
Process-Based Modeling of Phenology and Radial Growth in Pinus tabuliformis in Response to Climate Factors over a Cold and Semi-Arid Region

 ,
,
Abstract
1. Introduction
2. Materials and Methods
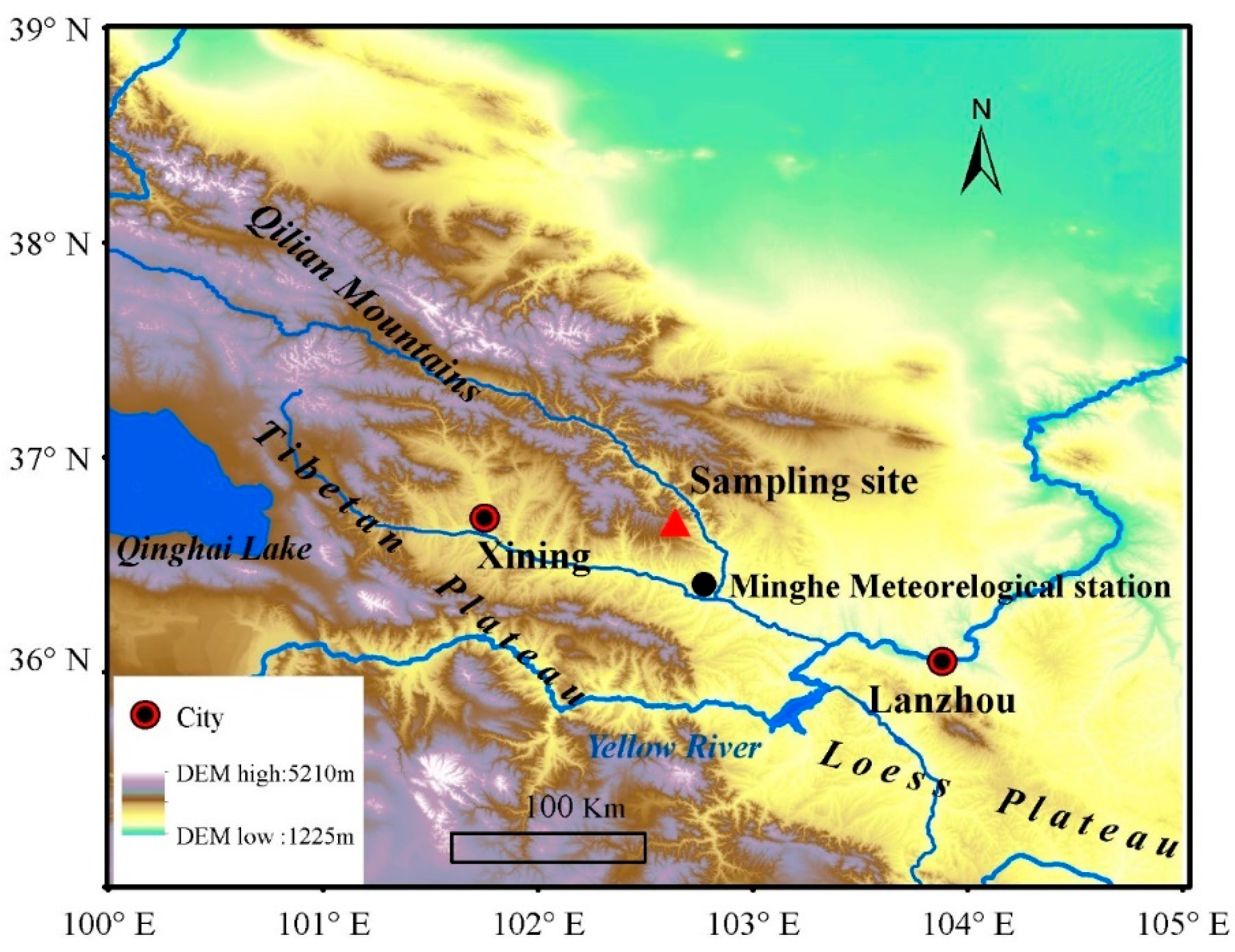
2.1. Study Region
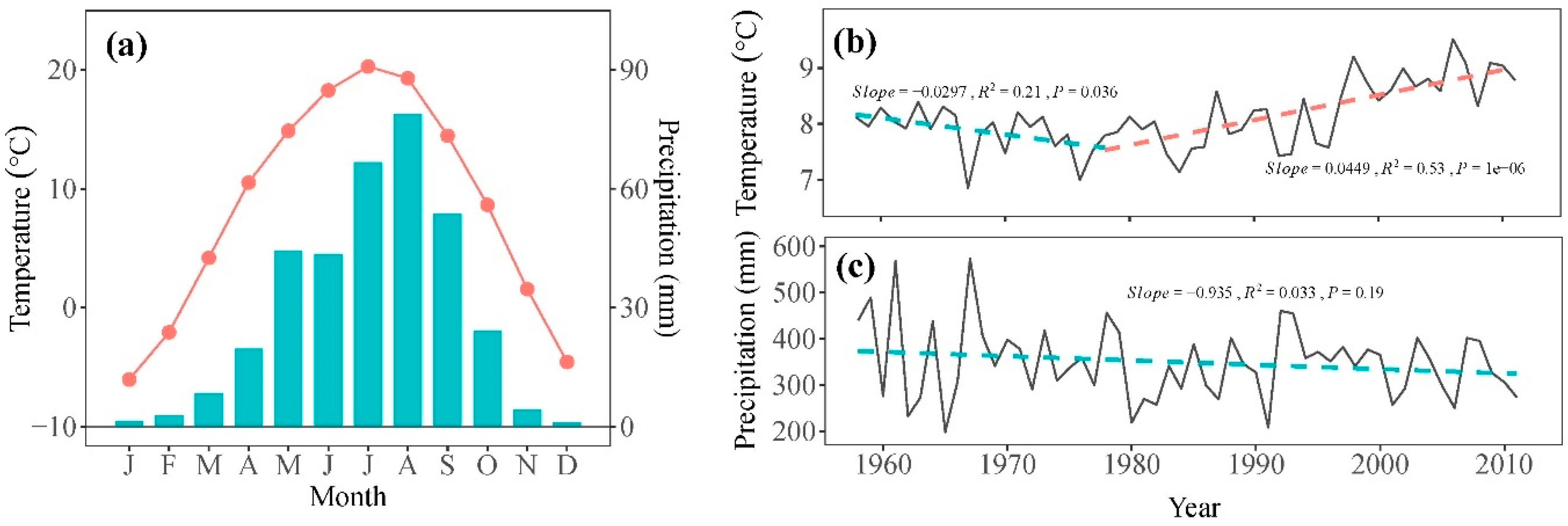
2.2. Meteorological Data
2.3. Tree-Ring Data
2.4. VS Model
2.5. Data Analysis and Statistics
3. Results
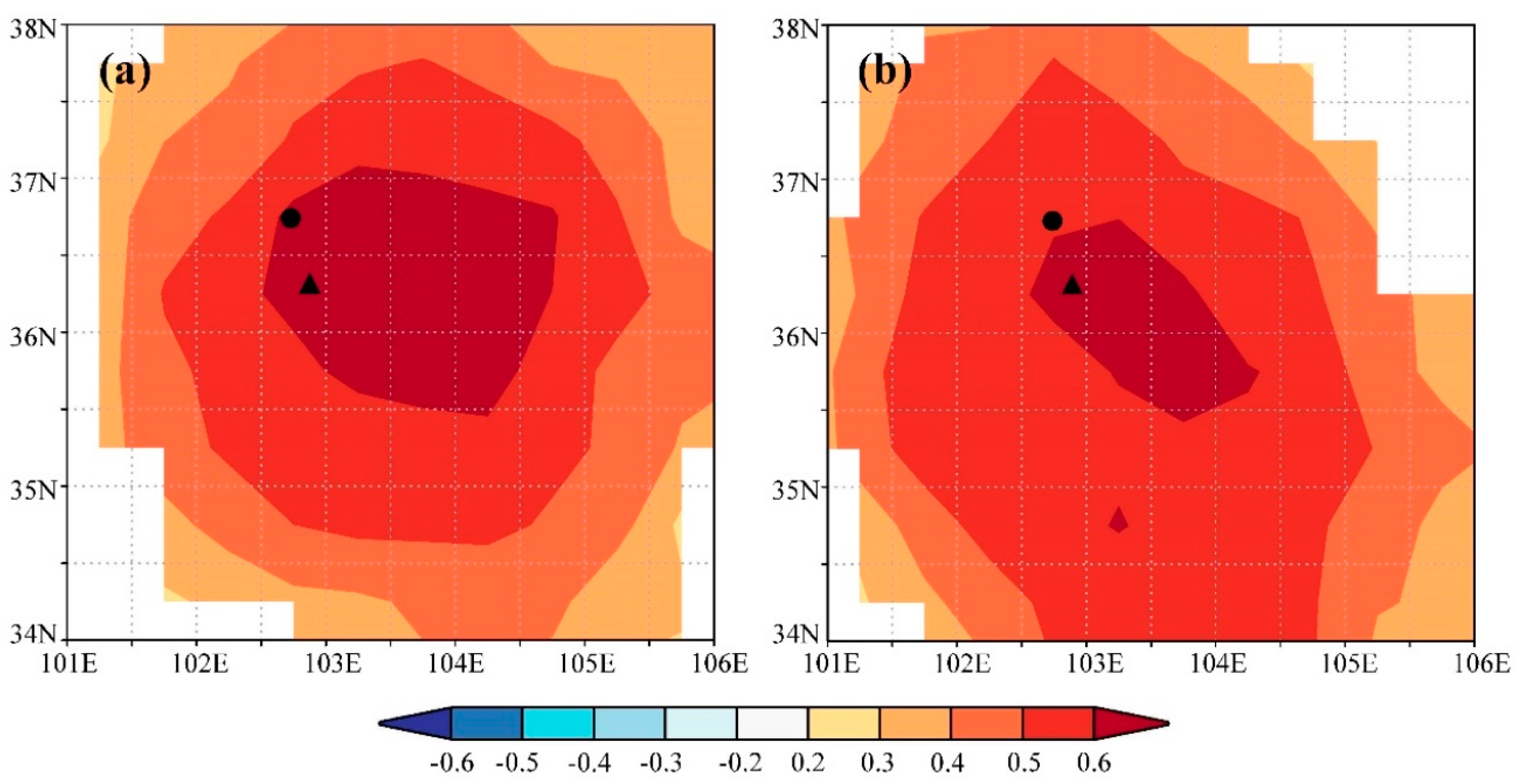
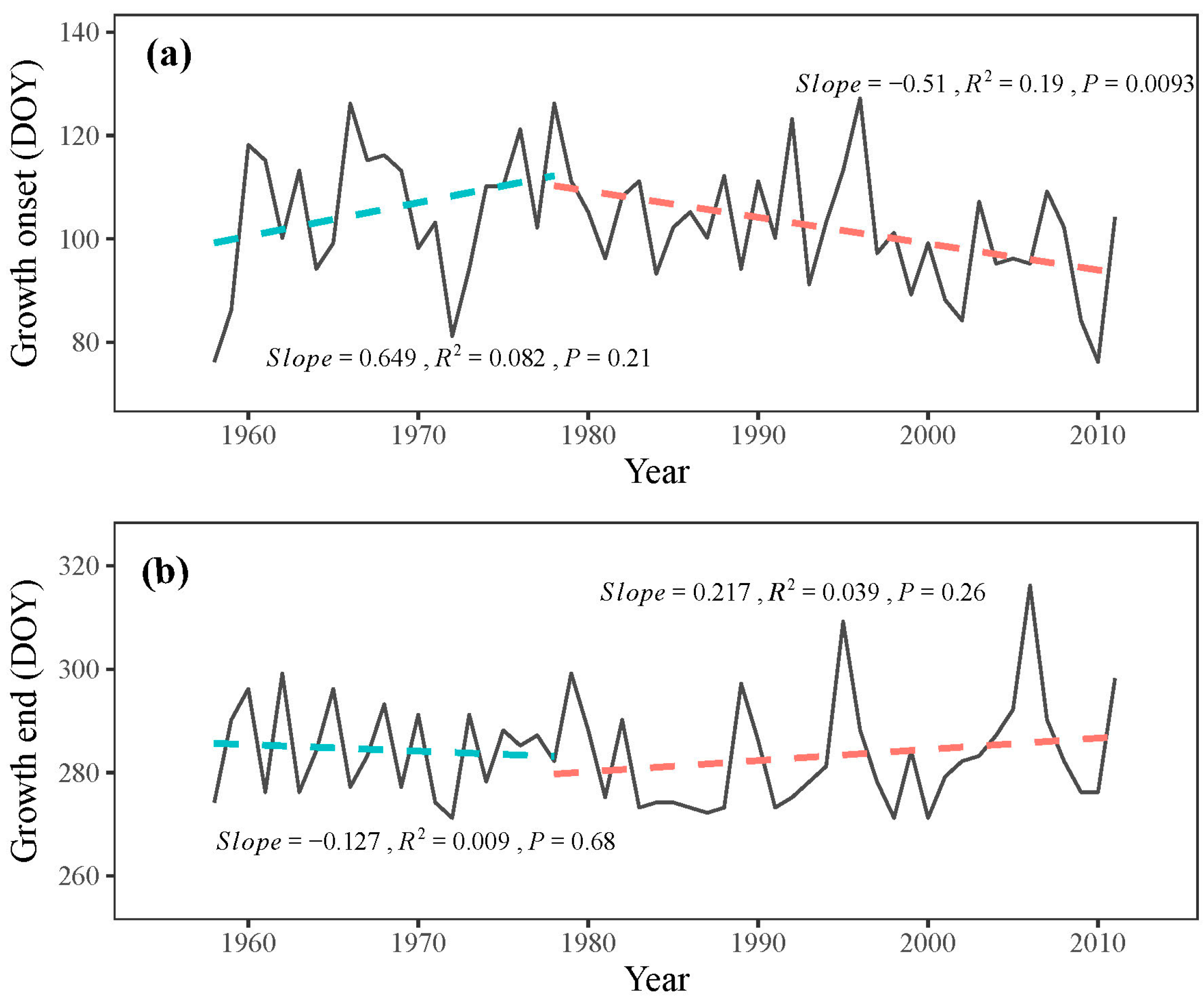
3.1. Tree-Ring and Climate Responses
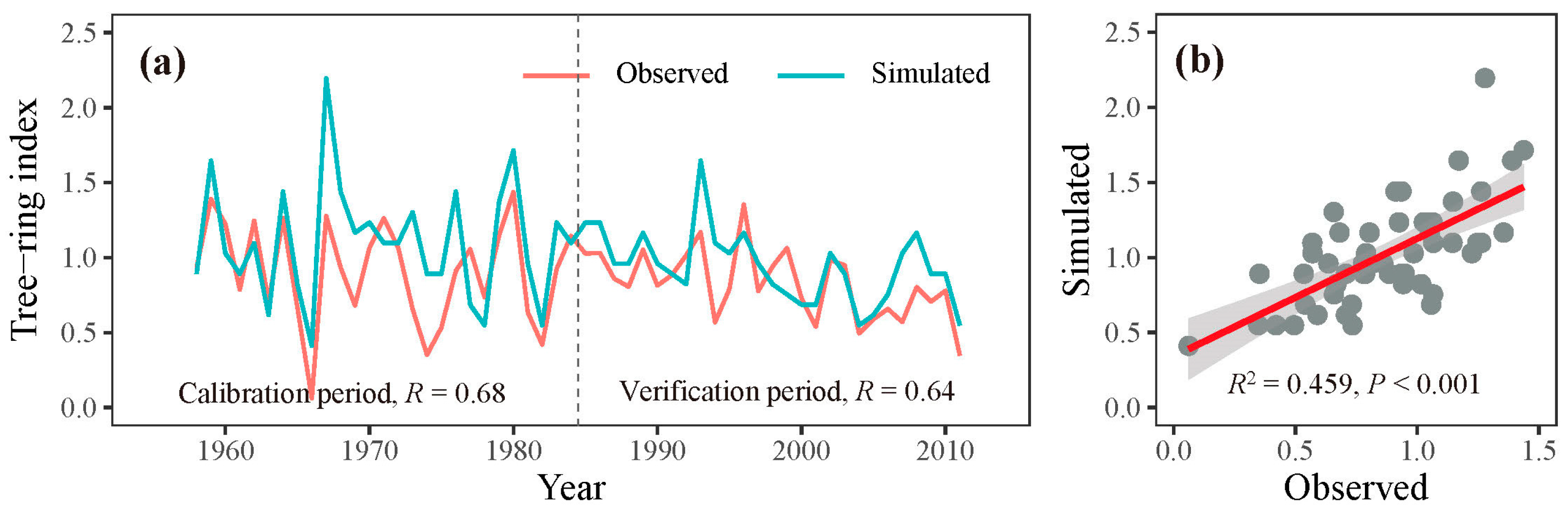
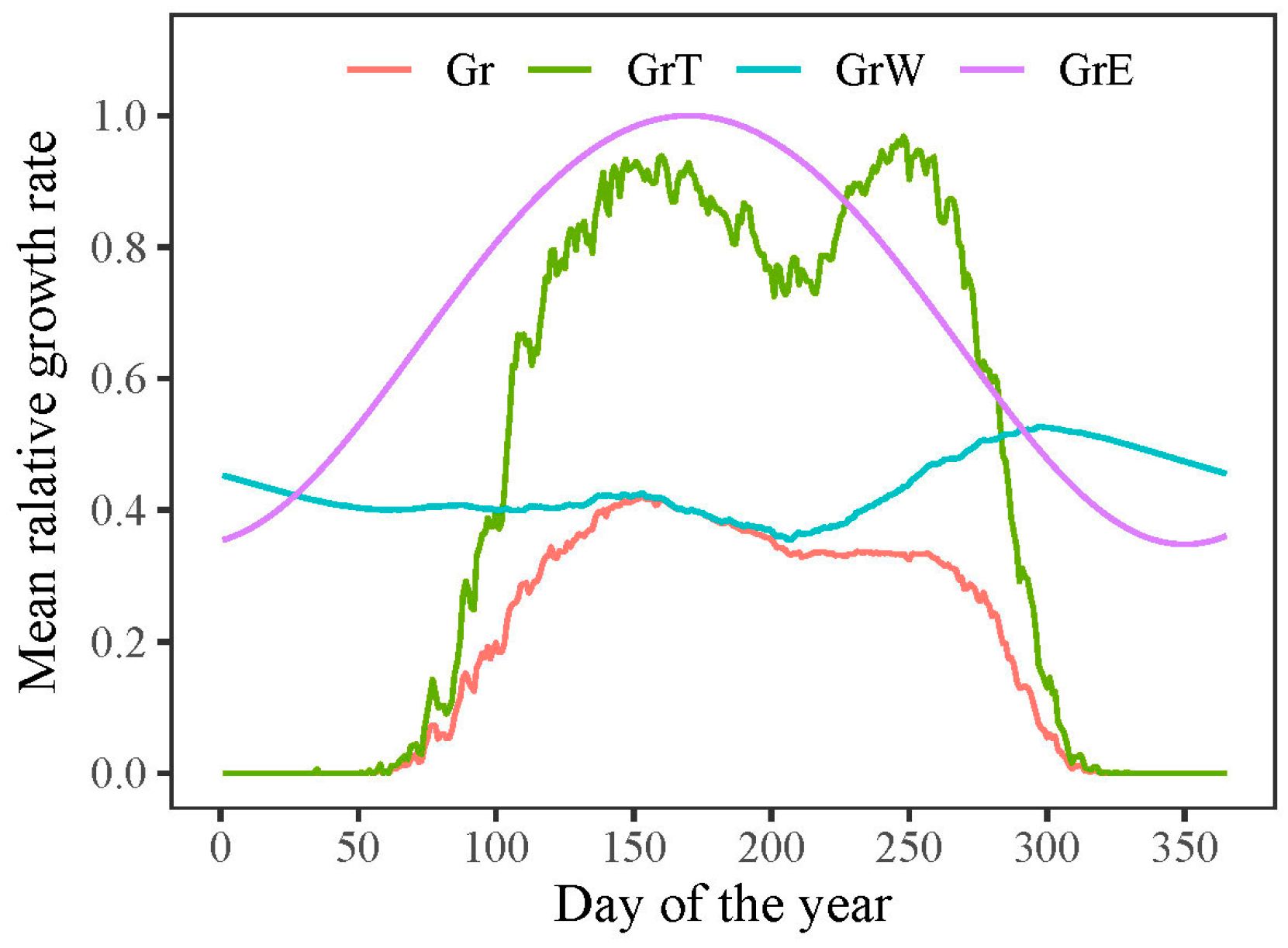
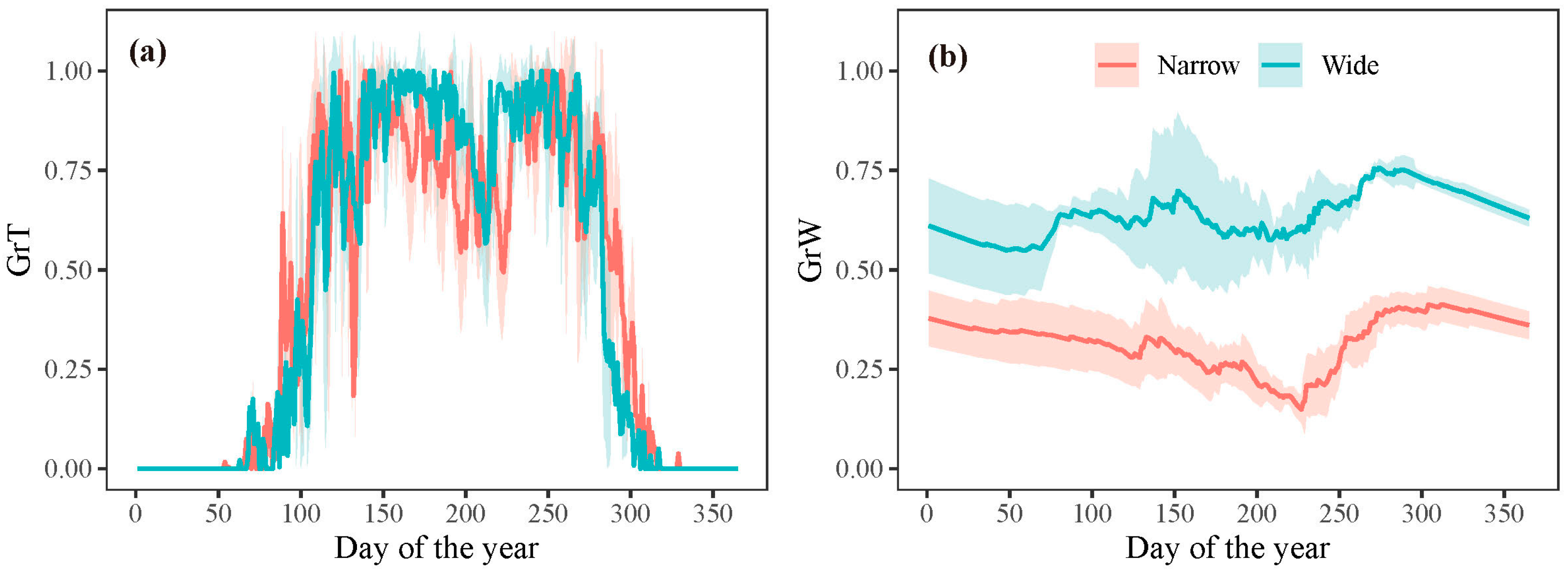
3.2. Process Model Analysis
3.3. Model Outputs of Phenology and Growth Rate of Trees
4. Discussion
4.1. Impacts of Precipitation on Tree Growth
4.2. Effects of Temperature on Phenology of Tree Growth
4.3. Influence of Water Availability on Intra-Annual Tree Growth
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Deslauriers, A.; Fonti, P.; Rossi, S.; Rathgeber, C.; Gričar, J. Ecophysiology and Plasticity of Wood and Phloem Formation. In Dendroecology, Tree-Ring Analyses Applied to Ecological Studies, AmorosoLori, M.M., Daniels, L.D., Baker, P.J., Camarero, J.J., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2017; pp. 13–33. [Google Scholar]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Pugh, T.A.M.; Rademacher, T.; Shafer, S.L.; Steinkamp, J.; Barichivich, J.; Beckage, B.; Haverd, V.; Harper, A.; Heinke, J.; Nishina, K.; et al. Understanding the uncertainty in global forest carbon turnover. Biogeosciences 2020, 17, 3961–3989. [Google Scholar] [CrossRef]
- IPCC. Summary for Policymakers. In Climate Change 2021: The Physical Science Basis; Working Group I Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Paris, France, 2021. [Google Scholar]
- Adams, H.D.; Macalady, A.K.; Breshears, D.D.; Allen, C.D.; Stephenson, N.L.; Saleska, S.R.; Huxman, T.E.; McDowell, N.G. Climate-induced tree mortality: Earth system consequences. Eos Trans. Am. Geophys. Union 2010, 91, 153–154. [Google Scholar] [CrossRef]
- Zhang, J.; Gou, X.; Manzanedo, R.D.; Zhang, F.; Pederson, N. Cambial phenology and xylogenesis of Juniperus przewalskii over a climatic gradient is influenced by both temperature and drought. Agric. For. Meteorol. 2018, 260–261, 165–175. [Google Scholar] [CrossRef]
- Fernandez-de-Una, L.; Aranda, I.; Rossi, S.; Fonti, P.; Canellas, I.; Gea-Izquierdo, G. Divergent phenological and leaf gas exchange strategies of two competing tree species drive contrasting responses to drought at their altitudinal boundary. Tree Physiol. 2018, 38, 1152–1165. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Anfodillo, T.; Čufar, K.; Cuny, H.E.; Deslauriers, A.; Fonti, P.; Frank, D.; Gricar, J.; Gruber, A.; Huang, J.G.; et al. Pattern of xylem phenology in conifers of cold ecosystems at the Northern Hemisphere. Glob. Chang. Biol. 2016, 22, 3804–3813. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Gou, X.; Pederson, N.; Zhang, F.; Niu, H.; Zhao, S.; Wang, F. Cambial phenology in Juniperus przewalskii along different altitudinal gradients in a cold and arid region. Tree Physiol. 2018, 38, 840–852. [Google Scholar] [CrossRef] [PubMed]
- Valentine, H.T.; Makela, A. Bridging process-based and empirical approaches to modeling tree growth. Tree Physiol. 2005, 25, 769–779. [Google Scholar] [CrossRef]
- Shishov, V.V.; Tychkov, I.I.; Popkova, M.I.; Ilyin, V.A.; Bryukhanova, M.V.; Kirdyanov, A.V. VS-oscilloscope: A new tool to parameterize tree radial growth based on climate conditions. Dendrochronologia 2016, 39, 42–50. [Google Scholar] [CrossRef]
- Evans, M.N.; Reichert, B.K.; Kaplan, A.; Anchukaitis, K.J.; Vaganov, E.A.; Hughes, M.K.; Cane, M.A. A forward modeling approach to paleoclimatic interpretation of tree-ring data. J. Geophys. Res.: Biogeosciences 2006, 111, G03008. [Google Scholar] [CrossRef]
- He, M.; Shishov, V.; Kaparova, N.; Yang, B.; Bräuning, A.; Grießinger, J. Process-based modeling of tree-ring formation and its relationships with climate on the Tibetan Plateau. Dendrochronologia 2017, 42, 31–41. [Google Scholar] [CrossRef]
- Yang, B.; He, M.; Shishov, V.; Tychkov, I.; Vaganov, E.; Rossi, S.; Ljungqvist, F.C.; Brauning, A.; Griessinger, J. New perspective on spring vegetation phenology and global climate change based on Tibetan Plateau tree-ring data. Proc. Natl. Acad. Sci. USA 2017, 114, 6966–6971. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Yang, B.; He, M.; Shishov, V. Intra-annual stem radial increment patterns of Chinese pine, Helan Mountains, Northern Central China. Trees 2019, 33, 751–763. [Google Scholar] [CrossRef]
- Zhang, J.; Gou, X.; Zhang, Y.; Lu, M.; Xu, X.; Zhang, F.; Liu, W.; Gao, L. Forward modeling analyses of Qilian Juniper (Sabina przewalskii) growth in response to climate factors in different regions of the Qilian Mountains, northwestern China. Trees-Struct Funct 2016, 30, 175–188. [Google Scholar] [CrossRef]
- Butto, V.; Shishov, V.; Tychkov, I.; Popkova, M.; He, M.; Rossi, S.; Deslauriers, A.; Morin, H. Comparing the Cell Dynamics of Tree-Ring Formation Observed in Microcores and as Predicted by the Vaganov-Shashkin Model. Front. Plant Sci. 2020, 11, 555055. [Google Scholar] [CrossRef] [PubMed]
- Jevšenak, J.; Tychkov, I.; Gričar, J.; Levanič, T.; Tumajer, J.; Prislan, P.; Arnič, D.; Popkova, M.; Shishov, V.V. Growth-limiting factors and climate response variability in Norway spruce (Picea abies L.) along an elevation and precipitation gradients in Slovenia. Int. J. Biometeorol. 2020, 65, 311–324. [Google Scholar] [CrossRef] [PubMed]
- Tumajer, J.; Begović, K.; Čada, V.; Jenicek, M.; Lange, J.; Mašek, J.; Kaczka, R.J.; Rydval, M.; Svoboda, M.; Vlček, L.; et al. Ecological and methodological drivers of non-stationarity in tree growth response to climate. Glob. Chang. Biol. 2022, 29, 462–476. [Google Scholar] [CrossRef] [PubMed]
- Tumajer, J.; Kašpar, J.; Kuželová, H.; Shishov, V.V.; Tychkov, I.I.; Popkova, M.I.; Vaganov, E.A.; Treml, V. Forward Modeling Reveals Multidecadal Trends in Cambial Kinetics and Phenology at Treeline. Front. Plant Sci. 2021, 12, 613643. [Google Scholar] [CrossRef] [PubMed]
- Breitenmoser, P.; Bronnimann, S.; Frank, D. Forward modelling of tree-ring width and comparison with a global network of tree-ring chronologies. Clim. Past 2014, 10, 437–449. [Google Scholar] [CrossRef]
- Touchan, R.; Shishov, V.V.; Meko, D.M.; Nouiri, I.; Grachev, A. Process based model sheds light on climate sensitivity of Mediterranean tree-ring width. Biogeosciences 2012, 9, 965–972. [Google Scholar] [CrossRef]
- Cao, Z.; Zhang, J.; Gou, X.; Wang, Y.; Sun, Q.; Yang, J.; Manzanedo, R.D.; Pederson, N. Increasing forest carbon sinks in cold and arid northeastern Tibetan Plateau. Sci. Total Environ. 2023, 905, 167168. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Gou, X.; Deng, Y.; Wang, Z.; Gu, F.; Wang, F. Increased growth of Qinghai spruce in northwestern China during the recent warming hiatus. Agric. For. Meteorol. 2018, 260-261, 9–16. [Google Scholar] [CrossRef]
- Liu, L.-Y.; Wang, X.-J.; Gou, X.-H.; Yang, M.-X.; Zhang, Z.-H. Projections of surface air temperature and precipitation in the 21st century in the Qilian Mountains, Northwest China, using REMO in the CORDEX. Adv. Clim. Chang. Res. 2022, 13, 344–358. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Y.; Li, Q.; Song, H.; Linderholm, H.W.; Leavitt, S.W.; Wang, R.; An, Z. Tree-ring stable carbon isotope-based May–July temperature reconstruction over Nanwutai, China, for the past century and its record of 20th century warming. Quat. Sci. Rev. 2014, 93, 67–76. [Google Scholar] [CrossRef]
- Chen, F.; Yuan, Y.; Zhang, R.; Qin, L. A tree-ring based drought reconstruction (AD 1760–2010) for the Loess Plateau and its possible driving mechanisms. Glob. Planet. Chang. 2014, 122, 82–88. [Google Scholar] [CrossRef]
- Zeng, Q.; Rossi, S.; Yang, B. Effects of Age and Size on Xylem Phenology in Two Conifers of Northwestern China. Front. Plant Sci. 2017, 8, 2264. [Google Scholar] [CrossRef] [PubMed]
- Wang, L. Intra-Annual Radial Growth of Three Coniferous Species and Their Responses to Climate Factors in the Eastern Qilian Mountains; Lanzhou University: Lanzhou, Chian, 2023. [Google Scholar]
- Zhang, J.; Gou, X.; Alexander, M.R.; Xia, J.; Wang, F.; Zhang, F.; Man, Z.; Pederson, N. Drought limits wood production of Juniperus przewalskii even as growing seasons lengthens in a cold and arid environment. Catena 2021, 196, 104936. [Google Scholar] [CrossRef]
- Stokes, M.; Smiley, T. An Introduction to Tree-Ring Dating; UA Press: Tucson, AZ, USA, 1968. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Cook, E.R. A Time-Series Analysis Approach to Tree-Ring Standardization; The University of Arizona: Tucson, AZ, USA, 1985. [Google Scholar]
- Vaganov, E.A.; Hughes, M.K.; Shashkin, A.V. Introduction and Factors Influencing the Seasonal Growth of Trees; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Fritts, H. Tree Rings and Climate; Academic Press: Cambridge, MA, USA, 1976. [Google Scholar]
- Liu, J.; Gou, X.; Gunina, A.; Long, X.-E.; Zhang, F.; Zhang, J. Soil nitrogen pool drives plant tissue traits in alpine treeline ecotones. For. Ecol. Manag. 2020, 477, 118490. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, F.; Gou, X.; Fonti, P.; Xia, J.; Cao, Z.; Liu, J.; Wang, Y.; Zhang, J. Seasonal variations in leaf-level photosynthesis and water use efficiency of three isohydric to anisohydric conifers on the Tibetan Plateau. Agric. For. Meteorol. 2021, 308-309, 108581. [Google Scholar] [CrossRef]
- Gao, L.L.; Gou, X.H.; Deng, Y.; Yang, M.X.; Zhao, Z.Q.; Cao, Z.Y. Dendroclimatic Response of Picea crassifolia along an Altitudinal Gradient in the Eastern Qilian Mountains, Northwest China. Arct. Antarct. Alp. Res. 2013, 45, 491–499. [Google Scholar] [CrossRef]
- Zhang, W.; Gou, X.; Liu, W.; Li, J.; Su, J.; Dilawar, N.; Zhu, F.; Xia, J.; Du, M.; Wang, L.; et al. Divergent tree radial growth patterns of Qinghai spruce (Picea crassifolia) at the alpine timberline along a moisture gradient in the Qilian mountains, Northwest China. Agric. For. Meteorol. 2023, 328, 109240. [Google Scholar] [CrossRef]
- Fang, K.Y.; Gou, X.H.; Chen, F.H.; Yang, M.X.; Li, J.B.; He, M.S.; Zhang, Y.; Tian, Q.H.; Peng, J.F. Drought variations in the eastern part of northwest China over the past two centuries: Evidence from tree rings. Clim. Res. 2009, 38, 129–135. [Google Scholar] [CrossRef]
- Gou, X.; Deng, Y.; Gao, L.; Chen, F.; Cook, E.; Yang, M.; Zhang, F. Millennium tree-ring reconstruction of drought variability in the eastern Qilian Mountains, northwest China. Clim. Dyn. 2015, 45, 1761–1770. [Google Scholar] [CrossRef]
- Zhang, J.; Alexander, M.R.; Gou, X.; Deslauriers, A.; Fonti, P.; Zhang, F.; Pederson, N. Extended xylogenesis and stem biomass production in Juniperus przewalskii Kom. during extreme late-season climatic events. Ann. For. Sci. 2020, 77, 99. [Google Scholar] [CrossRef]
- Huang, J.G.; Ma, Q.; Rossi, S.; Biondi, F.; Deslauriers, A.; Fonti, P.; Liang, E.; Makinen, H.; Oberhuber, W.; Rathgeber, C.B.K.; et al. Photoperiod and temperature as dominant environmental drivers triggering secondary growth resumption in Northern Hemisphere conifers. Proc. Natl. Acad. Sci. USA 2020, 117, 20645–20652. [Google Scholar] [CrossRef] [PubMed]
- Deslauriers, A.; Rossi, S.; Anfodillo, T.; Saracino, A. Cambial phenology, wood formation and temperature thresholds in two contrasting years at high altitude in southern Italy. Tree Physiol. 2008, 28, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Carraro, V. Evidence of threshold temperatures for xylogenesis in conifers at high altitudes. Oecologia 2007, 152, 1–12. [Google Scholar] [CrossRef]
- Rossi, S.; Girard, M.J.; Morin, H. Lengthening of the duration of xylogenesis engenders disproportionate increases in xylem production. Glob. Chang. Biol. 2014, 20, 2261–2271. [Google Scholar] [CrossRef] [PubMed]
- Ziaco, E.; Truettner, C.; Biondi, F.; Bullock, S. Moisture-driven xylogenesis in Pinus ponderosa from a Mojave Desert mountain reveals high phenological plasticity. Plant Cell Environ. 2018, 41, 823–836. [Google Scholar] [CrossRef] [PubMed]
- Balducci, L.; Cuny, H.E.; Rathgeber, C.B.; Deslauriers, A.; Giovannelli, A.; Rossi, S. Compensatory mechanisms mitigate the effect of warming and drought on wood formation. Plant Cell Environ. 2016, 39, 1338–1352. [Google Scholar] [CrossRef] [PubMed]
- Vieira, J.; Rossi, S.; Campelo, F.; Freitas, H.; Nabais, C. Xylogenesis of Pinus pinaster under a Mediterranean climate. Ann. For. Sci. 2014, 71, 71–80. [Google Scholar] [CrossRef]
- Steppe, K.; Sterck, F.; Deslauriers, A. Diel growth dynamics in tree stems: Linking anatomy and ecophysiology. Trends Plant Sci. 2015, 20, 335–343. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Yang, B.; Wang, Z.; Bräuning, A.; Pourtahmasi, K.; Oladi, R. Climatic forcing of xylem formation in Qilian juniper on the northeastern Tibetan Plateau. Trees 2016, 30, 923–933. [Google Scholar] [CrossRef]
- Ren, P.; Rossi, S.; Gricar, J.; Liang, E.; Cufar, K. Is precipitation a trigger for the onset of xylogenesis in Juniperus przewalskii on the north-eastern Tibetan Plateau? Ann. Bot. 2015, 115, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Liang, E.Y.; Shao, X.M.; Liu, X.H. Annual Precipitation Variation Inferred from Tree Rings since Ad 1770 for the Western Qilian Mts., Northern Tibetan Plateau. Tree-Ring Res. 2009, 65, 95–103. [Google Scholar] [CrossRef]
- Shao, X.; Xu, Y.; Yin, Z.Y.; Liang, E.; Zhu, H.; Wang, S. Climatic implications of a 3585-year tree-ring width chronology from the northeastern Qinghai-Tibetan Plateau. Quat. Sci. Rev. 2010, 29, 2111–2122. [Google Scholar] [CrossRef]
- Gou, X.; Zhou, F.; Zhang, Y.; Chen, Q.; Zhang, J. Forward modeling analysis of regional scale tree-ring patterns around the northeastern Tibetan Plateau, Northwest China. Biogeosciences Discuss. 2013, 10, 9969–9988. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Statistical Parameters | Value |
|---|---|
| Mean sensitivity (MS) | 0.507 |
| Inter-series correlation | 0.827 |
| Mean segment length (yr) | 84.3 |
| Absent rings (%) | 0.742 |
| All series Rbar | 0.613 |
| Standard deviation | 0.160 |
| Expressed population signal (EPS) | 0.978 |
| Signal-to-noise ratio (SNR) | 44.441 |
| Year of SSS > 0.85/yr (cores) | 1879 (4) |
| Parameter | Description (Units) | Value |
|---|---|---|
| Tmin | Minimum temperature for tree growth (°C) | 6 |
| Topt1 | Lower limit of optimal temperatures (°C) | 14 |
| Topt2 | Upper limit of optimal temperatures (°C) | 18 |
| Tmax | Maximum temperature for tree growth (°C) | 30 |
| Wmin | Minimum soil moisture for tree growth (v/v) | 0.04 |
| Wopt1 | Lower limit of optimal soil moisture (v/v) | 0.29 |
| Wopt2 | Upper limit of optimal soil moisture (v/v) | 0.8 |
| Wmax | Maximum soil moisture for tree growth (v/v) | 0.9 |
| Tbeg | Temperature sum for initiation of growth (°C), period = 10 days | 55 |
| Droot | Depth of root system (mm) | 1000 |
| Pmax | Maximum daily precipitation for saturated soil (mm) | 20 |
| k1 | Fraction of precipitation penetrating soil | 0.86 |
| k2 | First coefficient for calculation of transpiration (mm/day) | 0.12 |
| k3 | Second coefficient for calculation of transpiration (1/degree) | 0.176 |
| kr | Coefficient for water infiltration from soil | 0.002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Man, Z.; Zhang, J.; Liu, J.; Liu, L.; Yang, J.; Cao, Z. Process-Based Modeling of Phenology and Radial Growth in Pinus tabuliformis in Response to Climate Factors over a Cold and Semi-Arid Region. Plants 2024, 13, 980. https://doi.org/10.3390/plants13070980
Man Z, Zhang J, Liu J, Liu L, Yang J, Cao Z. Process-Based Modeling of Phenology and Radial Growth in Pinus tabuliformis in Response to Climate Factors over a Cold and Semi-Arid Region. Plants. 2024; 13(7):980. https://doi.org/10.3390/plants13070980
Chicago/Turabian StyleMan, Zihong, Junzhou Zhang, Junjun Liu, Li Liu, Jiqin Yang, and Zongying Cao. 2024. "Process-Based Modeling of Phenology and Radial Growth in Pinus tabuliformis in Response to Climate Factors over a Cold and Semi-Arid Region" Plants 13, no. 7: 980. https://doi.org/10.3390/plants13070980
APA StyleMan, Z., Zhang, J., Liu, J., Liu, L., Yang, J., & Cao, Z. (2024). Process-Based Modeling of Phenology and Radial Growth in Pinus tabuliformis in Response to Climate Factors over a Cold and Semi-Arid Region. Plants, 13(7), 980. https://doi.org/10.3390/plants13070980