

Phenotypic Characters and Inheritance Tendency of Agronomic Traits in F1 Progeny of Pear

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Data Collection and Analysis Methods
2.3. Statistical Analysis
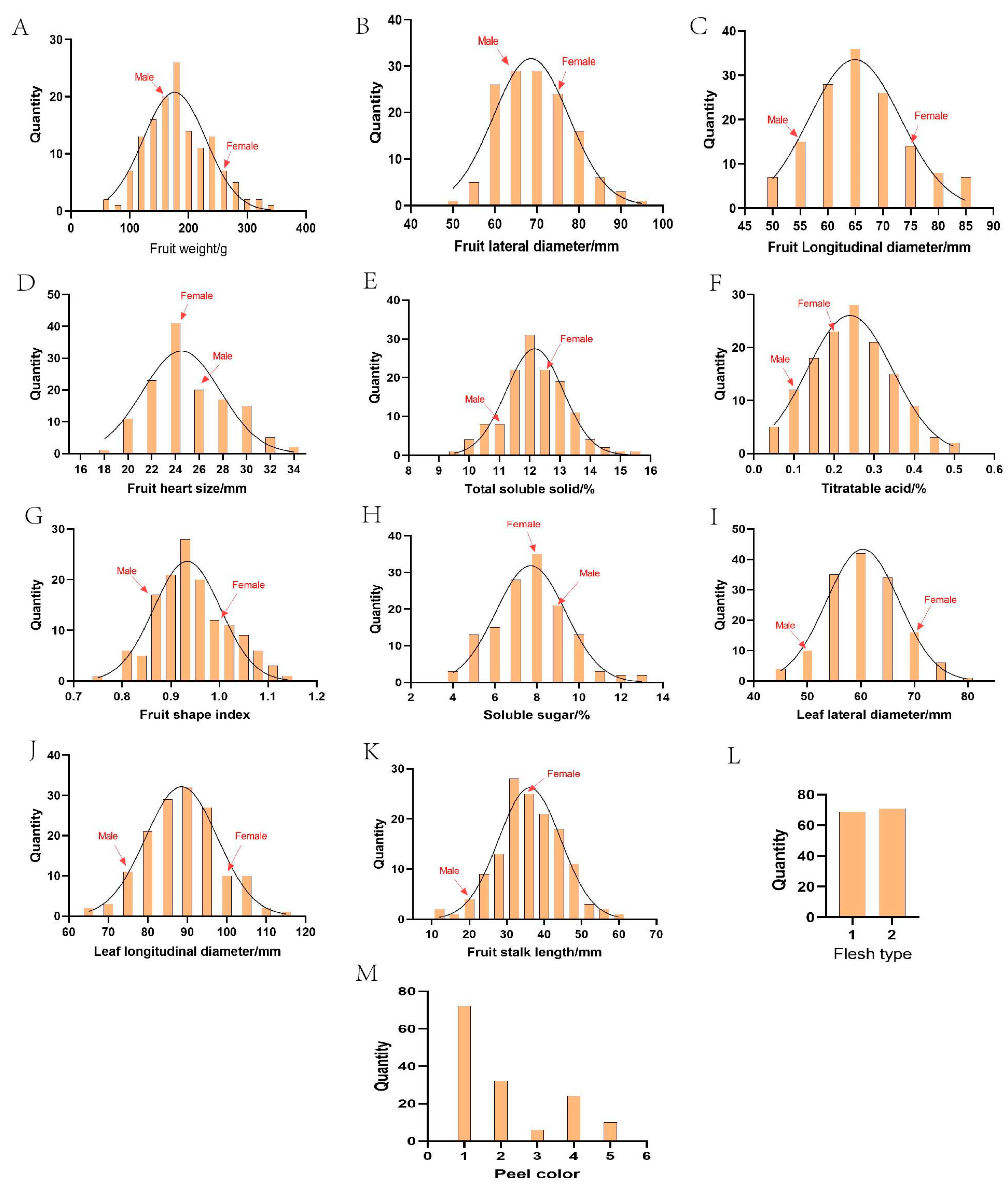
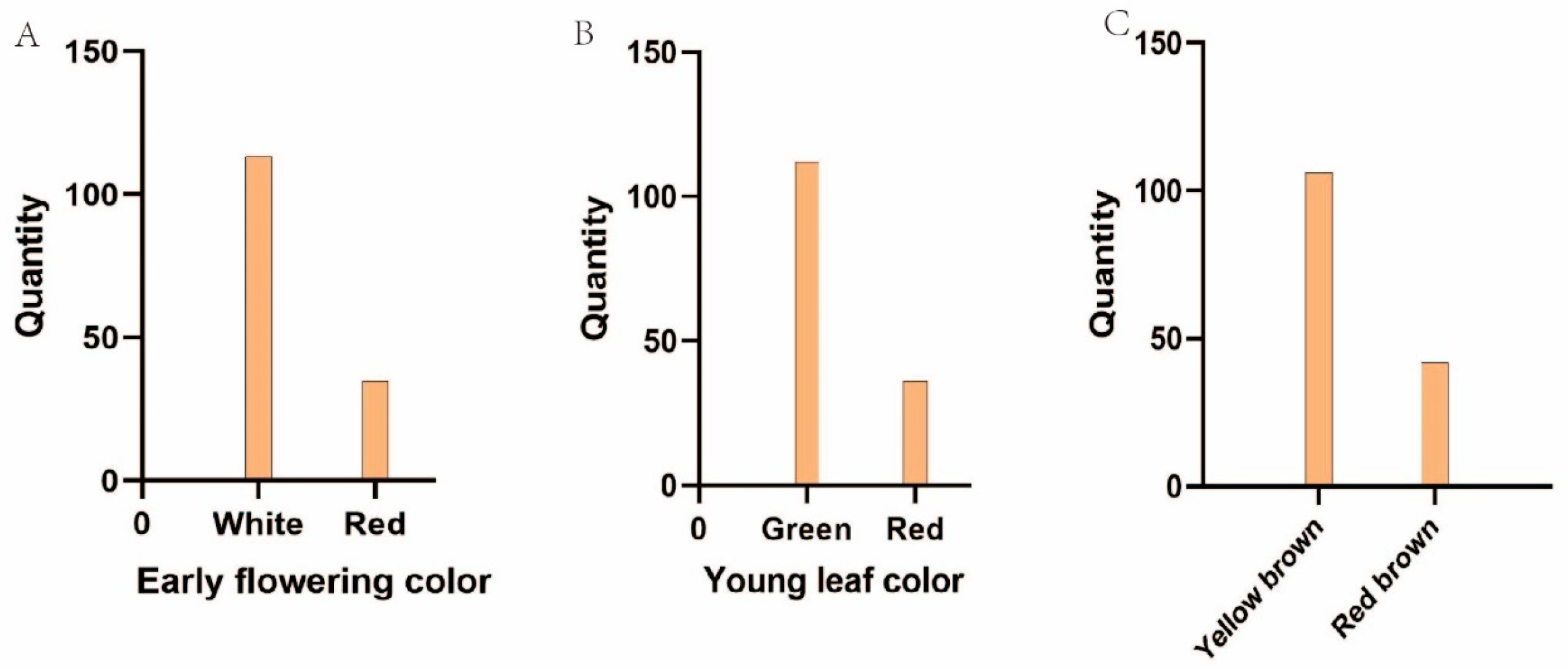
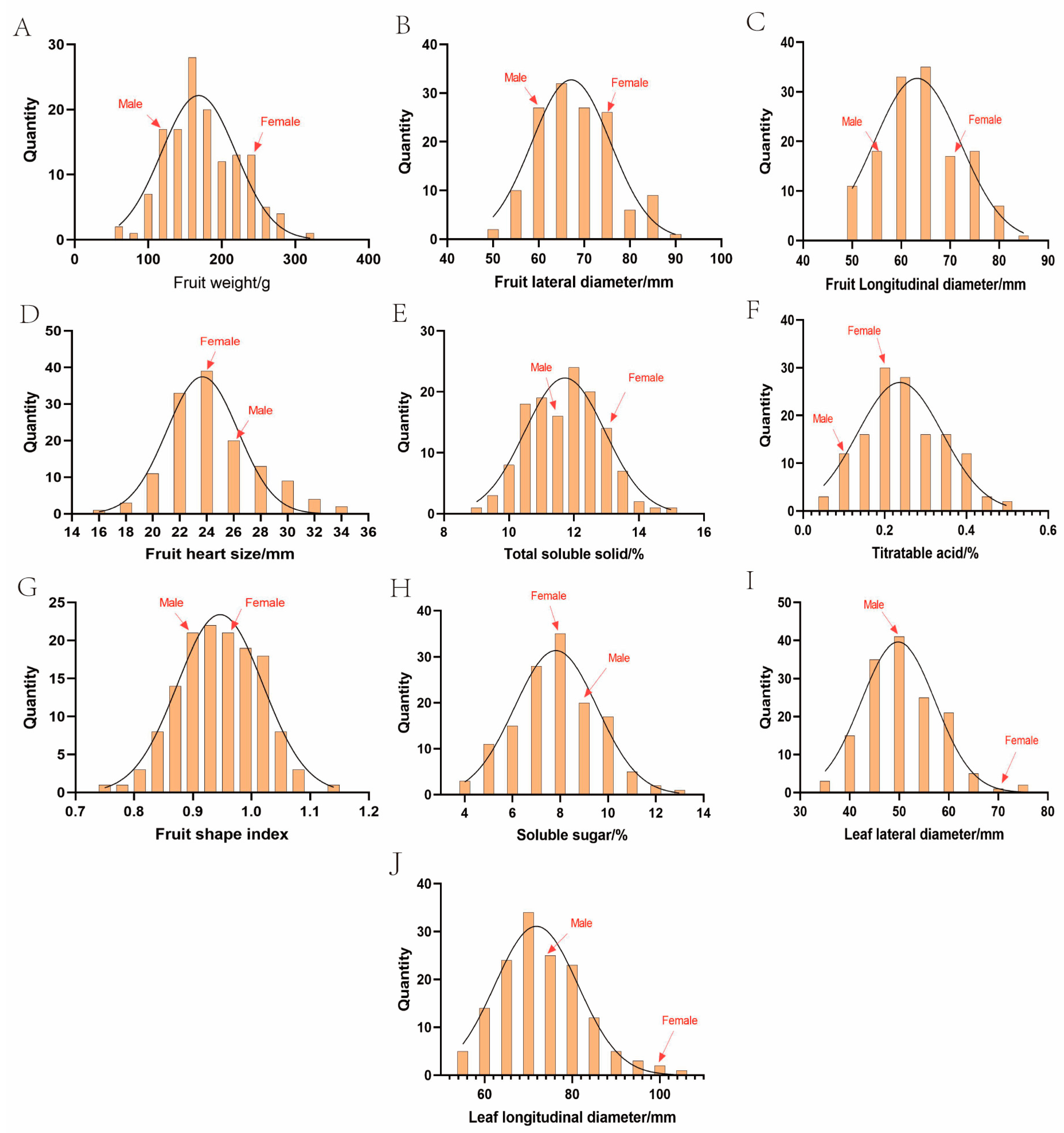
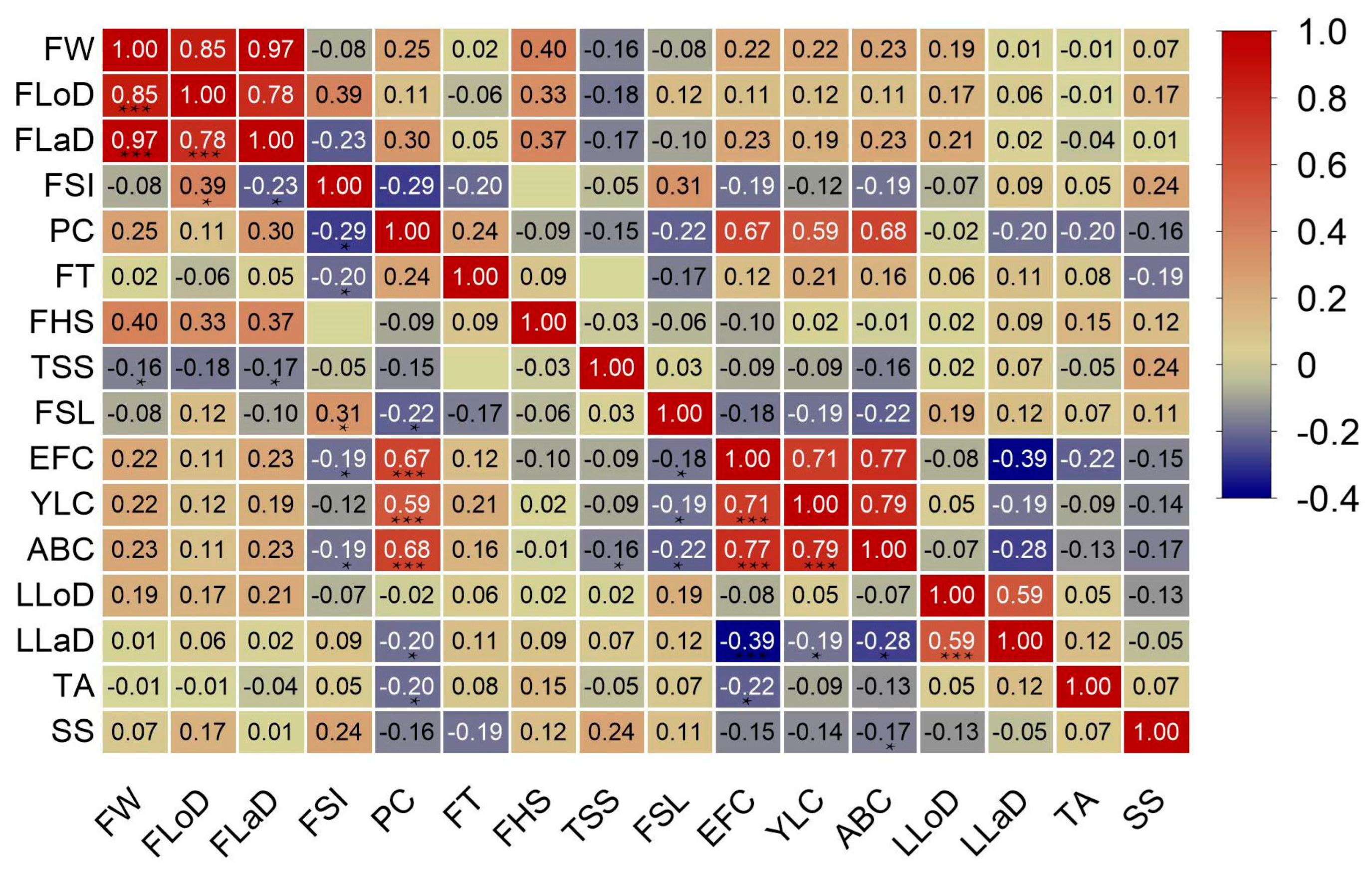
3. Results
3.1. Trait Differences Between Parents
3.2. Inheritance Analysis of Traits
3.3. Heterosis Analysis
3.4. Trait Correlation Analysis
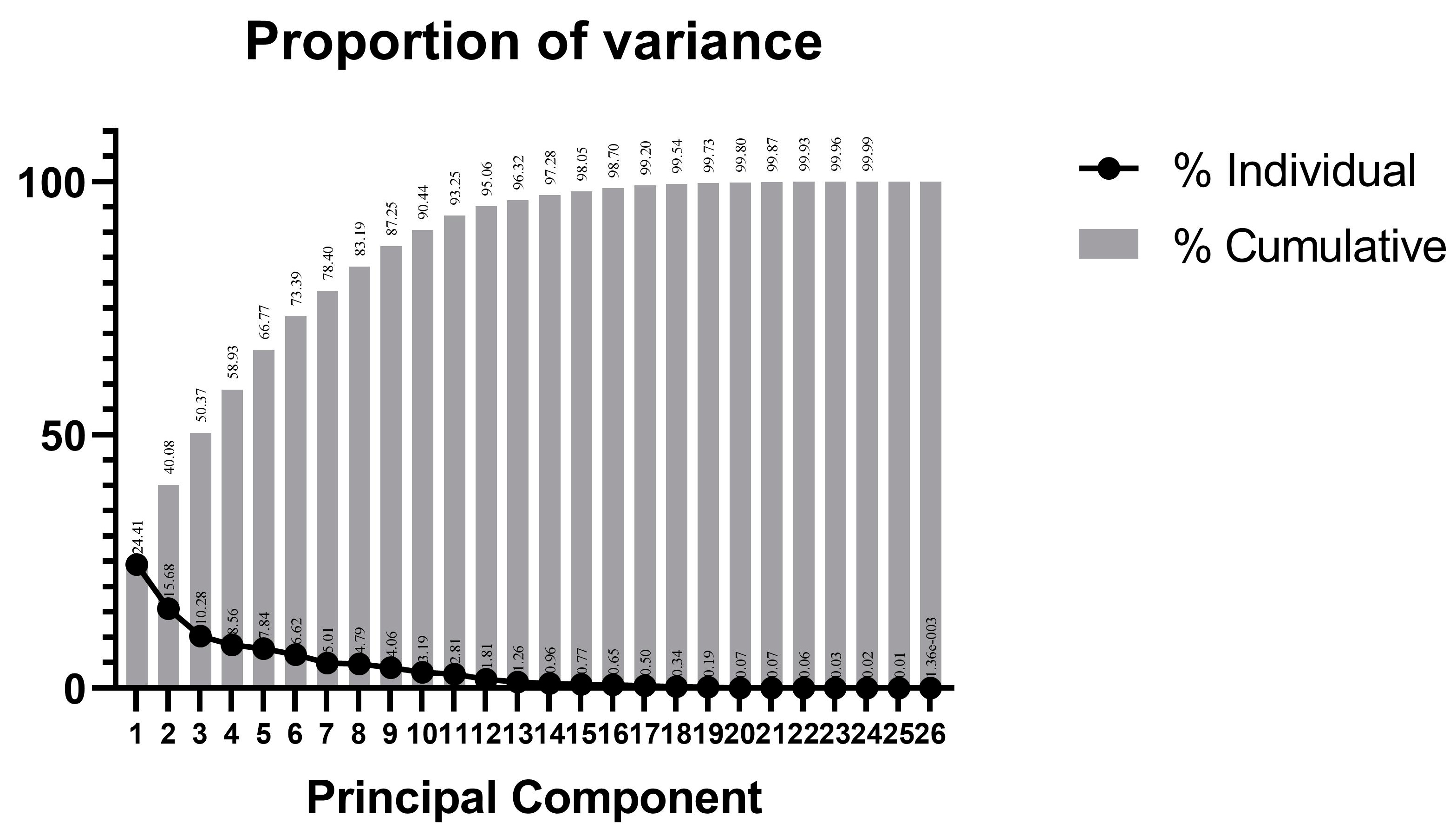
3.5. Principal Component Analysis of Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Huang, C.; Yu, B.; Teng, Y.; Su, J.; Shu, Q.; Cheng, Z. Effects of fruit bagging on coloring and related physiology, and qualities of red Chinese sand pears during fruit maturation. Sci. Hortic. 2009, 121, 149–158. [Google Scholar] [CrossRef]
- Wu, J.; Wang, Y.; Xu, J.; Korban, S.S.; Fei, Z.; Tao, S.; Ming, R.; Tai, S.; Khan, A.M.; Postman, J.D.; et al. Diversification and independent domestication of Asian and European pears. Genome Biol. 2018, 19, 77. [Google Scholar] [CrossRef] [PubMed]
- Qin, M.F.; Li, L.T.; Singh, J.; Sun, M.Y.; Bai, B.; Li, S.W.; Ni, J.P.; Zhang, J.Y.; Zhang, X.; Wei, W.L.; et al. Construction of a high-density bin-map and identification of fruit quality-related quantitative trait loci and functional genes in pear. Hortic. Res. 2022, 9, uhac141. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Kirk, C.; Deng, C.H.; Shirtliff, A.; Wiedow, C.; Qin, M.; Wu, J.; Brewer, L. Marker-trait associations and genomic predictions of interspecific pear (Pyrus) fruit characteristics. Sci. Rep. 2019, 9, 9072. [Google Scholar] [CrossRef]
- Wang, F.; Ou, C.Q.; Zhang, Y.J.; Ma, L.; Jiang, S.F. A New Red and Crisp Pear Cultivar ‘Huatong’. Acta Hortic. Sin. 2024, 51, 31–32. [Google Scholar]
- Zhang, Y.J.; Ou, C.Q.; Wang, F.; Ma, L.; Qi, L.Y.; Jiang, S.F. A New Mid-Maturing Red Cultivar ‘Xingli Yanhong’. Acta Hortic. Sin. 2024, 51, 29–30. [Google Scholar] [CrossRef]
- Tu, J.F.; Li, X.M.; Yang, F.C. A High Quality Early-Maturing Sand Pear New Cultivar ‘Mingjing’. Acta Hortic. Sin. 2024, 51, 15–16. [Google Scholar]
- Gu, C.; Xu, Y.; Wu, L.; Wang, X.; Qi, K.; Qiao, X.; Wang, Z.; Li, Q.; He, M.; Zhang, S. Long-read genome sequencing reveals the sequence characteristics of pear self-incompatibility locus. Mol. Hortic. 2025, 5, 13. [Google Scholar] [CrossRef]
- Gai, J.; Zhang, Y.M.; Wang, J. Genetic System of Quantitative Traits in Plants; Science Press: Beijing, China, 2003. (In Chinese) [Google Scholar]
- Dondini, L.; Pierantoni, L.; Ancarani, V.; D’Angelo, M.; Cho, K.H.; Shin, I.S.; Musacchi, S.; Kang, S.J.; Sansavini, S. The inheritance of the red colour character in European pear (Pyrus communis) and its map position in the mutated cultivar ‘Max Red Bartlett’. Plant Breed. 2008, 127, 524–526. [Google Scholar] [CrossRef]
- Oh, S.; Han, H.; Kim, D. A Novel Pear Scab (Venturia nashicola) Resistance Gene, Rvn3, from Interspecific Hybrid Pear (Pyrus pyrifolia × P. Communis). Plants 2021, 10, 2632. [Google Scholar] [CrossRef]
- Hwang, H.S.; Byeon, J.K.; Kim, W.C. Inheritance of Fruit Texture Traits in Oriental Pear (Pyrus pyrifolia var. culta Nakai). Wonye Kwahak Kisulchi = Korean J. Hortic. Sci. Technol. 2014, 32, 77–83. [Google Scholar]
- Liu, L.; Chen, C.X.; Zhu, Y.F. Maternal inheritance has impact on organic acid content in progeny of pear (Pyrus spp.) fruit. Euphytica 2016, 209, 305–321. [Google Scholar] [CrossRef]
- Jiao, C.Y.; Tao, K.Q.; Yu, Z.Y.; Shen, T.H. Study on genetic predisposition of Chinese plum fruit quality. Northen Hortic. 1999, 2, 23–25. [Google Scholar]
- Brown, A.G.; Harvey, D.M. The nature and inheritance of sweetness and acidity in the cultivated apple. Euphytica 1971, 20, 68–80. [Google Scholar] [CrossRef]
- Zou, L.M. Selection and Breeding of a New High-Quality, Storage-Resistant, Medium-Ripe Variety of ‘Yuluxiang’; Institute of Fruit Tree Research, Shanxi Academy of Agricultural Sciences: Taiyuan, China, 2003. [Google Scholar]
- Yue, W.; Zhang, H.; Liu, J. Breeding Report of a New Red Pear Cultivar ‘Xianghongli’. Agric. Sci. Technol. 2016, 17, 2569. [Google Scholar]
- Wang, L.; Wang, S.K.; Su, Y.L.; Chen, Y.; Wang, L.; Li, X.G.; Yang, J. Identification of Quantitative Trait Loci Associated with Fruit Quality Traits in Pear. Int. J. Agric. Biol. 2020, 23, 319–325. [Google Scholar]
- Kim, Y.; Oh, S.; Han, H.; Kim, D. QTL Analysis and CAPS Marker Development Linked with Russet in Pear (Pyrus spp.). Plants 2022, 11, 3196. [Google Scholar] [CrossRef]
- Cao, Y. Descriptors and Date Standard for Pear (Pyrus spp.); China Agriculture Press: Beijing, China, 2006. (In Chinese) [Google Scholar]
- Gao, J.F. Plant Physiology Experiment Guidance; Higher Education Press: Beijing, China, 2006; pp. 144–147. [Google Scholar]
- Li, H.S. Principles and Techniques of Plant Physiological and Biochemical Experiments; Higher Education Press: Beijing, China, 2000; pp. 195–197. [Google Scholar]
- Kaczmarska, E.; Gawroński, J.; Jabłońska-Ryś, E.; Zalewska-Korona, M.; Radzki, W.; Sławińska, A. Hybrid performance and heterosis in strawberry (Fragaria × ananassa Duchesne), regarding acidity, soluble solids and dry matter content in fruits. Plant Breed. 2016, 135, 232–238. [Google Scholar] [CrossRef]
- Li, J.; Zhang, M.; Li, X.; Khan, A.; Kumar, S.; Allan, A.C.; Lin-Wang, K.; Espley, R.V.; Wang, C.; Wang, R.; et al. Pear genetics: Recent advances, new prospects, and a roadmap for the future. Hortic. Res. 2022, 9, uhab040. [Google Scholar] [CrossRef]
- Teng, Y.; Tanabe, K.; Tamura, F.; Itai, A. Effects of Water Stress on Fruit Growth and the Partitioning of 13C-Assimilates in ‘Nijisseiki’ Pear Trees. J. Jpn. Soc. Hortic. Sci. 1999, 68, 1071–1078. [Google Scholar] [CrossRef]
- Li, N.; Kong, S.N.; Zhou, D.; Li, N.N.; Shang, J.L.; Wang, J.M.; Ma, S.W. Mapping and validation of a new quantitative trait locus (QTL) for fruit size in watermelon (Citrullus lanatus). Sci. Hortic. 2023, 318, 112054. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Z.S.; Yang, X.; Wang, Z.; Ma, L.; Tu, H.; Ma, Y.; Zhou, J.; Zhang, J.; Wang, H.; et al. Inheritance analysis of fruit-related traits in Chinese cherry [Cerasus pseudocerasus (Lindl.) G.Don] breeding progenies. Sci. Hortic. 2025, 307, 111519. [Google Scholar]
- Liu, N.W.; Wang, L.; Zhang, Z.Y.; Zhang, S.H.; Zhang, Q.; Wang, G.P.; Qin, L.; Cao, Q.Q. Analysis of inherited tendency of fruit characteristics in F1 group of reciprocal crossing between ‘Yanshanzaofeng’ and ‘Yanjing’ in Castanea mollissima. J. Beijing For. Univ. 2021, 43, 75–85. [Google Scholar]
- Peng, Z.; Zhao, C.; Li, S.; Guo, Y.; Xu, H.; Hu, G.; Liu, Z.; Chen, X.; Chen, J.; Lin, S.; et al. Integration of genomics, transcriptomics and metabolomics identifies candidate loci underlying fruit weight in loquat. Hortic. Res. 2022, 9, uhac037. [Google Scholar] [CrossRef]
- Morandi, B.; Losciale, P.; Manfrini, L.; Zibordi, M.; Anconelli, S.; Galli, F.; Pierpaoli, E.; Corelli Grappadelli, L. Increasing water stress negatively affects pear fruit growth by reducing first its xylem and then its phloem inflow. J. Plant Physiol. 2014, 171, 1500–1509. [Google Scholar] [CrossRef]
- Cyplik, A.; Bocianowski, J. Analytical and numerical comparisons of two methods of estimation of additive × additive interaction of QTL effects. Sci. Agric. 2012, 69, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Lozano, L.; Iglesias, I.; Puy, J.; Echeverria, G. Performance of an Expert Sensory Panel and Instrumental Measures for Assessing Eating Fruit Quality Attributes in a Pear Breeding Programme. Foods 2023, 12, 1426. [Google Scholar] [CrossRef]
- Luo, Y.; Chen, W.; Pan, Y.; Ge, L.; Wu, C.; Wang, J.; Liu, M.; Yan, F. Comparison and Genetic Variation Analysis of Important Fruit Traits in Jujube F1 Hybrids by Different Male Parents. Agronomy 2024, 14, 459. [Google Scholar] [CrossRef]
- Desnoues, E.; Baldazzi, V.; Génard, M.; Mauroux, J.; Lambert, P.; Confolent, C.; Quilot-Turion, B. Dynamic QTLs for sugars and enzyme activities provide an overview of genetic control of sugar metabolism during peach fruit development. J. Exp. Bot. 2016, 67, 3419–3431. [Google Scholar] [CrossRef]
- Altan, H.; Bircan, M.; Caliskan, O. Inheritance of earliness and fruit quality characteristics in five apricot progenies. Erwerbs-Obstbau 2022, 64, 591–601. [Google Scholar] [CrossRef]
- Lee, U.Y.; Kim, Y.K.; Shin, I.S.; Oh, K.S.; Jung, O.K.; Chun, J.P. Comparison of Fruit Development and Quality Indices According to Blossom Thinning on Early-season ‘Hanareum’ and Mid-season ‘Niitaka’ Pears. Hortic. Sci. Technol. 2015, 33, 486–491. [Google Scholar] [CrossRef]
- Li, J.; An, X.H.; Shao, J.Z.; Li, W.X.; Tian, Y. Inheritance of traits of hybrid seedlings from Wanbaihaitang × Huayue apple. Jiangsu Agric. Sci. 2023, 51, 169–175. [Google Scholar]
- Xu, J.H. The Comparison of Fruit Quality of Apple Cultivars and Genetic Tendency Analysis of Fruit Quality Characters in F1 Progenies. Master’s Thesis, Northwest A&F University, Xianyang, China, 2011. [Google Scholar]
- Zhao, Y.N.; Ou, C.Q.; Wang, F.; Zhang, Y.J.; Ma, L.; Jiang, S.L. Genetic Analysis and Correlation Study on Pear Fruit Characteristics in F1 Generation. Acta Agric. Broeali-Occident. Sin. 2018, 27, 1811–1818. [Google Scholar]
- Guo, K.L.; Liu, J.; Yu, X.M.; Yang, S. A preliminary study on genetic tendency of main fruit traits of pear crossbred offspring. J. Northwest AF Univ. (Nat. Sci. Ed.) 2024, 52, 97–108+119. [Google Scholar]
- Zhang, Y.; Li, S.; Gu, X.; Lei, D.; Zhao, B.; Tang, H.; Lin, Y.; Wang, Y.; Li, M.; Chen, Q. Anthocyanin Accumulation and Related Gene Expression Profile in ‘Red Zaosu’ Pear and Its Green Mutant. Agriculture 2021, 11, 898. [Google Scholar] [CrossRef]
- Tao, R.; Yu, W.; Gao, Y.; Ni, J.; Yin, L.; Zhang, X.; Li, H.; Wang, D.; Bai, S.; Teng, Y. Light-Induced Basic/Helix-Loop-Helix64 Enhances Anthocyanin Biosynthesis and Undergoes CONSTITUTIVELY PHOTOMORPHOGENIC1-Mediated Degradation in Pear. Plant Physiol. 2020, 184, 1684–1701. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Wang, S.; Yu, W.; Liao, Y.; Pan, C.; Zhang, M.; Tao, R.; Wei, J.; Gao, Y.; Wang, D.; et al. The ethylene-responsive transcription factor PpERF9 represses PpRAP2.4 and PpMYB114 via histone deacetylation to inhibit anthocyanin biosynthesis in pear. Plant Cell 2023, 35, 2271–2292. [Google Scholar] [CrossRef]
- Ma, Z.; Wei, C.; Cheng, Y.; Shang, Z.; Guo, X.; Guan, J. RNA-Seq Analysis Identifies Transcription Factors Involved in Anthocyanin Biosynthesis of ‘Red Zaosu’ Pear Peel and Functional Study of PpPIF8. Int. J. Mol. Sci. 2022, 23, 4798. [Google Scholar] [CrossRef]
- Xue, H.; Wang, S.; Yao, J.L.; Zhang, X.; Yang, J.; Wang, L.; Su, Y.; Chen, L.; Zhang, H.; Li, X. The genetic locus underlying red foliage and fruit skin traits is mapped to the same location in the two pear bud mutants ‘Red Zaosu’ and ‘Max Red Bartlett’. Hereditas 2018, 155, 25. [Google Scholar] [CrossRef]
- Reiland, H.; Slavin, J. Systematic Review of Pears and Health. Nutr. Today 2015, 50, 301–305. [Google Scholar] [CrossRef]
- Liu, Z.; Yang, S.; Hao, L.; Wang, H.; Zhang, J.; He, W.; Li, M.; Lin, Y.; Zhang, Y.; Chen, Q.; et al. Phenotypic Characters and Inheritance Tendency of Agronomic Traits in F1 Progeny of Chinese Cherry. Agronomy 2024, 14, 2862. [Google Scholar] [CrossRef]
- Alizadeh, K.; Fatholahi, S.; Teixeira da Silva, J.A. Variation in the fruit characteristics of local pear (Pyrus spp.) in the Northwest of Iran. Genet. Resour. Crop Evol. 2015, 62, 635–641. [Google Scholar] [CrossRef]
- Chenafi, A.; Monney, P.; Arrigoni, E.; Boudoukha, A.; Carlen, C. Influence of irrigation strategies on productivity, fruit quality and soil-plant water status of subsurface drip-irrigated apple trees. Fruits 2016, 71, 69–78. [Google Scholar] [CrossRef]
- Yang, G.; Sun, M.; Brewer, L.; Tang, Z.; Nieuwenhuizen, N.; Cooney, J.; Xu, S.; Sheng, J.; Andre, C.; Xue, C.; et al. Allelic variation of BBX24 is a dominant determinant controlling red coloration and dwarfism in pear. Plant Biotechnol. J. 2024, 22, 1468–1490. [Google Scholar] [CrossRef]
- Shen, F.; Huang, Z.; Zhang, B.; Wang, Y.; Zhang, X.; Wu, T.; Xu, X.; Zhang, X.; Han, Z. Mapping Gene Markers for Apple Fruit Ring Rot Disease Resistance Using a Multi-omics Approach. G3 2019, 9, 1663–1678. [Google Scholar] [CrossRef]
- Zheng, W.; Shen, F.; Wang, W.; Wu, B.; Wang, X.; Xiao, C.; Tian, Z.; Yang, X.; Yang, J.; Wang, Y.; et al. Quantitative trait loci-based genomics-assisted prediction for the degree of apple fruit cover color. Plant Genome 2020, 13, e20047. [Google Scholar] [CrossRef]
- Wen, G.; Wu, B.; Wang, Y.; Wu, T.; Han, Z.; Zhang, X. Natural variations in MdNAC18 exert major genetic effect on apple fruit harvest date by regulating ethylene biosynthesis genes. J. Integr. Plant Biol. 2024, 66, 2450–2469. [Google Scholar] [CrossRef]
- Liu, J.; Shen, F.; Xiao, Y.; Fang, H.; Qiu, C.; Li, W.; Wu, T.; Xu, X.; Wang, Y.; Zhang, X.; et al. Genomics-assisted prediction of salt and alkali tolerances and functional marker development in apple rootstocks. BMC Genom. 2020, 21, 550. [Google Scholar] [CrossRef]
- Zhou, Y.X.; Yue, Y.; Yu, C.L.; Zhang, Y.X.; Peng, J.Y. Fruit Quality Analysis, Excellent Line Selection and Quality Evaluation Methods of ‘Yuluxiang’ × ‘Xianghongli’ F1 Generation. Shandong Agric. Sci. 2024, 56, 24–33. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | FW/g | FLoD/mm | FLaD/mm | FSI | FSL/mm | TSS/% | TA/% | SS/% | FT | PC | FHS/mm | LLoD/mm | LLaD/mm | YLC | EFC | ABC | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2023 | Female | 256.96 a | 71.63 a | 75.07 a | 0.95 a | 37.95 a | 12.53 a | 0.20 a | 8.68 a | 2 | 2 | 24.76 a | 104.32 a | 72.16 a | 1 | 1 | 1 |
| Male | 165.47 b | 56.33 b | 63.75 b | 0.88 b | 18.66 b | 11.20 a | 0.11 b | 9.29 b | 1 | 5 | 26.11 a | 75.20 b | 50.15 b | 2 | 2 | 2 | |
| 2024 | Female | 243.52 a | 70.21 a | 72.35 a | 0.97 a | 13.02 a | 0.20 a | 8.24 a | 2 | 2 | 23.39 a | 101.58 a | 70.31 a | 1 | 1 | 1 | |
| Male | 136.23 b | 54.36 b | 60.68 b | 0.90 b | 11.47 b | 0.11 b | 9.17 b | 1 | 5 | 25.71 a | 74.31 b | 51.64 b | 2 | 2 | 2 |
| Trait | Abbr | Year | Sample Size | Minimum | Maximum | Mean | Standard Deviation | CV/% | Skewness | Kurtosis |
|---|---|---|---|---|---|---|---|---|---|---|
| Fruit weight/g | FW | 2023 | 140 | 71.93 | 364.33 | 185.10 | 59.94 | 32.38 | 0.48 | 0.34 |
| 2024 | 141 | 57.80 | 317.53 | 174.38 | 49.85 | 28.59 | 0.45 | 0.29 | ||
| Fruit longitudinal diameter/mm | FLoD | 2023 | 140 | 49.58 | 85.67 | 65.69 | 8.17 | 12.44 | 0.55 | 0.16 |
| 2024 | 141 | 48.63 | 83.76 | 64.18 | 7.95 | 12.39 | 0.39 | 0.49 | ||
| Fruit lateral diameter/mm | FLaD | 2023 | 140 | 52.27 | 92.82 | 69.74 | 8.42 | 12.08 | 0.38 | 0.46 |
| 2024 | 141 | 51.16 | 88.61 | 68.10 | 8.06 | 11.83 | 0.35 | 0.52 | ||
| Fruit shape index | FSI | 2023 | 140 | 0.76 | 1.14 | 0.94 | 0.08 | 7.96 | 0.27 | 0.44 |
| 2024 | 141 | 0.75 | 1.13 | 0.94 | 0.07 | 7.20 | 0.01 | 0.19 | ||
| Total soluble solids/% | TSS | 2023 | 134 | 9.70 | 15.50 | 12.17 | 1.06 | 8.69 | 0.15 | 0.37 |
| 2024 | 138 | 8.94 | 14.80 | 11.72 | 1.11 | 9.51 | 0 | 0.32 | ||
| Titratable acid/% | TA | 2023 | 136 | 0.03 | 0.55 | 0.19 | 0.10 | 54.42 | 0.32 | −0.38 |
| 2024 | 138 | 0.05 | 0.49 | 0.25 | 0.10 | 39.17 | 0.24 | −0.60 | ||
| Soluble sugar/% | SS | 2023 | 135 | 3.65 | 12.95 | 7.75 | 1.80 | 23.25 | 0.10 | −0.09 |
| 2024 | 137 | 4.03 | 12.76 | 7.91 | 1.75 | 22.13 | 0.06 | −0.25 | ||
| Fruit heart size/mm | FHS | 2023 | 135 | 17.30 | 33.61 | 25.09 | 3.32 | 13.22 | 0.37 | −0.19 |
| 2024 | 136 | 16.87 | 33.51 | 24.57 | 3.27 | 13.29 | 0.43 | −0.14 | ||
| Fruit stalk length/mm | FSL | 2023 | 138 | 13.60 | 58.17 | 35.97 | 8.45 | 23.50 | 0.05 | 0.01 |
| Leaf longitudinal diameter/mm | LLoD | 2023 | 141 | 62.84 | 114.31 | 88.81 | 9.02 | 10.17 | −0.07 | −0.06 |
| 2024 | 148 | 53.93 | 103.76 | 73.16 | 9.73 | 13.30 | 0.78 | 0.94 | ||
| Leaf lateral diameter/mm | LLaD | 2023 | 141 | 45.29 | 79.29 | 60.88 | 6.51 | 10.95 | 0.08 | −0.66 |
| 2024 | 148 | 34.57 | 74.08 | 50.71 | 7.39 | 14.58 | 0.60 | 0.39 |
| Trait | Year | Female | Male | MP | Mean | Hb2/% | Ta/% | Hm/% | OHP/% | BLP/% |
|---|---|---|---|---|---|---|---|---|---|---|
| FW/g | 2023 | 256.96 | 165.47 | 211.21 | 185.10 | 60.47 | 87.64 | −12.36 | 9.29 | 54.29 |
| 2024 | 243.52 | 136.23 | 189.88 | 174.38 | 55.23 | 91.84 | −8.16 | 11.35 | 22.70 | |
| FLoD/mm | 2023 | 71.63 | 56.92 | 64.28 | 65.76 | 27.17 | 97.84 | 2.21 | 20.00 | 3.57 |
| 2024 | 70.21 | 54.36 | 62.29 | 64.18 | 32.31 | 103.04 | 3.04 | 23.40 | 7.80 | |
| FLaD/mm | 2023 | 75.07 | 75.07 | 75.07 | 69.81 | 61.42 | 99.52 | 0.48 | 27.14 | 27.14 |
| 2024 | 72.35 | 60.68 | 66.52 | 68.10 | 46.78 | 102.38 | −31.90 | 36.17 | 21.28 | |
| FSI | 2023 | 0.95 | 0.89 | 0.92 | 0.94 | 34.25 | 102.23 | 2.23 | 41.43 | 19.29 |
| 2024 | 0.97 | 0.90 | 0.93 | 0.94 | 33.21 | 100.74 | 0.74 | 36.17 | 27.66 | |
| TSS/% | 2023 | 12.43 | 11.20 | 11.82 | 12.17 | 48.83 | 97.09 | 2.99 | 38.81 | 14.93 |
| 2024 | 13.02 | 11.47 | 12.25 | 11.72 | 52.91 | 95.71 | −4.29 | 12.32 | 39.13 | |
| TA/% | 2023 | 0.20 | 0.11 | 0.15 | 0.19 | 66.67 | 80.87 | 23.65 | 36.76 | 20.59 |
| 2024 | 0.20 | 0.11 | 0.16 | 0.25 | 51.84 | 159.24 | 59.24 | 60.87 | 10.14 | |
| SS/% | 2023 | 8.68 | 9.29 | 8.99 | 7.75 | 57.38 | 116.01 | −13.80 | 18.52 | 70.37 |
| 2024 | 8.24 | 9.17 | 8.17 | 7.91 | 49.25 | 90.87 | −9.13 | 23.36 | 58.39 | |
| FHS/mm | 2023 | 24.76 | 26.11 | 25.43 | 25.09 | 56.27 | 101.37 | −1.35 | 33.33 | 54.81 |
| 2024 | 23.39 | 25.71 | 24.55 | 24.57 | 43.82 | 100.08 | 0.08 | 33.09 | 42.65 | |
| FSL/mm | 2023 | 37.95 | 18.66 | 28.31 | 35.97 | 85.42 | 78.69 | 27.08 | 40.58 | 2.17 |
| LLoD/mm | 2023 | 104.32 | 75.20 | 89.76 | 88.81 | 69.22 | 101.07 | −1.06 | 4.05 | 6.76 |
| 2024 | 101.58 | 74.31 | 87.95 | 73.16 | 62.81 | 65.05 | −16.81 | 1.35 | 58.78 | |
| LLaD/mm | 2023 | 72.16 | 50.15 | 61.16 | 60.88 | 61.27 | 100.45 | −0.45 | 6.76 | 4.73 |
| 2024 | 70.31 | 51.64 | 60.98 | 50.71 | 55.47 | 83.17 | −16.83 | 1.35 | 55.41 |
| Principal Component | Eigenvalue | Contribution/% | Cumulative Percentage/% |
|---|---|---|---|
| PCA1 | 6.346 | 24.41% | 24.41% |
| PCA2 | 4.076 | 15.68% | 40.08% |
| PCA3 | 2.674 | 10.28% | 50.37% |
| PCA4 | 2.226 | 8.56% | 58.93% |
| PCA5 | 2.039 | 7.84% | 66.77% |
| PCA6 | 1.721 | 6.62% | 73.39% |
| Indexes | PCA1 | PCA2 | PCA3 | PCA4 | PCA5 | PCA6 |
|---|---|---|---|---|---|---|
| 2023FW | −0.946 | 0.019 | −0.121 | 0.096 | −0.129 | 0.081 |
| 2024FW | −0.945 | 0.076 | −0.100 | 0.085 | −0.107 | 0.076 |
| 2023FLoD | 0.059 | 0.525 | 0.538 | −0.495 | 0.071 | −0.214 |
| 2024FLoD | 0.062 | 0.448 | 0.568 | −0.494 | 0.089 | −0.208 |
| 2023FLaD | −0.406 | −0.718 | 0.108 | −0.019 | −0.013 | −0.212 |
| 2024FLaD | −0.064 | −0.244 | −0.318 | 0.166 | 0.035 | −0.191 |
| 2023FSI | −0.451 | 0.307 | −0.107 | 0.373 | 0.421 | 0.308 |
| 2024FSI | −0.460 | 0.309 | −0.089 | 0.383 | 0.410 | 0.309 |
| PC | 0.242 | 0.120 | 0.203 | 0.552 | −0.492 | −0.146 |
| FT | 0.239 | 0.055 | 0.175 | 0.598 | −0.473 | −0.278 |
| 2023FHS | 0.042 | 0.342 | 0.124 | −0.301 | −0.067 | −0.080 |
| 2924FHS | −0.371 | −0.730 | 0.229 | −0.031 | 0.021 | −0.273 |
| 2023TSS | −0.363 | −0.622 | 0.111 | 0.002 | 0.070 | −0.433 |
| 2024TSS | −0.384 | −0.713 | 0.142 | −0.033 | 0.128 | −0.343 |
| 2023FSL | −0.247 | 0.194 | −0.446 | −0.120 | −0.427 | −0.210 |
| EFC | −0.148 | 0.297 | −0.427 | −0.065 | −0.231 | −0.261 |
| YLC | −0.016 | 0.414 | −0.469 | −0.075 | −0.311 | −0.159 |
| ABC | 0.080 | 0.403 | −0.414 | −0.141 | −0.315 | −0.298 |
| 2023LLoD | 0.042 | 0.364 | −0.303 | 0.171 | 0.592 | −0.537 |
| 2024LLoD | 0.040 | 0.372 | −0.294 | 0.178 | 0.589 | −0.541 |
| 2023LLaD | −0.048 | 0.387 | 0.585 | 0.471 | −0.041 | −0.150 |
| 2024LLaD | −0.018 | 0.374 | 0.584 | 0.478 | −0.043 | −0.168 |
| 2023TA | −0.946 | 0.019 | −0.120 | 0.096 | −0.129 | 0.080 |
| 2024TA | −0.945 | 0.076 | −0.100 | 0.084 | −0.106 | 0.076 |
| 2023SS | 0.059 | 0.524 | 0.538 | −0.495 | 0.071 | −0.214 |
| 2024SS | 0.062 | 0.448 | 0.568 | −0.494 | 0.089 | −0.209 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Tang, M.; Li, J.; Chi, Y.; Wang, K.; Peng, J.; Zhang, Y. Phenotypic Characters and Inheritance Tendency of Agronomic Traits in F1 Progeny of Pear. Plants 2025, 14, 1491. https://doi.org/10.3390/plants14101491
Zhang X, Tang M, Li J, Chi Y, Wang K, Peng J, Zhang Y. Phenotypic Characters and Inheritance Tendency of Agronomic Traits in F1 Progeny of Pear. Plants. 2025; 14(10):1491. https://doi.org/10.3390/plants14101491
Chicago/Turabian StyleZhang, Xiaojie, Mengyue Tang, Jiamei Li, Yue Chi, Kexin Wang, Jianying Peng, and Yuxing Zhang. 2025. "Phenotypic Characters and Inheritance Tendency of Agronomic Traits in F1 Progeny of Pear" Plants 14, no. 10: 1491. https://doi.org/10.3390/plants14101491
APA StyleZhang, X., Tang, M., Li, J., Chi, Y., Wang, K., Peng, J., & Zhang, Y. (2025). Phenotypic Characters and Inheritance Tendency of Agronomic Traits in F1 Progeny of Pear. Plants, 14(10), 1491. https://doi.org/10.3390/plants14101491