


Effects of Bt Rice Straw Extract on Seed Germination and Plant Growth of Pakchoi: Novel Variables of Cropping System

Abstract
1. Introduction
2. Results
2.1. Effects of Different Rice Straw Extracts on Seed Germination Rate and Seedling Growth of Pakchoi
2.2. Effects of Different Rice Straw Extracts on Protective Enzyme Activities of Pakchoi Seedlings
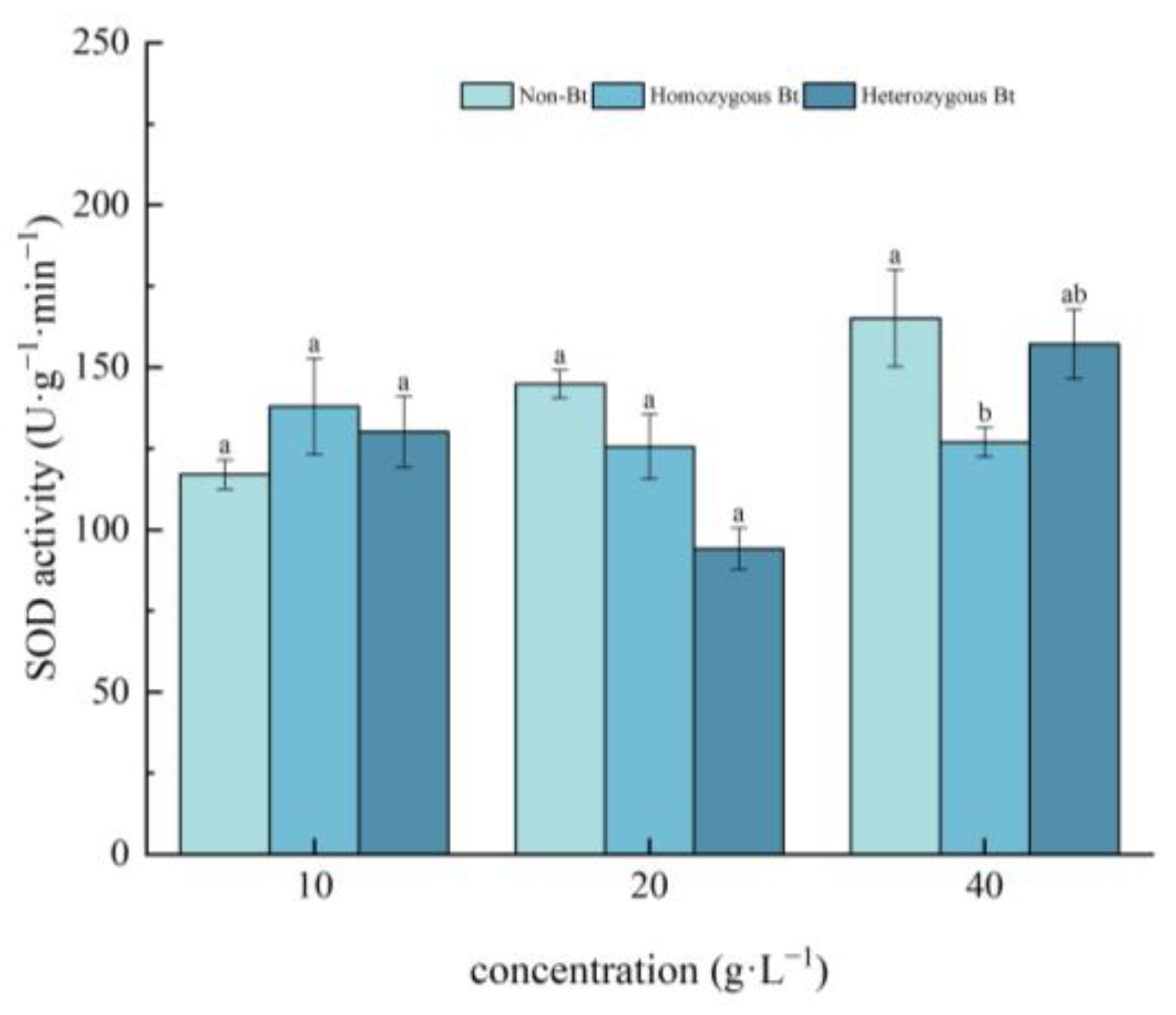
2.2.1. Effects on SOD Activity of Pakchoi Seedlings
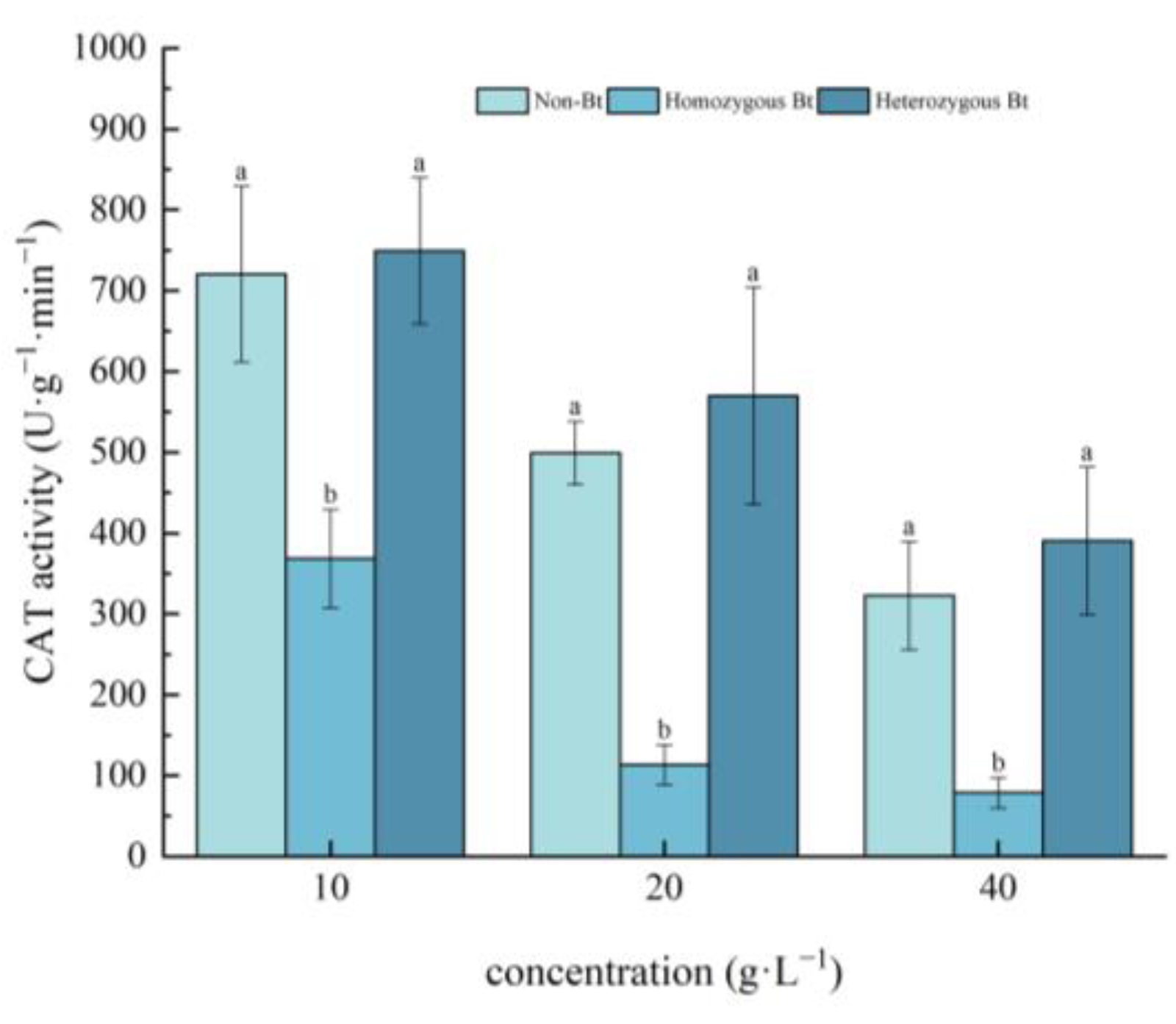
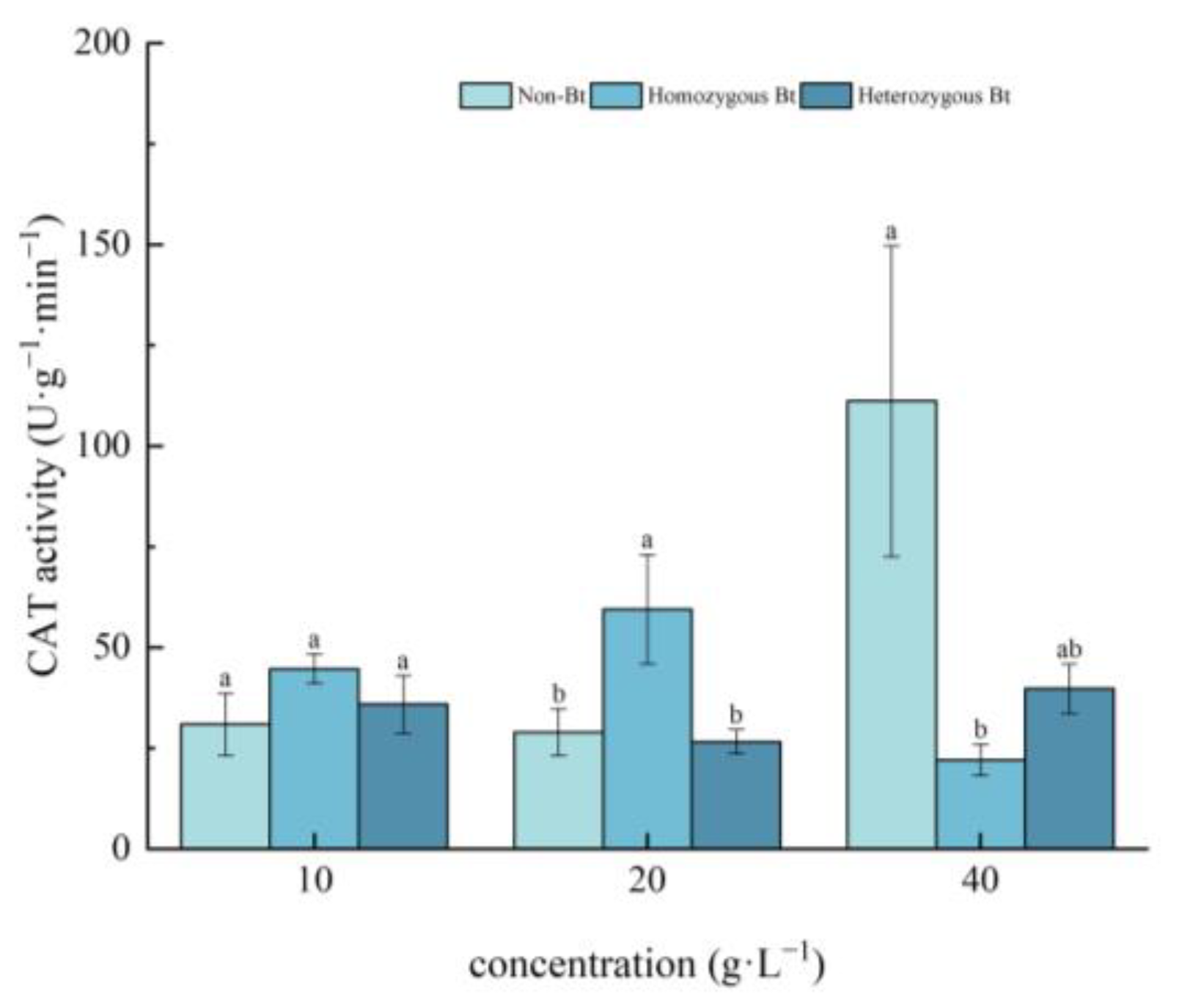
2.2.2. Effects on CAT Activity of Pakchoi Seedlings
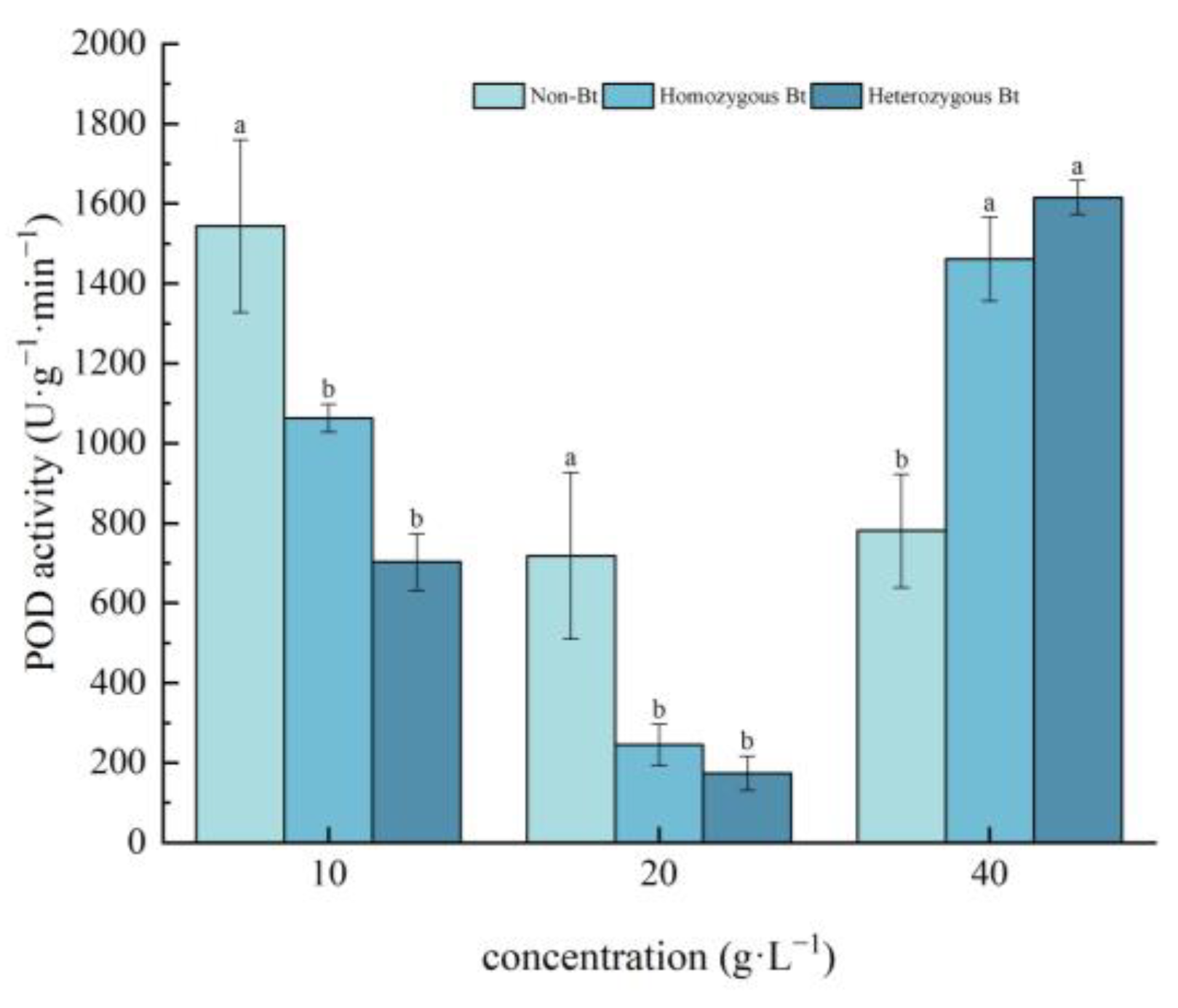
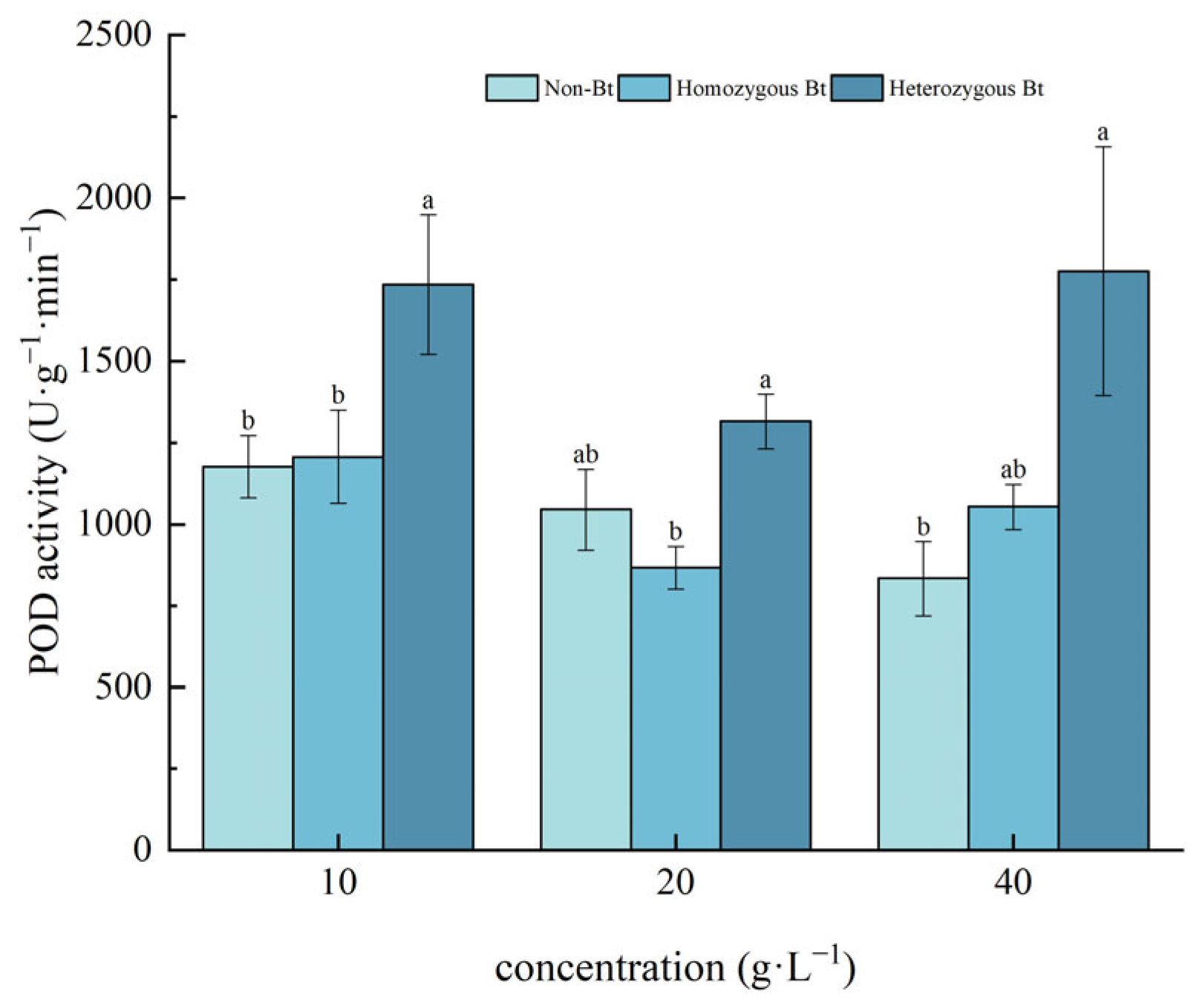
2.2.3. Effects on POD Activity of Pakchoi Seedlings
2.3. Effects of Different Rice Straw Extracts on Growth of Pakchoi
3. Discussion
4. Materials and Methods
4.1. Test Materials
4.2. Preparation of Extract
4.3. Seed Germination Test
4.4. Plant Growth Test
4.5. Measurement Items
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rayee, R.; Anh, L.H.; Khanh, T.D.; Xuan, T.D. Potential Momilactones in Rice Stress Tolerance and Health Advantages. Agronomy 2024, 14, 405. [Google Scholar] [CrossRef]
- Liu, Z.H.; Tian, Y.; Chen, H.N.; Zhou, Z.H.; Zheng, J.; Yang, X.H. Research Progress and Application Status of Transgenic Breeding in Rice. China Seed Ind. 2023, 11–17. [Google Scholar] [CrossRef]
- Liu, H.J. Screening of Endogenous miRNAs in Chilo suppressalis and Development of Transgenic Insect-Resistant Rice. Master’s Thesis, Huazhong Agricultural University, Wuhan, China, 2022. [Google Scholar]
- Agbio Investor. Global GM Crop Area Review. Available online: https://gm.agbioinvestor.com (accessed on 26 February 2025).
- Hu, Y.T.; Tian, C.B.; Feng, Y.J.; Ma, W.D.; Zhang, Y.J.; Yang, Q.; Zhang, X.R. Transgenic early japonica rice: Integration and expression characterization of stem borer resistance Bt gene. Gene 2024, 927, 148753. [Google Scholar] [CrossRef]
- Wang, K.X.; Zhang, K.R.; Cao, C.G.; Jiang, Y. Effect of Bt traits on transgenic rice’s growth and weed competitiveness. J. Integr. Agric. 2023, 22, 2346–2358. [Google Scholar] [CrossRef]
- Song, Z.Y.; Lin, X.F.; Yan, Y.Z.; Jin, Y.M. Overview of Research on Insect-Resistant Transgenic Rice and Its Safety. Agric. Technol. 2020, 40, 15–16. [Google Scholar]
- Li, C.M.; Han, G.J.; Liu, Q.; Qi, J.H.; Xu, J. Cultivation Characteristics and Insecticidal Activity Analysis of Bacillus thuringiensis Bt-8. J. South. Agric. 2016, 47, 2072–2077. [Google Scholar]
- Sujayanand, G.K.; Pandey, S.; Jagadeeswaran, R.; Chandra, A.; Kumar, V.; Dubey, S.; Dubey, J. Characterization of entomotoxic and nematotoxic genes from indigenous Bacillus thuringiensis strains and their biocontrol potential. Egypt. J. Biol. Pest Control 2023, 33, 88. [Google Scholar]
- Wang, Y.Y.; Dai, P.L.; Chen, X.P.; Romeis, J.; Shi, J.R.; Peng, Y.F.; Li, Y.H. Ingestion of Bt rice pollen does not reduce the survival or hypopharyngeal gland development of Apis mellifera adults. Environ. Toxicol. Chem. 2017, 36, 1243–1248. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wu, L.H.; Knauth, S.; Eickhorst, T. Degradation of transgenic Bt-rice straw incorporated with two different paddy soils. J. Environ. Manag. 2019, 244, 415–421. [Google Scholar] [CrossRef]
- Yaqoob, A.; Shahid, A.A.; Samiullah, T.R.; Rao, A.Q.; Khan, M.A.U.; Thair, S.; Mirza, S.A.; Husnain, T. Risk assessment of Bt crops on the non-target plant-associated insects and soil organisms. J. Sci. Food Agric. 2016, 96, 2613–2619. [Google Scholar] [CrossRef]
- Li, Y.J.; Wang, C.; Ge, L.; Hu, C.; Wu, G.G.; Sun, Y.; Song, L.L.; Wu, X.; Pan, A.H.; Xu, Q.Q.; et al. Environmental Behaviors of Bacillus thuringiensis (Bt) Insecticidal Proteins and Their Effects on Microbial Ecology. Plants 2022, 11, 1212. [Google Scholar] [CrossRef]
- Arpaia, S.; Birch, A.N.E.; Kiss, J.; Loon, J.J.A.V.; Messean, A.; Nuti, M.; Perry, J.N.; Sweet, J.B.; Tebbe, C.C. Assessing environmental impacts of genetically modified plants on non-target organisms: The relevance of in planta studies. Sci. Total Environ. 2017, 583, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Alengebawy, A.; Mohamed, B.A.; Ran, Y.; Yang, Y.; Pezzuolo, A.; Samer, M.; Ai, P. A comparative environmental life cycle assessment of rice straw-based bioenergy projects in China. Environ. Res. 2022, 212, 113404. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ding, Y.Q.; Yue, B.W. Research Progress on Direct Straw Returning Technology and Equipment in the Northeast Rice Region. China Rice 2024, 30, 37–42. [Google Scholar]
- Pathak, H.; Singh, R.; Bhatia, A.; Jain, N. Recycling of rice straw to improve wheat yield and soil fertility and reduce atmospheric pollution. Paddy Water Environ. 2006, 4, 111–117. [Google Scholar] [CrossRef]
- Chen, H.G.; Cao, Q.G.; Xiong, G.L.; Li, W.; Zhang, A.X.; Yu, H.S.; Wang, J.S. Composition of Wheat Rhizosphere Antagonistic Bacteria and Wheat Sharp Eyespot as Affected by Rice Straw Mulching. Pedosphere 2010, 20, 505–514. [Google Scholar] [CrossRef]
- Wang, F.R. Effects of Direct Straw Returning and Carbonization Returning on the Stoichiometric Ratios of Carbon, Nitrogen, and Phosphorus in Paddy Soil Aggregates and the Utilization of Nitrogen and Phosphorus Nutrients in Rice. Master’s Thesis, Gansu Agricultural University, Lanzhou, China, 2023. [Google Scholar]
- Che, Y.X.; Zhang, B.Y.; Liu, B.Y.; Wang, J.C.; Zhang, H.L. Effects of Straw Return Rate on Soil Physicochemical Properties and Yield in Paddy Fields. Agronomy 2024, 14, 1668. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, A.; Singh, S.P.; Ellur, R.K.; Choudhary, V.; Sarkel, S.; Singh, D.; Krishnan, S.G.; Nagarajan, M.; Vinod, K.K.; et al. Incorporation of blast resistance into “PRR78”, an elite Basmati rice restorer line, through marker assisted backcross breeding. Field Crop. Res. 2012, 128, 8–16. [Google Scholar] [CrossRef]
- Liu, Y.M.; Liu, Q.S.; Wang, Y.A.; Chen, X.P.; Song, X.Y.; Romeis, J.; Li, Y.H.; Peng, Y.F. Ingestion of Bt corn pollen containing Cry1Ab/2Aj or Cry1Ac does not harm Propylea japonica larvae. Sci. Rep. 2016, 6, 23507. [Google Scholar] [CrossRef]
- Wang, B.F.; Wu, F.C.; Yin, J.Q.; Jiang, Z.L.; Song, X.Y.; Reddy, V.P. Use of Taxonomic and Trait-Based Approaches to Evaluate the Effect of Bt maize Expressing Cry1Ie Protein on Non-Target Collembola: A Case Study in Northeast China. Insects 2021, 12, 88. [Google Scholar] [CrossRef]
- Li, P.; Ye, S.F.; Chen, J.; Wang, L.Y.; Li, Y.J.; Wu, G.G.; Song, L.L.; Wang, C.; Sun, Y.; Wang, J.B.; et al. Combined metagenomic and metabolomic analyses reveal that Bt rice planting alters soil C-N metabolism. ISME Commun. 2023, 3, 4. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Chen, G.W.; Chen, X.Y.; Wang, X.F.; Lu, Y.W.; Liang, Z.H.; Xu, J.F.; Peng, C. Analysis of the Unintended Effects of the Bacillus thuringiensis Insecticidal Protein in Genetically Modified Rice Using Untargeted Transcriptomics. Processes 2023, 11, 3202. [Google Scholar] [CrossRef]
- Li, C. Complexity and Density of Rhizosphere Microbiota in Bt Insect-Resistant Rice Remain Unchanged. China Science Daily, 9 August 2021. [Google Scholar]
- Qiu, L.M.; Liu, Q.Q.; Lin, R.K.; Shi, L.Q.; Zhan, Z.X. Research Progress on the Ecological Safety of Bt-Transgenic Rice on Major Non-Target Insects. Fujian J. Agric. Sci. 2018, 33, 326–333. [Google Scholar]
- Wu, Y.; Weng, Z.J.; Yan, H.X.; Yao, Z.T.; Li, Z.Z.; Sun, Y.J.; Ma, K.S.; Huil, J.J.; Zhang, D.L.; Ma, W.H.; et al. The microRNA-7322-5p/p38/Hsp19 axis modulates Chilo suppressalis cell-defences against Cry1Ca: An effective target for a stacked transgenic rice approach. Plant Biotechnol. J. 2023, 21, 1827–1838. [Google Scholar] [CrossRef]
- Sun, C. Determination and Analysis of the Hazard Quotient (HQ) of Three Bt Rice Varieties on Non-Target Organisms. Master’s Thesis, Zhejiang University, Hangzhou, China, 2019. [Google Scholar]
- Lu, H.H.; Wu, W.X.; Chen, Y.X.; Wang, H.L.; Devare, M.; Thies, J.E. Soil microbial community responses to Bt transgenic rice residue decomposition in a paddy field. J. Soils Sediments 2010, 10, 1598–1605. [Google Scholar] [CrossRef]
- Fang, H.; Dong, B.; Yan, H.; Tang, F.F.; Wang, B.C.; Yu, Y.L. Effect of vegetation of transgenic Bt rice lines and their straw amendment on soil enzymes, respiration, functional diversity and community structure of soil microorganisms under field conditions. J. Environ. Sci. 2012, 24, 1259–1270. [Google Scholar] [CrossRef] [PubMed]
- Raubuch, M.; Behr, K.; Roose, K.; Joergensen, R.G. Specific respiration rates, adenylates, and energy budgets of soil microorganisms after addition of transgenic Bt-maize straw. Pedobiologia 2010, 53, 191–196. [Google Scholar] [CrossRef]
- Čerevková, A.; Miklisová, D.; Szoboszlay, M.; Tebbe, C.C.; Cagan, L. The responses of soil nematode communities to Bt maize cultivation at four field sites across Europe. Soil Biol. Biochem. 2018, 119, 194–202. [Google Scholar] [CrossRef]
- Wang, S. Effects of Straw Returning on Seed Germination and Maize Seedling Growth. Master’s Thesis, Northeast Agricultural University, Harbin, China, 2021. [Google Scholar]
- Nakano, H.; Morita, S.; Shigemori, H.; Hasegawa, K. Plant Growth Inhibitory Compounds from Aqueous Leachate of Wheat Straw. Plant Growth Regul. 2006, 48, 215–219. [Google Scholar]
- Tang, Z.L.; Guo, R.Y.; Yang, X.; Min, J.Y.; He, P.Y.; Huang, X.Y.; Huang, K.F. Appropriate Concentration of Rice Straw Decomposition Liquid Prompted Germination and Seedling Growth of Common Buckwheat. J. Soil Sci. Plant Nutr. 2024, 24, 3861–3872. [Google Scholar] [CrossRef]
- Dong, S.Q.; Yuan, X.Y.; Hu, C.Y.; Wen, Y.Y.; Guo, P.Y.; Nie, L.Y.; Qiao, X.F. Effects of Foxtail Millet Straw Aqueous Extract on Allelopathy in Winter Wheat Seedlings. J. China Agric. Univ. 2015, 20, 31–38. [Google Scholar]
- Crawford, L.E.; Ii, M.M.W.; Wortman, S.E. An Early-Killed Rye (Secale cereale) Cover Crop Has Potential for Weed Management in Edamame (Glycine max). Weed Sci. 2018, 66, 502–507. [Google Scholar] [CrossRef]
- Yang, C.J.; Liu, R.Q.; Gu, Y.Y. Allelopathic Effects of Chokecherry Leaf Aqueous Extract on Four Plant Species. J. High. Norm. Sci. 2024, 44, 87–91. [Google Scholar]
- Tang, M.M.; Dong, N.; Liang, Z.H.; Zhang, T.H.; Li, S.M. Effects of Basil Extract on Wheat Seedling Growth. Anhui Agric. Bull. 2023, 29, 1–4. [Google Scholar]
- Li, B.; Wu, W.W.; Shen, W.Y.; Xiong, F.; Wang, K.H. Allelochemicals Released from Rice Straw Inhibit Wheat Seed Germination and Seedling Growth. Agronomy 2024, 14, 2376. [Google Scholar] [CrossRef]
- Xie, X.L.; Jiang, B.; Zhou, X.R.; Zhao, L.X.; Wang, J.H.; Shang, J. Allelopathic Effects of Aqueous Extract of Torreya fargesii Aril on Seed Germination and Seedling Growth of Radish, Mustard and Cabbage. Agric. Biotechnol. 2016, 5, 11–17. [Google Scholar]
- Qian, P.Z.; Sha, W.; Ma, T.Y.; Dong, J.H.; Wang, T.; Ao, J.; Zhang, J.; Zhang, M.J. Effects of Peony Seed Extract on Seed Germination and Seedling Growth of Two Plant Species. Mol. Plant Breed. 2023, 21, 642–648. [Google Scholar]
- Guo, H.G.; Ai, T.S.; Sun, Q.; Jin, D.; Sun, W.X.; Wu, Y.B. Effects of Different Crop Straw Extracts on Rapeseed Seed Germination and Seedling Growth. Mol. Plant Breed. 2024, 1–13. Available online: http://kns.cnki.net/kcms/detail/46.1068.S.20240514.1102.004.html (accessed on 26 February 2025).
- Feng, W.M.; Zhang, J.S.; Jiao, N.; Chen, H.F.; Yang, J.T.; Gao, D.D.; Guo, P.H. Effects of Codonopsis pilosula Straw Water Extract on Seed Germination and Seedling Growth of Medicinal Plants. Henan Agric. Sci. 2024, 53, 57–64. [Google Scholar]
- Liu, Y.P.; Xi, K.P.; Wang, H.J.; Wang, M.; Wu, L.L. Allelopathic Effects of Cotton Straw Water Extract on Lettuce. Shanxi Agric. Sci. 2018, 46, 1643–1645. [Google Scholar]
- Zhao, X.L.; Li, J.; Gu, W.R.; Ai, J.G.; Kong, L.Z.; Qiao, T.C.; Wei, S. Effects of Maize Straw Decomposition Solution on Seed Germination under Continuous Cropping. Crops 2013, 137–141. [Google Scholar] [CrossRef]
- Liu, X.M.; Meng, Q.M.; Wang, X.Q.; Wang, G.Q. Allelopathic Effects of Water Extracts from Different Parts of Maize Straw on Shepherd’s Purse. Hebei Agric. Sci. 2013, 17, 36–39. [Google Scholar]
- Shen, X.F.; Hu, X.Y.; Chen, Y.; Li, Y.B.; Luo, Q.Y. Effects of Rice Straw Extract on Seed Germination and Seedling Growth of Sweet Corn. Crops 2015, 158–160. [Google Scholar] [CrossRef]
- Li, F.Y.; Sun, X.F.; Feng, W.Q.; Qin, Y.S.; Wang, C.Q.; Tu, S.H. Allelopathic Effects of Rice Straw Water Extract on Wheat. Southwest China J. Agric. Sci. 2008, 21, 960–964. [Google Scholar]
- Zhang, B.; Yu, J.H.; Xie, J.M.; Feng, Z.; Zhang, G.B. Effects of Decomposed Maize Straw Water Extract on Germination Characteristics of Cucumber Seeds. J. Gansu Agric. Univ. 2012, 47, 82–87. [Google Scholar]
- Hua, Z.R.; Li, X.L. Allelopathic Effects of Maize Straw Water Extract on Shangmai 5226. J. Jiangxi Agric. 2017, 29, 28–32. [Google Scholar]
- Zhang, W.; Lu, L.Y.; Hu, L.Y.; Cao, W.; Sun, K.; Sun, Q.B.; Siddikee, A.; Shi, R.H.; Dai, C.C. Evidence for the Involvement of Auxin, Ethylene and ROS Signaling During Primary Root Inhibition of Arabidopsis by the Allelochemical Benzoic Acid. Plant Cell Physiol. 2018, 59, 1889–1904. [Google Scholar] [CrossRef]
- Liu, R.X.; Zhang, G.W.; Yang, C.Q.; Zhang, L.; Ni, W.C. Allelopathic Effects of Wheat Straw Extract and Decomposition Solution on Germination and Seedling Growth of Cotton. J. Cotton Sci. 2016, 28, 375–383. [Google Scholar]
- Elisante, F.; Tarimo, M.T.; Ndakidemi, P.A. Allelopathic effect of seed and leaf aqueous extracts of datura stramonium on leaf chlorophyll content, shoot and root elongation of Cenchrus ciliaris and Neonotonia wightii. Am. J. Plant Sci. 2013, 4, 2332. [Google Scholar] [CrossRef]
- Jaballah, S.B.; Zribi, I.; Haouala, R. Physiological and biochemical responses of two lentil varieties to chickpea (Cicer arietinum L.) aqueous extracts. Sci. Hortic. 2017, 225, 74–80. [Google Scholar] [CrossRef]
- Gong, Z.Y. Effects of Straw Residue Return on Cucumber Seedling Growth and Soil Microbial Community. Master’s Thesis, Northeast Agricultural University, Harbin, China, 2016. [Google Scholar]
- Chung, W.H. Unraveling new functions of superoxide dismutase using yeast model system: Beyond its conventional role in superoxide radical scavenging. J. Microbiol. 2017, 55, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Sui, L.H.; Huang, Y.Z.; Geng, C.M.; Yin, B.H. Physiological and visible injury responses in different growth stages of winter wheat to ozone stress and the protection of spermidine. Atmos. Pollut. Res. 2015, 6, 596–604. [Google Scholar] [CrossRef]
- Hu, M.J.; Zhu, Y.Y.; Liu, G.S.; Gao, Z.Y.; Li, M.; Su, Z.H.; Zhang, Z.K. Inhibition on anthracnose and induction of defense response by nitric oxide in pitaya fruit. Sci. Hortic. 2019, 245, 224–230. [Google Scholar] [CrossRef]
- Zhao, Y. Study on the Activity of Endogenous Inhibitory Substances in Paris polyphylla Seeds and Their Identification by GC-MS. Master’s Thesis, Sichuan Agricultural University, Ya’an, China, 2018. [Google Scholar]
- Chen, Y.; Wang, H.Y.; Lv, X.F.; Ma, H.J.; Li, Q.Z.; Cui, J.H. Effects of Corn Straw Extract on Seed Germination and Seedling Physiological Metabolism of Three Crops. Maize Sci. 2016, 24, 98–104. [Google Scholar]
- Wang, X.K. Principles and Techniques of Plant Physiology and Biochemistry Experiments, 3rd ed.; Higher Education Press: Beijing, China, 2015. [Google Scholar]
- Gao, J.F. Guide to Plant Physiology Experiments; Higher Education Press: Beijing, China, 2006. [Google Scholar]
- Li, H.S. Principles and Techniques of Plant Physiology and Biochemistry Experiments; Higher Education Press: Beijing, China, 2003. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Concentration (g·L−1) | Germination Rate (%) | Root Length (cm) | Seedling Height (cm) | Dry Weight (g) |
|---|---|---|---|---|---|
| T775 | 10 | 83 ± 3.42 a | 3.1 ± 0.22 b | 0.86 ± 0.01 a | 2.33 ± 0.14 a |
| F1 | 86 ± 4.16 a | 3.81 ± 0.25 a | 0.89 ± 0.11 a | 2.23 ± 0.08 a | |
| Tianyouhuazhan | 89 ± 1.92 a | 3.92 ± 0.14 a | 0.77 ± 0.05 a | 2.21 ± 0.1 a | |
| T775 | 20 | 77 ± 6.81 a | 3.09 ± 0.51 a | 1.13 ± 0.19 a | 2.04 ± 0.34 a |
| F1 | 78 ± 1.16 a | 4.63 ± 0.27 a | 1.02 ± 0.07 a | 2.53 ± 0.11 a | |
| Tianyouhuazhan | 82 ± 9.02 a | 3.61 ± 0.58 a | 1.02 ± 0.12 a | 3.26 ± 0.53 a | |
| T775 | 40 | 74 ± 2.58 a | 2.05 ± 0.13 ab | 0.88 ± 0.07 b | 2.2 ± 0.14 a |
| F1 | 77 ± 1.00 a | 2.95 ± 0.44 a | 1.16 ± 0.04 a | 2.53 ± 0.04 a | |
| Tianyouhuazhan | 56 ± 4.32 b | 1.41 ± 0.21 b | 0.75 ± 0.06 b | 2.53 ± 0.12 a |
| Treatment | Concentration (g·L−1) | Plant Height (cm) | Chlorophyll (mg·g−1) | Aboveground Dry Weight (g) | Underground Dry Weight (g) |
|---|---|---|---|---|---|
| T775 | 10 | 30.16 ± 1.38 a | 1.12 ± 0.21 a | 5.48 ± 0.49 a | 0.34 ± 0.04 a |
| F1 | 30.15 ± 1.41 a | 1.37 ± 0.09 a | 4.45 ± 0.43 ab | 0.23 ± 0.01 b | |
| Tianyouhuazhan | 27.2 ± 1.2 a | 1.06 ± 0.06 a | 4.11 ± 0.3 b | 0.21 ± 0.02 b | |
| T775 | 20 | 28.41 ± 1.89 ab | 1.03 ± 0.18 a | 4.02 ± 0.71 b | 0.34 ± 0.18 a |
| F1 | 26.95 ± 1.05 b | 1.2 ± 0.04 a | 4.35 ± 0.42 ab | 0.23 ± 0.04 a | |
| Tianyouhuazhan | 31.15 ± 1.08 a | 1.18 ± 0.12 a | 5.92 ± 0.62 a | 0.25 ± 0.03 a | |
| T775 | 40 | 29.38 ± 1.9 a | 1.06 ± 0.12 a | 4.48 ± 0.84 a | 0.19 ± 0.03 a |
| F1 | 27.89 ± 1.91 a | 1.24 ± 0.12 a | 4.34 ± 0.49 a | 0.18 ± 0.03 a | |
| Tianyouhuazhan | 29.84 ± 0.63 a | 1.53 ± 0.17 a | 5.04 ± 0.21 a | 0.25 ± 0.03 a |
| Index | Non-Bt | Homozygous Bt | Heterozygous Bt |
|---|---|---|---|
| Bt Protein (ng·g−1) | =LOD | 1405.21 ± 77.21 a | 1159.42 ± 38.97 b |
| Total Nitrogen (mg·L−1) | 5.58 ± 0.50 a | 4.29 ± 0.50 a | 5.01 ± 0.29 a |
| Total Phosphorus (mg·L−1) | 15.10 ± 0.65 b | 25.52 ± 1.86 a | 13.14 ± 1.02 b |
| Total Potassium (mg·L−1) | 71.19 ± 1.14 a | 24.62 ± 0.64 c | 31.46 ± 0.22 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Suo, W.; Pan, Y.; Feng, Y. Effects of Bt Rice Straw Extract on Seed Germination and Plant Growth of Pakchoi: Novel Variables of Cropping System. Plants 2025, 14, 1797. https://doi.org/10.3390/plants14121797
Zhang C, Suo W, Pan Y, Feng Y. Effects of Bt Rice Straw Extract on Seed Germination and Plant Growth of Pakchoi: Novel Variables of Cropping System. Plants. 2025; 14(12):1797. https://doi.org/10.3390/plants14121797
Chicago/Turabian StyleZhang, Chenning, Wenfang Suo, Yongfang Pan, and Yuanjiao Feng. 2025. "Effects of Bt Rice Straw Extract on Seed Germination and Plant Growth of Pakchoi: Novel Variables of Cropping System" Plants 14, no. 12: 1797. https://doi.org/10.3390/plants14121797
APA StyleZhang, C., Suo, W., Pan, Y., & Feng, Y. (2025). Effects of Bt Rice Straw Extract on Seed Germination and Plant Growth of Pakchoi: Novel Variables of Cropping System. Plants, 14(12), 1797. https://doi.org/10.3390/plants14121797