
New Insights into Sprout Production from Melon (Cucumis melo L. var. reticulatus) Seeds as By-Product of Fruit Processing


 ,
,
Abstract
1. Introduction
2. Results
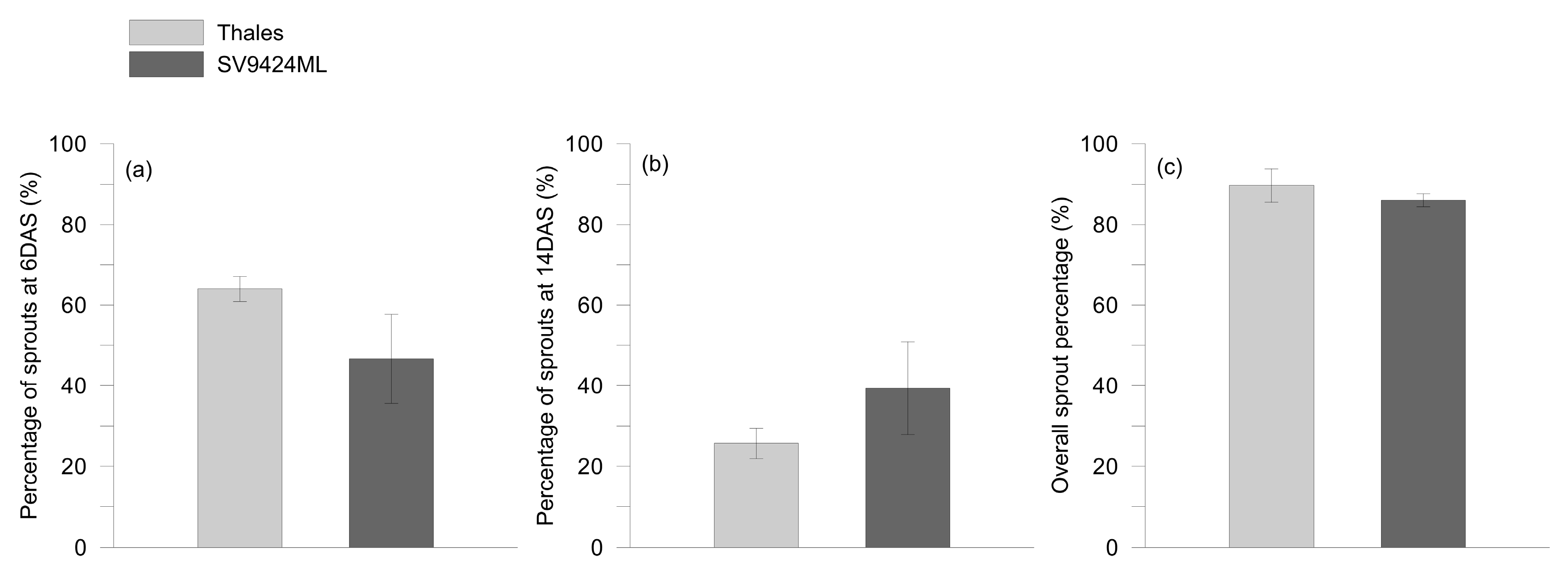
2.1. Germination and Growth Assessment
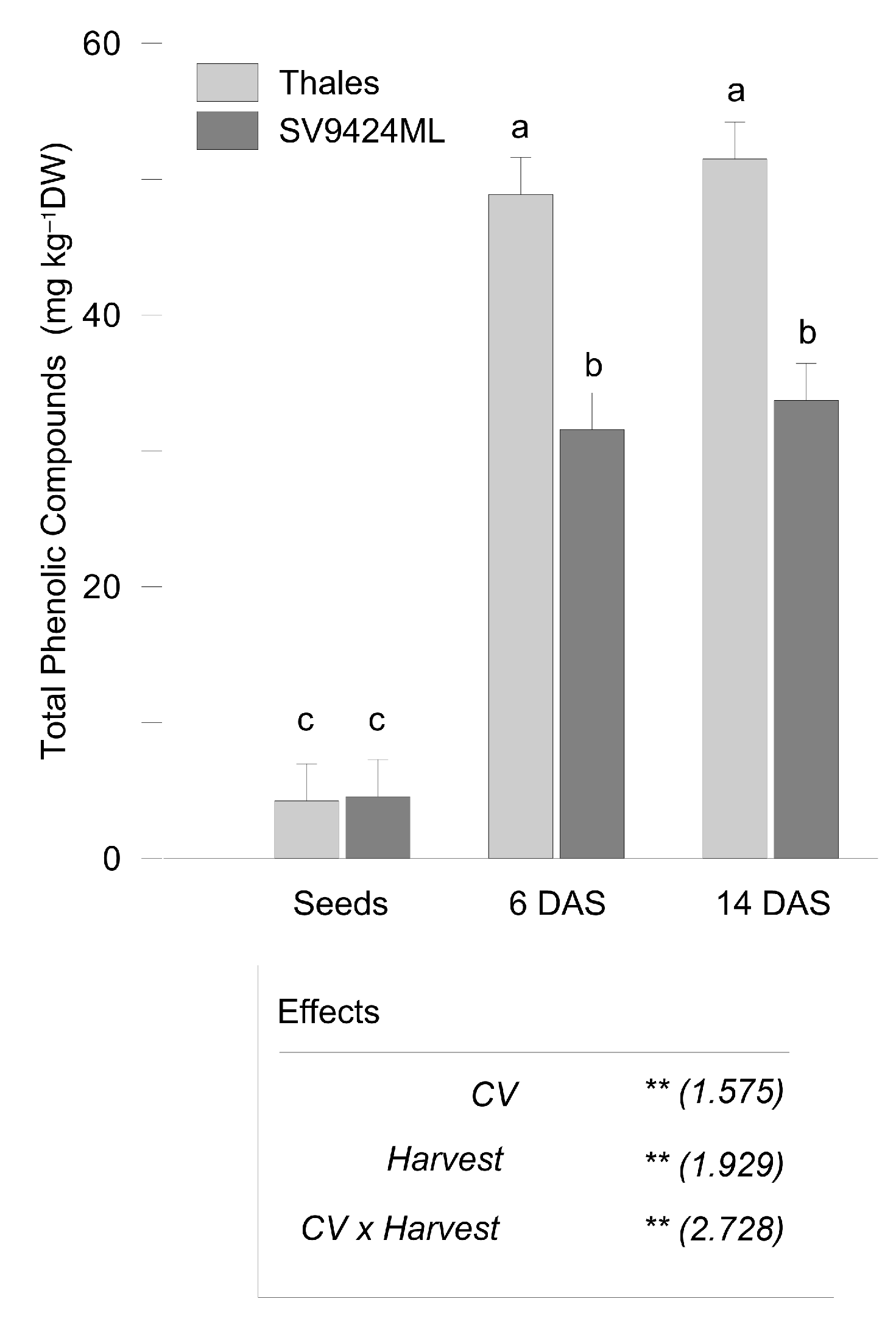
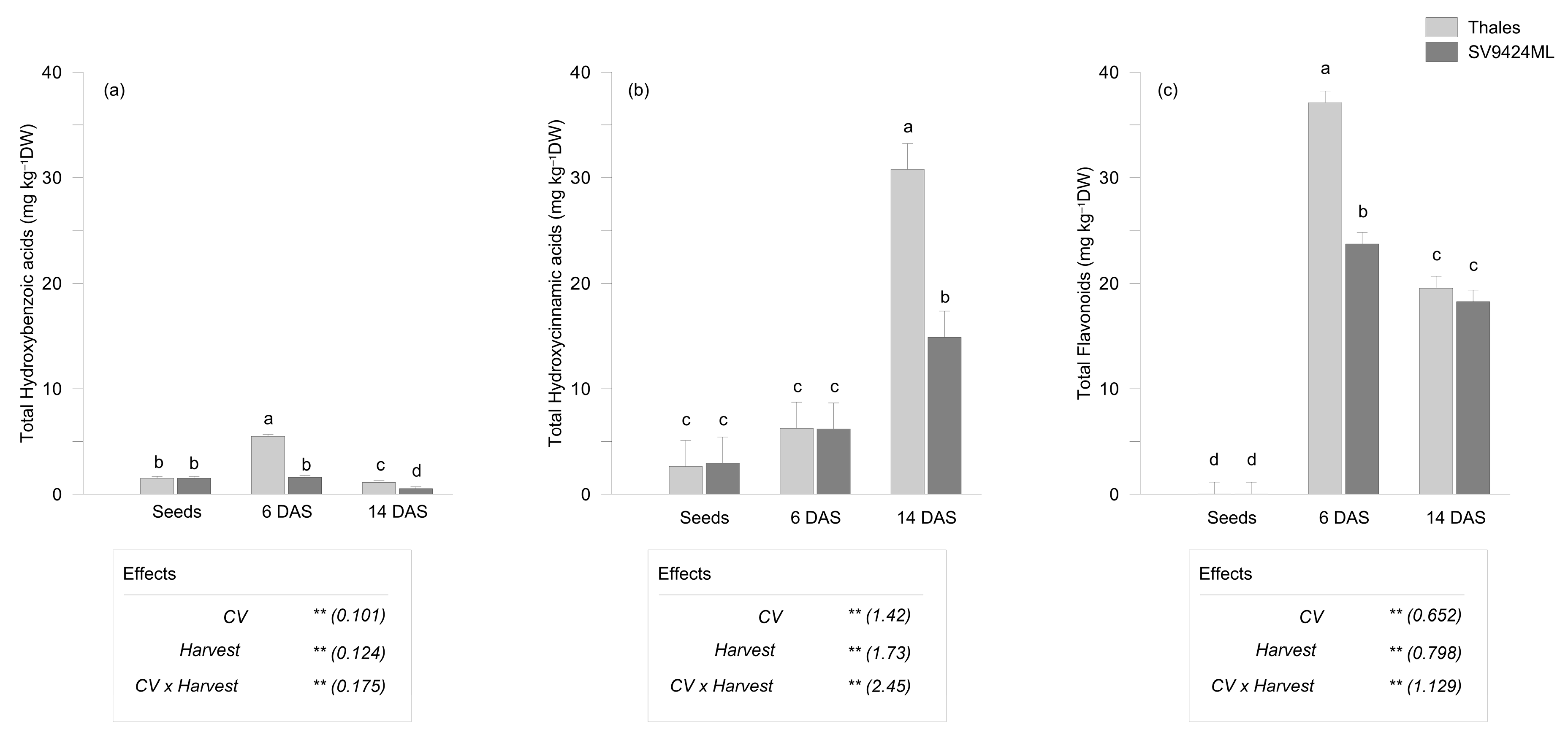
2.2. Phenolic Compounds Variation During Sprouting
3. Discussion
4. Materials and Methods
4.1. Plant Material and Sprouting Conditions
4.2. Chemicals
4.3. UPLC-ESI-MS/MS Analysis
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Rico, X.; Gullón, B.; Alonso, J.L.; Yáñez, R. Recovery of high value-added compounds from pineapple, melon, watermelon and pumpkin processing by-products: An overview. Food Res. Int. 2020, 132, 109086. [Google Scholar] [CrossRef]
- Silva, M.A.; Albuquerque, T.G.; Ferreira, D.M.; Alves, R.C.; Oliveira, M.B.P.; Costa, H.S. Nutritional and Bioactive Profiling of Cucumis melo L. By-Products: Towards a Circular Food Economy. Molecules 2025, 30, 1287. [Google Scholar] [CrossRef]
- Okuthe, G. Valorizing fruit and vegetable waste: The untapped potential for entrepreneurship in sub-saharan Africa—A systematic review. Recycling 2024, 9, 40. [Google Scholar] [CrossRef]
- de Medeiros, L.D.G.; de Carvalho, L.B.A.; Freitas, E.P.S.; Porto, D.L.; Aragão, C.F.S.; de Sousa Júnior, F.C.; da Silva Chaves Damasceno, K.S.F.; de Assis, C.F.; Araújo Morais, A.H.; Passos, T.S. Alternative flours from pulp melons (Cucumis melo L.): Seasonality influence on physical, chemical, technological parameters, and utilization in bakery product. Heliyon 2024, 10, e29609. [Google Scholar] [CrossRef]
- Silva, M.A.; Albuquerque, T.G.; Alves, R.C.; Oliveira, M.B.P.; Costa, H.S. Melon peel flour: Utilization as a functional ingredient in bakery products. Food Funct. 2024, 15, 1899–1908. [Google Scholar] [CrossRef]
- Mallek-Ayadi, S.; Bahloul, N.; Kechaou, N. Chemical composition and bioactive compounds of Cucumis melo L. seeds: Potential source for new trends of plant oils. Process Saf. Environ. Prot. 2018, 113, 68–77. [Google Scholar] [CrossRef]
- Galieni, A.; Falcinelli, B.; Stagnari, F.; Datti, A.; Benincasa, P. Sprouts and microgreens: Trends, opportunities, and horizons for novel research. Agronomy 2020, 10, 1424. [Google Scholar] [CrossRef]
- Zeb, A. Phenolic profile and antioxidant activity of melon (Cucumis melo L.) seeds from Pakistan. Foods 2016, 5, 67. [Google Scholar] [CrossRef]
- Khalid, W.; Ikram, A.; Rehan, M.; Afzal, F.A.; Ambreen, S.; Ahmad, M.; Aziz, A.; Sadiq, A. Chemical composition and health benefits of melon seed: A review. Pak. J. Agri. Res. 2021, 34, 309–317. [Google Scholar] [CrossRef]
- Saeed, F.; Afzaal, M.; Niaz, B.; Hussain, M.; Rasheed, A.; Raza, M.A.; Umar, M.; Khan, M.A.; Suleria, H.; Tufail, T.; et al. Comparative study of nutritional composition, antioxidant activity and functional properties of Cucumis melo and Citrullus lanatus seeds powder. Cogent Food Agric. 2024, 10, 2293517. [Google Scholar] [CrossRef]
- Kale, S.; Matthäus, B.; Aljuhaimi, F.; Ahmed, I.A.M.; Özcan, M.M.; Ghafoor, K.; Babiker, E.E.; Osman, M.A.; Gassem, M.A.; Alqah, H.A.S. Comparative study of the properties of 10 variety melon seeds and seed oils. J. Food Process. Preserv. 2020, 44, e14463. [Google Scholar] [CrossRef]
- Nerson, H. Seed production and germinability of cucurbit crops. Seed Sci. Biotechnol. 2007, 1, 1–10. [Google Scholar]
- Ebert, A.W. Sprouts and microgreens—Novel food sources for healthy diets. Plants 2022, 11, 57. [Google Scholar] [CrossRef]
- Bravi, E.; Falcinelli, B.; Mallia, G.; Marconi, O.; Royo-Esnal, A.; Benincasa, P. Effect of sprouting on the phenolic compounds, glucosinolates, and antioxidant activity of five Camelina sativa (L.) Crantz cultivars. Antioxidants 2023, 12, 1495. [Google Scholar] [CrossRef]
- Nerson, H. Fruit-set order affects seed yield and germinability in melon (Cucumis melo L.). J. Hortic. Sci. Biotechnol. 2004, 79, 985–990. [Google Scholar] [CrossRef]
- Amali, P.E.; Kortse, A.; Vange, T. The quality of egusi melon (Citrullus lanatus thumb matsum and Naki) seed derived from fruits harvested at different growth stages and different positions on the mother plant. Int. J. Sci. Res. Pub. 2013, 3, 1–7. [Google Scholar]
- Nerson, H. Effects of seed maturity, extraction practices and storage duration on germinability in watermelon. Sci. Hortic. 2002, 93, 245–256. [Google Scholar] [CrossRef]
- Marcos-Filho, J.; Bennett, M.A.; McDonald, M.B.; Evans, A.F.; Grassbaugh, E.M. Assessment of melon seed vigour by an automated computer imaging system compared to traditional procedures. Seed Sci. Technol. 2006, 34, 485–497. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, X.; Cui, Z.; Ding, Y.; Zhou, L.; Zhao, X. Cold plasma effects on the nutrients and microbiological quality of sprouts. Food Res. Int. 2022, 159, 111655. [Google Scholar] [CrossRef]
- Guragain, R.P.; Baniya, H.B.; Guragain, D.P.; Pradhan, S.P.; Subedi, D.P. From seed to sprout: Unveiling the potential of non-thermal plasma for optimizing cucumber growth. Heliyon 2023, 9, e21460. [Google Scholar] [CrossRef]
- Mallek-Ayadi, S.; Bahloul, N.; Kechaou, N. Phytochemical profile, nutraceutical potential and functional properties of Cucumis melo L. seeds. J. Sci. Food Agric. 2019, 99, 1294–1301. [Google Scholar] [CrossRef]
- Rolim, P.M.; Fidelis, G.P.; Padilha, C.E.A.; Santos, E.S.; Rocha, H.A.O.; Macedo, G.R. Phenolic profile and antioxidant activity from peels and seeds of melon (Cucumis melo L. var. reticulatus) and their antiproliferative effect in cancer cells. Braz. J. Med. Biol. Res. 2018, 51, e6069. [Google Scholar] [CrossRef]
- Araújo, K.T.A.; de Melo Queiroz, A.J.; de Figueirêdo, R.M. Germination on the nutritional properties of seeds of four melon varieties. Ciência Rural. 2023, 54, e20220307. [Google Scholar] [CrossRef]
- Ahmed, I.A.M.; Özcan, M.M.; Uslu, N.; Mohammed, B.M.; Albakry, Z. The effect of sprouting and roasting on bioactive compounds, phenolic and fatty acid profiles and some element contents of pumpkin seeds. J. Food Measurem. Characteriz. 2024, 18, 464–473. [Google Scholar] [CrossRef]
- Qian, H.; Liu, T.; Deng, M.; Miao, H.; Cai, C.; Shen, W.; Wang, Q. Effects of light quality on main health-promoting compounds and antioxidant capacity of Chinese kale sprouts. Food Chem. 2016, 196, 1232–1238. [Google Scholar] [CrossRef]
- Trasmundi, F.; Galieni, A.; Eugelio, F.; Fanti, F.; Benincasa, P.; Del Carlo, M.; Sergi, M.; Stagnari, F. Salt elicitation to enhance phytochemicals in durum wheat seedlings. J. Sci. Food Agric. 2024, 104, 249–256. [Google Scholar] [CrossRef]
- van Dam, N.M.; Tytgat, T.O.; Kirkegaard, J.A. Root and shoot glucosinolates: A comparison of their diversity, function and interactions in natural and managed ecosystems. Phytochem. Rev. 2009, 8, 171–186. [Google Scholar] [CrossRef]
- Martinez, V.; Mestre, T.C.; Rubio, F.; Girones-Vilaplana, A.; Moreno, D.A.; Mittler, R.; Rivero, R.M. Accumulation of flavonols over hydroxycinnamic acids favors oxidative damage protection under abiotic stress. Front. Plant Sci. 2016, 7, 838. [Google Scholar] [CrossRef]
- Saltveit, M.E. Synthesis and metabolism of phenolic compounds. In Fruit and Vegetable Phytochemicals: Chemistry and Human Health, 2nd ed.; Wiley Online Library: Hoboken, NJ, USA, 2017; pp. 115–124. [Google Scholar]
- Wang, Y.; Chantreau, M.; Sibout, R.; Hawkins, S. Plant cell wall lignification and monolignol metabolism. Front. Plant Sci. 2013, 4, 220. [Google Scholar] [CrossRef]
- Fahmy, M.I.; Sadek, M.A.; Abdou, K.; El-Dessouki, A.M.; El-Shiekh, R.A.; Khalaf, S.S. Orientin: A comprehensive review of a promising bioactive flavonoid. Inflammopharmacology 2025, 33, 1713–1728. [Google Scholar] [CrossRef]
- Sun, W.; Shahrajabian, M.H. Therapeutic potential of phenolic compounds in medicinal plants—Natural health products for human health. Molecules 2023, 28, 1845. [Google Scholar] [CrossRef]
- Benincasa, P.; Massoli, A.; Polegri, L.; Concezzi, L.; Onofri, A.; Tei, F. Optimising the use of plastic protective covers in field grown melon on a farm scale. Ital. J. Agron. 2014, 9, 8–18. [Google Scholar] [CrossRef]
- Oliva, E.; Viteritti, E.; Fanti, F.; Eugelio, F.; Pepe, A.; Palmieri, S.; Sergi, M.; Compagnone, D. Targeted and semi-untargeted determination of phenolic compounds in plant matrices by high performance liquid chromatography-tandem mass spectrometry. J. Chromatogr. A 2021, 1651, 462315. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 5 May 2024).

{kind=link}
{kind=link}
{kind=link}
| Cultivar | Harvest | RL (mm Sprout−1) | SL (mm Sprout−1) | FW (mg Sprout−1) | DM (%) |
|---|---|---|---|---|---|
| Thales | |||||
| Sprouts at 6 DAS | 101.3 ± 8.44 | 64.8 ± 2.52 | 240.2 ± 5.19 | 6.44 ± 0.025 | |
| Sprouts at 14 DAS | 126.2 ± 13.27 | 28.3 ± 1.09 | 180.4 ± 10.43 | 6.80 ± 0.001 | |
| Overall | 113.8 | 46.6 | 210.3 | 6.62 | |
| SV9424ML | |||||
| Sprouts at 6 DAS | 90.3 ± 2.18 | 68.7 ± 1.91 | 201.6 ± 13.73 | 6.31 ± 0.247 | |
| Sprouts at 14 DAS | 109.7 ± 7.30 | 23.0 ± 1.59 | 186.3 ± 2.40 | 5.82 ± 0.181 | |
| Overall | 100.0 | 45.8 | 193.9 | 6.07 | |
| F-test | |||||
| CV | n.s. | n.s. | n.s. | ** (0.153) | |
| Harvest | * (8.74) | ** (1.85) | ** (9.08) | n.s. | |
| CV × Harvest | n.s. | * (2.61) | * (12.84) | * (0.217) | |
| LSD | |||||
| CV | 19.03 | 4.03 | 19.79 | 0.33 | |
| Harvest | 19.03 | 4.03 | 19.79 | 0.33 | |
| CV × Harvest | 26.91 | 5.70 | 27.98 | 0.47 | |
| Effects | Hydroxybenzoic Acids (mg kg−1 DW × 102) | Hydroxycinnamic Acids (mg kg−1 DW ×102) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 4-OHBenzA | ProtA | GalA | VanA | SyrA | CafA | ChA | p-CouA | FerA | trans-CinA | |
| Thales | ||||||||||
| Seeds | 54.6 ± 0.49 | 30.5 ± 1.14 | 2.5 ± 0.09 | 36.6 ± 0.66 | 29.9 ± 0.71 | 30.5 ± 0.53 | 8.6 ± 0.26 | 133.5 ± 2.04 | 40.6 ± 0.57 | 51.6 ± 1.58 |
| Sprouts at 6 DAS | n.d. | 235.6 ± 20.71 | n.d. | 305.8 ± 3.68 | 6.5 ± 0.58 | 7.9 ± 0.10 | n.d. | 359.7 ± 8.17 | 216.0 ± 12.83 | 44.4 ± 4.35 |
| Sprouts at 14 DAS | n.d. | 25.0 ± 6.36 | n.d. | 83.2 ± 1.29 | 5.9 ± 0.72 | 13.3 ± 2.23 | n.d. | 2448.7 ± 322.23 | 582.6 ± 108.00 | 34.9 ± 4.90 |
| Overall | -- | 97.0 | -- | 141.9 | 14.1 | 17.2 | -- | 980.6 | 279.7 | 43.6 |
| SV9424ML | ||||||||||
| Seeds | 48.0 ± 2.51 | 37.9 ± 0.32 | 2.4 ± 0.08 | 35.1 ± 0.96 | 30.7 ± 0.87 | 40.2 ± 0.63 | 7.8 ± 0.23 | 145.5 ± 4.36 | 41.4 ± 0083 | 61.2 ± 3.81 |
| Sprouts at 6 DAS | n.d. | 1.02 ± 0.09 | n.d. | 154.8 ± 19.03 | 7.6 ± 0.97 | 7.6 ± 0.58 | n.d. | 350.5 ± 10.67 | 242.0 ± 13.22 | 20.9 ± 1.13 |
| Sprouts at 14 DAS | n.d. | 0.27 ± 0.02 | n.d. | 43.3 ± 1.19 | 12.7 ± 0.06 | 12.3 ± 1.96 | n.d. | 932.0 ± 82.56 | 530.4 ± 36.18 | 14.8 ± 1.25 |
| Overall | -- | 13.1 | -- | 77.7 | 17.0 | 20.1 | -- | 476.0 | 271.2 | 32.3 |
| F-test | ||||||||||
| CV | -- | ** (7.23) | -- | ** (6.50) | ** (0.58) | * (1.05) | -- | ** (110.98) | n.s. | ** (2.64) |
| Harvest | -- | ** (8.86) | -- | ** (7.96) | ** (0.71) | ** (1.28) | -- | ** (135.92) | ** (47.11) | ** (3.23) |
| CV × Harvest | -- | ** (12.52) | -- | ** (11.26) | ** (1.01) | ** (1.81) | -- | ** (192.23) | n.s. | ** (4.57) |
| LSD | ||||||||||
| CV | -- | 15.19 | -- | 13.65 | 1.22 | 2.20 | -- | 233.16 | 80.80 | 5.55 |
| Harvest | -- | 18.60 | -- | 16.72 | 1.50 | 2.69 | -- | 285.57 | 98.96 | 6.80 |
| CV × Harvest | -- | 26.31 | -- | 23.65 | 2.12 | 3.81 | -- | 403.85 | 139.96 | 9.61 |
| Effects | Flavones (mg kg−1 DW × 102) | Flavonols (mg kg−1 DW × 102) | Flavanones (mg kg−1 DW × 102) | ||||
|---|---|---|---|---|---|---|---|
| Api | Lut | Dios | Ori | Rut | Myr | Nar | |
| Thales | |||||||
| Seeds | n.d. | n.d. | n.d. | n.d. | 2.6 ± 0.35 | 1.4 ± 0.05 | 0.36 ± 0.036 |
| Sprouts at 6 DAS | 6.8 ± 0.40 | 261.4 ± 13.71 | 607.1 ± 48.12 | 2797.1 ± 64.58 | n.d. | n.d. | 38.2 ± 2.44 |
| Sprouts at 14 DAS | 5.3 ± 0.28 | 154.7 ± 18.86 | 462.7 ± 53.02 | 1299.6 ± 78.59 | n.d. | n.d. | 32.2 ± 4.52 |
| Overall | -- | -- | -- | -- | -- | -- | 23.6 |
| SV9424ML | |||||||
| Seeds | n.d. | n.d. | n.d. | n.d. | 2.3 ± 0.08 | 1.3 ± 0.02 | 0.12 ± 0.022 |
| Sprouts at 6 DAS | 4.3 ± 0.49 | 56.9 ± 7.24 | 257.4 ± 25.33 | 2026.6 ± 58.81 | n.d. | n.d. | 26.0 ± 1.57 |
| Sprouts at 14 DAS | 4.9 ± 0.09 | 200.5 ± 8.58 | 347.1 ± 19.82 | 1238.7 ± 58.42 | n.d. | n.d. | 33.7 ± 4.80 |
| Overall | -- | -- | -- | -- | -- | -- | 19.9 |
| F-test | |||||||
| CV | -- | -- | -- | -- | -- | -- | n.s. |
| Harvest | -- | -- | -- | -- | -- | -- | ** (2.94) |
| CV × Harvest | -- | -- | -- | -- | -- | -- | n.s. |
| LSD | |||||||
| CV | -- | -- | -- | -- | -- | -- | 5.04 |
| Harvest | -- | -- | -- | -- | -- | -- | 6.18 |
| CV × Harvest | -- | -- | -- | -- | -- | -- | 8.73 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galieni, A.; Falcinelli, B.; Stagnari, F.; Oliva, E.; Fanti, F.; Lorenzetti, M.C.; Benincasa, P. New Insights into Sprout Production from Melon (Cucumis melo L. var. reticulatus) Seeds as By-Product of Fruit Processing. Plants 2025, 14, 1896. https://doi.org/10.3390/plants14131896
Galieni A, Falcinelli B, Stagnari F, Oliva E, Fanti F, Lorenzetti MC, Benincasa P. New Insights into Sprout Production from Melon (Cucumis melo L. var. reticulatus) Seeds as By-Product of Fruit Processing. Plants. 2025; 14(13):1896. https://doi.org/10.3390/plants14131896
Chicago/Turabian StyleGalieni, Angelica, Beatrice Falcinelli, Fabio Stagnari, Eleonora Oliva, Federico Fanti, Maria Chiara Lorenzetti, and Paolo Benincasa. 2025. "New Insights into Sprout Production from Melon (Cucumis melo L. var. reticulatus) Seeds as By-Product of Fruit Processing" Plants 14, no. 13: 1896. https://doi.org/10.3390/plants14131896
APA StyleGalieni, A., Falcinelli, B., Stagnari, F., Oliva, E., Fanti, F., Lorenzetti, M. C., & Benincasa, P. (2025). New Insights into Sprout Production from Melon (Cucumis melo L. var. reticulatus) Seeds as By-Product of Fruit Processing. Plants, 14(13), 1896. https://doi.org/10.3390/plants14131896