
A Herpetosiphon llansteffanensis Strain from Forest Soil Exhibits Biocontrol Activity Against Pear Fire Blight

Abstract
1. Introduction
2. Results
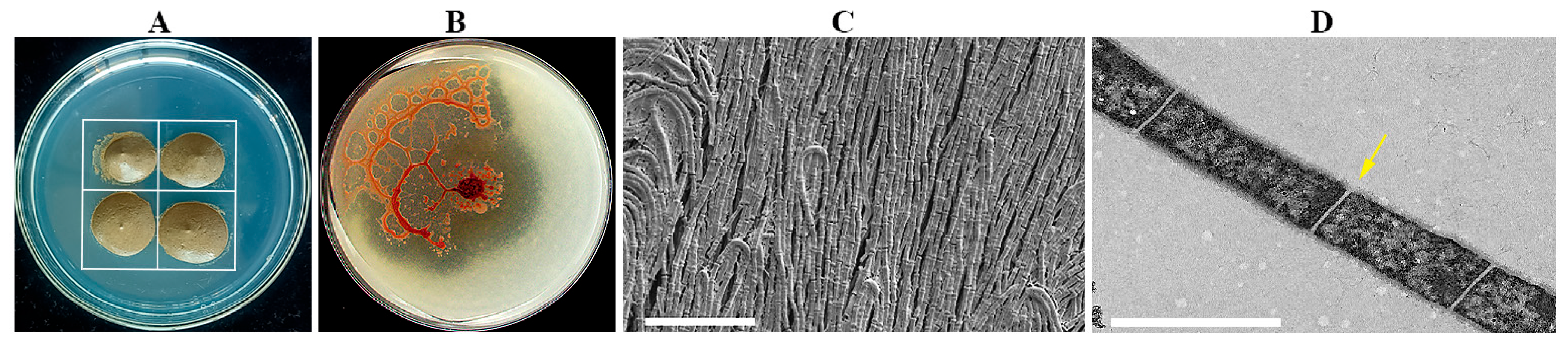
2.1. Characterization of Strain NSD29
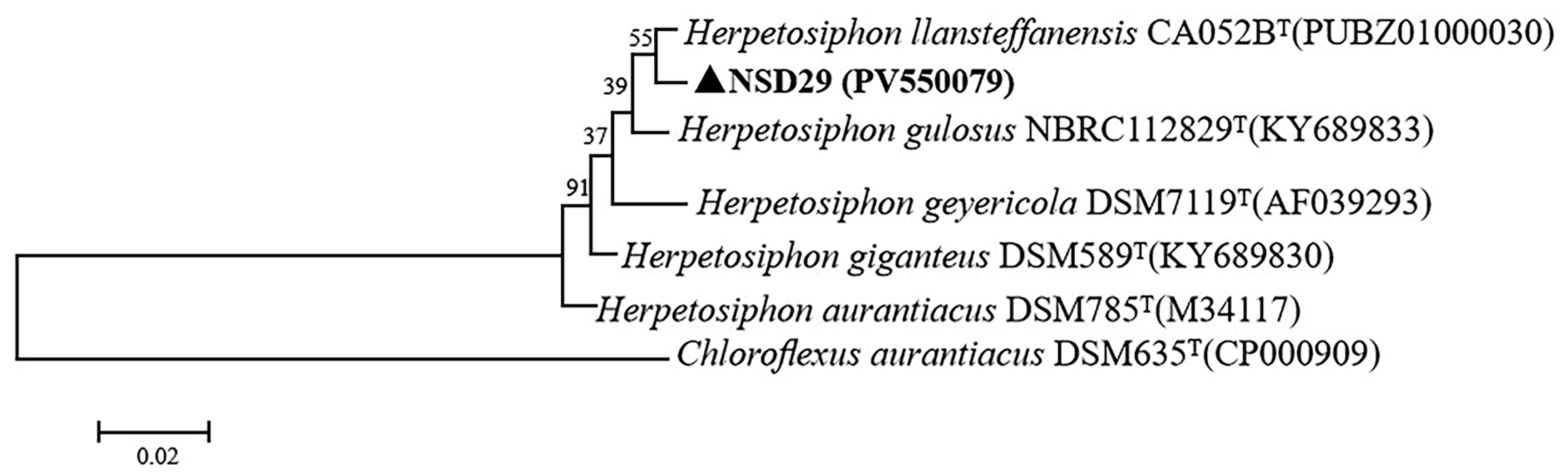
2.2. 16S rRNA Sequence Analysis
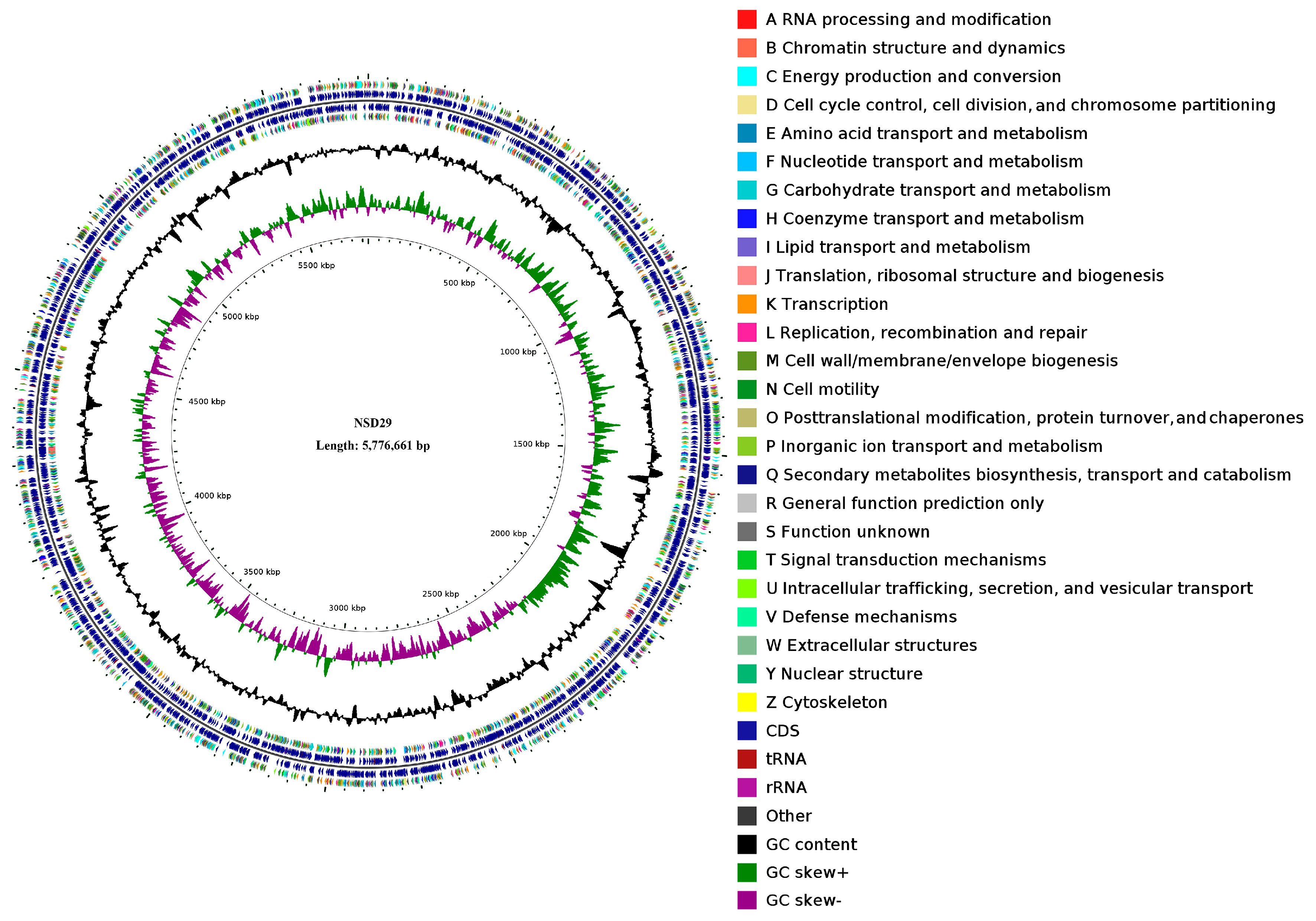
2.3. Whole-Genome Features of the Strain NSD29
2.4. Predatory Activity of the Strain NSD29 Against Plant Pathogenic Bacteria
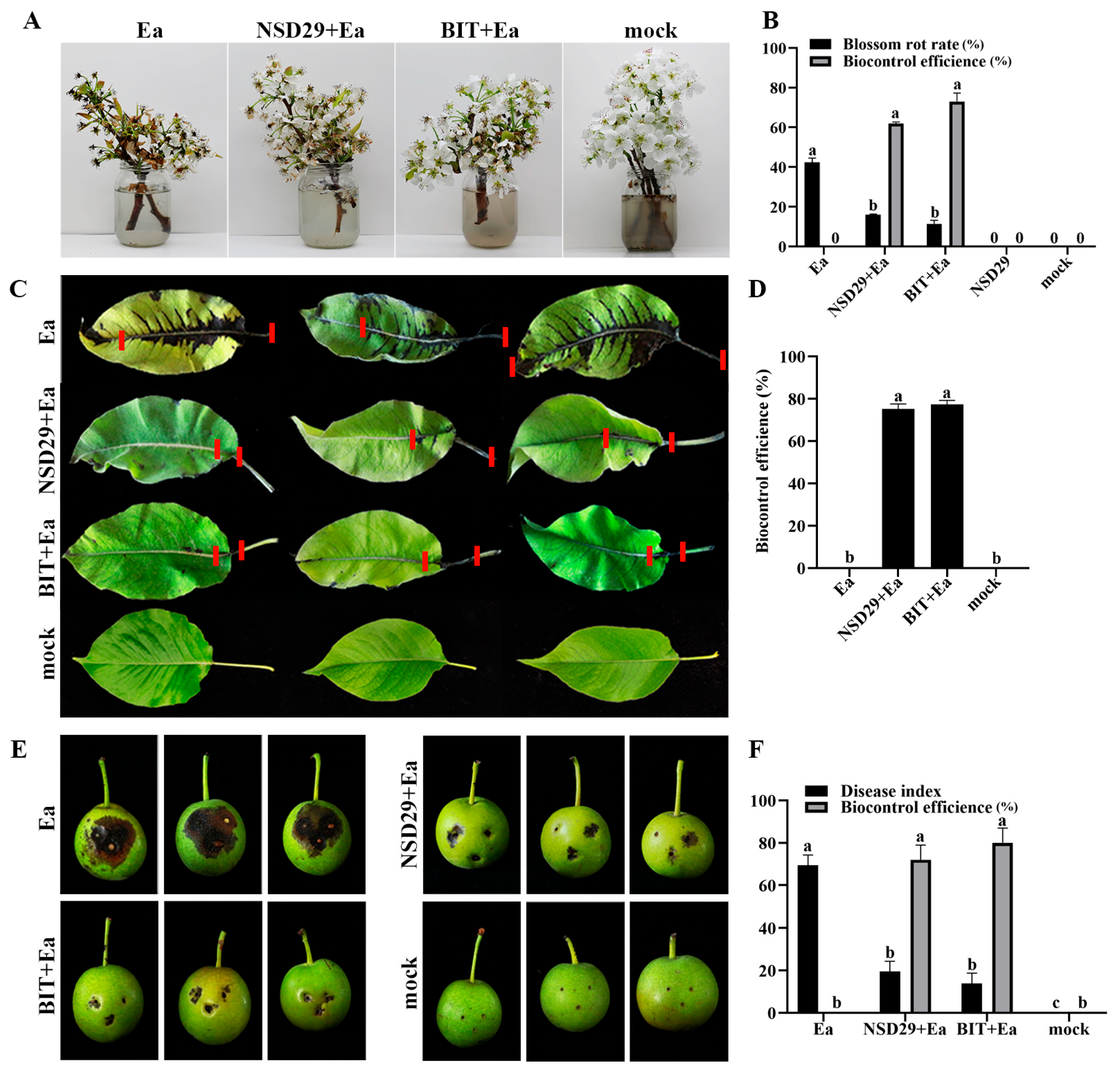
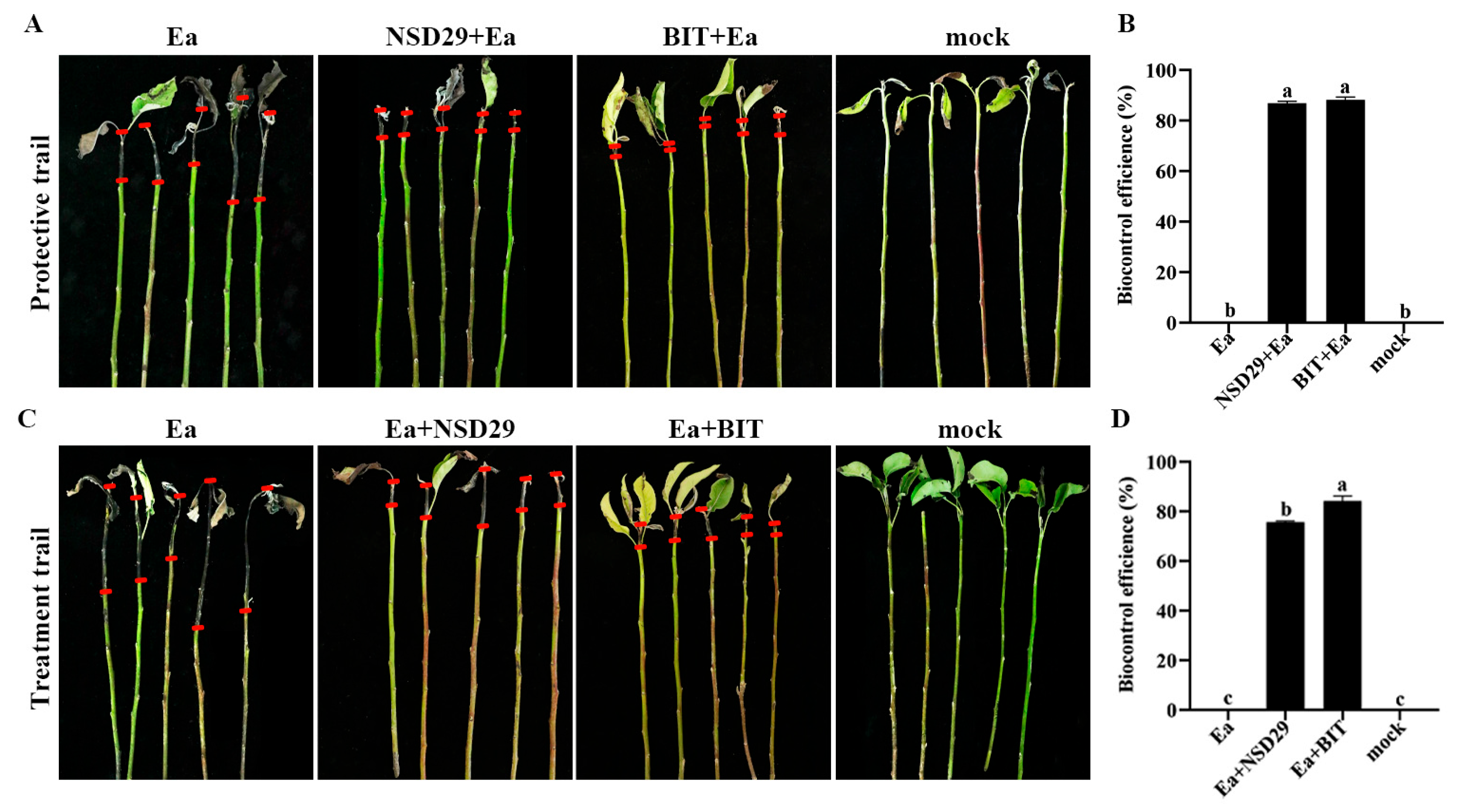
2.5. Evaluation of the Biocontrol Efficacy on Pear Fire Blight
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Isolation and Purification of Herpetosiphon
4.3. Morphological Observations
4.4. Biochemical Characterization
4.5. 16S rRNA Gene Analysis
4.6. Genome Sequencing and Analysis
4.7. Predatory Activity Against Plant Pathogens
4.8. Biological Assay of the Strain NSD29 Against Pear Fire Blight
4.8.1. Bacterial Preparation
4.8.2. Assay on Fragrant Pear Inflorescences
4.8.3. Assay on Detached Fragrant Pear Leaves
4.8.4. Assay on Detached Fragrant Pear Fruits
4.8.5. Assay on Detached Fragrant Pear Branches
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- García-Fernández, B.; Dolcet-Sanjuan, R.; Micheletti, D.; Antón-Díaz, M.J.; Solsona, C.; Fernández, M.; Abad, X.; Dapena, E. Susceptibility evaluation to fire blight and genome-wide associations within a collection of Asturian apple accessions. Plants 2023, 12, 4068. [Google Scholar] [CrossRef] [PubMed]
- Momol, M.T.; Momol, E.A.; Dankers, W. A severe outbreak of fire blight in woody ornamental Rosaceae plants in North Florida and South Georgia. Plant Dis. 2000, 84, 1153. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Sheng, Q.; Luo, M.; Ma, D.; Zhang, C. Occurrence status of fire blight on Korla fragrant pear in Xinjiang and the control proposals. Plant Prot. 2022, 48, 207–213. [Google Scholar] [CrossRef]
- Aktepe, B.P.; Aysan, Y. Biological control of fire blight disease caused by Erwinia amylovora on apple. Erwerbs-Obstbau 2023, 65, 645–654. [Google Scholar] [CrossRef]
- Slack, S.M.; Zeng, Q.; Outwater, C.A.; Sundin, G.W. Microbiological examination of Erwinia amylovora exopolysaccharide ooze. Phytopathology 2017, 107, 403–411. [Google Scholar] [CrossRef]
- Norelli, J.L.; Jones, A.L.; Aldwinckle, H.S. Fire blight management in the twenty-first century: Using new technologies that enhance host resistance in apple. Plant Dis. 2003, 87, 756–765. [Google Scholar] [CrossRef]
- Sun, W.; Gong, P.; Zhao, Y.; Ming, L.; Zeng, Q.; Liu, F. Current situation of fire blight in China. Phytopathology 2023, 113, 2143–2151. [Google Scholar] [CrossRef]
- Shtienberg, D.; Manulis-Sasson, S.; Zilberstaine, M.; Oppenheim, D.; Shwartz, H. The incessant battle against fire blight in pears: 30 years of challenges and successes in managing the disease in Israel. Plant Dis. 2015, 99, 1048–1058. [Google Scholar] [CrossRef]
- Liu, Y.; Baird, S.M.; Qiao, J.; Du, Y.; Lu, S.E. SecG is required for antibiotic activities of Pseudomonas sp. YL23 against Erwinia amylovora and Dickeya chrysanthemi. J. Basic Microbiol. 2015, 55, 617–624. [Google Scholar] [CrossRef]
- Gusberti, M.; Klemm, U.; Meier, M.S.; Maurhofer, M.; Hunger-Glaser, I. Fire blight control: The struggle goes on. A comparison of different fire blight control methods in Switzerland with respect to biosafety, efficacy and durability. Int. J. Environ. Res. Public Health 2015, 12, 11422–11447. [Google Scholar] [CrossRef]
- Pujol, M.; Badosa, E.; Manceau, C.; Montesinos, E. Assessment of the environmental fate of the biological control agent of fire blight, P. fluorescens EPS62e, on apple by culture and real-time PCR methods. Appl. Environ. Microbiol. 2006, 72, 2421–2427. [Google Scholar] [CrossRef]
- Kearns, L.P.; Mahanty, H.K. Antibiotic production by Erwinia herbicola Eh1087: Its role in inhibition of Erwinia amylovora and partial characterization of antibiotic biosynthesis genes. Appl. Environ. Microbiol. 1998, 64, 1837–1844. [Google Scholar] [CrossRef] [PubMed]
- Bibi, S.; Weis, K.; Kaur, A.; Bhandari, R.; Goss, E.; Jones, J.B.; Potnis, N. A brief e-valuation of a copper resistance mobile genetic Island in the bacterial leaf spot pathogen Xanthomonas euvesicatoria pv. perforans. Phytopathology 2023, 113, 1394–1398. [Google Scholar] [CrossRef] [PubMed]
- Acimovic, S.G.; Zeng, Q.; McGhee, G.C.; Sundin, G.W.; Wise, J.C. Control of fire blight (Erwinia amylovora) on apple trees with trunk-injected plant resistance inducers and antibiotics and assessment of induction of pathogenesis-related protein genes. Front. Plant Sci. 2015, 6, 16. [Google Scholar] [CrossRef]
- Russo, N.L.; Burr, T.J.; Breth, D.I.; Aldwinckle, H.S. Isolation of streptomycin-resistant isolates of Erwinia amylovora in New York. Plant Dis. 2008, 92, 714–718. [Google Scholar] [CrossRef] [PubMed]
- Schlathölter, I.; Jänsch, M.; Flachowsky, H.; Broggini, G.A.L.; Hanke, M.V.; Patocchi, A. Generation of advanced fire blight-resistant apple (Malus×domestica) selections of the fifth generation within 7 years of applying the early flowering approach. Planta 2018, 247, 1475–1488. [Google Scholar] [CrossRef]
- Born, Y.; Fieseler, L.; Thöny, V.; Leimer, N.; Duffy, B.; Loessner, M.J. Engineering of bacteriophages Y2: dpoL1-C and Y2: luxAB for efficient control and rapid detection of the fire blight pathogen, Erwinia amylovora. Appl. Environ. Microbiol. 2017, 83, e00341-17. [Google Scholar] [CrossRef]
- Jordi, C.; Montesinos, E. Lysozyme enhances the bactericidal effect of BP100 peptide against Erwinia amylovora, the causal agent of fire blight of rosaceous plants. BMC Microbiol. 2017, 17, 39. [Google Scholar] [CrossRef]
- Din, I.U.; Hu, L.; Jiang, Y.; Wei, J.; Afzal, M.; Sun, L. Bacterial lipopeptides are effective against pear fire blight. Microorganisms 2024, 12, 896. [Google Scholar] [CrossRef]
- Cui, Z.; Hu, L.; Zeng, L.; Meng, W.; Guo, D.; Sun, L. Isolation and characterization of Priestia megaterium KD7 for the biological control of pear fire blight. Front. Microbiol. 2023, 14, 1099664. [Google Scholar] [CrossRef]
- Kunz, S.; Zeng, Q.; Johnson, K.B. History, efficacy, orchard ecology, and mode of action of Aureobasidium pullulans, the microbial agent in blossom protect, for suppression of fire blight of pome fruit. J. Plant Pathol. 2024, 106, 871–881. [Google Scholar] [CrossRef]
- Zeng, Q.; Johnson, K.B.; Mukhtar, S.; Nason, S.; Huntley, R.; Millet, F.; Yang, C.H.; Hassani, M.A.; Zuverza-Mena, N.; Sundin, G.W. Aureobasidium pullulans from the fire blight biocontrol product, blossom protect, induces host resistance in apple flowers. Phytopathology 2023, 113, 1192–1201. [Google Scholar] [CrossRef]
- Nguyen, L.T.T.; Park, A.R.; Van Le, V.; Hwang, I.; Kim, J.C. Exploration of a multifunctional biocontrol agent Streptomyces sp. JCK-8055 for the management of apple fire blight. Appl. Microbiol. Biotechnol. 2024, 108, 49. [Google Scholar] [CrossRef] [PubMed]
- Xian, W.; Zhang, X.; Li, W. Research status and prospect on bacterial Phylum Chloroflexi. Acta Microbiol. Sin. 2020, 60, 1801–1820. [Google Scholar] [CrossRef]
- Livingstone, P.G.; Morphew, R.M.; Cookson, A.R.; Whitworth, D.E. Genome analysis, metabolic potential, and predatory capabilities of Herpetosiphon llansteffanense sp. nov. Appl. Environ. Microbiol. 2018, 84, e01040-18. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef]
- Holt, J.G.; Lewin, R.A. Herpetosiphon aurantiacus gen. et sp. n., a new filamentous gliding organism. J. Bacteriol. 1968, 95, 2407–2408. [Google Scholar] [CrossRef]
- Kiss, H.; Nett, M.; Domin, N.; Martin, K.; Maresca, J.A. Complete genome sequence of the filamentous gliding predatory bacterium Herpetosiphon aurantiacus type strain (114-95(T)). Stand. Genom. Sci. 2011, 31, 356–370. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Kage, H.; Martin, K.; Nett, M. Herpetosiphon gulosus sp. nov., a filamentous predatory bacterium isolated from sandy soil and Herpetosiphon giganteus sp. nov., nom. rev. Int. J. Syst. Evol. Microbiol. 2017, 67, 2476–2481. [Google Scholar] [CrossRef]
- Wei, B.; Cao, L.; Li, S.; Huang, D.; Zhou, J.; Guo, X. The role of endophytes in medical plants and the effect of endophytes on secondary metabolites. Chin. Agric. Sci. Bull. 2011, 27, 83–88. [Google Scholar] [CrossRef]
- Akomoneh, E.A.; Gestels, Z.; Abdellati, S.; Vereecken, K.; Bartholomeeusen, K. Genome mining uncovers NRPS and PKS clusters in rothia dentocariosa with inhibitory activity against Neisseria Species. Antibiotics 2023, 12, 1592. [Google Scholar] [CrossRef]
- Cortes, J.; Haydock, S.F.; Roberts, G.A.; Bevitt, D.J.; Leadlay, P.F. An unusually large multifunctional polypeptide in the erythromycin-producing polyketide synthase of Saccharopolyspora erythraea. Nature 1990, 348, 176–178. [Google Scholar] [CrossRef] [PubMed]
- Nett, M.; Erol, O.; Kehraus, S.; Kock, M.; Krick, A.; Eguereva, E. Siphonazole, an unusual metabolite from Herpetosiphon sp. Angew. Chem.-Int. Ed. 2006, 45, 3863–3867. [Google Scholar] [CrossRef] [PubMed]
- Schieferdecker, S.; Domin, N.; Hoffmeier, C.; Bryant, D.A.; Roth, M.; Nett, M. Structure and absolute configuration of auriculamide, a natural product from the predatory bacterium Herpetosiphon aurantiacus. Eur. J. Org. Chem. 2015, 2015, 3057–3062. [Google Scholar] [CrossRef]
- Lewin, R.A. New Herpetosiphon species (Flexibacterales). Can. J. Microbiol. 1970, 16, 517–520. [Google Scholar] [CrossRef]
- Quinn, G.R.; Skerman, V.B.D. Herpetosiphon—Nature’s scavenger? Curr. Microbiol. 1980, 4, 57–62. [Google Scholar] [CrossRef]
- Zeller, W.; Wolf, B.; Bonn, W.G. Studies on biological control of fire blight. VII Int. Workshop Fire Blight 1996, 411, 341–346. [Google Scholar] [CrossRef]
- Bahadou, S.A.; Ouijja, A.; Karfach, A.; Tahiri, A.; Lahlali, R. New potential bacterial antagonists for the biocontrol of fire blight disease (Erwinia amylovora) in Morocco. Microb. Pathog. 2018, 117, 7–15. [Google Scholar] [CrossRef]
- Sharifazizi, M.; Harighi, B.; Sadeghi, A. Evaluation of biological control of Erwinia amylovora, causal agent of fire blight disease of pear by antagonistic bacteria. Biol. Control 2017, 104, 28–34. [Google Scholar] [CrossRef]
- Han, J.; Dong, Z.; Ji, W.; Lv, W.; Luo, M.; Fu, B. From predator to protector: Myxococcus fulvus WCH05 emerges as a potent biocontrol agent for fire blight. Front. Microbiol. 2024, 15, 1378288. [Google Scholar] [CrossRef]
- Bai, X.; Han, J.; Sun, B.; Song, S.; Luo, M.; Sun, L. Screening and identification of biocontrol potential myxobacteria strains against fire blight and pear canker diseases and evaluation of indoor control efficacy. Chin. J. Biol. Control 2023, 39, 1384–1397. [Google Scholar] [CrossRef]
- Lü, T.; Xu, L.; Xi, H.; Han, J.; Luo, M. Characterization of the infectious colonization and expansion with GFP tagged strain of Erwinia amylovora in Kuerlexiangli pear (Pyrus sinkiangensis Yu) shoots. J. Fruit Sci. 2023, 40, 1692–1702. [Google Scholar] [CrossRef]
- Lee, N.; Reichenbach, H. The Genus Herpetosiphon. Prokaryotes 2006, 7, 854–877. [Google Scholar] [CrossRef]
- Zhang, X.; Yao, Q.; Cai, Z.; Xie, X.; Zhu, H. Isolation and identification of myxobacteria from saline-alkaline soils in Xinjiang, China. PLoS ONE 2013, 8, e70466. [Google Scholar] [CrossRef]
- Nair, R.R.; Vasse, M.; Wielgoss, S.; Sun, L.; Yu, Y.N.; Velicer, G.J. Bacterial predator-prey coevolution accelerates genome evolution and selects on virulence-associated prey defences. Nat. Commun. 2019, 10, 4301. [Google Scholar] [CrossRef] [PubMed]
- Yi, S.; Zhou, Y.; Zhang, X.; Yao, Q.; Li, H.; Zhu, H. Effects of different methods on the formation of fruiting bodies and isolation of myxobacteria. Acta Microbiol. Sin. 2021, 61, 923–934. [Google Scholar] [CrossRef]
- Kim, M.; Oh, H.S.; Park, S.C.; Chun, J. Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int. J. Syst. Evol. Microbiol. 2014, 64, 346–351. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Coil, D.; Jospin, G.; Darling, A.E. A5-miseq: An updated pipeline to assemble microbial genomes from Illumina MiSeq data. Bioinformatics 2015, 31, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Blake, J.D.; Cohen, F.E. Pairwise sequence alignment below the twilight zone. J. Mol. Biol. 2001, 307, 721–735. [Google Scholar] [CrossRef]
- Cantalapiedra, C.P.; HernándezPlaza, A.; Letunic, I.; Peer, B.; Jaime, H.C. eggNOG-mapper v2: Functional annotation, orthology assignments, and domain prediction at the metagenomic scale. Mol. Biol. Evol. 2021, 38, 5825–5829. [Google Scholar] [CrossRef]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef]
- Kalvari, I.; Argasinska, J.; Quinones-Olvera, N.; Nawrocki, E.P.; Rivas, E.; Eddy, S.R. Rfam 13.0: Shifting to a genome-centric resource for non-coding RNA families. Nucleic Acids Res. 2018, 46, D335–D342. [Google Scholar] [CrossRef]
- Richter, M.; Rossello-Mora, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Augustijn, H.E.; Reitz, Z.L.; Biermann, F.; Alanjary, M. AntiSMASH 7.0: New and improved predictions for detection, regulation, chemical structures and visualisation. Nucleic Acids Res. 2023, 51, W46–W50. [Google Scholar] [CrossRef]
- Stothard, P.; Grant, J.R.; Van, D.G. Visualizing and comparing circular genomes using the CGView family of tools. Brief. Bioinform. 2019, 20, 1576–1582. [Google Scholar] [CrossRef]
- Medhioub, I.; Cheffi, M.; Tounsi, S. Study of Bacillus velezensis OEE1 potentialities in the biocontrol against Erwinia amylovora, causal agent of fire blight disease of rosaceous plants. Biol. Control 2022, 167, 104842. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biochemical Substrates | H. llansteffanensis CA052BT | H. aurantiacus DSM785T | H. geysericola DSM7119T | H. gulosus NBRC 112829T | H. giganteus DSM589T | NSD29 |
|---|---|---|---|---|---|---|
| Esculin | + | − | + | + | + | + |
| Arginine | − | + | − | + | + | + |
| Gelatin | + | − | − | − | + | + |
| Glucose | + | + | − | +/− | +/− | − |
| Arabinose | − | + | + | + | + | − |
| Sucrose | − | + | + | + | + | + |
| Rhamnose | − | + | + | + | + | − |
| Mannitol | − | + | + | + | + | − |
| Characteristic | H. llansteffanensis CA052BT | H. aurantiacus DSM785T | H. geysericola DSM7119T | NSD29 |
|---|---|---|---|---|
| Size (Mbp) | 6.14 | 6.79 | 6.24 | 5.78 |
| No. of contigs | 170 | 1 (+2 plasmids) | 46 | 1 (+4 plasmids) |
| %GC content | 50.8 | 50.9 | 50.7 | 50.89 |
| t1PKS-NRPS | 2 | 3 | 2 | 2 |
| Thiopeptide | 1 | 1 | 1 | 1 |
| Terpene | 2 | 2 | 2 | 2 |
| Bacteriocin | 2 | 1 | 2 | − |
| NRPS | 2 | 4 | 2 | 2 |
| t3PKS | 1 | 1 | 1 | 1 |
| t1PKS | − | 1 | − | − |
| Lantipeptide-t1PKS-NRPS | − | 1 | − | − |
| Code | Value | % | Description |
|---|---|---|---|
| B | 2 | 0.05 | Chromatin structure and dynamics |
| C | 277 | 6.87 | Energy production and conversion |
| D | 41 | 1.02 | Cell cycle control, cell division, and chromosome partitioning |
| E | 255 | 6.32 | Amino acid transport and metabolism |
| F | 131 | 3.25 | Nucleotide transport and metabolism |
| G | 256 | 6.35 | Carbohydrate transport and metabolism |
| H | 192 | 4.76 | Coenzyme transport and metabolism |
| I | 128 | 3.17 | Lipid transport and metabolism |
| J | 186 | 4.61 | Translation, ribosomal structure, and biogenesis |
| K | 400 | 9.92 | Transcription |
| L | 199 | 4.93 | Replication, recombination, and repair |
| M | 275 | 6.82 | Cell wall/membrane/envelope biogenesis |
| N | 33 | 0.82 | Cell motility |
| O | 123 | 3.05 | Post-translational modification, protein turnover, and chaperones |
| P | 208 | 5.16 | Inorganic ion transport and metabolism |
| Q | 110 | 2.73 | Secondary metabolites biosynthesis, transport, and catabolism |
| S | 707 | 17.53 | Function unknown |
| T | 333 | 8.26 | Signal transduction mechanisms |
| U | 85 | 2.11 | Intracellular trafficking, secretion, and vesicular transport |
| V | 88 | 2.18 | Defense mechanisms |
| W | 2 | 0.05 | Extracellular structures |
| Z | 2 | 0.05 | Cytoskeleton |
| Strains (GenBank Accession Number) | 16S rRNA Identity (%) | ANI (%) | dDDH (%) |
|---|---|---|---|
| H. llansteffanensis CA052BT (PUBZ01000117) | 99.2 | 96.1 | 66.2 |
| H. giganteus DSM589T (NZ_JAFBCZ010000001) | 99.2 | 91.7 | 45.3 |
| H. gulosus NBRC112829T (BAABRU010000059) | 99.2 | 84.5 | 28.1 |
| H. geysericola DSM7119T (LGKP01000036) | 98.2 | 91.1 | 43.2 |
| H. aurantiacus DSM785T (CP000875) | 95.5 | 84.4 | 28.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lv, W.; Wang, R.; Ji, W.; Fu, B.; Luo, M.; Han, J. A Herpetosiphon llansteffanensis Strain from Forest Soil Exhibits Biocontrol Activity Against Pear Fire Blight. Plants 2025, 14, 1564. https://doi.org/10.3390/plants14111564
Lv W, Wang R, Ji W, Fu B, Luo M, Han J. A Herpetosiphon llansteffanensis Strain from Forest Soil Exhibits Biocontrol Activity Against Pear Fire Blight. Plants. 2025; 14(11):1564. https://doi.org/10.3390/plants14111564
Chicago/Turabian StyleLv, Wen, Ruiyue Wang, Wenbo Ji, Benzhong Fu, Ming Luo, and Jian Han. 2025. "A Herpetosiphon llansteffanensis Strain from Forest Soil Exhibits Biocontrol Activity Against Pear Fire Blight" Plants 14, no. 11: 1564. https://doi.org/10.3390/plants14111564
APA StyleLv, W., Wang, R., Ji, W., Fu, B., Luo, M., & Han, J. (2025). A Herpetosiphon llansteffanensis Strain from Forest Soil Exhibits Biocontrol Activity Against Pear Fire Blight. Plants, 14(11), 1564. https://doi.org/10.3390/plants14111564