3.8 | |||||||||
| 4.5 | 4.3 | 3.9 | 3.8 | 0.1 | 4 | 0.3 | 0 | 3.6 | |||||||||
| 4.6 | 4.1 | 4.4 | 3.9 | 0.1 | 3.3 | 3.6 | 0 | 3.4 | |||||||||
| H. odorata subsp. baltica | |||||||||||||||||
| 1015766 | 55 | 19 | 32 | 24 | 24 | 17 | 19 | 14 | 5.3 | 4.8 | 5.2 | 4.6 | 0 | 4.3 | 4.6 | 0 | 4.2 |
| 5.2 | 4.9 | 5.1 | 4.9 | 0 | 4 | 4.5 | 0 | 4.1 | |||||||||
| 5 | 4.1 | 4.8 | 4.3 | 0 | 4.1 | 4 | 0 | 4.2 | |||||||||
| 4.9 | 4.6 | 4.8 | 4.4 | 0 | 3.9 | 4.1 | 0 | 4 | |||||||||
| 5.1 | 4.8 | 5 | 4.1 | 0 | 4.1 | 3.8 | 0 | 3.6 | |||||||||
| 5.1 | 4.5 | 4.9 | 4.3 | 0 | 4.1 | 4.1 | 0 | 4.2 | |||||||||
| 1015766 | 67 | 18 | 31 | 24 | 28 | 19 | 22 | 16 | 6.2 | 5.1 | 6.1 | 4.5 | 0.1 | 4 | 4.3 | 0 | 4.2 |
| 6.3 | 5.6 | 6.2 | 4.8 | 0.1 | 4.1 | 4.3 | 0 | 4.1 | |||||||||
| 6.2 | 5.2 | 6.1 | 4.6 | 0 | 4 | 4.1 | 0 | 4 | |||||||||
| 5.8 | 5.6 | 5.6 | 4.5 | 0 | 4.2 | 4.2 | 0 | 4.2 | |||||||||
| 6.1 | 4.9 | 6 | 4.6 | 0 | 4 | 4.1 | 0 | 4 | |||||||||
| 5.5 | 4.8 | 5.4 | 4.4 | 0 | 4.1 | 4.1 | 0 | 3.6 | |||||||||
| 1015766 | 48 | 18 | 29 | 24 | 21 | 17 | 16 | 11 | 5 | 4.2 | 4.9 | 4.5 | 0 | 3.9 | 4.4 | 0 | 4.4 |
| 5.1 | 4.8 | 5 | 4.4 | 0 | 4 | 4.1 | 0 | 4.1 | |||||||||
| 4.6 | 4.2 | 4.5 | 4.2 | 0 | 3.8 | 4 | 0 | 3.7 | |||||||||
| 5.1 | 4.7 | 5 | 4.8 | 0 | 4 | 4.2 | 0 | 4.1 | |||||||||
| 5.1 | 4.7 | 4.9 | 4.6 | 0 | 4 | 4.5 | 0 | 4 | |||||||||
| 5.1 | 4.2 | 4.7 | 4.8 | 0 | 4 | 4.2 | 0 | 4.1 | |||||||||
| 486885 | 42 | 17 | 26 | 19 | 18 | 13 | 12 | 9 | 5.2 | 5.1 | 5.1 | 4.9 | 0 | 4.2 | 4.1 | 0 | 4 |
| 5.3 | 4.5 | 5.1 | 4.6 | 0 | 4 | 4.1 | 0 | 4.1 | |||||||||
| 5.2 | 4.8 | 5.1 | 4.7 | 0 | 3.8 | 4.2 | 0 | 4 | |||||||||
| 5.2 | 5 | 5.1 | 4.7 | 0 | 4.1 | 4.2 | 0 | 4 | |||||||||
| 5.1 | 4.8 | 5 | 4.3 | 0 | 3.8 | 3.8 | 0 | 3.6 | |||||||||
| 5.2 | 4.8 | 4.9 | 4.6 | 0 | 3.9 | 4.4 | 0 | 4 | |||||||||
| 486885 | 72 | 19 | 28 | 27 | 28 | 16 | 22 | 16 | 6.1 | 5 | 5.8 | 5.1 | 0 | 4.3 | 4.6 | 0 | 4.4 |
| 6.1 | 5 | 5.9 | 4.9 | 0 | 4.1 | 4.3 | 0 | 4.1 | |||||||||
| 6.2 | 5 | 6 | 5.1 | 0 | 4.4 | 4.7 | 0 | 4.2 | |||||||||
| 5.8 | 5.6 | 5.4 | 4.9 | 0 | 4.3 | 4.5 | 0 | 3.9 | |||||||||
| 6 | 5.8 | 5.9 | 5.2 | 0 | 4.2 | 4.7 | 0 | 4.3 | |||||||||
| 5.1 | 4.9 | 5 | 4.3 | 0 | 3.9 | 3.9 | 0 | 4 | |||||||||
| 486885 | 61 | 16 | 35 | 32 | 24 | 16 | 21 | 17 | 5.1 | 5 | 4.5 | 4.5 | 0 | 3.9 | 3.8 | 0 | 3.7 |
| 6.1 | 5.2 | 5.9 | 4.9 | 0 | 4.6 | 4.7 | 0 | 4.4 | |||||||||
| 5.2 | 5 | 5.1 | 4.6 | 0 | 4.1 | 4.1 | 0 | 4 | |||||||||
| 5.8 | 5.5 | 5.7 | 5.2 | 0 | 4.2 | 4.7 | 0 | 4.4 | |||||||||
| 6 | 5.1 | 5.9 | 4.7 | 0 | 4.1 | 4.1 | 0 | 4.1 | |||||||||
| 5.4 | 4.8 | 5.3 | 4.8 | 0 | 3.9 | 4.3 | 0 | 4.1 | |||||||||
| 9395 | 68 | 19 | 26 | - | 24 | 22 | 19 | 12 | 5.6 | 5 | 5.5 | 4.8 | 0 | 4 | 4.2 | 0 | 4.1 |
| 5.1 | 4.7 | 5 | 4.1 | 0 | 4.2 | 4.3 | 0 | 4 | |||||||||
| 5.9 | 5 | 5.8 | 4.7 | 0 | 4.3 | 4.3 | 0 | 3.5 | |||||||||
| 6.1 | 5.2 | 6 | 5 | 0 | 4.8 | 4.6 | 0 | 4.3 | |||||||||
| 5.1 | 4.8 | 5.1 | 4.6 | 0 | 4.5 | 4 | 0 | 3.9 | |||||||||
| 5.7 | 5.2 | 5.6 | 5 | 0 | 4.4 | 4.5 | 0 | 4.5 | |||||||||
| 9395 | 73 | 20 | 36 | 32 | 31 | 21 | 19 | 16 | 5.2 | 5 | 5.1 | 4.8 | 0 | 4.1 | 4.1 | 0 | 4.1 |
| 5.1 | 5 | 4.7 | 4.6 | 0 | 4 | 4 | 0 | 4 | |||||||||
| 5.3 | 5.1 | 5.2 | 4.7 | 0 | 4 | 4.1 | 0 | 3.9 | |||||||||
| 5.5 | 5 | 5.4 | 4.7 | 0 | 4.4 | 4.1 | 0 | 4.3 | |||||||||
| 6.1 | 5.5 | 5.8 | 4.9 | 0 | 3.9 | 4.3 | 0 | 4.1 | |||||||||
| 5.5 | 5 | 5.4 | 4.6 | 0 | 4 | 4.3 | 0 | 4.5 | |||||||||
| 9395 | 61 | 14 | 31 | 25 | 21 | 18 | 20 | 13 | 5.3 | 5.1 | 5.2 | 4.8 | 0 | 4.1 | 4.3 | 0 | 4 |
| 5.1 | 4.8 | 4.9 | 4.1 | 0 | 3.9 | 4.1 | 0 | 4.2 | |||||||||
| 5 | 4.8 | 4.9 | 4.7 | 0 | 4 | 4.2 | 0 | 3.9 | |||||||||
| 4.8 | 4.3 | 4.7 | 4.1 | 0 | 3.8 | 3.9 | 0 | 4.1 | |||||||||
| 5.1 | 5 | 5 | 4.6 | 0 | 3.9 | 4.2 | 0 | 4.1 | |||||||||
| 5.2 | 4.9 | 5.1 | 4.6 | 0 | 4 | 4.1 | 0 | 3.9 | |||||||||
| 9386 | 62 | 16 | 35 | 21 | 25 | 18 | 17 | 15 | 5.4 | 5.1 | 5.3 | 4.7 | 0 | 3.9 | 4.1 | 0 | 3.9 |
| 5.2 | 4.9 | 5.1 | 4.7 | 0 | 3.8 | 4.2 | 0 | 3.9 | |||||||||
| 5.7 | 5.2 | 5.6 | 4.6 | 0 | 4.3 | 4.1 | 0 | 4.4 | |||||||||
| 5.4 | 5.1 | 5.2 | 4.6 | 0 | 3.9 | 4.2 | 0 | 4.1 | |||||||||
| 5.4 | 4.9 | 5.3 | 4.7 | 0 | 4.2 | 4.2 | 0 | 4.3 | |||||||||
| 5.5 | 5.1 | 5.4 | 4.7 | 0 | 3.9 | 4.1 | 0 | 4.4 | |||||||||
| 9386 | 53 | 19 | 31 | 21 | 24 | 17 | 19 | 15 | 6.1 | 5.8 | 5.9 | 5.2 | 0 | 2.5 | 4.6 | 0 | 4.5 |
| 6.8 | 6.3 | 6.4 | 5.6 | 0 | 4.3 | 5.2 | 0 | 4.5 | |||||||||
| 6.1 | 5.8 | 5.9 | 5.1 | 0 | 4.3 | 4.8 | 0 | 4.3 | |||||||||
| 6.2 | 6.1 | 5.7 | 5.2 | 0 | 4.8 | 4.5 | 0 | 4.2 | |||||||||
| 6.4 | 6.1 | 6.3 | 5.2 | 0 | 4.7 | 4.8 | 0 | 4.4 | |||||||||
| 6.5 | 6.1 | 6.2 | 5.7 | 0 | 4.6 | 5.1 | 0 | 4.5 | |||||||||
| 9369 | 52 | 16 | 34 | 26 | 25 | 18 | 15 | 11 | 5.9 | 5.6 | 5.8 | 4.9 | 0 | 4.3 | 4.7 | 0 | 4.2 |
| 5.9 | 5.1 | 5.8 | 5 | 0 | 4.2 | 4.8 | 0 | 4.5 | |||||||||
| 5.5 | 5 | 5.4 | 5.1 | 0 | 4.3 | 4.8 | 0 | 4.6 | |||||||||
| 6.2 | 5.3 | 6.1 | 5.2 | 0 | 4.4 | 4.8 | 0 | 4.3 | |||||||||
| 5.7 | 5.6 | 5.6 | 5.3 | 0 | 4.2 | 4.6 | 0 | 4.5 | |||||||||
| 6.1 | 5.8 | 6.1 | 5 | 0 | 4.4 | 4.7 | 0 | 4.3 | |||||||||
| 9369 | 63 | 16 | 34 | 26 | 24 | 19 | 21 | 14 | 5.3 | 5.1 | 5.2 | 4.8 | 0 | 4.1 | 4.3 | 0 | 4 |
| 5.1 | 4.8 | 4.9 | 4.1 | 0 | 4 | 4.4 | 0 | 4.2 | |||||||||
| 5.2 | 4.6 | 5.1 | 4.7 | 0 | 4 | 4.2 | 0 | 3.9 | |||||||||
| 4.8 | 4.7 | 4.7 | 4.1 | 0 | 3.8 | 4.1 | 0 | 4.1 | |||||||||
| 5.2 | 4.8 | 5.1 | 4.6 | 0 | 3.8 | 4.2 | 0 | 4.1 | |||||||||
| 5.2 | 4.9 | 5.1 | 4.6 | 0 | 4 | 4.1 | 0 | 3.9 | |||||||||
| 9303 | 61 | 19 | 31 | 24 | 24 | 22 | 16 | 14 | 6.2 | 5.6 | 6.1 | 5.1 | 0 | 2.5 | 4.6 | 0 | 4.5 |
| 5.8 | 5.6 | 5.8 | 4.9 | 0 | 4.3 | 4.7 | 0 | 4.2 | |||||||||
| 5.4 | 5.1 | 5.3 | 5.1 | 0 | 4.2 | 4.7 | 0 | 4.6 | |||||||||
| 5.5 | 5.2 | 5.4 | 5.2 | 0 | 4.2 | 4.6 | 0 | 4.5 | |||||||||
| 5.5 | 5 | 5.4 | 5.1 | 0 | 4.3 | 4.8 | 0 | 4.6 | |||||||||
| 5.9 | 5.6 | 5.8 | 4.9 | 0 | 4.3 | 4.7 | 0 | 4.2 | |||||||||
| 9303 | 59 | 19 | 36 | 28 | 27 | 21 | 21 | 18 | 5.9 | 5.6 | 5.8 | 5.2 | 0 | 4.2 | 4.6 | 0 | 4.5 |
| 5.8 | 5.5 | 5.7 | 4.9 | 0 | 4.3 | 4.7 | 0 | 4.2 | |||||||||
| 5.4 | 5.1 | 5.3 | 5.2 | 0 | 4.2 | 4.6 | 0 | 4.5 | |||||||||
| 5.6 | 5.2 | 5.5 | 5.2 | 0 | 4.2 | 4.7 | 0 | 4.5 | |||||||||
| 5.3 | 5.1 | 5.2 | 5.2 | 0 | 4.2 | 4.5 | 0 | 4.3 | |||||||||
| 5.5 | 5.1 | 5.4 | 5.1 | 0 | 4.3 | 4.7 | 0 | 4.5 | |||||||||
| H. hirta subsp. arctica | |||||||||||||||||
| 9358 | 50 | 17 | 28 | 20 | 18 | 17 | 14 | 13 | 4.9 | 4.1 | 4.8 | 3.8 | 0.4 | 3.5 | 3.8 | 0.2 | 3.1 |
| 4.7 | 4.1 | 4.6 | 4 | 0.6 | 3.1 | 3.7 | 0.2 | 3.5 | |||||||||
| 4.6 | 4.1 | 4.5 | 3.9 | 0.4 | 3.4 | 3.7 | 0.2 | 3.6 | |||||||||
| 4.7 | 4.1 | 4.6 | 3.7 | 0.6 | 3.1 | 3.3 | 0.4 | 3.2 | |||||||||
| 5.1 | 4 | 5 | 3.8 | 0.1 | 3.6 | 3.5 | 0 | 3.5 | |||||||||
| 4.9 | 4.2 | 4.8 | 3.8 | 0.6 | 3.1 | 3.3 | 0.6 | 3.2 | |||||||||
| 9358 | 55 | 15 | 40 | 29 | 33 | 22 | 17 | 14 | 5.3 | 4.9 | 5.2 | 4.2 | 0.2 | 3.9 | 4 | 0.1 | 3.9 |
| 5.2 | 5.1 | 5 | 4.1 | 0.3 | 4 | 4.1 | 0.2 | 3.9 | |||||||||
| 5.1 | 4.9 | 5 | 4.1 | 0.1 | 3.5 | 3.9 | 0.1 | 3.8 | |||||||||
| 4.8 | 4.5 | 4.9 | 4 | 0.2 | 4 | 3.8 | 0.1 | 4 | |||||||||
| 5.1 | 4.1 | 5 | 4 | 0.4 | 3.8 | 3.6 | 0.4 | 3.7 | |||||||||
| 5.1 | 4.6 | 5 | 4.1 | 0.5 | 3.6 | 3.9 | 0.3 | 3.7 | |||||||||
| 9358 | 75 | 22 | 36 | 32 | 33 | 18 | 26 | 24 | 5 | 4.5 | 4.9 | 4 | 0.3 | 3.2 | 4.2 | 0.2 | 3.8 |
| 5 | 4.2 | 4.9 | 3.9 | 0.2 | 3.9 | 4 | 0.1 | 3.6 | |||||||||
| 5.1 | 4.3 | 5 | 4 | 0.1 | 4 | 3.8 | 0 | 3.9 | |||||||||
| 5.2 | 5 | 5.1 | 4.1 | 0.2 | 4.6 | 4 | 0.1 | 3.5 | |||||||||
| 4.9 | 4.6 | 4.9 | 3.9 | 0.2 | 3.4 | 3.4 | 0.1 | 3.5 | |||||||||
| 5 | 4.1 | 4.8 | 4 | 0.1 | 3.5 | 4.1 | 0 | 3.3 | |||||||||
| 9393 | 70 | 20 | 45 | 40 | 38 | 35 | 25 | 18 | 6.2 | 6.1 | 6 | 5.1 | 0.2 | 4.2 | 4.9 | 0.1 | 4 |
| 6.3 | 4.8 | 6.2 | 5 | 0.1 | 4.6 | 5 | 0 | 4.7 | |||||||||
| 6.4 | 6.4 | 6 | 5.3 | 0.4 | 5 | 4.8 | 0.2 | 5 | |||||||||
| 6.5 | 6.2 | 6.5 | 5.4 | 0.4 | 4.6 | 5.1 | 0.3 | 4.4 | |||||||||
| 6.4 | 5.9 | 6.3 | 5.2 | 0.2 | 4.3 | 4.9 | 0.1 | 3.9 | |||||||||
| 6.2 | 5.6 | 6.1 | 5.4 | 0.1 | 4.2 | 5.1 | 0 | 4.4 | |||||||||
| 9393 | 75 | 21 | 45 | 25 | 30 | 21 | 28 | 18 | 6.2 | 5.8 | 6.1 | 5.1 | 0.1 | 4.8 | 5.1 | 0.2 | 4.8 |
| 6.2 | 5.6 | 6.1 | 5.4 | 0.1 | 4.2 | 5.1 | 0 | 4.4 | |||||||||
| 6.6 | 6.9 | 6.2 | 5.1 | 0.4 | 4.2 | 4.8 | 0.1 | 4.8 | |||||||||
| 6.8 | 6.7 | 6.8 | 5.5 | 0.1 | 4.8 | 5 | 0.1 | 4.6 | |||||||||
| 6.3 | 6 | 6.2 | 5.4 | 0.6 | 4.5 | 4.8 | 0.3 | 4.5 | |||||||||
| 6.4 | 6.2 | 6.3 | 5.5 | 0.3 | 5 | 5.1 | 0.1 | 5.1 | |||||||||
| 9393 | 43 | 13 | 19 | 16 | 20 | 16 | 12 | 11 | 5.2 | 5 | 5.1 | 4.8 | 0.2 | 3.7 | 4 | 0.1 | 3.4 |
| 5.1 | 4.8 | 5 | 4.2 | 0.2 | 3.9 | 4.1 | 0.1 | 3.8 | |||||||||
| 5 | 4.9 | 4.9 | 4.3 | 0.1 | 3.9 | 4.1 | 0 | 3.9 | |||||||||
| 4.6 | 4.2 | 4.5 | 4.3 | 0.2 | 3.7 | 4 | 0.1 | 3.8 | |||||||||
| 4.9 | 4.5 | 4.8 | 4.2 | 0.2 | 3.5 | 4.1 | 0.2 | 3.8 | |||||||||
| 6.6 | 6.9 | 6.2 | 5.1 | 0.4 | 4.2 | 4.8 | 0.1 | 4.8 | |||||||||
| 522684 | 60 | 18 | 30 | 20 | 30 | 28 | 12 | 11 | 5.3 | 5 | 5.2 | 4.4 | 0.7 | 4.1 | 3.9 | 0.6 | 3.8 |
| 5.8 | 5.2 | 5.7 | 4.3 | 0.5 | 4 | 4 | 0.3 | 3.9 | |||||||||
| 5.4 | 5.1 | 5.3 | 4.7 | 0.8 | 4 | 4 | 0.7 | 3.8 | |||||||||
| 5.5 | 4.9 | 5.4 | 4.2 | 0.8 | 3.7 | 4 | 0.6 | 3.8 | |||||||||
| 5.2 | 4.8 | 5.1 | 3.9 | 0.8 | 3.5 | 3.9 | 1 | 3.7 | |||||||||
| 5.9 | 5.1 | 5.8 | 4.3 | 0.5 | 3.8 | 4.2 | 1 | 3.8 | |||||||||
| 522684 | 90 | 27 | 42 | 30 | 28 | 18 | 26 | 18 | 6.1 | 5.7 | 6 | 5 | 0.7 | 4.1 | 4 | 0.8 | 4.4 |
| 5.2 | 4.9 | 5.1 | 4.8 | 0.6 | 4 | 4.2 | 1.1 | 4.1 | |||||||||
| 5.2 | 5 | 5.1 | 5 | 0.4 | 4.3 | 4.2 | 0.4 | 4.1 | |||||||||
| 5.3 | 4.8 | 5.2 | 4.2 | 0.4 | 4.1 | 4.5 | 0.8 | 4.2 | |||||||||
| 5.2 | 5 | 5 | 4.8 | 0.5 | 4.3 | 4.7 | 0.9 | 4.3 | |||||||||
| 5.2 | 4.5 | 5.1 | 4.7 | 0.2 | 3.8 | 4.5 | 0.8 | 4.1 | |||||||||
| 522684 | 95 | 26 | 45 | 30 | 30 | 20 | 20 | 19 | 5.3 | 4.7 | 5.2 | 4.8 | 0.7 | 4.2 | 4.3 | 0.9 | 4.1 |
| 5.3 | 4.9 | 5.1 | 4.7 | 0.6 | 4.1 | 4.9 | 1 | 4.2 | |||||||||
| 5.6 | 5 | 5.5 | 5 | 0.9 | 4 | 4.8 | 1 | 4.1 | |||||||||
| 6 | 5.4 | 5.9 | 4.9 | 0.7 | 4.1 | 4.7 | 1 | 4.1 | |||||||||
| 5.5 | 5 | 5.4 | 4.7 | 0.6 | 4.1 | 4.4 | 0.5 | 4.2 | |||||||||
| 5.5 | 5.2 | 5.4 | 5 | 1 | 4.3 | 4.9 | 1 | 4.2 | |||||||||
| 9383 | 51 | 15 | 35 | 23 | 19 | 18 | 15 | 13 | 5.5 | 5 | 5.4 | 4.5 | 0.9 | 3.8 | 4.2 | 0.6 | 3.8 |
| 5 | 4.8 | 4.9 | 4.6 | 0.6 | 4.1 | 4.5 | 0.3 | 3.9 | |||||||||
| 5.6 | 4.3 | 5.5 | 4.4 | 0.2 | 4.1 | 3.9 | 0 | 3.8 | |||||||||
| 5.9 | 5.3 | 5.8 | 4.5 | 0.8 | 3.5 | 4.2 | 0.7 | 3.9 | |||||||||
| 5.2 | 4.6 | 5.1 | 4.2 | 0.4 | 3.7 | 3.9 | 1 | 3.8 | |||||||||
| 5.7 | 4.9 | 5.6 | 4.2 | 0.6 | 3.8 | 4.3 | 0.6 | 4 | |||||||||
| 9383 | 65 | 20 | 28 | 24 | 20 | 17 | 19 | 15 | 5.3 | 5.2 | 5.2 | 4.1 | 0.3 | 3.6 | 4.1 | 0.2 | 3.7 |
| 6 | 5.1 | 5.9 | 4.5 | 0.1 | 4 | 4 | 0.1 | 3.7 | |||||||||
| 6.1 | 5.5 | 6 | 4.9 | 0.1 | 3.9 | 4.3 | 0.6 | 4 | |||||||||
| 6 | 4.9 | 5.9 | 4.5 | 0.1 | 4 | 4 | 0.1 | 3.9 | |||||||||
| 5.8 | 3.7 | 5.7 | 4.1 | 0.4 | 3.6 | 4.1 | 0.6 | 3.8 | |||||||||
| 5.5 | 4.8 | 5.4 | 4.1 | 0.4 | 3.8 | 4.1 | 0.7 | 3.8 | |||||||||
| 9383 | 50 | 16 | 28 | 20 | 20 | 16 | 16 | 15 | 5.2 | 5 | 5.1 | 4.3 | 0.1 | 3.9 | 4.1 | 0.6 | 4 |
| 5.3 | 5 | 5.2 | 4.3 | 0 | 3.8 | 4 | 0.1 | 3.9 | |||||||||
| 5.2 | 4.7 | 5.1 | 4.1 | 0.3 | 3.7 | 3.9 | 0.1 | 3.9 | |||||||||
| 5.2 | 4.9 | 5.1 | 4.1 | 0.3 | 3.9 | 4 | 0.1 | 3.7 | |||||||||
| 5.9 | 4.7 | 5.8 | 4.1 | 0.1 | 3.5 | 4.1 | 0.1 | 3.8 | |||||||||
| 5.6 | 4.4 | 5.5 | 4.3 | 0.2 | 4.1 | 3.9 | 0.1 | 3.8 | |||||||||
| 9384 | 56 | 18 | 36 | 25 | 21 | 18 | 16 | 14 | 5.6 | 5.2 | 5.4 | 4.5 | 0.9 | 3.8 | 4.2 | 0.6 | 3.8 |
| 5.2 | 4.8 | 5.1 | 4.6 | 0.6 | 4.2 | 4.5 | 0.5 | 3.9 | |||||||||
| 5.6 | 4.3 | 5.5 | 4.4 | 0.2 | 4.1 | 3.9 | 0.1 | 3.8 | |||||||||
| 6.1 | 5.3 | 5.9 | 4.5 | 0.9 | 3.5 | 4.2 | 0.7 | 3.9 | |||||||||
| 5.6 | 4.6 | 5.5 | 4.2 | 0.6 | 3.7 | 3.9 | 1 | 3.9 | |||||||||
| 5.9 | 5.1 | 5.8 | 4.2 | 0.8 | 3.8 | 4.3 | 0.7 | 4 | |||||||||
| 9384 | 60 | 20 | 30 | 25 | 23 | 18 | 19 | 11 | 5.3 | 4.5 | 5.2 | 4.1 | 0.8 | 3.9 | 4.1 | 0.9 | 3.8 |
| 6.1 | 5.6 | 6.1 | 4.8 | 0.5 | 4.1 | 4.3 | 0.1 | 4.1 | |||||||||
| 5 | 5.1 | 5.4 | 4.1 | 0.5 | 3.4 | 3.8 | 0.3 | 3.7 | |||||||||
| 5.3 | 5.1 | 5.2 | 4.5 | 0.4 | 4.1 | 4 | 0.3 | 4.2 | |||||||||
| 5.5 | 5.2 | 5.4 | 4.1 | 0.6 | 3.9 | 3.8 | 0.6 | 4 | |||||||||
| 5.7 | 5.6 | 5.8 | 4.3 | 0.9 | 3.7 | 4 | 0.4 | 3.9 | |||||||||
| 9384 | 70 | 23 | 24 | 20 | 20 | 14 | 18 | 15 | 5.6 | 5.2 | 5.5 | 4.2 | 0.8 | 4 | 4.1 | 0.7 | 3.9 |
| 5.4 | 5.6 | 5.3 | 4.1 | 0.6 | 3.9 | 3.9 | 0.8 | 4 | |||||||||
| 5.2 | 4.9 | 5.1 | 4.4 | 0.1 | 3.8 | 3.9 | 0.2 | 3.8 | |||||||||
| 5.9 | 5.9 | 5.7 | 4.5 | 0.8 | 3.8 | 4.1 | 0.9 | 3.8 | |||||||||
| 5.8 | 5.1 | 5.7 | 4.3 | 0.8 | 3.4 | 4.2 | 0.8 | 3.7 | |||||||||
| 5.5 | 5.1 | 5.3 | 4.4 | 0.8 | 3.8 | 4.1 | 0.7 | 3.9 | |||||||||
| H. hirta subsp. praetermina | |||||||||||||||||
| 1221679 | 51 | 17 | 26 | 21 | 22 | 13 | 17 | 12 | 5.1 | 4.5 | 4.9 | 4.1 | 0 | 3.5 | 4 | 0.1 | 3.9 |
| 4.9 | 4.4 | 4.8 | 4.1 | 0.2 | 3.5 | 3.7 | 0.1 | 3.2 | |||||||||
| 4.5 | 4.2 | 4.4 | 4 | 0 | 3.4 | 3.8 | 0 | 3.3 | |||||||||
| 4.9 | 4.3 | 4.8 | 4 | 0 | 3.9 | 4 | 0 | 4 | |||||||||
| 4.2 | 4.1 | 4.1 | 4 | 0 | 3.9 | 3.7 | 0 | 4 | |||||||||
| 4.7 | 4.6 | 4.3 | 4 | 0 | 3.6 | 4 | 0 | 3.9 | |||||||||
| 1221679 | 62 | 20 | 32 | 24 | 26 | 18 | 18 | 13 | 4.6 | 4 | 4.5 | 4 | 0.1 | 3.2 | 3.9 | 0.1 | 3.9 |
| 5 | 4.7 | 4.9 | 3.7 | 0 | 3.8 | 3.9 | 0 | 3.9 | |||||||||
| 4.7 | 4.6 | 4.5 | 3.9 | 0 | 3.5 | 3.6 | 0 | 3.8 | |||||||||
| 4.4 | 4 | 4.3 | 4 | 0 | 3.8 | 3.8 | 0 | 3.2 | |||||||||
| 4.2 | 4.1 | 4 | 3.7 | 0 | 3.5 | 3.8 | 0 | 3.8 | |||||||||
| 4.4 | 4 | 4.3 | 4 | 0 | 3.8 | 3.7 | 0.1 | 4 | |||||||||
| 1221679 | 84 | 23 | 41 | 31 | 29 | 21 | 27 | 18 | 5.6 | 5.2 | 5.5 | 4.7 | 0 | 4 | 4.4 | 0.1 | 3.5 |
| 5.6 | 5.5 | 5.3 | 4.2 | 0 | 3.7 | 4 | 0.1 | 3.8 | |||||||||
| 5.4 | 5.3 | 5.2 | 4.2 | 0 | 3.8 | 4 | 0 | 3.5 | |||||||||
| 6 | 5.1 | 5.9 | 4.2 | 0.2 | 4 | 4.1 | 0.1 | 3.5 | |||||||||
| 5.3 | 5.1 | 5.2 | 4.7 | 0.1 | 3.8 | 4.5 | 0 | 3.9 | |||||||||
| 5.6 | 5.3 | 5.4 | 4.1 | 0.1 | 3.5 | 4.2 | 0.1 | 3.8 | |||||||||
| 1221679 | 74 | 22 | 41 | 31 | 28 | 19 | 11 | 17 | 5.2 | 5 | 5.1 | 4.4 | 0 | 3.5 | 3.9 | 0 | 3.2 |
| 6 | 5.2 | 5.9 | 4.6 | 0 | 4.1 | 4.3 | 0 | 3.9 | |||||||||
| 5 | 4.9 | 4.9 | 4.5 | 0 | 3.7 | 4.1 | 0.1 | 3.5 | |||||||||
| 5.1 | 4.8 | 5 | 4.2 | 0 | 3.4 | 4 | 0.1 | 3.6 | |||||||||
| 6.1 | 5.1 | 6 | 4.3 | 0.1 | 4.1 | 4.1 | 0.3 | 4.1 | |||||||||
| 5.6 | 5.3 | 5.5 | 4.2 | 0 | 4.1 | 4.1 | 0 | 3.5 | |||||||||
| 1245158 | 55 | 12 | 36 | 26 | 26 | 17 | 19 | 14 | 5.3 | 4.9 | 5.2 | 4.6 | 0 | 4.1 | 4.1 | 0 | 3.6 |
| 4.6 | 4.3 | 4.5 | 4.5 | 0 | 3.2 | 4 | 0 | 3.3 | |||||||||
| 4.8 | 4.6 | 4.7 | 3.8 | 0 | 3.2 | 3.8 | 0 | 3.6 | |||||||||
| 4.2 | 3.8 | 4.1 | 3.8 | 0 | 3.2 | 3.5 | 0 | 3.3 | |||||||||
| 5.7 | 4.7 | 5.6 | 4.1 | 0 | 3.5 | 4.1 | 0 | 3.9 | |||||||||
| 5.2 | 4.9 | 5.1 | 4.1 | 0.1 | 3.6 | 4 | 0.2 | 3.4 | |||||||||
| 1245158 | 65 | 18 | 35 | 28 | 24 | 16 | 21 | 17 | 5.4 | 5.3 | 5.1 | 4 | 0 | 3.7 | 4 | 0 | 3.8 |
| 5.1 | 4.8 | 5 | 4.2 | 0 | 3.9 | 3.9 | 0.1 | 3.9 | |||||||||
| 4.6 | 4.2 | 4.5 | 4 | 0 | 3.1 | 3.7 | 0 | 3.2 | |||||||||
| 4.6 | 4.1 | 4.5 | 4 | 0 | 3.8 | 3.8 | 0 | 3.2 | |||||||||
| 4.2 | 3.9 | 4.1 | 3.5 | 0 | 3.2 | 3.5 | 0 | 3.6 | |||||||||
| 4.8 | 4.7 | 5 | 4.1 | 0 | 3.7 | 3.9 | 0 | 3.7 | |||||||||
| 1245158 | 54 | 17 | 31 | 22 | 26 | 18 | 21 | 16 | 4.3 | 4.1 | 4.2 | 3.6 | 0 | 3.2 | 3.6 | 0 | 3.3 |
| 5.2 | 5 | 5.1 | 4.2 | 0 | 3.5 | 4.1 | 0 | 3.6 | |||||||||
| 5.1 | 4.1 | 5 | 4.2 | 0 | 3.6 | 4.1 | 0 | 3.5 | |||||||||
| 5.3 | 4.9 | 5.2 | 4.1 | 0.2 | 3.9 | 4 | 0.1 | 3.7 | |||||||||
| 4.5 | 4.2 | 4.4 | 2.7 | 0 | 2.6 | 3.5 | 0.4 | 3.2 | |||||||||
| 5.5 | 4.9 | 5.4 | 4.6 | 0.1 | 4.3 | 4.1 | 0 | 4.1 | |||||||||
| 1245158 | 51 | 18 | 29 | 26 | 22 | 18 | 21 | 14 | 4.9 | 4.5 | 4.8 | 4.2 | 0 | 4.1 | 4 | 0 | 3.4 |
| 4.3 | 2.7 | 4.1 | 3.7 | 0 | 3.1 | 3.8 | 0 | 3.2 | |||||||||
| 4.9 | 4.2 | 4.7 | 4.1 | 0 | 3.9 | 7 | 0 | 3.3 | |||||||||
| 4.3 | 3.9 | 4.1 | 3.8 | 0 | 3.2 | 3.6 | 0 | 3.6 | |||||||||
| 5.1 | 4.6 | 4.9 | 4.3 | 0.1 | 3.4 | 4.1 | 0.1 | 3.6 | |||||||||
| 4.8 | 4.1 | 4.5 | 4 | 0 | 3.9 | 3.8 | 0.1 | 3.2 | |||||||||
| 1245158 | 80 | 22 | 32 | 30 | 28 | 24 | 24 | 17 | 5.4 | 5.3 | 5.1 | 4 | 0 | 3.7 | 4 | 0 | 3.8 |
| 5.1 | 4.8 | 5 | 4.2 | 0 | 3.9 | 3.9 | 0.1 | 3.9 | |||||||||
| 4.6 | 4.2 | 4.5 | 4 | 0 | 3.1 | 3.7 | 0 | 3.2 | |||||||||
| 4.6 | 4.1 | 4.5 | 4 | 0 | 3.8 | 3.8 | 0 | 3.2 | |||||||||
| 4.2 | 3.9 | 4.1 | 3.5 | 0 | 3.2 | 3.5 | 0 | 3.6 | |||||||||
| 4.8 | 4.7 | 5 | 4.1 | 0 | 3.7 | 3.9 | 0 | 3.7 | |||||||||
| 767323 | 71 | 22 | 48 | 39 | 36 | 24 | 29 | 26 | 5.3 | 4.9 | 5.2 | 4.6 | 0 | 4.1 | 4.1 | 0 | 3.6 |
| 4.6 | 4.3 | 4.5 | 4.5 | 0 | 3.2 | 4 | 0 | 3.3 | |||||||||
| 4.8 | 4.6 | 4.7 | 3.8 | 0 | 3.2 | 3.8 | 0 | 3.6 | |||||||||
| 4.2 | 3.8 | 4.1 | 3.8 | 0 | 3.2 | 3.5 | 0 | 3.3 | |||||||||
| 5.7 | 4.7 | 5.6 | 4.1 | 0 | 3.5 | 4.1 | 0 | 3.9 | |||||||||
| 5.2 | 4.9 | 5.1 | 4.1 | 0.1 | 3.6 | 4 | 0.2 | 3.4 | |||||||||
| H. hirta subsp. hirta | |||||||||||||||||
| 9358 | 70 | 22 | 38 | 34 | 28 | 24 | 21 | 18 | 5.1 | 4.8 | 4.9 | 4.9 | 0.9 | 4 | 5 | 1 | 4.3 |
| 5.6 | 4.8 | 5.5 | 5.1 | 0.9 | 4 | 4.8 | 0.7 | 3.9 | |||||||||
| 5.5 | 5.1 | 5.5 | 5.2 | 0.9 | 4.3 | 5.1 | 0.8 | 4.1 | |||||||||
| 5.5 | 5.4 | 5.4 | 5 | 0.9 | 4 | 4.6 | 0.8 | 4 | |||||||||
| 4.9 | 5.1 | 4.8 | 4.8 | 0.8 | 4.1 | 4.5 | 0.9 | 4 | |||||||||
| 5.1 | 4.7 | 5 | 4.9 | 0.9 | 4 | 4.2 | 0.8 | 4.1 | |||||||||
| 9358 | 90 | 32 | 48 | 37 | 36 | 25 | 21 | 17 | 5.2 | 4.9 | 5.1 | 4.9 | 0.9 | 4.2 | 4.6 | 0.9 | 4.1 |
| 5.3 | 4.9 | 5.2 | 4.9 | 0.8 | 4.2 | 4.6 | 0.7 | 4.1 | |||||||||
| 6 | 5.2 | 5.9 | 5.2 | 1 | 4.5 | 4.9 | 0.9 | 4.1 | |||||||||
| 5.9 | 5 | 5.8 | 4.5 | 0.6 | 4.1 | 5 | 1.1 | 4.1 | |||||||||
| 5.8 | 5.6 | 5.7 | 5 | 0.8 | 4.2 | 5 | 0.9 | 4.2 | |||||||||
| 5.5 | 5 | 5.4 | 4.9 | 0.6 | 4.5 | 4.8 | 0.6 | 4.3 | |||||||||
| 9358 | 65 | 20 | 30 | 25 | 20 | 18 | 19 | 11 | 5 | 4.7 | 4.9 | 4.1 | 0.8 | 4 | 4.1 | 0.6 | 3.9 |
| 4.8 | 4.3 | 4.7 | 4.1 | 0.6 | 3.8 | 3.8 | 0.5 | 3.7 | |||||||||
| 5.2 | 4.6 | 5.1 | 4.2 | 0.8 | 3.7 | 4.2 | 0.6 | 3.8 | |||||||||
| 5.1 | 4.6 | 5 | 4.2 | 0.6 | 4.2 | 4.1 | 0.8 | 4.1 | |||||||||
| 4.3 | 4.2 | 4.1 | 4 | 0.9 | 3.5 | 3.8 | 0.9 | 3.7 | |||||||||
| 4.9 | 4.7 | 4.8 | 4.2 | 0.8 | 3.8 | 4 | 0.8 | 3.9 | |||||||||
| 9358 | 75 | 18 | 32 | 28 | 27 | 19 | 14 | 17 | 4.7 | 4.2 | 4.6 | 4.2 | 0.9 | 3.5 | 4.1 | 0.7 | 3.6 |
| 5 | 4.5 | 4.9 | 4.6 | 0.7 | 4.1 | 4.3 | 0.6 | 4 | |||||||||
| 4.5 | 4 | 4.4 | 4.1 | 0.8 | 3.2 | 4.2 | 0.9 | 3.2 | |||||||||
| 4.8 | 4.6 | 4.7 | 4.3 | 0.5 | 4 | 3.9 | 0.6 | 3.8 | |||||||||
| 5.1 | 4.7 | 5 | 4.4 | 0.7 | 3.9 | 4 | 0.4 | 3.6 | |||||||||
| 4.8 | 4.2 | 4.6 | 4.5 | 0.9 | 3.9 | 4.5 | 0.8 | 4 | |||||||||
| 570435 | 68 | 20 | 31 | 25 | 29 | 18 | 19 | 16 | 5.6 | 5.7 | 5.5 | 4.5 | 0 | 4 | 4.7 | 0.2 | 4 |
| 5.2 | 4.9 | 5.1 | 4 | 0.4 | 4 | 4.2 | 0.3 | 3.9 | |||||||||
| 5.2 | 5.1 | 5.5 | 4.6 | 0.8 | 3.8 | 4.2 | 0.6 | 3.7 | |||||||||
| 5.2 | 5 | 5.1 | 4.5 | 0.8 | 3.9 | 4.3 | 0.8 | 4.1 | |||||||||
| 6.1 | 5.3 | 6 | 5 | 0.4 | 4.7 | 3.9 | 1 | 4 | |||||||||
| 570435 | 58 | 18 | 28 | 19 | 17 | 16 | 16 | 11 | 6.1 | 5.6 | 6 | 4.9 | 0.7 | 3.9 | 4.9 | 1 | 3.8 |
| 5.4 | 4.8 | 5.3 | 4.9 | 1.1 | 4 | 4.9 | 1 | 4.1 | |||||||||
| 5.9 | 5.1 | 5.8 | 4.7 | 1.1 | 4 | 4.9 | 0.2 | 4 | |||||||||
| 5.3 | 4.6 | 5.2 | 4.2 | 0 | 3.9 | 4.2 | 0.3 | 3.9 | |||||||||
| 6.3 | 5.3 | 6.2 | 4.4 | 0.2 | 3.9 | 4.2 | 0.4 | 3.9 | |||||||||
| 570435 | 68 | 20 | 40 | 28 | 28 | 18 | 20 | 14 | 4.6 | 4.2 | 4.5 | 4.1 | 0.4 | 3.7 | 4 | 0.5 | 3.6 |
| 4.5 | 4.1 | 4.4 | 3.8 | 0 | 3.9 | 4.1 | 0.4 | 3.6 | |||||||||
| 4.5 | 4.3 | 4.4 | 4.4 | 0.7 | 3.8 | 4.1 | 0.6 | 3.8 | |||||||||
| 5 | 4.3 | 4.9 | 4.1 | 0.6 | 3.8 | 4.2 | 0.4 | 3.9 | |||||||||
| 4.2 | 4 | 4.1 | 4.2 | 0.6 | 3.6 | 4.1 | 0.3 | 3.8 | |||||||||
| 4.9 | 4.3 | 4.8 | 4.4 | 0.6 | 3.8 | 4.2 | 0.6 | 3.9 | |||||||||
| 505794 | 70 | 20 | 35 | 30 | 28 | 19 | 20 | 16 | 5.1 | 4.3 | 5 | 4.5 | 0.9 | 3.6 | 4.3 | 1 | 3.5 |
| 5.2 | 5 | 5.1 | 4.4 | 0.4 | 3.7 | 4.3 | 0.7 | 4.1 | |||||||||
| 5 | 4.2 | 4.9 | 4.6 | 0.4 | 3.9 | 4.5 | 0.5 | 4.1 | |||||||||
| 5.2 | 4.6 | 5.1 | 4.7 | 0.8 | 3.9 | 4.1 | 0.4 | 4.2 | |||||||||
| 5.2 | 4.8 | 5.1 | 4.6 | 0.4 | 3.3 | 4.2 | 0.3 | 4 | |||||||||
| 5.3 | 4.8 | 5.1 | 4.5 | 0.6 | 3.5 | 4.2 | 0.4 | 4 | |||||||||
| 505794 | 120 | 40 | 40 | 33 | 30 | 22 | 22 | 20 | 6 | 5.7 | 5.9 | 5 | 0.7 | 4.6 | 5.1 | 0.7 | 4.1 |
| 6.1 | 5.7 | 6 | 5 | 0.9 | 4.4 | 4.8 | 0.6 | 4.5 | |||||||||
| 5.4 | 5.2 | 5.3 | 5.2 | 0.6 | 4.3 | 5 | 0.5 | 4.1 | |||||||||
| 5.5 | 5.1 | 5.4 | 4.8 | 0.1 | 4.5 | 4.7 | 0.6 | 4.2 | |||||||||
| 5.3 | 4.9 | 5.2 | 4.5 | 0.4 | 4 | 4.5 | 0.3 | 4.2 | |||||||||
| 5.7 | 5 | 5.6 | 4.8 | 0.7 | 4.3 | 4.9 | 0.6 | 4.5 | |||||||||
| 505794 | 115 | 30 | 55 | 40 | 35 | 24 | 30 | 25 | 5 | 4.5 | 4.9 | 4.1 | 0.3 | 4 | 4.1 | 0.6 | 4 |
| 5 | 4.5 | 4.9 | 4.2 | 0.6 | 4.1 | 4.1 | 0.8 | 3.9 | |||||||||
| 5.3 | 4.9 | 5.2 | 4.6 | 0.6 | 4.1 | 4.4 | 0.7 | 4.1 | |||||||||
| 5.3 | 5 | 5.2 | 4 | 1.2 | 3.8 | 3.6 | 0.9 | 3.2 | |||||||||
| 5.1 | 4.8 | 5 | 4.2 | 0.8 | 3.8 | 4.1 | 0.7 | 3.8 | |||||||||
| 4.4 | 4.1 | 4 | 4.1 | 0.4 | 3.8 | 3.9 | 0.1 | 3.1 | |||||||||
| 487263 | 50 | 16 | 30 | 27 | 28 | 18 | 17 | 19 | 5.3 | 4.7 | 5.2 | 4.6 | 0.3 | 4 | 4 | 0.5 | 4.1 |
| 4.9 | 4.2 | 4.8 | 4.1 | 0.1 | 3.8 | 4 | 0.6 | 4 | |||||||||
| 6 | 5.1 | 5.9 | 4.5 | 0 | 4.1 | 4.5 | 0.7 | 3.8 | |||||||||
| 5.2 | 4.6 | 5.1 | 4.6 | 0.5 | 4 | 3.9 | 0.1 | 4 | |||||||||
| 4.7 | 4 | 4.6 | 4.1 | 0.2 | 3.8 | 4 | 0.7 | 3.9 | |||||||||
| 5.2 | 5.1 | 5.1 | 4.7 | 0.6 | 4.1 | 4.4 | 0.5 | 4.1 | |||||||||
| 487263 | 60 | 18 | 25 | 18 | 18 | 16 | 22 | 18 | 5.3 | 4.5 | 5.2 | 4.9 | 0.1 | 3 | 4.7 | 0.1 | 3.3 |
| 5 | 4.5 | 4.9 | 4.6 | 0.4 | 4 | 4.2 | 0.1 | 3.9 | |||||||||
| 5 | 4.6 | 4.9 | 4.8 | 0.6 | 3.3 | 4.8 | 0.7 | 4.1 | |||||||||
| 5.1 | 4.3 | 4.9 | 4.9 | 0.8 | 4.2 | 4.7 | 0.6 | 4.1 | |||||||||
| 4.8 | 4.1 | 4.7 | 4.5 | 0.6 | 3.8 | 4.1 | 0.1 | 4.1 | |||||||||
| 5.1 | 4.3 | 5 | 4.3 | 0.5 | 4 | 4.1 | 0.7 | 3.9 | |||||||||
| 487263 | 80 | 25 | 32 | 24 | 30 | 22 | 22 | 18 | 5.1 | 4.5 | 5 | 4.6 | 0.8 | 3.9 | 4.5 | 0.6 | 4 |
| 4.5 | 4 | 4.2 | 4.1 | 0.3 | 3.6 | 4.1 | 0.3 | 4.1 | |||||||||
| 4.7 | 4.4 | 4.6 | 4.8 | 0.5 | 4 | 4.5 | 0.2 | 4.1 | |||||||||
| 4.8 | 4.1 | 4.7 | 4.3 | 0.6 | 4 | 4.4 | 0.5 | 4 | |||||||||
| 4.8 | 4 | 4.7 | 4.1 | 0.5 | 3.9 | 4.1 | 0.6 | 3.8 | |||||||||
| 5.1 | 3.9 | 5 | 4.4 | 0.3 | 3.9 | 4.1 | 0.4 | 3.9 | |||||||||
| 487262 | 100 | 22 | 32 | 28 | 34 | 24 | 24 | 19 | 5.2 | 3.9 | 5.1 | 4.5 | 0.4 | 4.3 | 4.9 | 0.8 | 4.4 |
| 5.6 | 4.9 | 5.6 | 4.9 | 1.1 | 4.2 | 4.9 | 1.2 | 4.4 | |||||||||
| 5.9 | 5.7 | 5.8 | 4.8 | 0.6 | 4.2 | 4.8 | 0.9 | 4.1 | |||||||||
| 5.5 | 5 | 5.4 | 4.8 | 0.6 | 4.2 | 4.9 | 0.8 | 4.2 | |||||||||
| 5.4 | 5.1 | 5.3 | 4.8 | 0.5 | 4.1 | 4.5 | 0.8 | 4.2 | |||||||||
| 5.3 | 4.9 | 5.2 | 4.7 | 0.2 | 4.5 | 4.6 | 0.8 | 4 | |||||||||
| 487262 | 120 | 36 | 51 | 38 | 32 | 22 | 25 | 23 | 4.7 | 4.3 | 4.6 | 4.2 | 0.6 | 3.9 | 4.1 | 0.6 | 3.9 |
| 4.8 | 4.2 | 4.6 | 4.2 | 0.7 | 4 | 4.7 | 0.5 | 4.3 | |||||||||
| 4.4 | 3.9 | 4.3 | 4.5 | 0.7 | 4.1 | 4.8 | 1.2 | 3.9 | |||||||||
| 5.9 | 5.3 | 5.8 | 5.1 | 0.5 | 4.1 | 4.8 | 0.8 | 4.3 | |||||||||
| 5.1 | 4.9 | 5 | 4.9 | 0.5 | 4.2 | 4.9 | 0.9 | 4.1 | |||||||||
| 6 | 5.3 | 5.9 | 5 | 0.3 | 4.3 | 4.8 | 0.5 | 4.2 | |||||||||
Appendix B
- Data from the herbarium sheets examined.
- Hierochloë odorata (L.) Wahlenb. subsp. odorata G. Weim.
- 9359: Flora von Hinterpommern. 23.05.1886. leg. A. Paul
- 9364: Plantae Finlandle Exsiccatae. 10.06.1909. Savonia borealis, Kuopio, Mustalahti. leg.: K. Linkola
- 9395: HerbariumFlora Russicae Sobr. 11.07.1904. Fennica. Prov. Savonia borealis, par. Jorois, Järvikylä. Lindberg
- 492192: Herbarium AAU. 03.06.1995. Flora Jutlandica Exsiccata. leg.: Peter Lörken
- 9365: Flora silesiacae exciccata.No.523. 1874. A. Callier
- 9363: Fl. uplas. 1899. 32 (1820). – Holcus odoratus L: Sp. pl. ed. I. 1048 (17580) -
- Hieroch. borealis R. et Syst. II: 513 (1817). var. machrantha Grunell in Bull. Mosc. a. 1869. No 1. 137; v.sp. Fennia, pr. Helsingfors. leg.: Cobr. V.M. Sohin.
- 9361: Plantae Finlandle Exsiccatae. 30.05.1940. Hattula, Kuopio, Parola. leg.: A. Wagelius
- Hierochloë odorata (L.) Wahlenb. subsp. baltica G. Weim.
- 1015766: 17.06.1927. Södermanland, Paroccia Örno. leg.: Erik Asplund
- 486885: Plantae ex. Herb. Univ. Helsingiensis 16.06.1962. Taipalsaari, Pöniälä.Leg.: Aune Haakana
- 9395: Herbarium Flora Russiaca – 11.07.1904. (Var. firma F.Nulander, Spicileg. pl. fenn. II. 1. (1844). – H. LIndb. in Pl. Finland. excicc. No 99! – H. odorata Walb. v. supra No 195.) Fennica, Prov. Savonia borealis, par. Jorolia, Järvikylä. leg.: S.G. Lindberga
- 9386: Plantae Finlandle Excicatae- 1879. (Hierochloë odorata (L.) Wahlenb. var. firma F.Nyl.) Satakinta, par. Birkkala. leg.: Hedv. et Hj. Hjelt.
- 9369: 23.05.1873. Königsberg: Wiese bei Schönbusch. leg.: C. Baenitz.
- 9303: Planta Neerlandicae – Loc. canal near Apeldoorn. 05. 1884. leg.: H.J. Ankersmit
- Hierochloë hirta (Schrank) Borbás subsp. praetermissa (G. Weim.) B. Walln.
- 1245158: 17.06.1964. Sweden. Västergötland. Dalum. C. 800 m S of Silarp, along brook near road. leg.: Weimarck, A.; Weimarck, G.
- 1221679: 17.06.1964. Sweden. Västergötland. Dalum. C. 800 m S of Silarp, along brook near road. leg.: Weimarck, A.; Weimarck, G.
- 767323: 17.06.1964. Itakien, Südtirol, Pustertal. 20.07.1995. leg.: B. Wallnöfer
- Hierochloë hirta (Schrank) Borbás subsp. arctica (Presl) G. Weim.
- 9358: Herbarium F. Bruns. Flora Hamburgensis. 18.05.1887.
- 9393: Herbarium pf the University of Michigen. 22.06.1933. Douglas Lake. leg.: J.H. Ehlers
- 522684: Flora Fennica 10.06.19959. Pikkalara. Leg.: Paavo S. Jokela
- 9383: 27.05.1897. Novgorod, leg.: A. Kolmowskij
- 9384: 05.1900. Janissejsk, leg.: N. Martjonov
- Hierochloë hirta (Schrank) Borbás subsp. hirta G. Weim.
- 570435: 06.05.1922. Hütteldorf. leg.: J. Schaffer
- 487262: Flora Fennica 24.06.1959. Kumpulainen. Leg.: Paavo S. Jokela
- 505794: Flora Fennica 24.06.1959. Enontakiö. Leg.: Paavo S. Jokela
- 487263: Flora Fennica 06.06.1959. Ostrobottnia. Leg.: Paavo S. Jokela
References
- Watson, L.; MacFarlane, T.D.; Dallwitz, M.J. The Grass Genera of the World. Available online: https://www.delta-intkey.com/grass/www/hierochl.htm (accessed on 10 December 2024).
- Lema-Suárez, I.L.; Sahuquillo, E.; Manuel, P. A revision of Hierochloë ect. Monoecia (Anthoxanthinae, Pooideae, Poaceae). Phytotaxa 2021, 478, 92–104. [Google Scholar] [CrossRef]
- Wu, Z.Y.; Raven, P.H.; Hong, D.Y. (Eds.) Flora of China. Vol. 22 (Poaceae); Date of publication of the printed volume: 16; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MA, USA, 2006. [Google Scholar]
- Allred, K.W.; Barkworth, M. Anthoxanthum Linnaeus. Flora N. Am. 2007, 24, 758–764. [Google Scholar]
- Edgar, E.; Connor, H.E. Flora of New Zealand Volume V: Gramineae (Grasses). Nord. J. Bot. 2000, 20, 442. [Google Scholar] [CrossRef]
- Cope, T.; Gray, A. Grasses of the British Isles; Tebbs, M., Illustrations; Ashton, P., Ed.; Botanical Society of Britain & Ireland: Hertfordshire, UK, 2009; ISBN 9780901158420. [Google Scholar]
- de Lange, P.J.; James, C.J. New combinations in Anthoxanthum (Poaceae) for Aotearoa/New Zealand taxa earlier placed in Hierochloe. Ukr. Bot. J. 2024, 81, 259–262. [Google Scholar] [CrossRef]
- Iwatsuki, K.; Boufford, D.E.; Ohba, H. (Eds.) Flora of Japan; Kodansha Ltd.: Tokyo, Japan, 2020; Volume IVa. [Google Scholar]
- Allred, K.W.; Barkworth, M.E. Anthoxanthum L. & Hierochloë R.Br. In Flora of North America; Barkworth, M.E., Capels, K.M., Long, S., Anderton, L.K., Piep, M.B., Eds.; Oxford University Press: New York, NY, USA, 2007; Volume XXIV. [Google Scholar]
- Connor, H.E.; Renvoize, S.A. Hierochloe Quebrada (Poaceae), a New Species from Peru and Notes on Floral Biology in South American Species. Kew Bull. 2009, 64, 727–734. [Google Scholar] [CrossRef]
- Available online: https://powo.science.kew.org/taxon/urn:lsid:ipni.org:names:389034-1/general-information (accessed on 4 February 2025).
- Clayton, W.D.; Vorontsova, M.S.; Harman, K.T.; Williamson, H. Onwards: GRASSBASE—The Online World Grass Flora. 2006. Available online: http://www.kew.org/data/grasses-db.html (accessed on 15 February 2025).
- Govaerts, R.; Nic Lughadha, E.; Black, N.; Turner, R.; Paton, A. The World Checklist of Vascular Plants, a continuously updated resource for exploring global plant diversity. Sci. Data 2021, 8, 215. [Google Scholar] [CrossRef] [PubMed]
- Soreng, R.J.; Peterson, P.M.; Davidse, G.; Judziewicz, E.J.; Zuloaga, F.O.; Filgueiras, T.S.; Morrone, O. Catalogue of New World Grasses (Poaceae): IV. Subfamily Pooideae. Contrib. U. S. Natl. Herb. 2003, 48, 1–730. [Google Scholar]
- Schouten, Y.; Veldkamp, J.F. A revision of Anthoxanthum including Hierochloë (Gramineae) in Malesia and Thailand. Blumea 1985, 30, 319–351. [Google Scholar]
- Wu, Z.L.; Phillips, S.M. Anthoxanthum L. In Flora of China; Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China, 2006; Volume XXII. [Google Scholar]
- Zhao, Y.Z.; Zhao, L.Q.; Rui, C. (Eds.) Flora Intramongolica, 3rd ed.; Typis Intramongolicae Popularis: Huhhot, China, 2019; Volume 6. [Google Scholar]
- Chepinoga, V.V.; Seregin, A.P.; Barkalov, V.Y.; Ebel, A.L.; Efimov, P.G.; Friesen, N.V.; Gontcharov, A.A.; Kechaykin, A.A.; Knyazev, M.S.; Korobkov, A.A.; et al. New combinations and new names in vascular plants of Asian Russia. Botanica Pacifica. 2023, 12, 120–133. [Google Scholar] [CrossRef]
- Clayton, W.D.; Renvoize, S. Genera Graminum: Grasses of the world. Kew Bull. 1986, 13, 1–389. [Google Scholar]
- Connor, H.E. Floral biology of Australian species of Hierochloë (Gramineae). Aust. J. Bot. 2008, 50, 166–176. [Google Scholar] [CrossRef]
- Connor, H.E. Flowers and floral biology of the holy grasses (Hierochloë and Anthoxanthum: Aveneae, Gramineae). Flora 2012, 207, 323–333. [Google Scholar] [CrossRef]
- Edgar, E.; Connor, H.E. Flora of New Zealand, 2nd ed.; Manaaki Whenua Press: Lincoln, New Zealand, 2010; Volume V. [Google Scholar]
- Nowak, A.; Świsłowski, P.; Świerszcz, S.; Nowak, S.; Rajfur, M.; Wacławek, W. Ecovoltaics—A truly ecological and green source of renewable goods. Ecol. Chem. Eng. 2023, 30, 315–332. [Google Scholar] [CrossRef]
- Tutin, T.G. Anthoxanthum L. & Hierochloë R.Br. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1980; Volume V, pp. 229–230. [Google Scholar]
- Pimentel, M.; Sahuquillo, E.; Torrecilla, Z.; Popp, M.; Catala’n, P.; Brochmann, C. Hybridization and long-distance colonization at different time scales: Towards resolution of long-term controversies in the sweet vernal grasses (Anthoxanthum). Ann. Bot. 2013, 112, 1015–1030. [Google Scholar] [CrossRef] [PubMed]
- Lema-Suarez, I.; Balbuena, E.S.; Estevez-Perez, G.; Loureiro, J.; Castro, S.; Pereira, M.P. A taxonomic and evolutionary review of the South American Hierochloe section Monoecia (Poaceae: Anthoxanthinae). Bot. J. Linn. Soc. 2018, 186, 389. [Google Scholar] [CrossRef]
- Villalobos, N.I.; Finot, V.L.; Ruiz, E.; Penailillo, P.; Collado, G. Morphometric and taxonomic study of the native Chilean species of genus Anthoxanthum (Poaceae, Pooideae, Poeae, Anthoxanthinae). Darwiniana Nueva Ser. 2019, 7, 93–136. [Google Scholar] [CrossRef]
- Conert, H.J. Hierochloë. In Illustrierte Flora von Mitteleuropa; Hegi, G., Ed.; Paul Parey: Berlin/Hamburg, Germany, 1985; Volume: 1/3, pp. 166–175. [Google Scholar]
- Conert, H.J. Pareys Gräserbuch. In Die Gräser Deutschlands Erkennen und Bestimmen; Paul Parey Verlag: Berlin/Hamburg, Germany, 2000. [Google Scholar]
- de Lange, P.J.; Parris, B.S. New combinations in Cranfillia (Blechnaceae: Polypodiopsida) for recent segregates of the Blechnum vulcanicum complex. Telopea 2019, 22, 153–156. [Google Scholar] [CrossRef]
- Ford, M. The return of Holy Grass, a century unrecorded in the North. Trilepidea 2023, 228, 10–12. [Google Scholar]
- Ford, M.; Anthoxanthum Redolens Fact Sheet (Content Continuously Updated). New Zealand Plant Conservation Network. Available online: https://www.nzpcn.org.nz/flora/species/anthoxanthum-redolens/ (accessed on 31 May 2024).
- Bączek, K.; Angielczyk, M.; Przybył, J.; Ejdys, M.; Geszprych, A.; Weglarz, Z. Functional traits of selected clones of southern sweet-grass (Hierochloë australis/schrad./roem. & schult.). Herba Pol. 2014, 60, 23–33. [Google Scholar] [CrossRef]
- Saarela, J.M.; Liu, Q.; Peterson, P.M.; Soreng, R.J.; Paszko, B. Phylogenetics of the Grass ‘Aveneae-Type Plastid DNA Clade’ (Poaceae: Pooideae, Poeae) Based on Plastid and Nuclear Ribosomal DNA Sequence Data. In Diversity, Phylogeny and Evolution in the Monocotyledons; Seberg, Q., Peterson, G., Baxford, A.S., Davis, J.I., Eds.; Aarhus University Press: Aarhus, Denmark, 2010; pp. 557–586. [Google Scholar]
- Quintanar, A.; Castroviejo, S.; Catalán, P. Phylogeny of the tribe Aveneae (Pooideae, Poaceae) inferred from plastid trnT-F and nuclear ITS sequences. Am. J. Bot. 2007, 94, 1554–1569. [Google Scholar] [CrossRef] [PubMed]
- Tkach, N.; Schneider, J.; Döring, E.; Wölk, A.; Hochbach, A.; Nissen, J.; Winterdeld, G.; Meyer, S.; Gabriel, J.; Hoffmann, M.H.; et al. Phylogenetic lineages and the role of hybridization as driving force of evolution in grass supertribe Poodae. Taxon 2020, 69, 234–277. [Google Scholar] [CrossRef]
- Manuel, P.; Sahuquillo, E. Relationships between the close congeners Anthoxanthum odoratum and A. alpinum (Poaceae, Pooideae) assessed by morphological and molecular methods. Bot. J. Linn. Soc. 2008, 156, 237–252. [Google Scholar] [CrossRef]
- Talar-Krasa, M.; Wolski, K.; Radkowski, A.; Khachatryan, K.; Bujak, H.; Bocianowski, J. Effects of a Plasma Water and Biostimulant on Lawn Functional Value. Agronomy 2021, 11, 254. [Google Scholar] [CrossRef]
- Ciesielska, A.; Ciesielski, W.; Khachatryan, K.; Koloczek, H.; Kulawik, D.; Oszczeda, Z.; Soroka, J.A.; Tomasik, P. Structure and physicochemical properties of water treated under carbon dioxide with low-temperature low-pressure glow plasma of low frequency. Water 2020, 12, 1920. [Google Scholar] [CrossRef]
- Prończuk, S.; Prończuk, M. Evaluation of the response of perennial ryegrass (Lolium perenne L.) cultivars to temporary shading in turf maintenance. Biul. IHAR 2008, 248, 134–135. [Google Scholar]
- Laudański, Z.; Prończuk, M.; Prończuk, M. Proposition of turf characters synthesis for assessment of Festuca spp. cultivars value. Bull. Plant Breed. Acclim. Inst. 2004, 233, 181–193. [Google Scholar] [CrossRef]
- Borbás, V. A Balaton Tavának és Partmellékének Növényföldrajza és Edényes Növényzete; A Balaton Tud. Tanulm. Eredm; Hornyánszky Viktor Csász. és Kir. Udvari Könyvnyomdája: Budapest, Hungary, 1900; Volume 2, pp. 1–432. [Google Scholar]
- Jávorka, S. Magyar flóra (Flora hungarica) Magyarország virágos és edényes virágtalan növényeinek meghatározó kézikönyve; I–III Studium: Budapest, Hungary, 1925. [Google Scholar]
- Jávorka, S.; Soó, R. A Magyar Növényvilág Kézikönyve; Akadémiai Kiadó: Budapest, Hungary, 1951; Volume I–II. [Google Scholar]
- Soó, R. A Magyar Flóra és Vegetáció Rendszertani-Növényföldrajzi Kézikönyve V. (Synopsis Systematico-Geobotanica Florae Vegetationisque Hungariae VI.); Akadémiai Kiadó: Budapest, Hungary, 1980. [Google Scholar]
- Soó, R.; Kárpáti, Z. Növényhatározó II; Tankönyvkiadó: Budapest, Hungary, 1968. [Google Scholar]
- Simon, T. A Magyarországi Edényes Flóra Határozója; Nemzeti Tankönyvkiadó: Budapest, Hungary, 2000. [Google Scholar]
- Penksza, K. Trisetum–Aranyzab, Deschampsia–Sédbúza, Aira–Lengefű, Hierochloë–Szentperje, Anthoxanthum–Borjúpázsit, Holcus–Selyemperje, Corynephorus–Ezüstperje. In Új Magyar Fűvészkönyv; Király, G., Ed.; Aggteleki Nemzeti Park Igazgatóság: Jósvafő, Hungary, 2009; pp. 527–528. [Google Scholar]
- Simonkai, L. Erdély Edényes Flórája; Studium: Budapest, Hungary, 1886. [Google Scholar]
- Somlyay, L.; Penksza, K.; Pintér, I. A Hierochloë Repens Magyarországon; Botanikai Szakosztály: Budapest, Hungary, 1997. [Google Scholar]
- Penksza, K.; Engloner, A.; Bauer, L.; Asztalos, J. Összehasonlító morfológiai vizsgálatok a Hierochloë nemzetségben I. Kanitzia 1999, 7, 59–66. [Google Scholar]
- Penksza, K.; Ruprecht, E. Some corrections to the identification and data about the occurence of Hierochloe repens (Host) Simonkai in Romania. Contrib. Bot. 2002, 37, 7–11. [Google Scholar]
- Weimarck, G. Variation and taxonomy of Hierochloë (Gramineae) in the northern Hemisphere. Bot. Not. 1971, 124, 129–175. [Google Scholar]
- Weimarck, G. Apomixis and sexuality in Hierochloë australis and in Swedish H. odorata on different polyploidy levels. Bot. Not. 1967, 116, 172–176. [Google Scholar]
- Weimarck, G. Apomixis and sexuality in Hierochloë alpina (Gramineae) from Finland and Greenland and in H. monticola from Greenland. Bot. Not. 1970, 123, 495–504. [Google Scholar]
- Weimarck, G. Hierochloë hirta subsp. praetermissa subsp. nova (Poaceae), an Asiatic—E European taxon extending to N and C Europe. Symb. Bot. Upsal. 1987, 27, 175–181. [Google Scholar]
- Wallnöfer, B. Die Hierochloë odorata-Verwandtschaft in Österreich und im nordöstlichen Italien. Ann. Naturhist. Mus. Wien. B 2007, 108, 249–263. [Google Scholar]
- Wallnöfer, B. Gefäßpflanzen der Moore und Feuchtgebiete Südtirols, dargestellt in 215 Verbreitungskarten. In Kataster der Moore und Feuchtgebiete Südtirols; Tätigkeitsberichte des Biologischen Landeslabors der Autonomen Provinz Bozen-Südtirol, Biologisches Labor Autonome Provinz Bozen Südtirol: Bolzano Alto Adige, Italy, 1991; Volume 6, pp. 75–152. [Google Scholar]
- Wallnöfer, B. Fünfzig bemerkenswerte Pflanzenarten Südtirols. Verh. Zool.-Bot. Ges. Osterr. 1988, 125, 69–123. [Google Scholar]
- Perić, R.; Panjković, B.; Stojšić, V.; Rilak, S. Hierochloë repens (Host) Simonk. (Gramineae) in Serbia. Bot. Serbica 2017, 41, 71–77. [Google Scholar]
- Gęsiński, K.; Ratyńska, H. Phytocoenotic description of habitats occupied by Hierochloë repens (Host) Simonkai: A new species for the flora of Bydgoszcz. Plant Div. Evol. 2011, 129, 59–70. [Google Scholar] [CrossRef]
- Wisskirchen, R.; Haupler, H. Standardliste der Farn- und Blütenpflanzen Deutschlands; Eugen Ulmer: Stuttgart, Germany, 1998. [Google Scholar]
- Jäger, E.J.; Werner, K. (Hrsg.). In Exkursionsflora von Deutschland, 4; Gefäßpflanzen: Kritischer Band. 10. Aufl.; Spektrum Akademischer Verlag (Elsevier): München, Germany, 2005. [Google Scholar]
- Fischer, M.A. (Hrsg.). In Exkursionsflora für Österreich, Liechtenstein und Südtirol. 2; Biologiezentrum der OÖ Landesmuseen: Linz, Austria, 2025. [Google Scholar]
- Buttler, K.P.; Thieme, M. Mitarbeiter Florenliste von Deutschland—Gefäßpflanzen. Version 4. 2012. Available online: http://www.kp-buttler.de/florenliste/index.htm (accessed on 4 February 2025).
- Available online: https://europlusmed.org/cdm_dataportal/taxon/af4d4350-2947-45b4-9967-ee2bfda782 (accessed on 4 February 2025).
- Karlsson, T. Förteckning över svenska kärlväxter. Sven. Bot. Tidskr. Utgifven Af Sven. Bot. Föreningen 1998, 91, 241–560. [Google Scholar]
- Tzvelev, N. Notae de gramineis florae URSS, 7. Nov. Sist. Vyssh. Rast. 1973, 10, 79–98. [Google Scholar]
- Tikhomirov, V.N. Genus Hierochloe (Poaceae) in the Flora of Belarus (Problems of studying and preserving the flora of Eurasia). In Proceedings of the All-Russian Conference with International Participation, Dedicated to the Memory of L.V. Bardunov (1932–2008), Irkutsk, Russia, 15–19 September 2010; Publishing house of the Institute of Geography named after V.B. Sochava, Siberian Branch of the Russian Academy of Sciences. V.B. Sochava SB RAS: Irkutsk, Russia, 2010; pp. 270–271. [Google Scholar]
- The International Plant Names Index Collaborators. International Plant Names Index. Checklist Dataset 2019. Accessed via GBIF.org. Available online: https://doi.org/10.15468/uhllmw (accessed on 4 February 2025).
- Cite Taxon Page as WFO Hierochloe Odorata Subsp. Praetermissa G.Weim. Available online: http://www.worldfloraonline.org/taxon/wfo-0000917941 (accessed on 4 February 2025).
- Wahlenberg, G. Flora Upsaliensis Enumerans Plantas Circa Upsaliam Sponte Crescentes; Typis et impensis R. Acad.; Typographorum: Uppsala, Sweden, 1820; pp. 1–495. [Google Scholar]
- Schrank, v.P. Baierische Flora; Verlag München: München, Germany, 1789. [Google Scholar]
- Greenacre, M.; Groenen, P.J.F.; Hastie, T.; D’Enza, A.I.; Markos, A.; Tuzhilina, E. Principal component analysis. Nat. Rev. Methods Primers 2022, 2, 100. [Google Scholar] [CrossRef]
- Lumivero. XLSTAT Statistical and Data Analysis Solution. 2024. Available online: https://www.xlstat.com (accessed on 4 February 2025).




| H. odorata ssp. odorata | H. odorata ssp. baltica | H. hirta ssp. arctica | H. hirta ssp. praetermissa | H. hirta ssp. hirta | |
|---|---|---|---|---|---|
| 1 | 80.1 ± 12 a | 59.8 ± 8.7 a | 64.3 ± 15.0 ab | 64.9 ± 12.2 abc | 80.6 ± 23.1 bc |
| 2 | 24.3 ± 5.0 b | 17.7 ± 1.7 a | 19.4 ± 4.0 ab | 19.0 ± 3.6 ab | 23.8 ± 7.3 b |
| 3 | 40.0 ± 8.7 a | 31.7 ± 3.4 a | 34.1 ± 8.2 a | 35.1 ± 6.3 a | 36.5 ± 8.8 a |
| 4 | 29.0 ± 6.5 a | 25.2 ± 3.6 a | 25.3 ± 6.1 a | 27.4 ± 4.9 a | 28.9 ± 6.5 a |
| 5 | 28.4 ± 5.9 a | 24.5 ± 3.2 a | 25.5 ± 6.4 a | 26.5 ± 3.6 a | 28.0 ± 5.7 a |
| 6 | 19.6 ± 4.8 a | 18.3 ± 2.5 a | 19.7 ± 5.3 a | 18.4 ± 3.2 a | 21.3 ± 3.1 a |
| 7 | 22.9 ± 6.0 a | 18.6 ± 2.9 a | 18.9 ± 5.2 a | 20.0 ± 5.3 a | 20.8 ± 3.9 a |
| 8 | 16.5 ± 4.1 a | 14.1 ± 2.5 a | 15.3 ± 3.6 a | 15.9 ± 3.5 a | 17.5 ± 3.8 a |
| 9 | 4.7 ± 0.6 a | 5.5 ± 0.5 c | 5.5 ± 0.5 c | 5.0 ± 0.5 ab | 5.2 ± 0.5 b |
| 10 | 4.3 ± 0.6 a | 5.1 ± 0.4 c | 5.0 ± 0.6 c | 4.6 ± 0.5 b | 4.7 ± 0.5 b |
| 11 | 4.5 ± 0.6 a | 5.4 ± 0.5 c | 5.4 ± 0.5 c | 4.9 ± 0.5 b | 5.1 ± 0.5 b |
| 12 | 3.9 ± 0.4 a | 4.8 ± 0.3 d | 4.5 ± 0.5 c | 4.1 ± 0.3 b | 4.5 ± 0.3 c |
| 13 | 0.1 ± 0.1 b | 0.0 ± 0.0 a | 0.4 ± 0.3 c | 0.0 ± 0.1 ab | 0.6 ± 0.3 c |
| 14 | 3.5 ± 0.4 a | 4.1 ± 0.3 c | 3.9 ± 0.4 b | 3.6 ± 0.3 a | 4.0 ± 0.3 b |
| 15 | 3.7 ± 0.6 a | 4.4 ± 0.3 c | 4.2 ± 0.4 b | 4.0 ± 0.4 a | 4.4 ± 0.4 c |
| 16 | 0.0 ± 0.0 ab | 0.0 ± 0.0 a | 0.4 ± 0.3 c | 0.0 ± 0.1 b | 0.6 ± 0.3 d |
| 17 | 3.5 ± 0.4 a | 4.2 ± 0.2 c | 3.9 ± 0.4 b | 3.6 ± 0.3 a | 4.0 ± 0.3 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Penksza, K.; Szabó-Szöllösi, T.; Neményi, A.; Sipos, L.; Szentes, S.; Wagenhoffer, Z.; Palla, B.; Balogh, D.Á.; Saláta-Falusi, E. Evaluating Inflorescence Morphology in Two Species and Subspecies of the Genus Hierochloë R. Brown. Plants 2025, 14, 2270. https://doi.org/10.3390/plants14152270
Penksza K, Szabó-Szöllösi T, Neményi A, Sipos L, Szentes S, Wagenhoffer Z, Palla B, Balogh DÁ, Saláta-Falusi E. Evaluating Inflorescence Morphology in Two Species and Subspecies of the Genus Hierochloë R. Brown. Plants. 2025; 14(15):2270. https://doi.org/10.3390/plants14152270
Chicago/Turabian StylePenksza, Károly, Tünde Szabó-Szöllösi, András Neményi, László Sipos, Szilárd Szentes, Zsombor Wagenhoffer, Balázs Palla, Dániel Ákos Balogh, and Eszter Saláta-Falusi. 2025. "Evaluating Inflorescence Morphology in Two Species and Subspecies of the Genus Hierochloë R. Brown" Plants 14, no. 15: 2270. https://doi.org/10.3390/plants14152270
APA StylePenksza, K., Szabó-Szöllösi, T., Neményi, A., Sipos, L., Szentes, S., Wagenhoffer, Z., Palla, B., Balogh, D. Á., & Saláta-Falusi, E. (2025). Evaluating Inflorescence Morphology in Two Species and Subspecies of the Genus Hierochloë R. Brown. Plants, 14(15), 2270. https://doi.org/10.3390/plants14152270