
Anticholinesterase Activity of Methanolic Extract of Amorpha fruticosa Flowers and Isolation of Rotenoids and Putrescine and Spermidine Derivatives

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Isolation and Identification of Chemical Compounds and Quantification of Compound 5
2.2. Inhibition of Acetylcholinesterase (AChE) and Butyrylcholinesterase (BuChE)
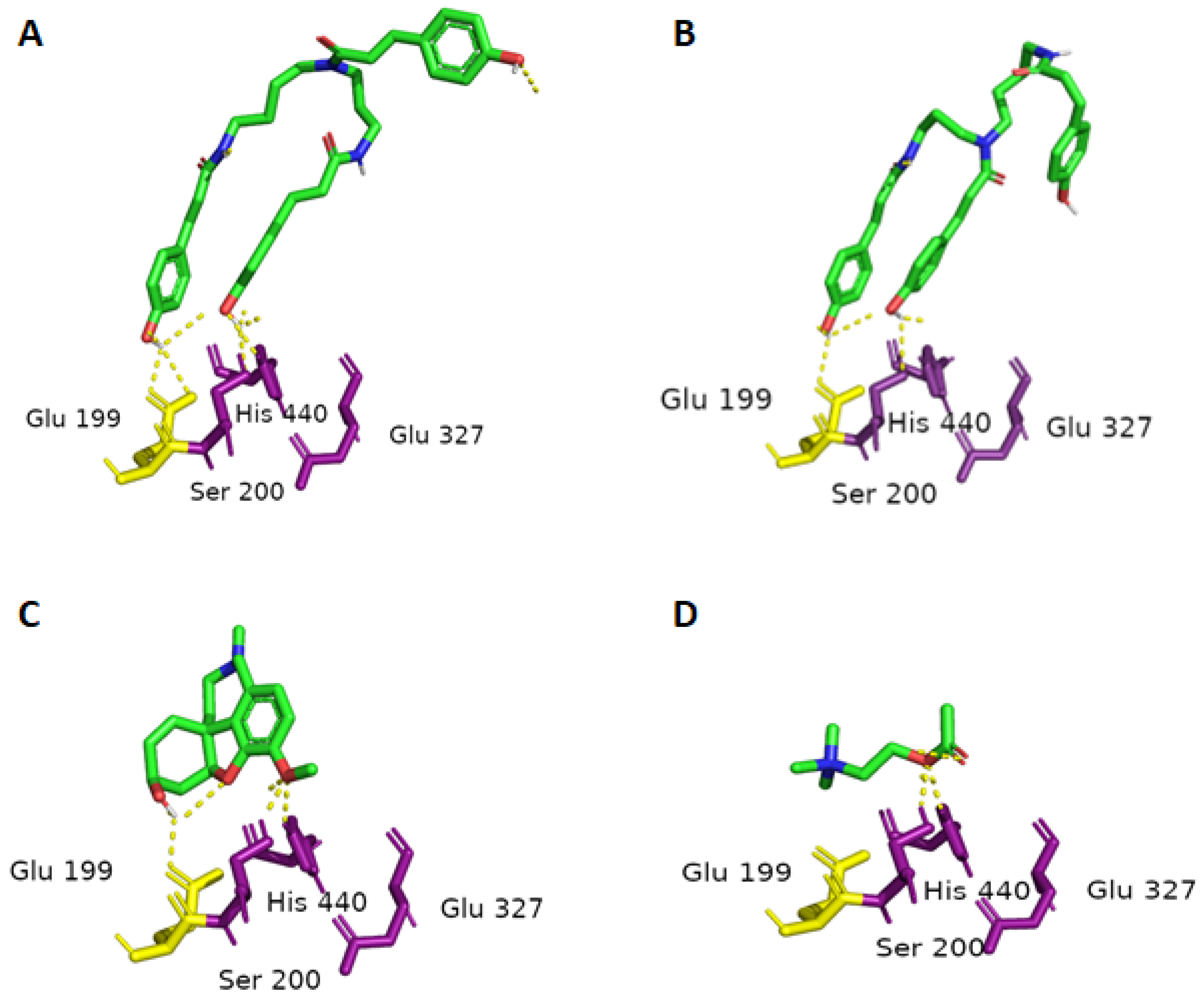
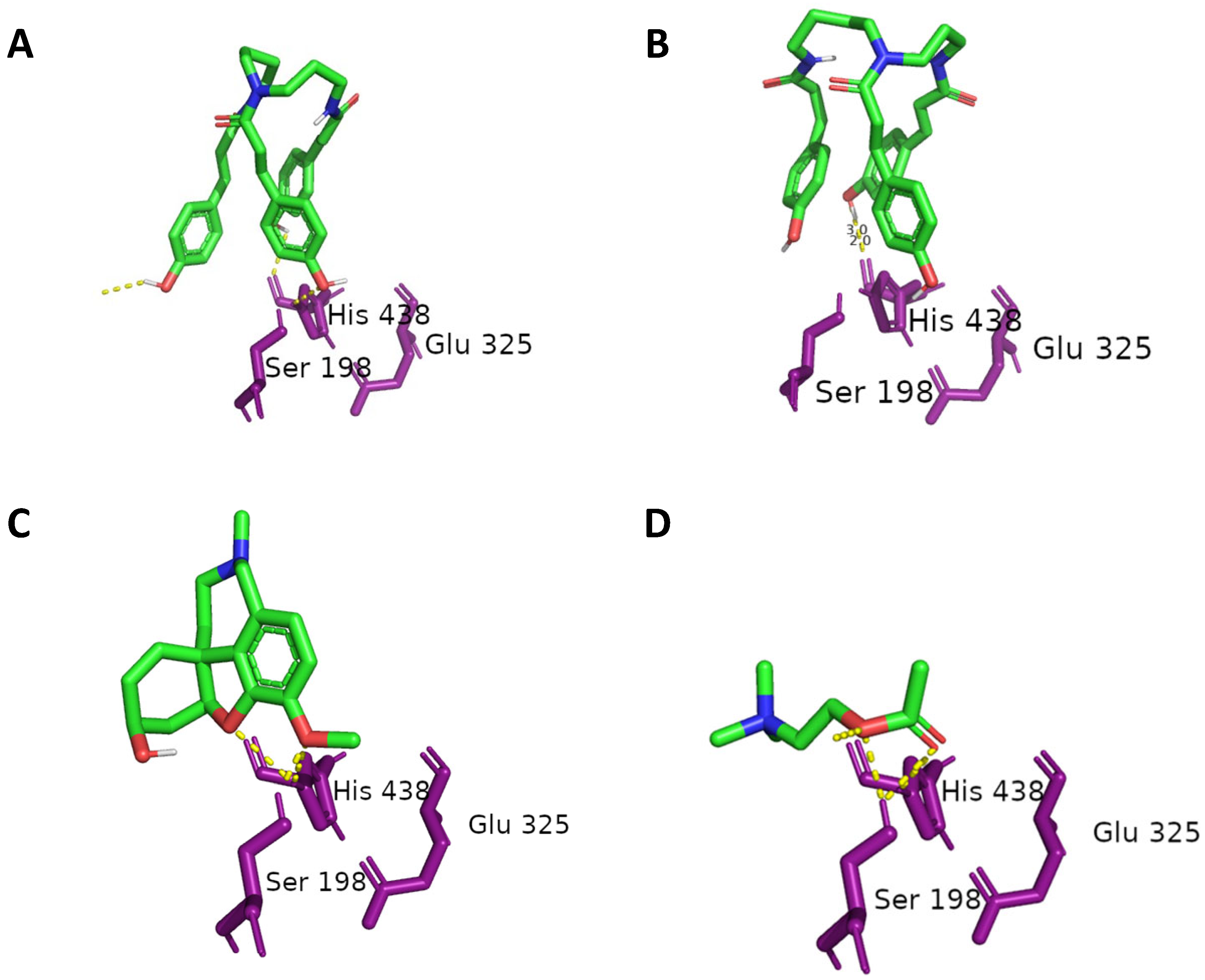
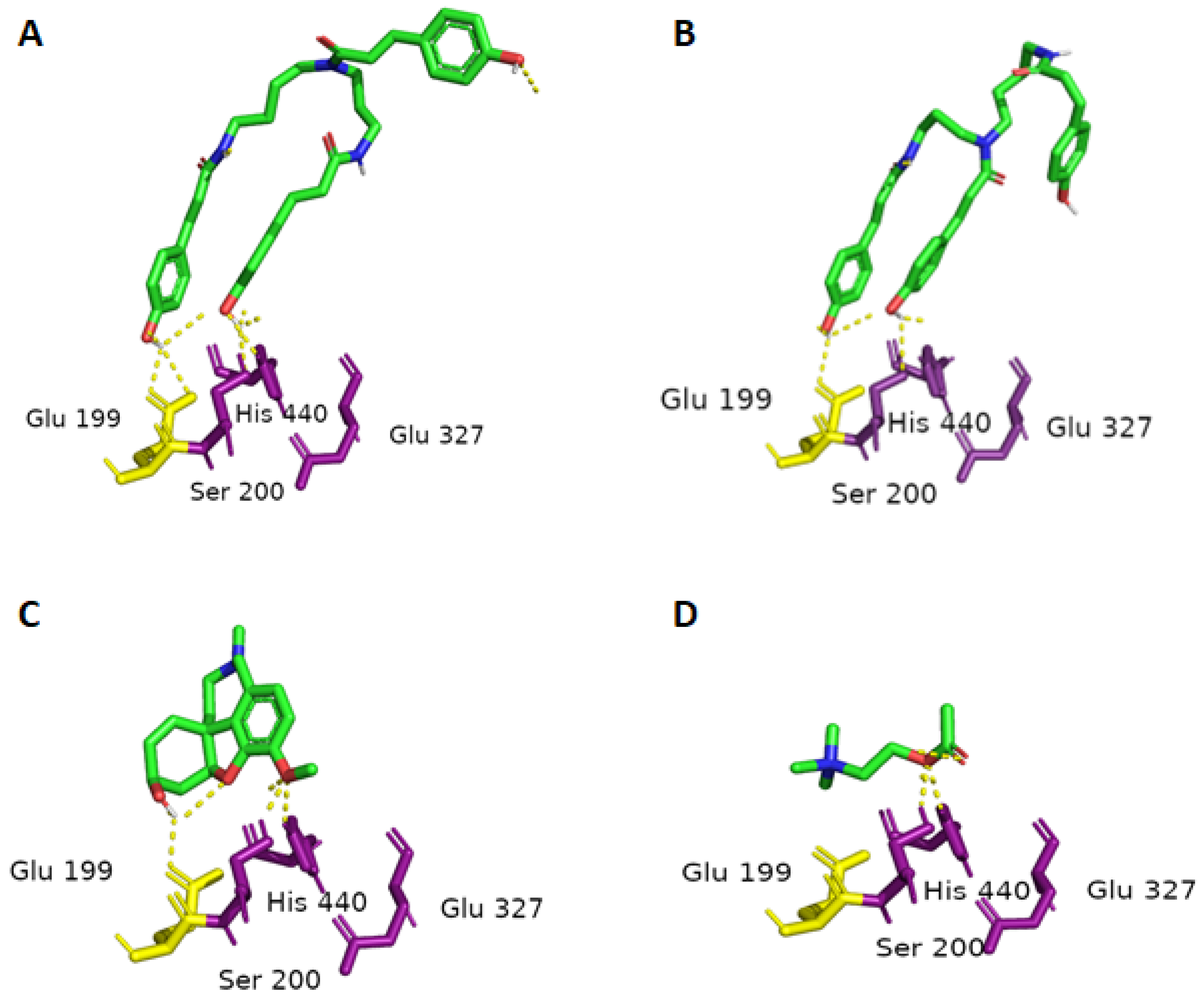
2.3. Molecular Docking
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Plant Material
4.3. Extraction and Isolation
4.4. Quantification of Compound 5 by HPLC-DAD
4.5. Anticholinesterase Activity
4.6. Molecular Docking
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kozuharova, E.; Benbassat, N.; Ionkova, I. The invasive alien species Amorpha fruticosa in Bulgaria and its potential as economically prospective source of valuable essential oil. Pharmacia 2020, 67, 357–362. [Google Scholar] [CrossRef]
- Kozuharova, E.; Matkowski, A.; Woźniak, D.; Simeonova, R.; Naychov, Z.; Malainer, C.; Mocan, A.; Nabavi, S.M.; Atanasov, A.G. Amorpha fruticosa—A Noxious Invasive Alien Plant in Europe or a Medicinal Plant against Metabolic Disease? Front. Pharmacol. 2017, 8, 333. [Google Scholar] [CrossRef] [PubMed]
- Lis, A.; Góra, J. Essential Oil of Amorpha fruticosa L. J. Essent. Oil Res. 2001, 13, 340–342. [Google Scholar] [CrossRef]
- Konoshima, T.; Terada, H.; Kokumai, M.; Kozuka, M.; Tokuda, H.; Estes, J.R.; Li, L.; Wang, H.-K.; Lee, K.-H. Studies on Inhibitors of Skin Tumor Promotion, XII. Rotenoids from Amorpha fruticosa. J. Nat. Prod. 1993, 56, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Muharini, R.; Díaz, A.; Ebrahim, W.; Mándi, A.; Kurtán, T.; Rehberg, N.; Kalscheuer, R.; Hartmann, R.; Orfali, R.S.; Lin, W.; et al. Antibacterial and Cytotoxic Phenolic Metabolites from the Fruits of Amorpha fruticosa. J. Nat. Prod. 2017, 80, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Zheleva-Dimitrova, D. Antioxidant and acetylcholinesterase inhibition properties of Amorpha fruticosa L. and Phytolacca americana L. Pharmacogn. Mag. 2013, 9, 109. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y.; Kim, P.S.; Park, J.; Yoo, E.S.; Baik, K.U.; Kim, Y.-K.; Park, M.H. Inhibitor of tumor necrosis factor-α production in lipopolysaccharide-stimulated RAW264.7 cells from Amorpha fruticosa. J. Ethnopharmacol. 2000, 70, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Dat, N.T.; Lee, J.-H.; Lee, K.; Hong, Y.-S.; Kim, Y.H.; Lee, J.J. Phenolic Constituents of Amorpha fruticosa That Inhibit NF-κB Activation and Related Gene Expression. J. Nat. Prod. 2008, 71, 1696–1700. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.W.; Kim, J.Y.; Lee, S.H.; Im, S.Y.; Lee, G.; Park, K.H. New Class of Tyrosinase Inhibitors, Rotenoids, from Amorpha fruticosa. ACS Omega 2023, 8, 31870–31879. [Google Scholar] [CrossRef]
- Weidner, C.; De Groot, J.C.; Prasad, A.; Freiwald, A.; Quedenau, C.; Kliem, M.; Witzke, A.; Kodelja, V.; Han, C.-T.; Giegold, S.; et al. Amorfrutins are potent antidiabetic dietary natural products. Proc. Natl. Acad. Sci. USA 2012, 109, 7257–7262. [Google Scholar] [CrossRef] [PubMed]
- Bassard, J.-E.; Ullmann, P.; Bernier, F.; Werck-Reichhart, D. Phenolamides: Bridging polyamines to the phenolic metabolism. Phytochemistry 2010, 71, 1808–1824. [Google Scholar] [CrossRef] [PubMed]
- Kyselka, J.; Bleha, R.; Dragoun, M.; Bialasová, K.; Horáčková, Š.; Schätz, M.; Sluková, M.; Filip, V.; Synytsya, A. Antifungal Polyamides of Hydroxycinnamic Acids from Sunflower Bee Pollen. J. Agric. Food Chem. 2018, 66, 11018–11026. [Google Scholar] [CrossRef] [PubMed]
- Roumani, M.; Duval, R.E.; Ropars, A.; Risler, A.; Robin, C.; Larbat, R. Phenolamides: Plant specialized metabolites with a wide range of promising pharmacological and health-promoting interests. Biomed. Pharmacother. 2020, 131, 110762. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Gu, C.; Liu, J. Nature spermidine and spermine alkaloids: Occurrence and pharmacological effects. Arab. J. Chem. 2022, 15, 104367. [Google Scholar] [CrossRef]
- Nugroho, A.; Choi, J.S.; Hong, J.-P.; Park, H.-J. Anti-acetylcholinesterase activity of the aglycones of phenolic glycosides isolated from Leonurus japonicus. Asian Pac. Trop. Biomed. 2017, 7, 849–854. [Google Scholar] [CrossRef]
- Greig, N.H.; Lahiri, D.K.; Sambamurti, K. Butyrylcholinesterase: An Important New Target in Alzheimer’s Disease Therapy. Int. Psychogeriatr. 2002, 14, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.B.; Liu, Q.; Ahn, J.H.; Jo, Y.H.; Turk, A.; Hong, I.P.; Han, S.M.; Hwang, B.Y.; Lee, M.K. Polyamine derivatives from the bee pollen of Quercus mongolica with tyrosinase inhibitory activity. Bioorg. Chem. 2018, 81, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.W.; Lee, S.K.; Kim, E.O.; Oh, J.H.; Yoon, K.S.; Parris, N.; Hicks, K.B.; Moreau, R.A. Antioxidant and Antimelanogenic Activities of Polyamine Conjugates from Corn Bran and Related Hydroxycinnamic Acids. J. Agric. Food Chem. 2007, 55, 3920–3925. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.-S.; Lü, L.; Yang, Y.-J.; Zhang, J.-L.; Zhang, P.-C. New spermidines from the florets of Carthamus tinctorius. J. Asian Nat. Prod. Res. 2008, 10, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Qin, G.-W.; Gai, Y.; Guo, L.-H. Structural Identification of a New Tri-p-coumaroylspermidine with Serotonin Transporter Inhibition from Safflower. Chem. Pharm. Bull. 2010, 58, 950–952. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.-M.; Nakamura, N.; Hattori, M. Inhibitory Effects on HIV-1 Protease of Tri-p-coumaroylspermidine from Artemisia caruifolia and Related Amides. Chem. Pharm. Bull. 2001, 49, 915–917. [Google Scholar] [CrossRef] [PubMed]
- Dagne, E.; Yenesew, A.; Waterman, P.G. Flavonoids and isoflavonoids from Tephrosia fulvinervis and Tephrosia pentaphylla. Phytochemistry 1989, 28, 3207–3210. [Google Scholar] [CrossRef]
- Lin, L.-J.; Ruangrungsi, N.; Cordell, G.A.; Shieh, H.-L.; You, M.; Pezzuto, J.M. 6-Deoxyclitoriacetal from Clitoria macrophylla. Phytochemistry 1992, 31, 4329–4331. [Google Scholar] [CrossRef]
- Terada, H.; Kokumai, M.; Konoshima, T.; Kozuka, M.; Haruna, M.; Ito, K.; Estes, J.R.; Li, L.; Wang, H.-K.; Lee, K.-H. Structural Elucidation and Chemical Conversion of Amorphispironone, a Novel Spironone from Amorpha fruticosa, to Rotenoids. Chem. Pharm. Bull. 1993, 41, 187–190. [Google Scholar] [CrossRef]
- Andrei, C.C.; Vieira, P.C.; Fernandes, J.B.; Da Silva, M.F.D.G.F.; Rodrigues^Fo, E. Dimethylchromene rotenoids from Tephrosia candida. Phytochemistry 1997, 46, 1081–1085. [Google Scholar] [CrossRef]
- Colletier, J.-P.; Royant, A.; Specht, A.; Sanson, B.; Nachon, F.; Masson, P.; Zaccai, G.; Sussman, J.L.; Goeldner, M.; Silman, I.; et al. Use of a “caged” analogue to study the traffic of choline within acetylcholinesterase by kinetic crystallography. Acta Cryst. D 2007, 63, 1115–1128. [Google Scholar] [CrossRef] [PubMed]
- Bajda, M.; Więckowska, A.; Hebda, M.; Guzior, N.; Sotriffer, C.A.; Malawska, B. Structure-Based Search for New Inhibitors of Cholinesterases. Int. J. Mol. Sci. 2013, 14, 5608–5632. [Google Scholar] [CrossRef] [PubMed]
- Khongkarat, P.; Ramadhan, R.; Phuwapraisirisan, P.; Chanchao, C. Safflospermidines from the bee pollen of Helianthus annuus L. exhibit a higher in vitro antityrosinase activity than kojic acid. Heliyon 2020, 6, e03638. [Google Scholar] [CrossRef] [PubMed]
- Vukašinović, E.L.; Kebert, M.; Radišić, P.; Đorđievski, S.; Čelić, T.V.; Pihler, I.; Kojić, D.; Purać, J. Polyamine profiling in honey bee products: A contribution to functional nutrition. J. Food Compos. Anal. 2024, 125, 105856. [Google Scholar] [CrossRef]
- Zhou, Z.-Q.; Fan, H.-X.; He, R.-R.; Xiao, J.; Tsoi, B.; Lan, K.-H.; Kurihara, H.; So, K.-F.; Yao, X.-S.; Gao, H. Lycibarbarspermidines A–O, New Dicaffeoylspermidine Derivatives from Wolfberry, with Activities against Alzheimer’s Disease and Oxidation. J. Agric. Food Chem. 2016, 64, 2223–2237. [Google Scholar] [CrossRef]
- Kim, E.O.; Min, K.J.; Kwon, T.K.; Um, B.H.; Moreau, R.A.; Choi, S.W. Anti-inflammatory activity of hydroxycinnamic acid derivatives isolated from corn bran in lipopolysaccharide-stimulated Raw 264.7 macrophages. Food Chem. Toxicol. 2012, 50, 1309–1316. [Google Scholar] [CrossRef]
- Szwajgier, D.; Borowiec, K. Phenolic acids from malt are efficient acetylcholinesterase and butyrylcholinesterase inhibitors: Phenolic acids as efficient AChE and BChE inhibitors. J. Inst. Brew. 2012, 118, 40–48. [Google Scholar] [CrossRef]
- Tu, Y.; Wu, C.; Kang, Y.; Li, Q.; Zhu, C.; Li, Y. Bioactivity-guided identification of flavonoids with cholinesterase and β-amyloid peptide aggregation inhibitory effects from the seeds of Millettia pachycarpa. Bioorg. Med. Chem. Lett. 2019, 29, 1194–1198. [Google Scholar] [CrossRef]
- Malaník, M.; Farková, V.; Křížová, J.; Kresová, A.; Šmejkal, K.; Kašparovský, T.; Dadáková, K. Comparison of Metabolic Profiles of Fruits of Arctium lappa, Arctium minus, and Arctium tomentosum. Plant Foods Hum. Nutr. 2024. [Google Scholar] [CrossRef]
- Fan, P.; Terrier, L.; Hay, A.-E.; Marston, A.; Hostettmann, K. Antioxidant and enzyme inhibition activities and chemical profiles of Polygonum sachalinensis F.Schmidt ex Maxim (Polygonaceae). Fitoterapia 2010, 81, 124–131. [Google Scholar] [CrossRef]
- Kubínová, R.; Švajdlenka, E.; Jankovská, D. Anticholinesterase, antioxidant activity and phytochemical investigation into aqueous extracts from five species of Agrimonia genus. Nat. Prod. Res. 2016, 30, 1174–1177. [Google Scholar] [CrossRef]
- Dallakyan, S.; Olson, A.J. Small-Molecule Library Screening by Docking with PyRx. In Chemical Biology: Methods and Protocols; Hempel, J.E., Williams, C.H., Hong, C.C., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2015; pp. 243–250. ISBN 978-1-4939-2269-7. [Google Scholar]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Schrodinger, L. The PyMOL Molecular Graphics System; Version 2.5; DeLano Scientific LLC: San Francisco, CA, USA, 2010. [Google Scholar]

{kind=link}
{kind=link}
| Compound | % Inhibition of AChE | % Inhibition of BuChE |
|---|---|---|
| 1 | 9.1 ± 3.2 | 8.1 ± 5.9 |
| 2 | 1.1 ± 0.7 | 4.8 ± 3.3 |
| 3 | 14.1 ± 3.4 | 4.5 ± 2.5 |
| 4 | 26.6 ± 2.5 | 19.1 ± 2.1 |
| 5 | 47.9 ± 2.1 | 43.8 ± 1.9 |
| 6 | 17.9 ± 5.9 | 23.8 ± 5.2 |
| 7 | 37.9 ± 5.0 | 25.6 ± 5.6 |
| 8 | N.I. | 37.6 ± 2.7 |
| 9 | N.I. | 23.6 ± 2.6 |
| 10 | 22.4 ± 4.9 | 22.8 ± 5.9 |
| galantamine | 82.1 ± 0.9 | 62.8 ± 0.3 |
| Compound | Binding Affinity to AChE [Kcal/mol] | Binding Affinity to BuChE [Kcal/mol] |
|---|---|---|
| trans-trans-trans | −10.8 | −9.4 |
| trans-trans-cis | −10.4 | −9.6 |
| trans-cis-trans | −10.8 | −10.4 |
| trans-cis-cis | −11.3 | −10.3 |
| cis-trans-trans | −11.3 | −10.2 |
| cis-trans-cis | −11.3 | −9.5 |
| cis-cis-trans | −11.4 | −9.9 |
| cis-cis-cis | −11.2 | −10.0 |
| trans-trans | −9.8 | −8.4 |
| trans-cis | −9.6 | −9.6 |
| cis-trans | −10.0 | −8.7 |
| cis-cis | −10.1 | −8.5 |
| Galantamine | −10.5 | −8.6 |
| Acetylcholine | −5.1 | −4.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jankovská, D.; Jurčová, N.; Kubínová, R.; Václavík, J.; Švajdlenka, E.; Mascellani, A.; Maršík, P.; Bouzková, K.; Malaník, M. Anticholinesterase Activity of Methanolic Extract of Amorpha fruticosa Flowers and Isolation of Rotenoids and Putrescine and Spermidine Derivatives. Plants 2024, 13, 1181. https://doi.org/10.3390/plants13091181
Jankovská D, Jurčová N, Kubínová R, Václavík J, Švajdlenka E, Mascellani A, Maršík P, Bouzková K, Malaník M. Anticholinesterase Activity of Methanolic Extract of Amorpha fruticosa Flowers and Isolation of Rotenoids and Putrescine and Spermidine Derivatives. Plants. 2024; 13(9):1181. https://doi.org/10.3390/plants13091181
Chicago/Turabian StyleJankovská, Dagmar, Nikol Jurčová, Renata Kubínová, Jiří Václavík, Emil Švajdlenka, Anna Mascellani, Petr Maršík, Kateřina Bouzková, and Milan Malaník. 2024. "Anticholinesterase Activity of Methanolic Extract of Amorpha fruticosa Flowers and Isolation of Rotenoids and Putrescine and Spermidine Derivatives" Plants 13, no. 9: 1181. https://doi.org/10.3390/plants13091181
APA StyleJankovská, D., Jurčová, N., Kubínová, R., Václavík, J., Švajdlenka, E., Mascellani, A., Maršík, P., Bouzková, K., & Malaník, M. (2024). Anticholinesterase Activity of Methanolic Extract of Amorpha fruticosa Flowers and Isolation of Rotenoids and Putrescine and Spermidine Derivatives. Plants, 13(9), 1181. https://doi.org/10.3390/plants13091181