
Integrative Identification of Chloroplast Metabolism-Related RETICULATA-RELATED Genes in Soybean

 , and
, and
Abstract
1. Introduction
2. Results
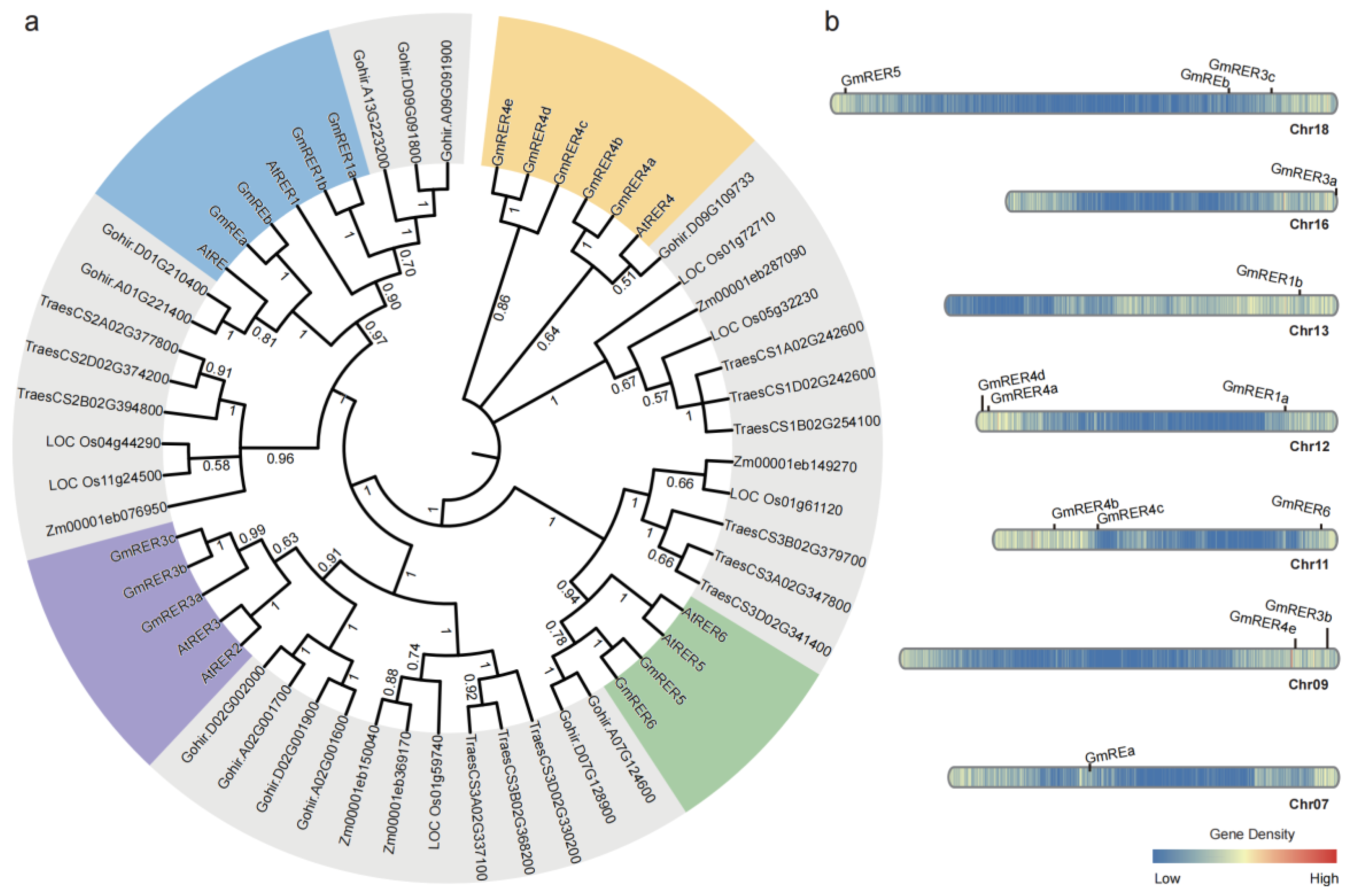
2.1. Identification of the GmRER Gene Family
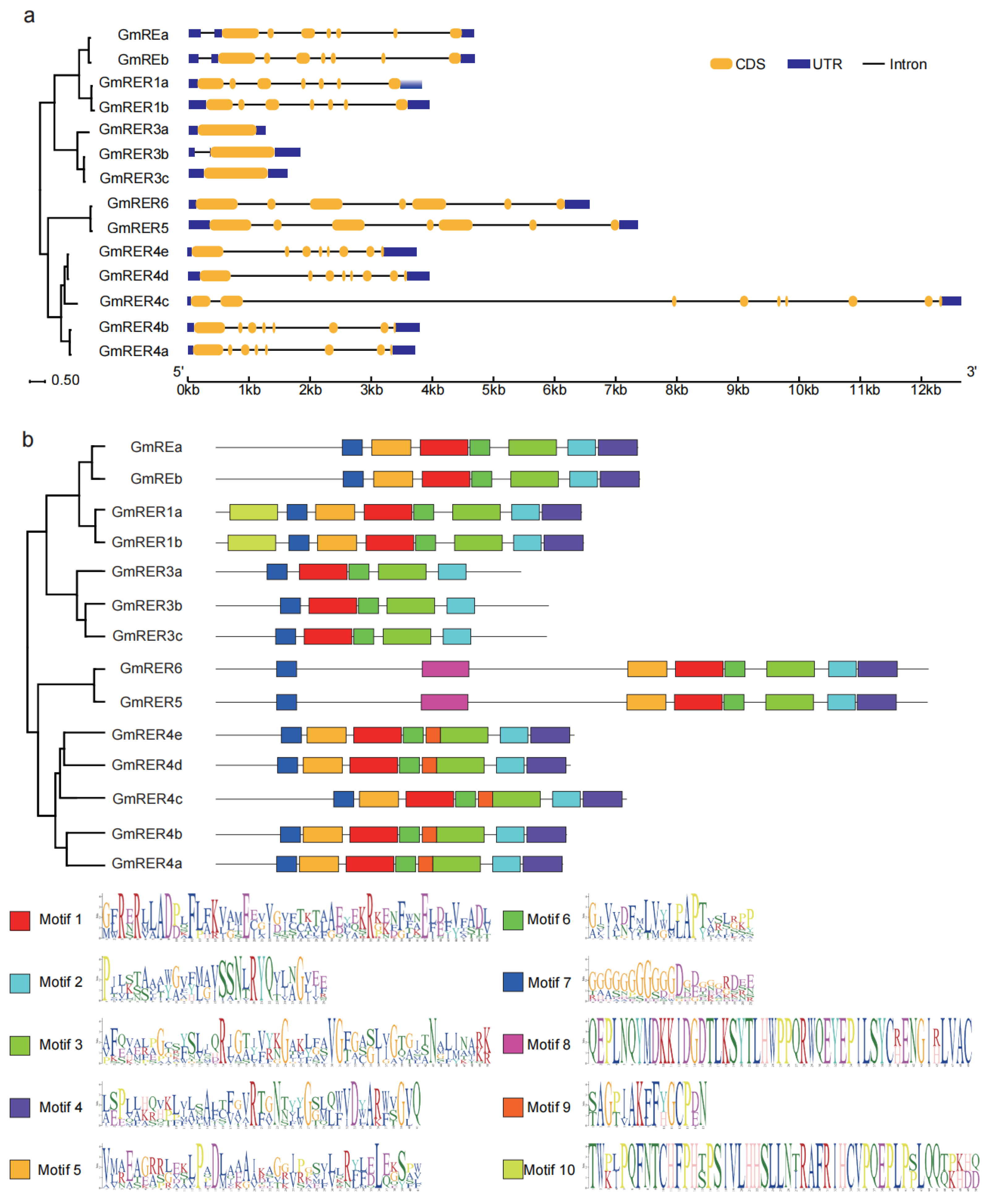
2.2. Architectural Diversity of GmRERs in Gene Structures and Conserved Motifs
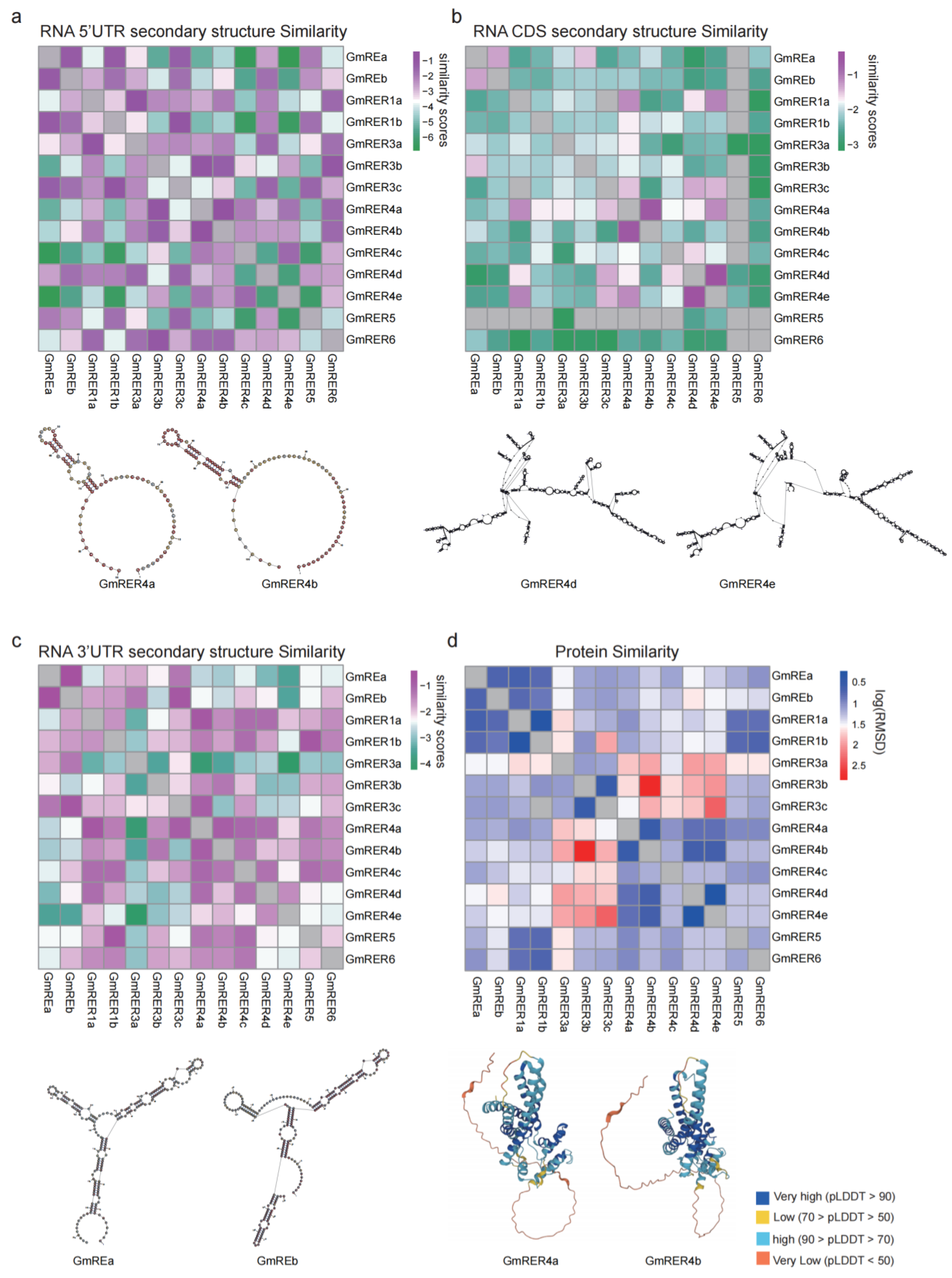
2.3. RNA Secondary Structure and Protein Structure Prediction of GmRERs
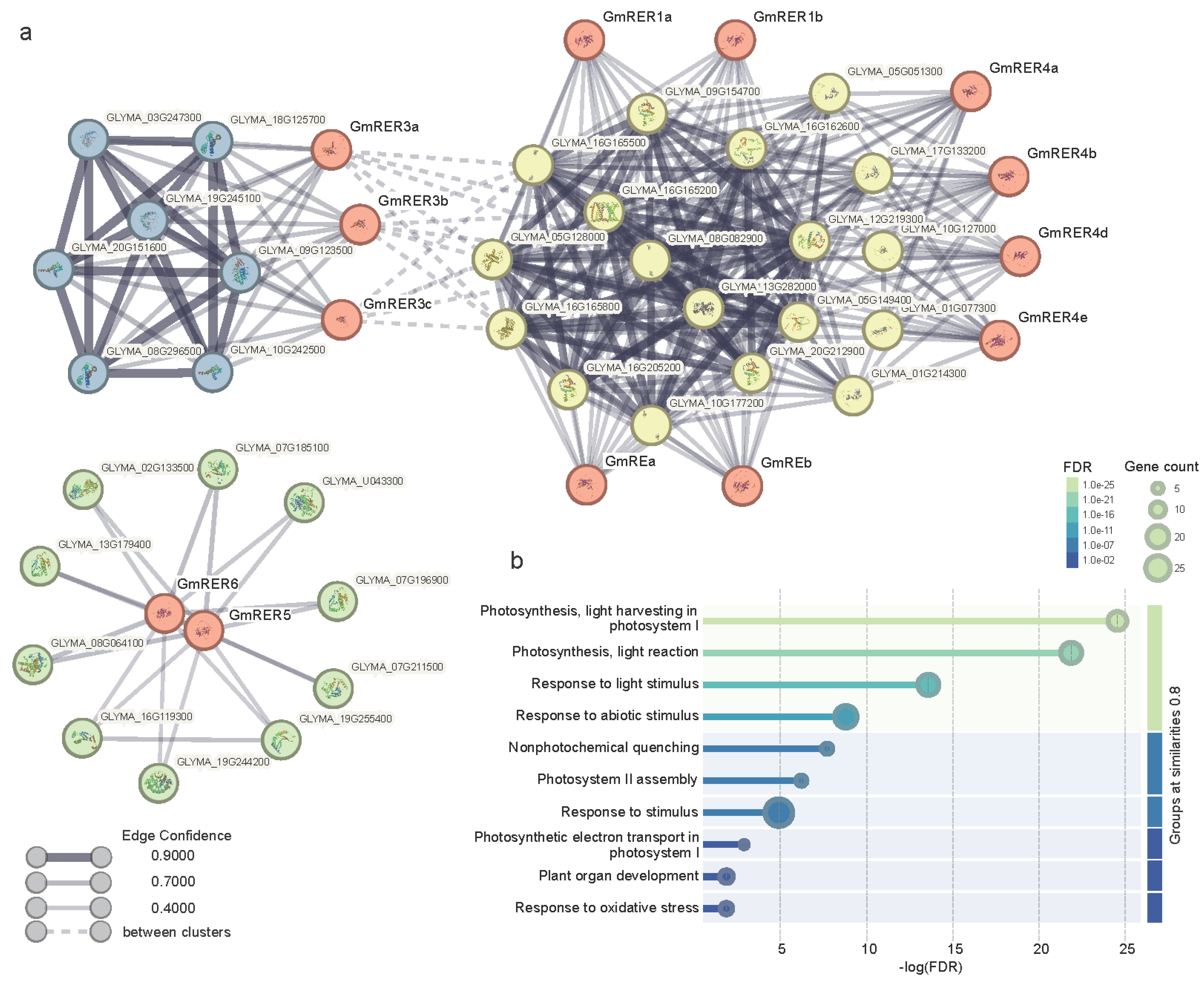
2.4. Modular Functionality of GmRER Paralogs via Subcellular Specialization and Lineage-Specific PPI Networks
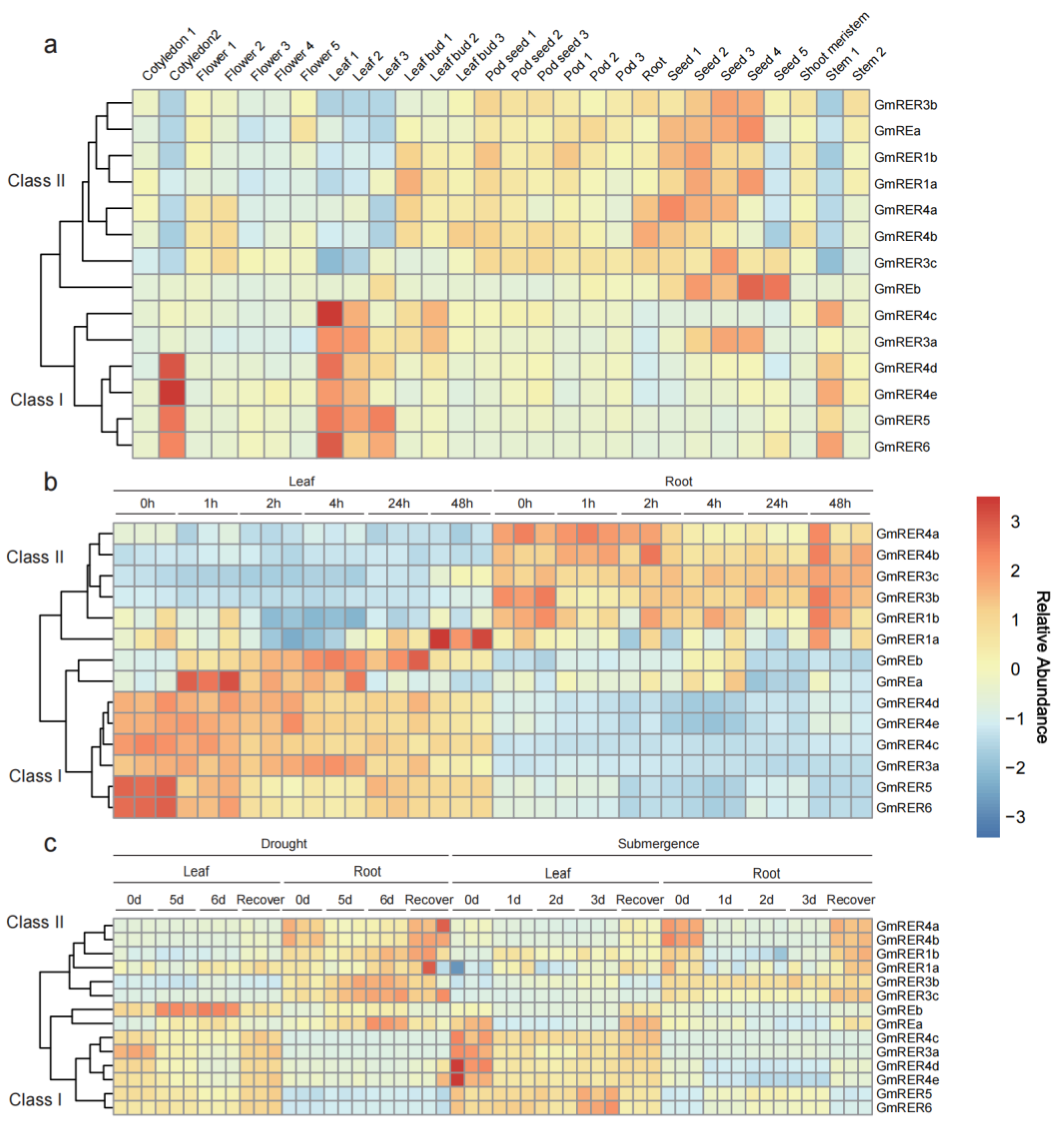
2.5. Spatiotemporal Expression Dynamics Define Functional Classes of GmRERs
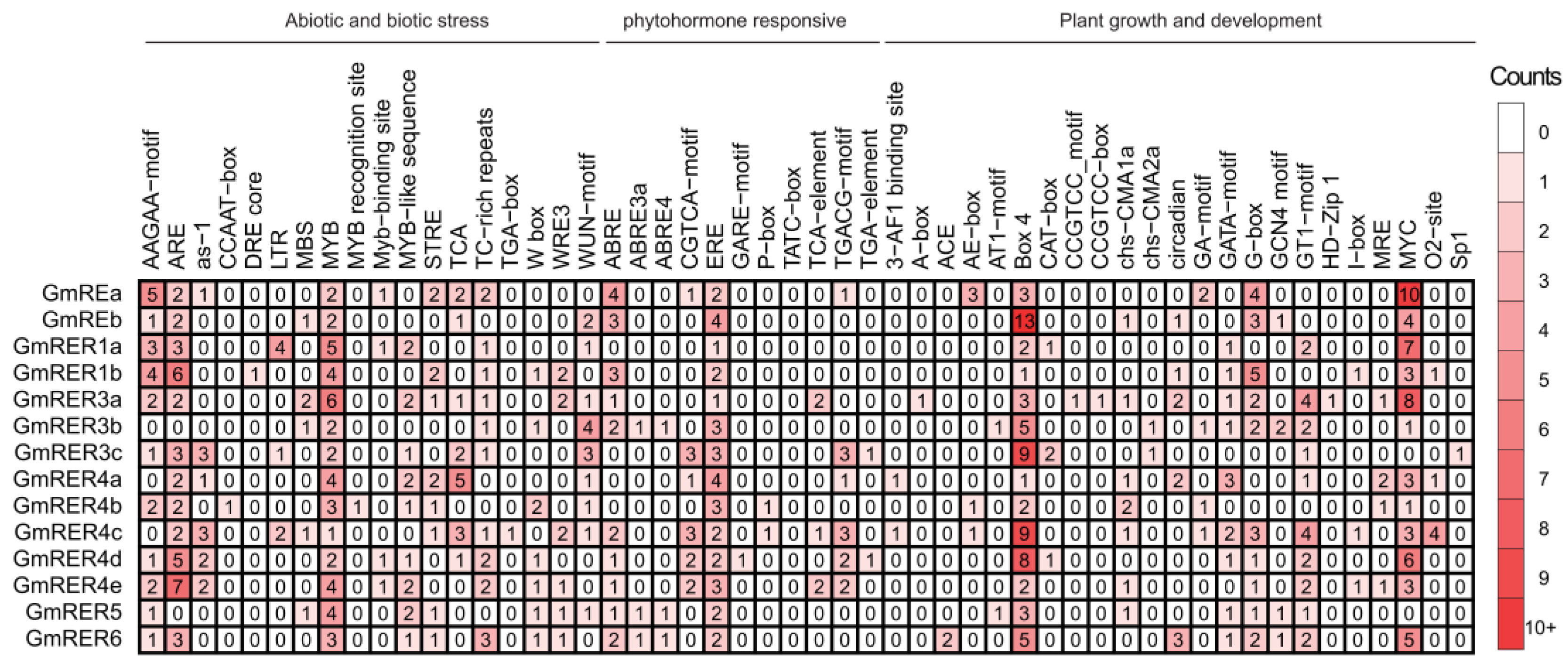
2.6. Cis-Regulatory Modules Conveyed Roles of GmRERs in Stress and Development
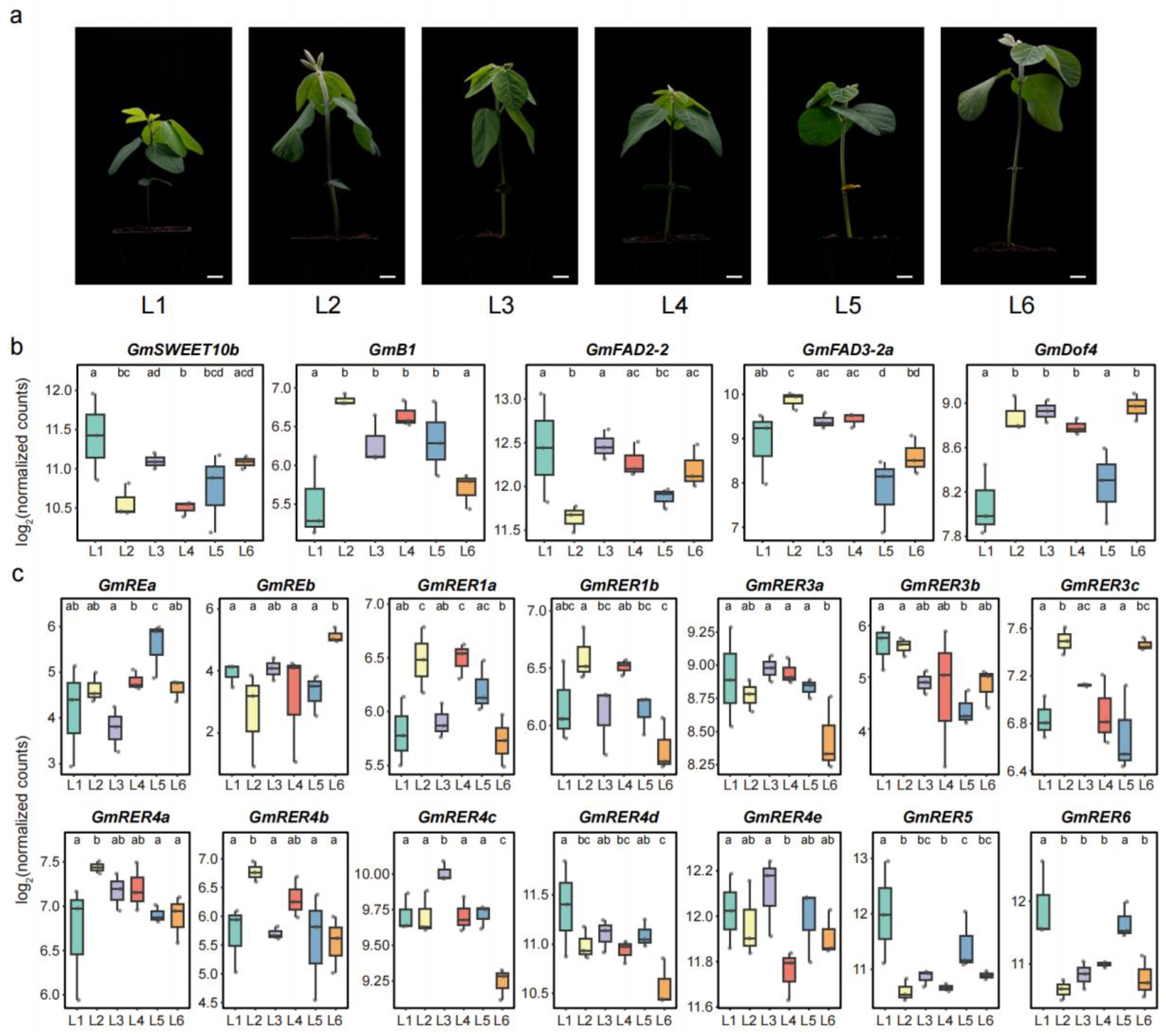
2.7. Expression Patterns of GmRERs in Six Northern Spring Soybean Landraces
3. Discussion
3.1. Evolutionary Conservation and Functional Dynamics in the GmRER Family
3.2. Structural Diversification Coupled with Regulatory Plasticity in GmRERs
3.3. Functional Implications of GmRERs in Leaf Lipid Metabolism
4. Materials and Methods
4.1. Growth Conditions
4.2. Genome-Wide Identification of GmRER Genes
4.3. Molecular Evolution Analysis and Chromosomal Locations
4.4. Gene Structure and Conserved Motif Annotation
4.5. Cis-Element Profiling and Promoter Analysis
4.6. Protein Interaction Network Construction
4.7. Expression Profiling and Data Visualization
4.8. RNA Extraction and Sequencing
4.9. RNA-seq Data Processing, Mapping, and Differential Expression Analysis
4.10. RNA Secondary Structure and Protein Similarity Assessment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABA | Abscisic acid |
| ABRE | Abscisic acid-responsive element |
| CDS | Coding sequence |
| CRISPR | Clustered Regularly Interspaced Short Palindromic Repeats |
| ERE | Ethylene-responsive element |
| FPKM | Fragments Per Kilobase of transcript per Million mapped reads |
| GO | Gene Ontology |
| MYB | MYeloBlastosis transcription factor family |
| PPI | Protein–protein interaction |
| RER | RETICULATA-RELATED |
| RMSD | Root-mean-square deviation |
| UTR | Untranslated region |
| WGD | Whole-genome duplication |
References
- Graham, P.H.; Vance, C.P. Legumes: Importance and Constraints to Greater Use. Plant Physiol. 2003, 131, 872–877. [Google Scholar] [CrossRef]
- Hartman, G.L.; West, E.D.; Herman, T.K. Crops That Feed the World 2. Soybean—Worldwide Production, Use, and Constraints Caused by Pathogens and Pests. Food Sec. 2011, 3, 5–17. [Google Scholar] [CrossRef]
- Pagano, M.C.; Miransari, M. The Importance of Soybean Production Worldwide. In Abiotic and Biotic Stresses in Soybean Production; Miransari, M., Ed.; Academic Press: San Diego, CA, USA, 2016; pp. 1–26. ISBN 978-0-12-801536-0. [Google Scholar]
- Tian, Z.; Nepomuceno, A.L.; Song, Q.; Stupar, R.M.; Liu, B.; Kong, F.; Ma, J.; Lee, S.-H.; Jackson, S.A. Soybean2035: A Decadal Vision for Soybean Functional Genomics and Breeding. Mol. Plant 2025, 18, 245–271. [Google Scholar] [CrossRef]
- al Amin, N.; Ahmad, N.; Wu, N.; Pu, X.; Ma, T.; Du, Y.; Bo, X.; Wang, N.; Sharif, R.; Wang, P. CRISPR-Cas9 Mediated Targeted Disruption of FAD2–2 Microsomal Omega-6 Desaturase in Soybean (Glycine max.L). BMC Biotechnol. 2019, 19, 9. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, T.; Watanabe, S.; Takagi, Y.; Anai, T. A Novel GmFAD3-2a Mutant Allele Developed through TILLING Reduces α-Linolenic Acid Content in Soybean Seed Oil. Breed. Sci. 2014, 64, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-W.; Zhang, B.; Hao, Y.-J.; Huang, J.; Tian, A.-G.; Liao, Y.; Zhang, J.-S.; Chen, S.-Y. The Soybean Dof-Type Transcription Factor Genes, GmDof4 and GmDof11, Enhance Lipid Content in the Seeds of Transgenic Arabidopsis Plants. Plant J. 2007, 52, 716–729. [Google Scholar] [CrossRef]
- Liu, Y.-F.; Li, Q.-T.; Lu, X.; Song, Q.-X.; Lam, S.-M.; Zhang, W.-K.; Ma, B.; Lin, Q.; Man, W.-Q.; Du, W.-G.; et al. Soybean GmMYB73 Promotes Lipid Accumulation in Transgenic Plants. BMC Plant Biol. 2014, 14, 73. [Google Scholar] [CrossRef]
- Zhang, Y.-Q.; Lu, X.; Zhao, F.-Y.; Li, Q.-T.; Niu, S.-L.; Wei, W.; Zhang, W.-K.; Ma, B.; Chen, S.-Y.; Zhang, J.-S. Soybean GmDREBL Increases Lipid Content in Seeds of Transgenic Arabidopsis. Sci. Rep. 2016, 6, 34307. [Google Scholar] [CrossRef]
- Manan, S.; Ahmad, M.Z.; Zhang, G.; Chen, B.; Haq, B.U.; Yang, J.; Zhao, J. Soybean LEC2 Regulates Subsets of Genes Involved in Controlling the Biosynthesis and Catabolism of Seed Storage Substances and Seed Development. Front. Plant Sci. 2017, 8, 1604. [Google Scholar] [CrossRef]
- Lu, X.; Li, Q.-T.; Xiong, Q.; Li, W.; Bi, Y.-D.; Lai, Y.-C.; Liu, X.-L.; Man, W.-Q.; Zhang, W.-K.; Ma, B.; et al. The Transcriptomic Signature of Developing Soybean Seeds Reveals the Genetic Basis of Seed Trait Adaptation during Domestication. Plant J. 2016, 86, 530–544. [Google Scholar] [CrossRef]
- Marchive, C.; Nikovics, K.; To, A.; Lepiniec, L.; Baud, S. Transcriptional Regulation of Fatty Acid Production in Higher Plants: Molecular Bases and Biotechnological Outcomes. Eur. J. Lipid Sci. Technol. 2014, 116, 1332–1343. [Google Scholar] [CrossRef]
- Xu, C.; Shanklin, J. Triacylglycerol Metabolism, Function, and Accumulation in Plant Vegetative Tissues*. Annu. Rev. Plant Biol. 2016, 67, 179–206. [Google Scholar] [CrossRef]
- Pyc, M.; Cai, Y.; Greer, M.S.; Yurchenko, O.; Chapman, K.D.; Dyer, J.M.; Mullen, R.T. Turning Over a New Leaf in Lipid Droplet Biology. Trends Plant Sci. 2017, 22, 596–609. [Google Scholar] [CrossRef]
- Wan, S.; Truong-Trieu, V.M.; Ward, T.; Whalen, J.K.; Altosaar, I. Advances in the Use of Genetically Modified Plant Biomass for Biodiesel Generation. Biofuels Bioprod. Biorefining 2017, 11, 749–764. [Google Scholar] [CrossRef]
- Rennie, B.D.; Tanner, J.W. Fatty Acid Composition of Oil from Soybean Leaves Grown at Extreme Temperatures. J. Am. Oil Chem. Soc. 1991, 68, 1O6–108. [Google Scholar] [CrossRef]
- Park, Y.J.; Lee, J.S.; Park, S.; Kim, Y.J.; Mani, V.; Lee, K.; Kwon, S.J.; Park, S.U.; Kim, J.K. Metabolite Changes in Soybean (Glycine Max) Leaves during the Entire Growth Period. ACS Omega 2023, 8, 41718–41727. [Google Scholar] [CrossRef]
- Shimada, T.L.; Hayashi, M.; Hara-Nishimura, I. Membrane Dynamics and Multiple Functions of Oil Bodies in Seeds and Leaves. Plant Physiol. 2018, 176, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Lagunas, B.; Román, Á.; Andreu, V.; Picorel, R.; Alfonso, M. A Temporal Regulatory Mechanism Controls the Different Contribution of Endoplasmic Reticulum and Plastidial ω-3 Desaturases to Trienoic Fatty Acid Content during Leaf Development in Soybean (Glycine Max Cv Volania). Phytochemistry 2013, 95, 158–167. [Google Scholar] [CrossRef]
- Pérez-Bueno, M.L.; Pineda, M.; Barón, M. Phenotyping Plant Responses to Biotic Stress by Chlorophyll Fluorescence Imaging. Front. Plant Sci. 2019, 10, 1135. [Google Scholar] [CrossRef]
- Leister, D. Retrograde Signaling in Plants: From Simple to Complex Scenarios. Front. Plant Sci. 2012, 3, 135. [Google Scholar] [CrossRef]
- Pérez-Pérez, J.M.; Esteve-Bruna, D.; González-Bayón, R.; Kangasjärvi, S.; Caldana, C.; Hannah, M.A.; Willmitzer, L.; Ponce, M.R.; Micol, J.L. Functional Redundancy and Divergence within the Arabidopsis RETICULATA-RELATED Gene Family. Plant Physiol. 2013, 162, 589–603. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, L.; Qu, L.-J. Plant Mediator Complex and Its Critical Functions in Transcription Regulation. J. Integr. Plant Biol. 2016, 58, 106–118. [Google Scholar] [CrossRef]
- Kesten, C.; García-Moreno, Á.; Amorim-Silva, V.; Menna, A.; Castillo, A.G.; Percio, F.; Armengot, L.; Ruiz-Lopez, N.; Jaillais, Y.; Sánchez-Rodríguez, C.; et al. Peripheral Membrane Proteins Modulate Stress Tolerance by Safeguarding Cellulose Synthases. Sci. Adv. 2022, 8, eabq6971. [Google Scholar] [CrossRef] [PubMed]
- Lundquist, P.K.; Rosar, C.; Bräutigam, A.; Weber, A.P.M. Plastid Signals and the Bundle Sheath: Mesophyll Development in Reticulate Mutants. Mol. Plant 2014, 7, 14–29. [Google Scholar] [CrossRef] [PubMed]
- Price, C.A.; Weitz, J.S. Costs and Benefits of Reticulate Leaf Venation. BMC Plant Biol. 2014, 14, 234. [Google Scholar] [CrossRef]
- Schubert, M.; Petersson, U.A.; Haas, B.J.; Funk, C.; Schröder, W.P.; Kieselbach, T. Proteome Map of the Chloroplast Lumen of Arabidopsis Thaliana *. J. Biol. Chem. 2002, 277, 8354–8365. [Google Scholar] [CrossRef]
- Peltier, J.-B.; Emanuelsson, O.; Kalume, D.E.; Ytterberg, J.; Friso, G.; Rudella, A.; Liberles, D.A.; Söderberg, L.; Roepstorff, P.; von Heijne, G.; et al. Central Functions of the Lumenal and Peripheral Thylakoid Proteome of Arabidopsis Determined by Experimentation and Genome-Wide Prediction. Plant Cell 2002, 14, 211–236. [Google Scholar] [CrossRef]
- Pérez-Pérez, J.M.; Candela, H.; Robles, P.; López-Torrejón, G.; del Pozo, J.C.; Micol, J.L. A Role for AUXIN RESISTANT3 in the Coordination of Leaf Growth. Plant Cell Physiol. 2010, 51, 1661–1673. [Google Scholar] [CrossRef]
- Lynch, M.; Force, A. The Probability of Duplicate Gene Preservation by Subfunctionalization. Genetics 2000, 154, 459–473. [Google Scholar] [CrossRef]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.-H. Evolution of Gene Duplication in Plants. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef]
- Freeling, M.; Scanlon, M.J.; Fowler, J.E. Fractionation and Subfunctionalization Following Genome Duplications: Mechanisms That Drive Gene Content and Their Consequences. Curr. Opin. Genet. Dev. 2015, 35, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tan, X.; Paterson, A.H. Different Patterns of Gene Structure Divergence Following Gene Duplication in Arabidopsis. BMC Genom. 2013, 14, 652. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome Sequence of the Palaeopolyploid Soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef]
- Adams, K.L.; Wendel, J.F. Polyploidy and Genome Evolution in Plants. Curr. Opin. Plant Biol. 2005, 8, 135–141. [Google Scholar] [CrossRef]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N. Phytozome: A Comparative Platform for Green Plant Genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef]
- Heinig, U.; Gutensohn, M.; Dudareva, N.; Aharoni, A. The Challenges of Cellular Compartmentalization in Plant Metabolic Engineering. Curr. Opin. Biotechnol. 2013, 24, 239–246. [Google Scholar] [CrossRef]
- Zoschke, R.; Bock, R. Chloroplast Translation: Structural and Functional Organization, Operational Control, and Regulation. Plant Cell 2018, 30, 745–770. [Google Scholar] [CrossRef] [PubMed]
- Woodson, J.D. Chloroplast Stress Signals: Regulation of Cellular Degradation and Chloroplast Turnover. Curr. Opin. Plant Biol. 2019, 52, 30–37. [Google Scholar] [CrossRef]
- Noctor, G.; Lelarge-Trouverie, C.; Mhamdi, A. The Metabolomics of Oxidative Stress. Phytochemistry 2015, 112, 33–53. [Google Scholar] [CrossRef]
- Chan, K.X.; Phua, S.Y.; Crisp, P.; McQuinn, R.; Pogson, B.J. Learning the Languages of the Chloroplast: Retrograde Signaling and Beyond. Annu. Rev. Plant Biol. 2016, 67, 25–53. [Google Scholar] [CrossRef]
- Tamang, B.G.; Li, S.; Rajasundaram, D.; Lamichhane, S.; Fukao, T. Overlapping and Stress-Specific Transcriptomic and Hormonal Responses to Flooding and Drought in Soybean. Plant J. 2021, 107, 100–117. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Zhou, Z.; Wang, Z.; Li, W.; Fang, C.; Wu, M.; Ma, Y.; Liu, T.; Kong, L.-A.; Peng, D.-L.; et al. Global Dissection of Alternative Splicing in Paleopolyploid Soybean. Plant Cell 2014, 26, 996–1008. [Google Scholar] [CrossRef] [PubMed]
- Belamkar, V.; Weeks, N.T.; Bharti, A.K.; Farmer, A.D.; Graham, M.A.; Cannon, S.B. Comprehensive Characterization and RNA-Seq Profiling of the HD-Zip Transcription Factor Family in Soybean (Glycine Max) during Dehydration and Salt Stress. BMC Genom. 2014, 15, 950. [Google Scholar] [CrossRef]
- Juven-Gershon, T.; Hsu, J.-Y.; Theisen, J.W.M.; Kadonaga, J.T. The RNA Polymerase II Core Promoter—The Gateway to Transcription. Curr. Opin. Cell Biol. 2008, 20, 253–259. [Google Scholar] [CrossRef]
- Narusaka, Y.; Nakashima, K.; Shinwari, Z.K.; Sakuma, Y.; Furihata, T.; Abe, H.; Narusaka, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Interaction between Two Cis-Acting Elements, ABRE and DRE, in ABA-Dependent Expression of Arabidopsis rd29A Gene in Response to Dehydration and High-Salinity Stresses. Plant J. 2003, 34, 137–148. [Google Scholar] [CrossRef]
- Zhang, M.; Qin, S.; Yan, J.; Li, L.; Xu, M.; Liu, Y.; Zhang, W. Genome-Wide Identification and Analysis of TCP Family Genes in Medicago Sativa Reveal Their Critical Roles in Na+/K+ Homeostasis. BMC Plant Biol. 2023, 23, 301. [Google Scholar] [CrossRef] [PubMed]
- Parwez, R.; Aftab, T.; Gill, S.S.; Naeem, M. Abscisic Acid Signaling and Crosstalk with Phytohormones in Regulation of Environmental Stress Responses. Environ. Exp. Bot. 2022, 199, 104885. [Google Scholar] [CrossRef]
- Wang, G.-Y.; Shi, J.-L.; Ng, G.; Battle, S.L.; Zhang, C.; Lu, H. Circadian Clock-Regulated Phosphate Transporter PHT4;1 Plays an Important Role in Arabidopsis Defense. Mol. Plant 2011, 4, 516–526. [Google Scholar] [CrossRef]
- Zhao, S.; Gao, H.; Jia, X.; Li, X.; Mao, K.; Ma, F. The γ-Clade HD-Zip I Transcription Factor MdHB-7 Regulates Salt Tolerance in Transgenic Apple (Malus Domestica). Plant Soil. 2021, 463, 509–522. [Google Scholar] [CrossRef]
- Hyten, D.L.; Song, Q.; Zhu, Y.; Choi, I.-Y.; Nelson, R.L.; Costa, J.M.; Specht, J.E.; Shoemaker, R.C.; Cregan, P.B. Impacts of Genetic Bottlenecks on Soybean Genome Diversity. Proc. Natl. Acad. Sci. USA 2006, 103, 16666–16671. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, M.; Feng, F.; Tian, Z. Toward a “Green Revolution” for Soybean. Mol. Plant 2020, 13, 688–697. [Google Scholar] [CrossRef]
- Zhou, Z.; Jiang, Y.; Wang, Z.; Gou, Z.; Lyu, J.; Li, W.; Yu, Y.; Shu, L.; Zhao, Y.; Ma, Y.; et al. Resequencing 302 Wild and Cultivated Accessions Identifies Genes Related to Domestication and Improvement in Soybean. Nat. Biotechnol. 2015, 33, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Liu, X.; Shen, D.; Miao, H.; Xie, B.; Li, X.; Zeng, P.; Wang, S.; Shang, Y.; Gu, X.; et al. A Genomic Variation Map Provides Insights into the Genetic Basis of Cucumber Domestication and Diversity. Nat. Genet. 2013, 45, 1510–1515. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Zhao, H.; Ren, L.; Song, W.; Zeng, B.; Guo, J.; Wang, B.; Liu, Z.; Chen, J.; Li, W.; et al. Genome-Wide Genetic Changes during Modern Breeding of Maize. Nat. Genet. 2012, 44, 812–815. [Google Scholar] [CrossRef] [PubMed]
- Hufford, M.B.; Xu, X.; van Heerwaarden, J.; Pyhäjärvi, T.; Chia, J.-M.; Cartwright, R.A.; Elshire, R.J.; Glaubitz, J.C.; Guill, K.E.; Kaeppler, S.M.; et al. Comparative Population Genomics of Maize Domestication and Improvement. Nat. Genet. 2012, 44, 808–811. [Google Scholar] [CrossRef]
- Wang, S.; Liu, S.; Wang, J.; Yokosho, K.; Zhou, B.; Yu, Y.-C.; Liu, Z.; Frommer, W.B.; Ma, J.F.; Chen, L.-Q.; et al. Simultaneous Changes in Seed Size, Oil Content and Protein Content Driven by Selection of SWEET Homologues during Soybean Domestication. Natl. Sci. Rev. 2020, 7, 1776–1786. [Google Scholar] [CrossRef]
- Zhang, D.; Sun, L.; Li, S.; Wang, W.; Ding, Y.; Swarm, S.A.; Li, L.; Wang, X.; Tang, X.; Zhang, Z.; et al. Elevation of Soybean Seed Oil Content through Selection for Seed Coat Shininess. Nat. Plants 2018, 4, 30–35. [Google Scholar] [CrossRef]
- Zhao, X.; Fu, X.; Yin, C.; Lu, F. Wheat Speciation and Adaptation: Perspectives from Reticulate Evolution. aBIOTECH 2021, 2, 386–402. [Google Scholar] [CrossRef]
- Birchler, J.A.; Veitia, R.A. Gene Balance Hypothesis: Connecting Issues of Dosage Sensitivity across Biological Disciplines. Proc. Natl. Acad. Sci. USA 2012, 109, 14746–14753. [Google Scholar] [CrossRef]
- Li, C.; Ng, C.K.-Y.; Fan, L.-M. MYB Transcription Factors, Active Players in Abiotic Stress Signaling. Environ. Exp. Bot. 2015, 114, 80–91. [Google Scholar] [CrossRef]
- He, H.; Zhang, Y.; Xu, S.; Zhang, X.; Yang, X.; Cheng, Y. Genetic Transformation of GmFBX322 Gene and Salt Tolerance Physiology in Soybean. PLoS ONE 2024, 19, e0307706. [Google Scholar] [CrossRef] [PubMed]
- Varshney, R.K.; Bohra, A.; Yu, J.; Graner, A.; Zhang, Q.; Sorrells, M.E. Designing Future Crops: Genomics-Assisted Breeding Comes of Age. Trends Plant Sci. 2021, 26, 631–649. [Google Scholar] [CrossRef] [PubMed]
- Vu, L.D.; Xu, X.; Zhu, T.; Pan, L.; van Zanten, M.; de Jong, D.; Wang, Y.; Vanremoortele, T.; Locke, A.M.; van de Cotte, B.; et al. The Membrane-Localized Protein Kinase MAP4K4/TOT3 Regulates Thermomorphogenesis. Nat. Commun. 2021, 12, 2842. [Google Scholar] [CrossRef]
- Watson, S.J.; Sowden, R.G.; Jarvis, P. Abiotic Stress-Induced Chloroplast Proteome Remodelling: A Mechanistic Overview. J. Exp. Bot. 2018, 69, 2773–2781. [Google Scholar] [CrossRef]
- Ye, H.; Roorkiwal, M.; Valliyodan, B.; Zhou, L.; Chen, P.; Varshney, R.K.; Nguyen, H.T. Genetic Diversity of Root System Architecture in Response to Drought Stress in Grain Legumes. J. Exp. Bot. 2018, 69, 3267–3277. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.; Li, J.; Yahya, M.; Sher, A.; Ma, C.; Wang, X.; Qiu, L. Research Progress and Perspective on Drought Stress in Legumes: A Review. Int. J. Mol. Sci. 2019, 20, 2541. [Google Scholar] [CrossRef]
- Gonzalez-Rizzo, S.; Laporte, P.; Crespi, M.; Frugier, F. Legume Root Architecture: A Peculiar Root System. In Annual Plant Reviews Online; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2018; pp. 239–287. ISBN 978-1-119-31299-4. [Google Scholar]
- Wang, Z.; Yung, W.-S.; Gao, Y.; Huang, C.; Zhao, X.; Chen, Y.; Li, M.-W.; Lam, H.-M. From Phenotyping to Genetic Mapping: Identifying Water-Stress Adaptations in Legume Root Traits. BMC Plant Biol. 2024, 24, 749. [Google Scholar] [CrossRef]
- Jiang, G.-L.; Mireku, P.; Song, Q. Utilization of Natural Hybridization and Intra-Cultivar Variations for Improving Soybean Yield, Seed Weight, and Agronomic Traits. Crop Sci. 2024, 64, 3174–3184. [Google Scholar] [CrossRef]
- Rasmusson, D.C.; Phillips, R.L. Plant Breeding Progress and Genetic Diversity from De Novo Variation and Elevated Epistasis. Crop Sci. 1997, 37, 303–310. [Google Scholar] [CrossRef]
- Fasoula, V.A.; Boerma, H.R. Intra-Cultivar Variation for Seed Weight and Other Agronomic Traits within Three Elite Soybean Cultivars. Crop Sci. 2007, 47, 367–373. [Google Scholar] [CrossRef]
- Haun, W.J.; Hyten, D.L.; Xu, W.W.; Gerhardt, D.J.; Albert, T.J.; Richmond, T.; Jeddeloh, J.A.; Jia, G.; Springer, N.M.; Vance, C.P.; et al. The Composition and Origins of Genomic Variation among Individuals of the Soybean Reference Cultivar Williams 821[W][OA]. Plant Physiol. 2011, 155, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.; Binns, D.; Chang, H.-Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-Scale Protein Function Classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A Toolkit for Detection and Evolutionary Analysis of Gene Synteny and Collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER Web Server: Interactive Sequence Similarity Searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An Upgraded Gene Feature Visualization Server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME Suite: Tools for Motif Discovery and Searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. 20 Years of the SMART Protein Domain Annotation Resource. Nucleic Acids Res. 2018, 46, D493–D496. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L.L. Predicting Transmembrane Protein Topology with a Hidden Markov Model: Application to Complete Genomes1. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Salvatore, M.; Emanuelsson, O.; Winther, O.; von Heijne, G.; Elofsson, A.; Nielsen, H. Detecting Sequence Signals in Targeting Peptides Using Deep Learning. Life Sci. Alliance 2019, 2, e201900429. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING Database in 2021: Customizable Protein–Protein Networks, and Functional Characterization of User-Uploaded Gene/Measurement Sets. Nucleic Acids Res. 2020, 49, D605–D612. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the Unification of Biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Kovaka, S.; Zimin, A.V.; Pertea, G.M.; Razaghi, R.; Salzberg, S.L.; Pertea, M. Transcriptome Assembly from Long-Read RNA-Seq Alignments with StringTie2. Genome Biol. 2019, 20, 278. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-Based Genome Alignment and Genotyping with HISAT2 and HISAT-Genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, R.; Bernhart, S.H.; Höner zu Siederdissen, C.; Tafer, H.; Flamm, C.; Stadler, P.F.; Hofacker, I.L. ViennaRNA Package 2.0. Algorithms Mol. Biol. 2011, 6, 26. [Google Scholar] [CrossRef]
- Schirmer, S.; Giegerich, R. Forest Alignment with Affine Gaps and Anchors, Applied in RNA Structure Comparison. Theor. Comput. Sci. 2013, 483, 51–67. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making Protein Folding Accessible to All. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef]
- Schrödinger, L.; DeLano, W. PyMOL, (version 3.1.4.1); DeLano Scientific LLC: Palo Alto, CA, USA, 2020.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Arabidopsis Gene ID | Name | Type of Duplicattion | Location (Mb) | Protein Length (aa) | Protein pI | ProteinMW (Kda) | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|
| RE | AT2G37860 | GmREa | WGD | 16.276–16.281 | 444 | 4.813 | 47.64 | Chloroplast |
| GmREb | WGD | 45.830–45.836 | 446 | 5.688 | 47.94 | Chloroplast | ||
| RER1 | AT5G22790 | GmRER1a | WGD | 35.553–35.558 | 385 | 7.5 | 41.37 | Chloroplast |
| GmRER1b | WGD | 40.846–40.851 | 387 | 8.021 | 41.53 | Chloroplast | ||
| RER3 | AT3G08640 | GmRER3a | WGD | 38.020–38.022 | 321 | 10.196 | 34.04 | Chloroplast |
| GmRER3b | WGD | 49.235–49.238 | 350 | 8.476 | 36.94 | Chloroplast | ||
| GmRER3c | WGD | 50.736–50.739 | 348 | 9.149 | 36.92 | Chloroplast | ||
| RER4 | AT5G12470 | GmRER4a | WGD | 1.403–1.408 | 365 | 8.461 | 38.8 | Chloroplast |
| GmRER4b | WGD | 7.082–7.087 | 369 | 9.474 | 39.38 | Chloroplast | ||
| GmRER4c | Dispersed | 12.077–12.090 | 432 | 9.722 | 46.96 | Chloroplast | ||
| GmRER4d | WGD | 0.697–0.702 | 373 | 8.944 | 39.82 | Chloroplast | ||
| GmRER4e | WGD | 45.552–45.556 | 377 | 9.156 | 40.16 | Chloroplast | ||
| RER5 | AT2G40400 | GmRER5 | WGD | 1.682–1.690 | 748 | 8.777 | 82.64 | Plasma membrane |
| RER6 | AT3G56140 | GmRER6 | WGD | 37.799–37.807 | 749 | 8.44 | 82.63 | Plasma membrane |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Q.; Niu, L.; Gong, X.; Xing, Q.; Liang, J.; Lang, J.; Wang, T.; Yang, X. Integrative Identification of Chloroplast Metabolism-Related RETICULATA-RELATED Genes in Soybean. Plants 2025, 14, 1516. https://doi.org/10.3390/plants14101516
Dong Q, Niu L, Gong X, Xing Q, Liang J, Lang J, Wang T, Yang X. Integrative Identification of Chloroplast Metabolism-Related RETICULATA-RELATED Genes in Soybean. Plants. 2025; 14(10):1516. https://doi.org/10.3390/plants14101516
Chicago/Turabian StyleDong, Qianli, Lu Niu, Xiyu Gong, Qianlong Xing, Jie Liang, Jun Lang, Tianya Wang, and Xiangdong Yang. 2025. "Integrative Identification of Chloroplast Metabolism-Related RETICULATA-RELATED Genes in Soybean" Plants 14, no. 10: 1516. https://doi.org/10.3390/plants14101516
APA StyleDong, Q., Niu, L., Gong, X., Xing, Q., Liang, J., Lang, J., Wang, T., & Yang, X. (2025). Integrative Identification of Chloroplast Metabolism-Related RETICULATA-RELATED Genes in Soybean. Plants, 14(10), 1516. https://doi.org/10.3390/plants14101516