
Calliviminone A from Callistemon citrinus Induces PANC-1 Pancreatic Cancer Cell Death by Targeting the PI3K/Akt/mTOR Pathway

 ,
,  , , , , , and
, , , , , and 
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
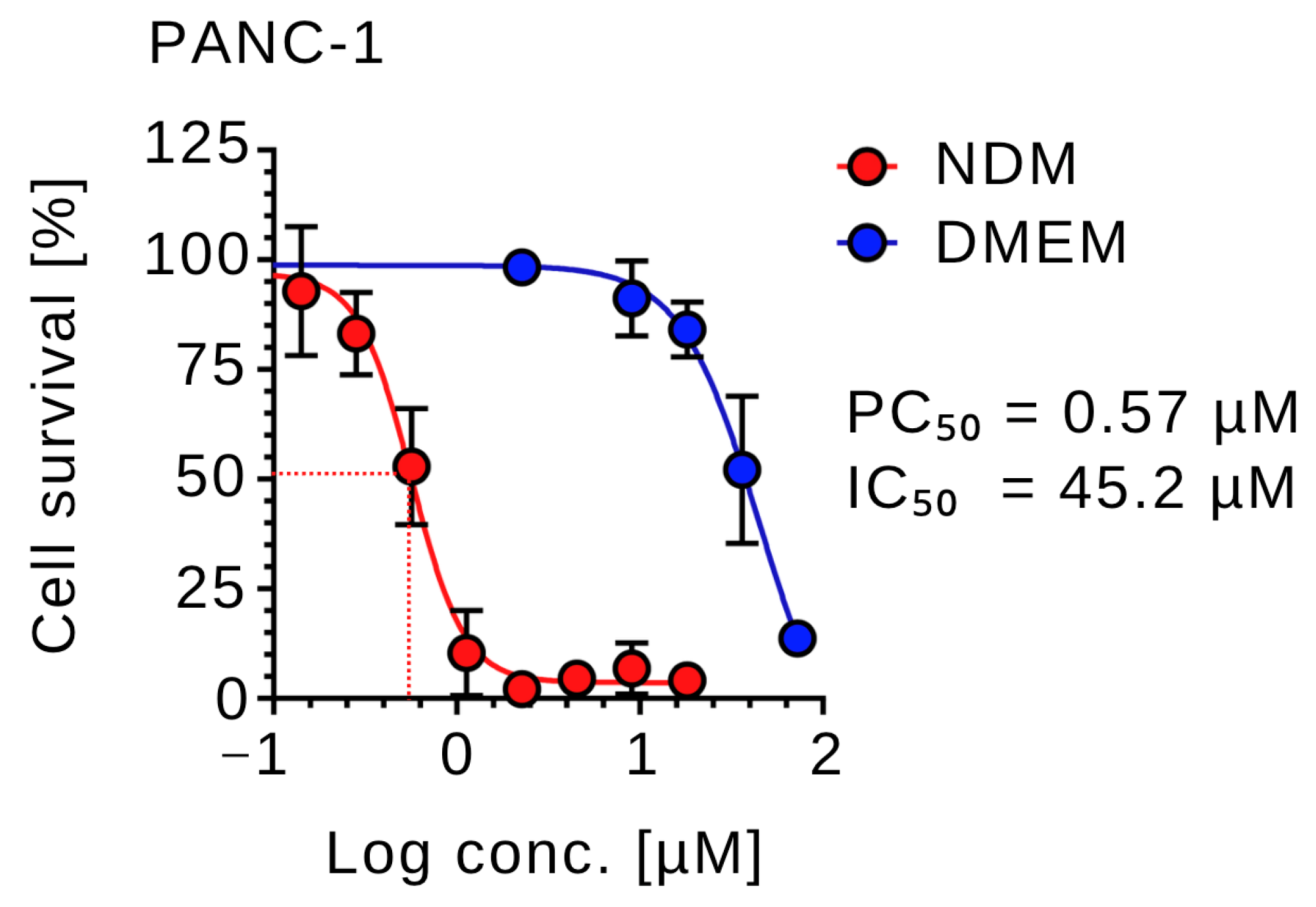
2.1. Anti-Austerity Activity of Calliviminone A (CVM-A) Against PANC-1 Human Pancreatic Cancer Cells
2.2. Real-Time Live-Cell Imaging of Calliviminone (CVM-A) A-Induced Morphological Changes and Cell Death in PANC-1 Pancreatic Cancer Cells in Nutrient-Deprived Medium (NDM)
2.3. Live-/Dead-Cell Imaging Analysis of Calliviminone A (CVM-A)-Induced Cytotoxicity in Nutrient-Deprived Medium (NDM)
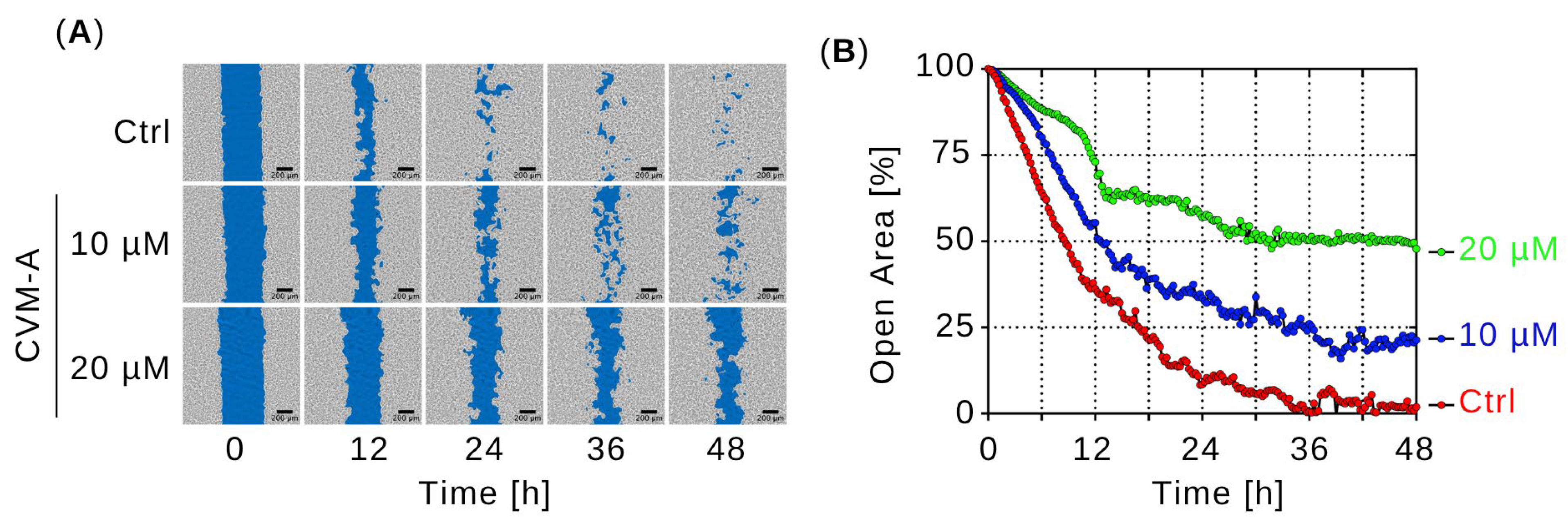
2.4. Real-Time Cell Migration Assay of Calliviminone A (CVM-A)-Induced Inhibition of PANC-1 Pancreatic Cancer Cell Migration in Normal Nutrient Medium (DMEM)
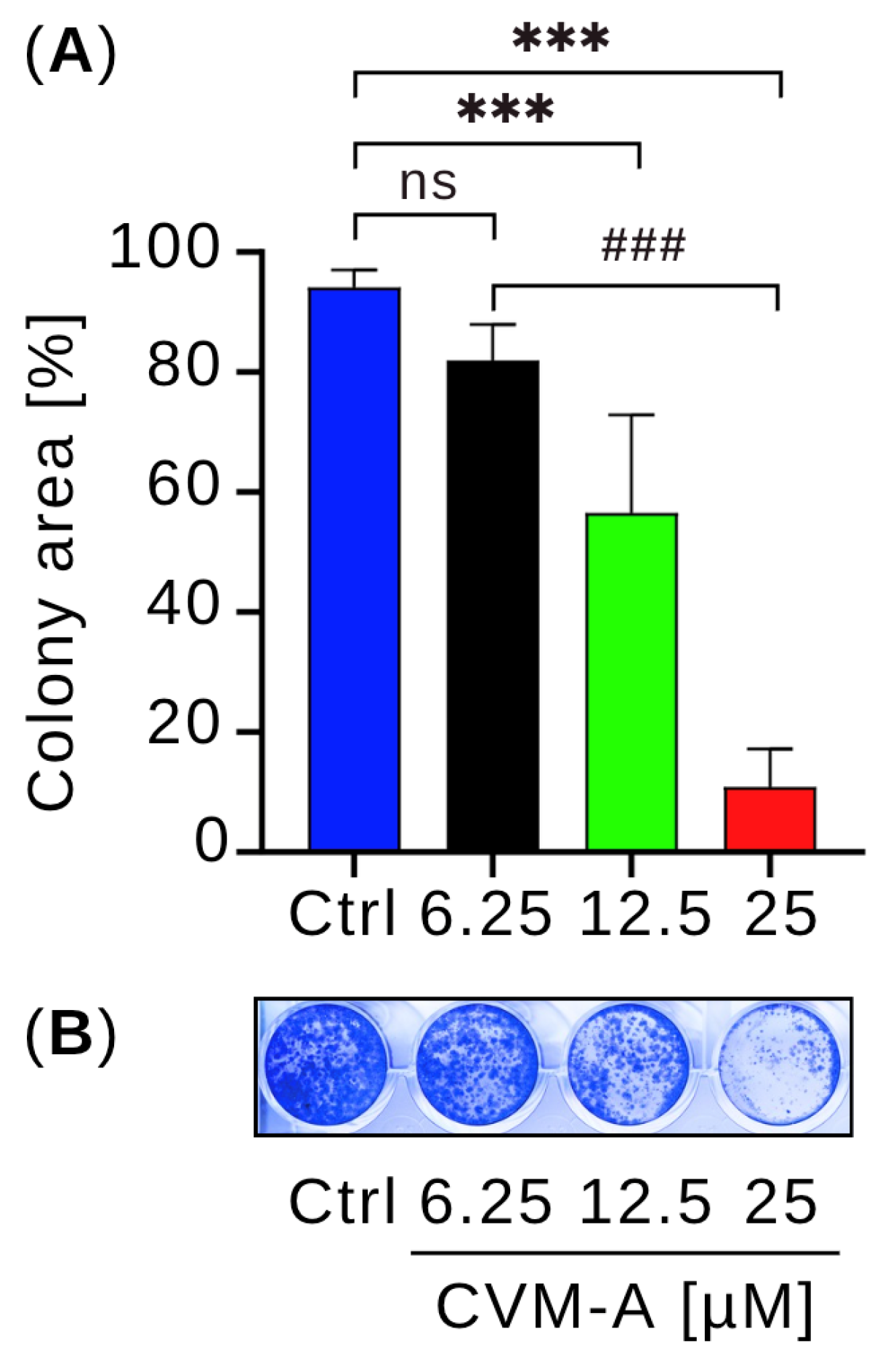
2.5. Calliviminone A (CVM-A)-Mediated Inhibition of PANC-1 Pancreatic Cancer Cell Colony Formation in Normal Nutrient Medium (DMEM)
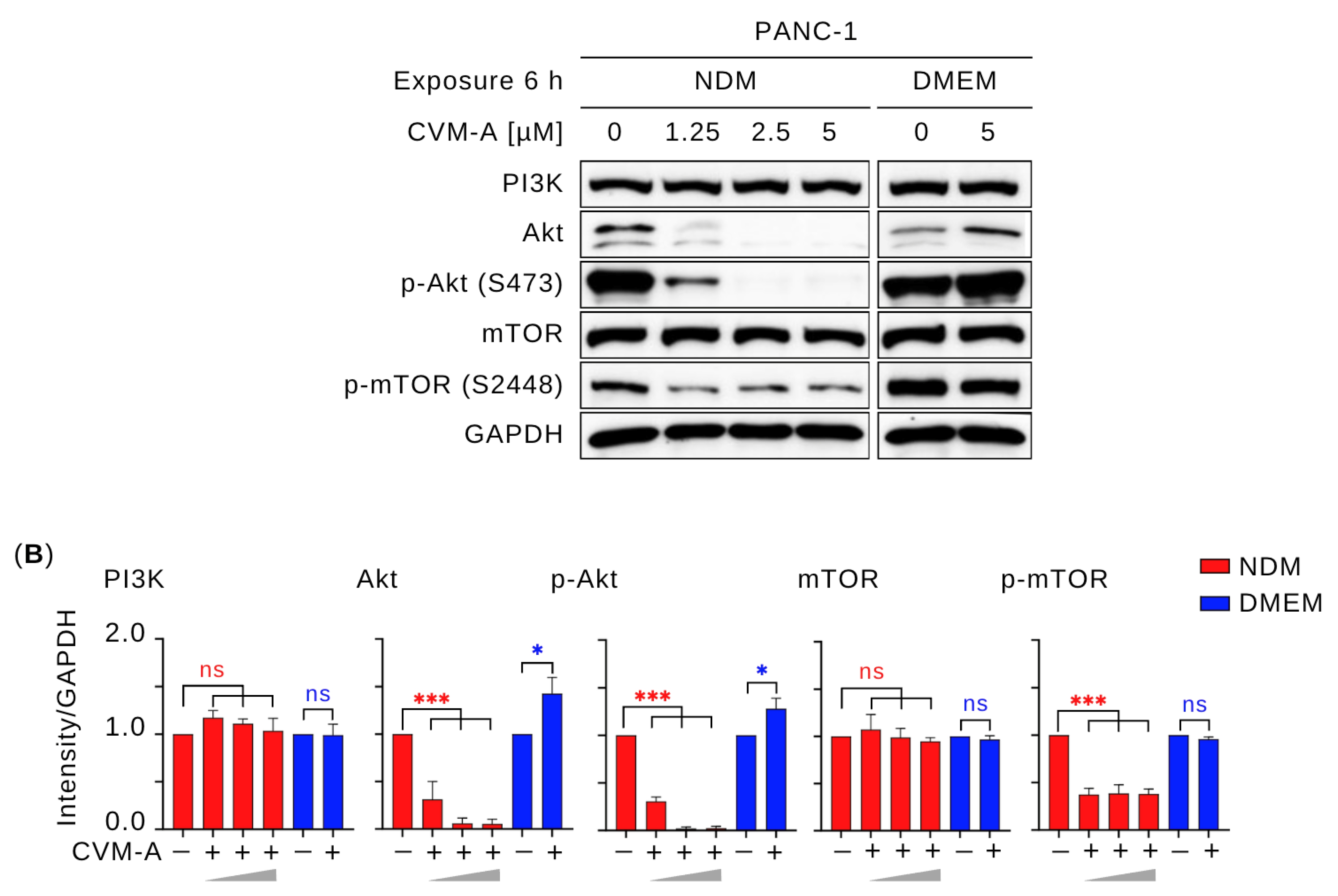
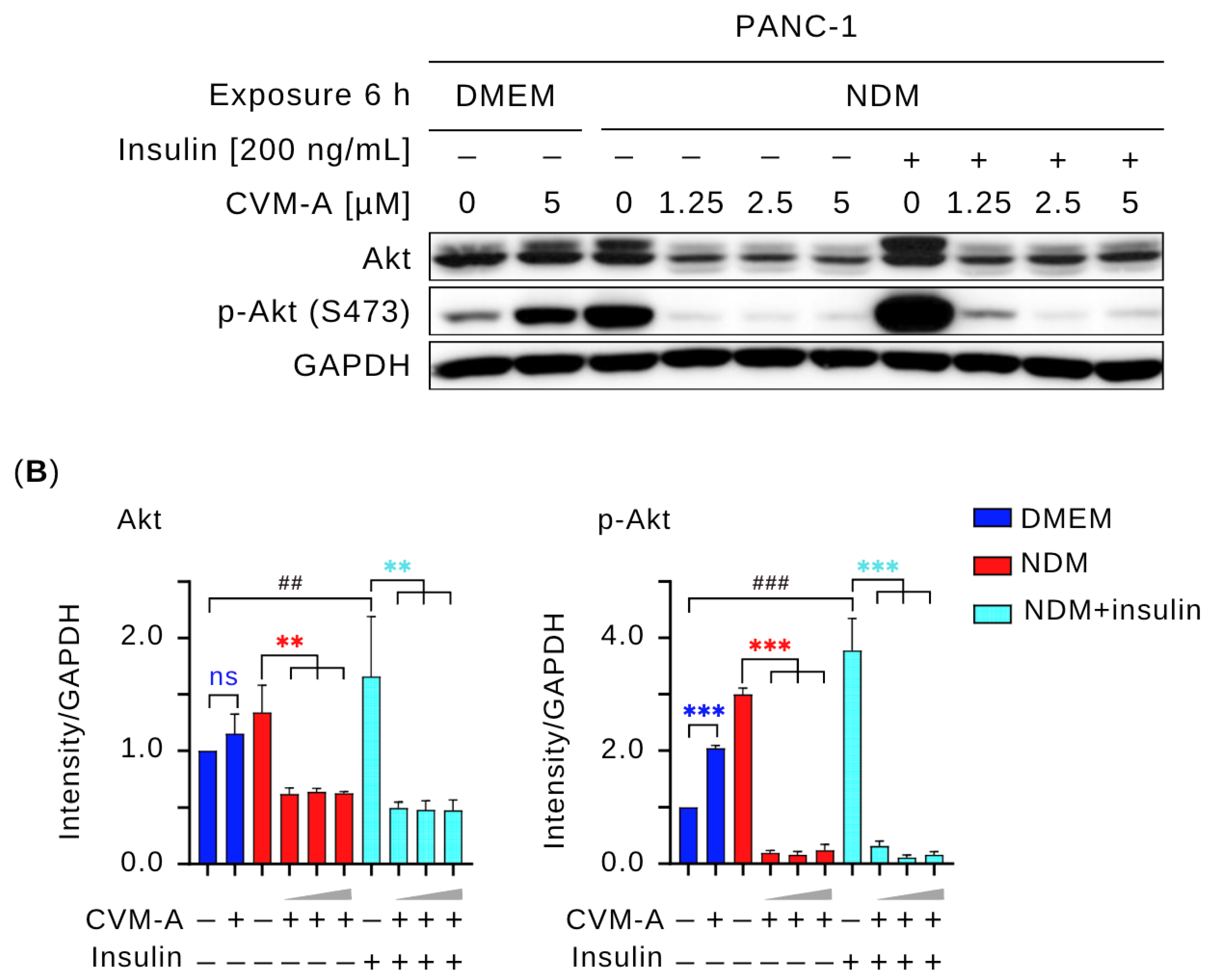
2.6. Calliviminone A (CVM-A) Inhibits Insulin-Induced PI3K/Akt/mTOR Activation in PANC-1 Pancreatic Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture
4.3. Preferential Cytotoxicity Assay
4.4. Live-Cell Imaging
4.5. Morphological Changes
4.6. Cell Migration Assay
4.7. Colony Formation Assay
4.8. Western Blot Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tsai, P.Y.; Lee, M.-S.; Jadhav, U.; Naqvi, I.; Madha, S.; Adler, A.; Mistry, M.; Naumenko, S.; Lewis, C.A.; Hitchcock, D.S.; et al. Adaptation of pancreatic cancer cells to nutrient deprivation is reversible and requires glutamine synthetase stabilization by mTORC1. Proc. Natl. Acad Sci. USA 2021, 118, e2003014118. [Google Scholar] [CrossRef]
- Michetti, F.; Cirone, M.; Strippoli, R.; D’Orazi, G.; Cordani, M. Mechanistic insights and therapeutic strategies for targeting autophagy in pancreatic ductal adenocarcinoma. Discov. Oncol. 2025, 16, 592. [Google Scholar] [CrossRef]
- Mizrahi, J.D.; Surana, R.; Valle, J.W.; Shroff, R.T. Pancreatic cancer. Lancet 2020, 395, 2008–2020. [Google Scholar] [CrossRef] [PubMed]
- Halbrook, C.J.; Lyssiotis, C.A.; Pasca Di Magliano, M.; Maitra, A. Pancreatic cancer: Advances and challenges. Cell 2023, 186, 1729–1754. [Google Scholar] [CrossRef] [PubMed]
- Qadir, R.M.A.B.; Umair, M.B.; Tariq, U.B.; Ahmad, A.; Kiran, W.; Shahid, M.H. Unraveling pancreatic cancer: Epidemiology, risk factors, and global trends. Cureus 2024, 16, e72816. [Google Scholar] [CrossRef]
- Zottl, J.; Sebesta, C.G.; Tomosel, E.; Sebesta, M.C.; Sebesta, C. Unraveling the burden of pancreatic cancer in the 21st century: Trends in incidence, mortality, survival, and key contributing factors. Cancers 2025, 17, 1607. [Google Scholar] [CrossRef]
- Leiphrakpam, P.D.; Chowdhury, S.; Zhang, M.; Bajaj, V.; Dhir, M.; Are, C. Trends in the global incidence of pancreatic cancer and a brief review of its histologic and molecular subtypes. J. Gastrointest. Cancer 2025, 56, 71. [Google Scholar] [CrossRef] [PubMed]
- Pancreatic Cancer Statistics. World Cancer Research Fund. Available online: https://www.wcrf.org/preventing-cancer/cancer-statistics/pancreatic-cancer-statistics (accessed on 1 July 2025).
- Nienhüser, H.; Büchler, M.W.; Schneider, M. Resection of recurrent pancreatic cancer: Who can benefit? Visc. Med. 2021, 38, 42–48. [Google Scholar] [CrossRef]
- Crippa, S.; Belfiori, G.; Bissolati, M.; Partelli, S.; Pagnanelli, M.; Tamburrino, D.; Gasparini, G.; Rubini, C.; Zamboni, G.; Falconi, M. Recurrence after surgical resection of pancreatic cancer: The importance of postoperative complications beyond tumor biology. HPB 2021, 23, 1666–1673. [Google Scholar] [CrossRef]
- Okusaka, T.; Furuse, J. Recent advances in chemotherapy for pancreatic cancer: Evidence from Japan and recommendations in guidelines. J. Gastroenterol. 2020, 55, 369–382. [Google Scholar] [CrossRef]
- Conroy, T.; Desseigne, F.; Ychou, M.; Bouché, O.; Guimbaud, R.; Bécouarn, Y.; Adenis, A.; Raoul, J.L.; Gourgou-Bourgade, S.; De La Fouchardière, C.; et al. FOLFIRINOX versus gemcitabine for metastatic pancreatic cancer. N. Engl. J. Med. 2011, 364, 1817–1825. [Google Scholar] [CrossRef] [PubMed]
- Di Costanzo, F.; Di Costanzo, F.; Antonuzzo, L.; Mazza, E.; Giommoni, E. Optimizing first-line chemotherapy in metastatic pancreatic cancer: Efficacy of FOLFIRINOX versus nab-paclitaxel plus gemcitabine. Cancers 2023, 15, 416. [Google Scholar] [CrossRef] [PubMed]
- Nichetti, F.; Rota, S.; Ambrosini, P.; Pircher, C.; Gusmaroli, E.; Busset, M.D.D.; Pusceddu, S.; Sposito, C.; Coppa, J.; Morano, F.; et al. NALIRIFOX, FOLFIRINOX, and gemcitabine with nab-paclitaxel as first-line chemotherapy for metastatic pancreatic cancer. JAMA Netw. Open 2024, 7, e2350756. [Google Scholar] [CrossRef]
- Cho, I.R.; Kang, H.; Jo, J.H.; Lee, H.S.; Chung, M.J.; Park, J.Y.; Park, S.W.; Song, S.Y.; An, C.; Park, M.S.; et al. FOLFIRINOX vs. gemcitabine/nab-paclitaxel for treatment of metastatic pancreatic cancer: Single-center cohort study. World J. Gastrointest. Oncol. 2020, 12, 182–194. [Google Scholar] [CrossRef]
- Principe, D.R.; Underwood, P.W.; Korc, M.; Trevino, J.G.; Munshi, H.G.; Rana, A. The current treatment paradigm for pancreatic ductal adenocarcinoma and barriers to therapeutic efficacy. Front. Oncol. 2021, 11, 688377. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Feng, Z.; Miao, R.; Liu, X.; Liu, C.; Liu, Z. Prognosis and survival analysis of patients with pancreatic cancer: Retrospective experience of a single institution. World J. Surg. Oncol. 2022, 20, 11. [Google Scholar] [CrossRef]
- De Santis, M.C.; Bockorny, B.; Hirsch, E.; Cappello, P.; Martini, M. Exploiting pancreatic cancer metabolism: Challenges and opportunities. Trends Mol. Med. 2024, 30, 592–604. [Google Scholar] [CrossRef]
- Wu, H.; Fu, M.; Wu, M.; Cao, Z.; Zhang, Q.; Liu, Z. Emerging mechanisms and promising approaches in pancreatic cancer metabolism. Cell Death Dis. 2024, 15, 553. [Google Scholar] [CrossRef]
- Wang, S.; Zheng, Y.; Yang, F.; Zhu, L.; Zhu, X.Q.; Wang, Z.F.; Wu, X.L.; Zhou, C.H.; Yan, J.Y.; Hu, B.Y.; et al. The molecular biology of pancreatic adenocarcinoma: Translational challenges and clinical perspectives. Signal Transduct. Target. Ther. 2021, 6, 249. [Google Scholar] [CrossRef]
- Ansardamavandi, A.; Nikfarjam, M.; He, H. PAK in pancreatic cancer-associated vasculature: Implications for therapeutic response. Cells 2023, 12, 2692. [Google Scholar] [CrossRef]
- Awale, S.; Lu, J.; Kalauni, S.K.; Kurashima, Y.; Tezuka, Y.; Kadota, S.; Esumi, H. Identification of arctigenin as an antitumor agent having the ability to eliminate the tolerance of cancer cells to nutrient starvation. Cancer Res. 2006, 66, 1751–1757. [Google Scholar] [CrossRef] [PubMed]
- Awale, S.; Nakashima, E.M.N.; Kalauni, S.K.; Tezuka, Y.; Kurashima, Y.; Lu, J.; Esumi, H.; Kadota, S. Angelmarin, a novel anti-cancer agent able to eliminate the tolerance of cancer cells to nutrient starvation. Bioorg. Med. Chem. Lett. 2006, 16, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Phan, N.D.; Omar, A.M.; Sun, S.; Maneenet, J.; Dibwe, D.F.; Sato, M.; Kalauni, S.K.; Toyooka, N.; Fujii, T.; Awale, S. Abietane diterpenes from Abies spectabilis and their anti-pancreatic cancer activity against the MIA PaCa-2 cell line. Bioorg. Med. Chem. Lett. 2022, 66, 128723. [Google Scholar] [CrossRef]
- Phan, N.D.; Omar, A.M.; Takahashi, I.; Baba, H.; Okumura, T.; Imura, J.; Okada, T.; Toyooka, N.; Fujii, T.; Awale, S. Nicolaioidesin C: An antiausterity agent shows promising antitumor activity in a pancreatic cancer xenograft mouse model. J. Nat. Prod. 2023, 86, 1402–1410. [Google Scholar] [CrossRef]
- Awale, S.; Baba, H.; Phan, N.D.; Kim, M.J.; Maneenet, J.; Sawaki, K.; Kanda, M.; Okumura, T.; Fujii, T.; Okada, T.; et al. Targeting pancreatic cancer with novel plumbagin derivatives: Design, synthesis, molecular mechanism, in vitro and in vivo evaluation. J. Med. Chem. 2023, 66, 8054–8065. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, T.; Phan, N.D.; Maneenet, J.; Yamagishi, M.; Nishikawa, Y.; Okada, T.; Okumura, T.; Toyooka, N.; Fujii, T.; Awale, S. Targeting pancreatic cancer with novel nicolaioidesin C derivatives: Molecular mechanism, in vitro, and in vivo evaluations. J. Med. Chem. 2024, 67, 14313–14328. [Google Scholar] [CrossRef]
- Awale, S.; Maneenet, J.; Phan, N.D.; Nguyen, H.H.; Fujii, T.; Ihmels, H.; Soost, D.; Tajuddeen, N.; Feineis, D.; Bringmann, G. Toyaburgine, a synthetic N-biphenyl-dihydroisoquinoline inspired by related N,C-coupled naphthylisoquinoline alkaloids, with high in vivo efficacy in preclinical pancreatic cancer models. Chem. Biol. 2025, 20, 917–929. [Google Scholar] [CrossRef]
- Tawila, A.M.; Sun, S.; Kim, M.J.; Omar, A.M.; Dibwe, D.F.; Ueda, J.; Toyooka, N.; Awale, S. Highly potent antiausterity agents from Callistemon citrinus and their mechanism of action against the PANC-1 human pancreatic cancer cell line. J. Nat. Prod. 2020, 83, 2221–2232. [Google Scholar] [CrossRef] [PubMed]
- Tawila, A.M.; Sun, S.; Kim, M.J.; Omar, A.M.; Dibwe, D.F.; Ueda, J.; Toyooka, N.; Awale, S. Chemical constituents of Callistemon citrinus from Egypt and their antiausterity activity against PANC-1 human pancreatic cancer cell line. Bioorg. Med. Chem. Lett. 2020, 30, 127352. [Google Scholar] [CrossRef]
- Tawila, A.M.; Omar, A.M.; Phan, N.D.; Takahashi, I.; Maneenet, J.; Awale, S. New callistrilone epimers from Callistemon citrinus and their antiausterity activity against the PANC-1 human pancreatic cancer cell line. Tetrahedron Lett. 2022, 100, 153881. [Google Scholar] [CrossRef]
- Tawila, A.M.; Sun, S.; Kim, M.J.; Omar, A.M.; Dibwe, D.F.; Awale, S. A Triterpene lactone from Callistemon citrinus inhibits the PANC-1 human pancreatic cancer cells viability through suppression of unfolded protein response. Chem. Biodivers. 2020, 17, e2000495. [Google Scholar] [CrossRef]
- Wu, L.; Luo, J.; Zhang, Y.; Zhu, M.; Wang, X.; Luo, J.; Yang, M.; Yu, B.; Yao, H.; Dai, Y.; et al. Isolation and biomimetic synthesis of (±)-calliviminones A and B, two novel diels–alder adducts, from Callistemon viminalis. Tetrahedron Lett. 2015, 56, 229–232. [Google Scholar] [CrossRef]
- He, Y.; Sun, M.M.; Zhang, G.G.; Yang, J.; Chen, K.S.; Xu, W.W.; Li, B. Targeting PI3K/Akt signal transduction for cancer therapy. Signal Transduct. Target. Ther. 2021, 6, 425. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Wang, Y.; Zhou, C.; Mei, W.; Zeng, C. PI3K/Akt/mTOR pathway and its role in cancer therapeutics: Are we making headway? Front. Oncol. 2022, 12, 819128. [Google Scholar] [CrossRef]
- Chen, Z.; Han, F.; Du, Y.; Shi, H.; Zhou, W. Hypoxic microenvironment in cancer: Molecular mechanisms and therapeutic interventions. Signal Transduct. Target. Ther. 2023, 8, 70. [Google Scholar] [CrossRef]
- Ngu, S.F.; Ngan, H.Y.; Chan, K.K. Role of adjuvant and post-surgical treatment in gynaecological cancer. Best Pract. Res. Clin. Obstet. Gynaecol. 2022, 78, 2–13. [Google Scholar] [CrossRef]
- Yang, J.; Liu, Y.; Liu, S. The role of epithelial-mesenchymal transition and autophagy in pancreatic ductal adenocarcinoma invasion. Cell Death Dis. 2023, 14, 506. [Google Scholar] [CrossRef] [PubMed]
- Maneenet, J.; Tawila, A.M.; Omar, A.M.; Phan, N.D.; Ojima, C.; Kuroda, M.; Sato, M.; Mizoguchi, M.; Takahashi, I.; Awale, S. Chemical constituents of Callistemon subulatus and their anti-pancreatic cancer activity against human PANC-1 cell line. Plants 2022, 11, 2466. [Google Scholar] [CrossRef]
- Park, W.; Chawla, A.; O’Reilly, E.M. Pancreatic cancer: A review. JAMA 2021, 326, 851–862. [Google Scholar] [CrossRef]
- Bengtsson, A.; Andersson, R.; Ansari, D. The actual 5-year survivors of pancreatic ductal adenocarcinoma based on real-world data. Sci. Rep. 2020, 10, 16425. [Google Scholar] [CrossRef]
- Guo, Z.; Ashrafizadeh, M.; Zhang, W.; Zou, R.; Sethi, G.; Zhang, X. Molecular profile of metastasis, cell plasticity and EMT in pancreatic cancer: A pre-clinical connection to aggressiveness and drug resistance. Cancer Metastasis Rev. 2024, 43, 29–53. [Google Scholar] [CrossRef]
- Alabaş, E.; Özçimen, A.A. The supression of migration and metastasis via inhibition of vascular endothelial growth factor in pancreatic adenocarcinoma cells applied danusertib. Turk. J. Gastroenterol. 2024, 35, 150–157. [Google Scholar] [CrossRef]
- Massagué, J.; Ganesh, K. Metastasis-initiating cells and ecosystems. Cancer Discov. 2021, 11, 971–994. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chen, J.; Ren, Y.; Liu, S.; Ba, Y.; Zuo, A.; Luo, P.; Cheng, Q.; Xu, H.; Han, X. Multi-stage mechanisms of tumor metastasis and therapeutic strategies. Sig. Transduct. Target. Ther. 2024, 9, 270. [Google Scholar] [CrossRef] [PubMed]
- Stanciu, S.; Ionita-Radu, F.; Stefani, C.; Miricescu, D.; Stanescu-Spinu, I.I.; Greabu, M.; Ripszky Totan, A.; Jinga, M. Targeting PI3K/AKT/mTOR signaling pathway in pancreatic cancer: From molecular to clinical aspects. Int. J. Mol. Sci. 2022, 23, 10132. [Google Scholar] [CrossRef] [PubMed]
- Glaviano, A.; Foo, A.S.C.; Lam, H.Y.; Yap, K.C.H.; Jacot, W.; Jones, R.H.; Eng, H.; Nair, M.G.; Makvandi, P.; Geoerger, B.; et al. PI3K/AKT/mTOR signaling transduction pathway and targeted therapies in cancer. Mol. Cancer 2023, 22, 138. [Google Scholar] [CrossRef]
- Ouissam, A.J.; Hind, C.; Sami Aziz, B.; Said, A. Inhibition of the PI3K/AKT/mTOR pathway in pancreatic cancer: Is it a worthwhile endeavor? Ther. Adv. Med. Oncol. 2024, 16, 17588359241284911. [Google Scholar] [CrossRef]
- Kearney, A.L.; Norris, D.M.; Ghomlaghi, M.; Kin Lok Wong, M.; Humphrey, S.J.; Carroll, L.; Yang, G.; Cooke, K.C.; Yang, P.; Geddes, T.A.; et al. Akt phosphorylates insulin receptor substrate to limit PI3K-mediated PIP3 synthesis. eLife 2021, 10, e66942. [Google Scholar] [CrossRef]
- Miao, R.; Fang, X.; Wei, J.; Wu, H.; Wang, X.; Tian, J. Akt: A Potential drug target for metabolic syndrome. Front Physiol. 2022, 13, 822333. [Google Scholar] [CrossRef]
- Tsai, P.J.; Lai, Y.H.; Manne, R.K.; Tsai, Y.-S.; Sarbassov, D.; Lin, H.K. Akt: A key transducer in cancer. J. Biomed. Sci. 2022, 29, 76. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maneenet, J.; Tawila, A.M.; Nguyen, H.H.; Phan, N.D.; Monthakantirat, O.; Daodee, S.; Boonyarat, C.; Khamphukdee, C.; Chulikhit, Y.; Awale, S. Calliviminone A from Callistemon citrinus Induces PANC-1 Pancreatic Cancer Cell Death by Targeting the PI3K/Akt/mTOR Pathway. Plants 2025, 14, 2074. https://doi.org/10.3390/plants14132074
Maneenet J, Tawila AM, Nguyen HH, Phan ND, Monthakantirat O, Daodee S, Boonyarat C, Khamphukdee C, Chulikhit Y, Awale S. Calliviminone A from Callistemon citrinus Induces PANC-1 Pancreatic Cancer Cell Death by Targeting the PI3K/Akt/mTOR Pathway. Plants. 2025; 14(13):2074. https://doi.org/10.3390/plants14132074
Chicago/Turabian StyleManeenet, Juthamart, Ahmed M. Tawila, Hung Hong Nguyen, Nguyen Duy Phan, Orawan Monthakantirat, Supawadee Daodee, Chantana Boonyarat, Charinya Khamphukdee, Yaowared Chulikhit, and Suresh Awale. 2025. "Calliviminone A from Callistemon citrinus Induces PANC-1 Pancreatic Cancer Cell Death by Targeting the PI3K/Akt/mTOR Pathway" Plants 14, no. 13: 2074. https://doi.org/10.3390/plants14132074
APA StyleManeenet, J., Tawila, A. M., Nguyen, H. H., Phan, N. D., Monthakantirat, O., Daodee, S., Boonyarat, C., Khamphukdee, C., Chulikhit, Y., & Awale, S. (2025). Calliviminone A from Callistemon citrinus Induces PANC-1 Pancreatic Cancer Cell Death by Targeting the PI3K/Akt/mTOR Pathway. Plants, 14(13), 2074. https://doi.org/10.3390/plants14132074