


Genome-Wide Investigation of Class III Peroxidase Genes in Brassica napus Reveals Their Responsiveness to Abiotic Stresses

 ,
,  , and
, and 
Abstract
1. Introduction
2. Results
2.1. Characterization of BnPOD Gene Family
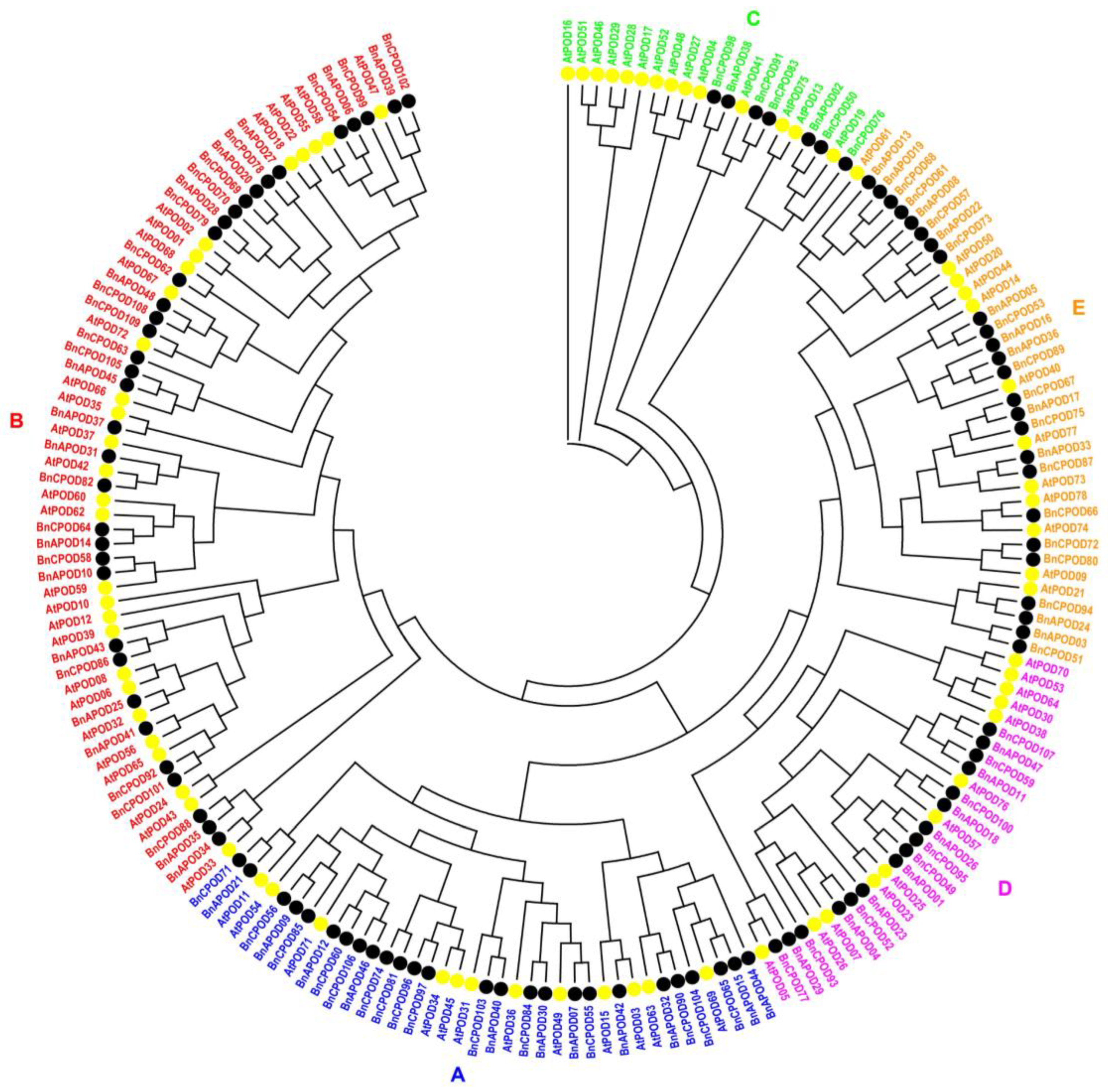
2.2. Phylogeny of POD Genes in B. napus
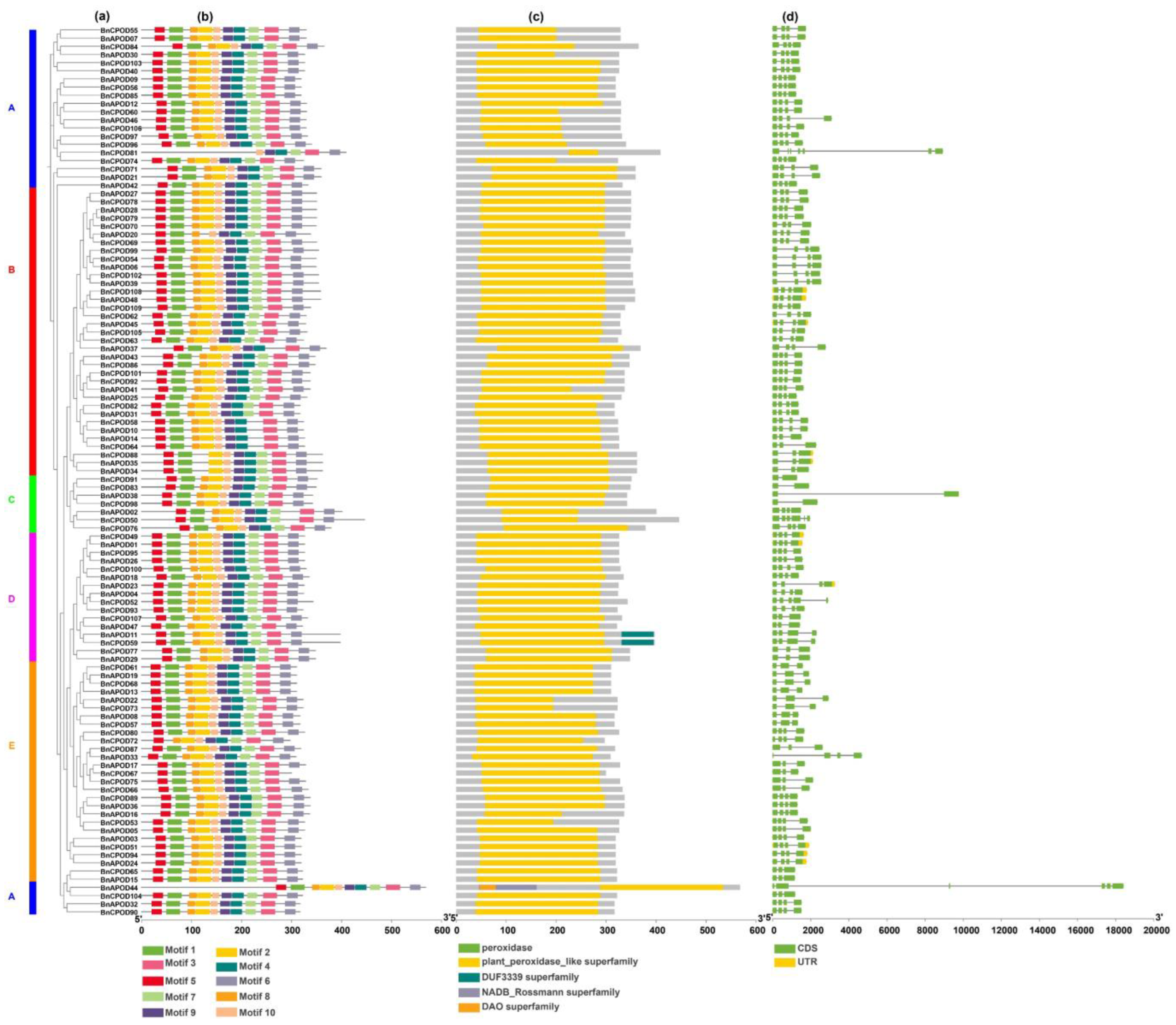
2.3. Gene Structural Analysis of BnPOD Proteins
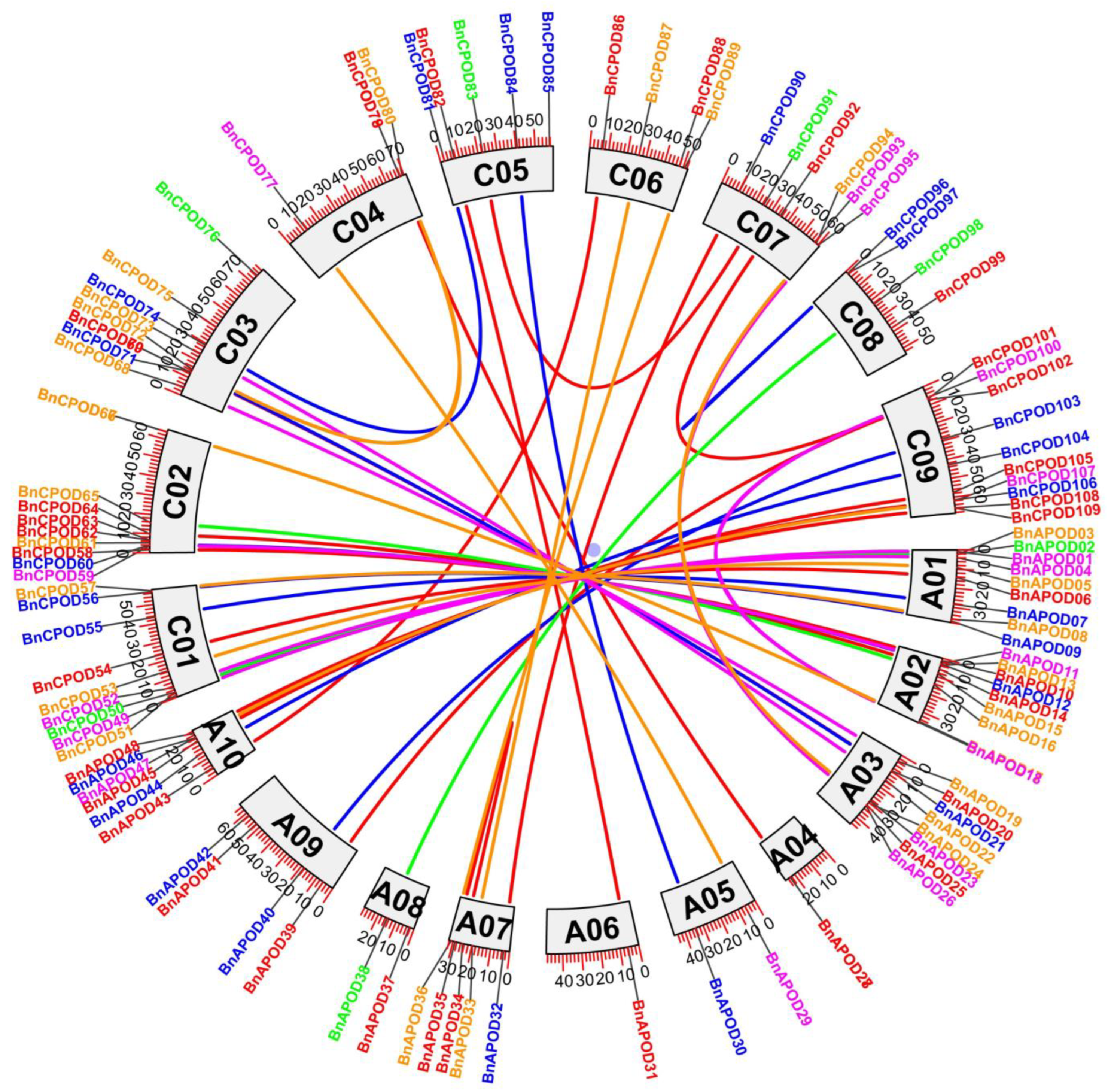
2.4. Chromosomal Distribution and Duplication Events of POD Genes
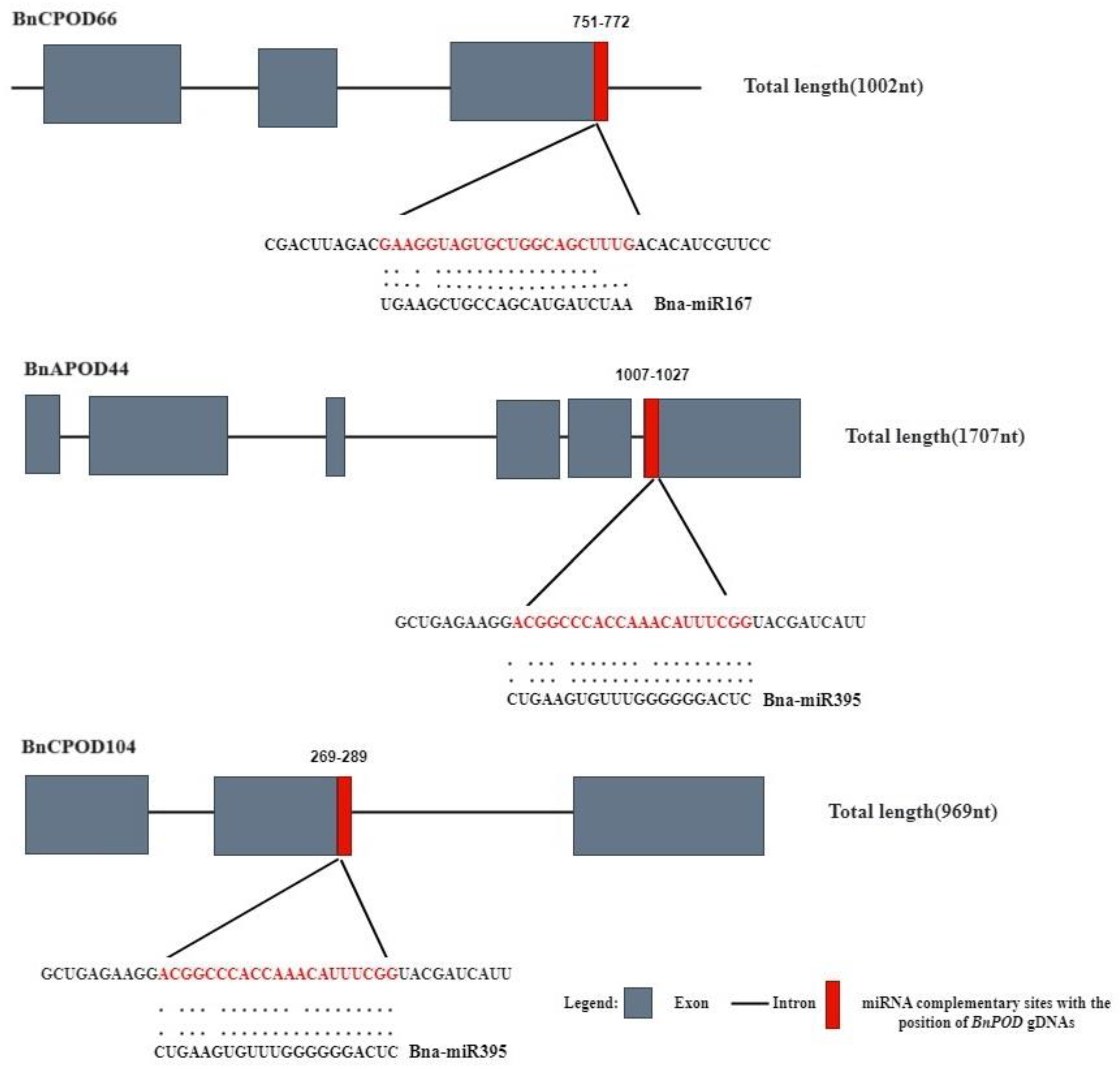
2.5. miRNA-Mediated BnPOD Regulation
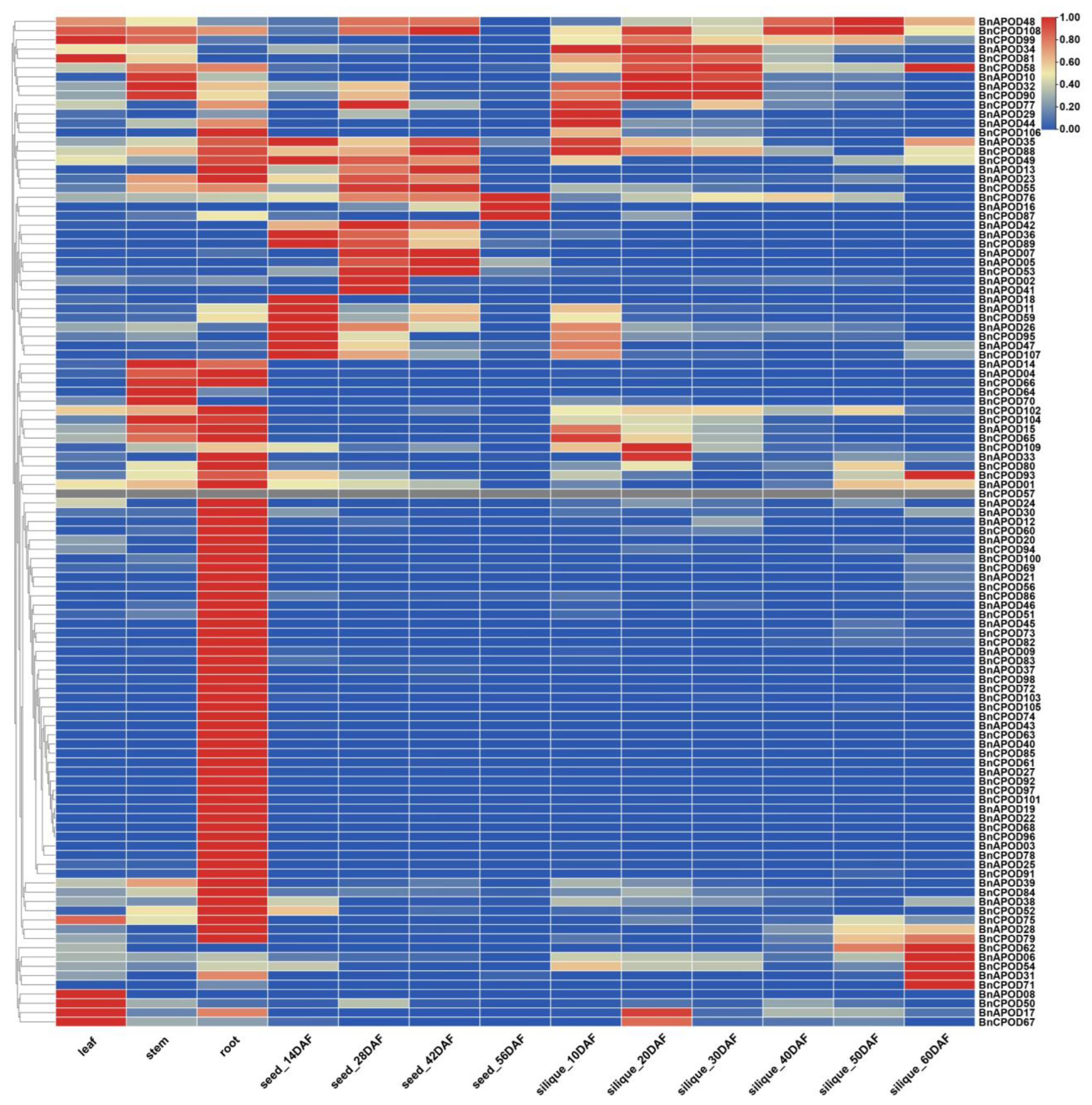
2.6. Expression Profiling of BnPOD Genes in Various Tissues
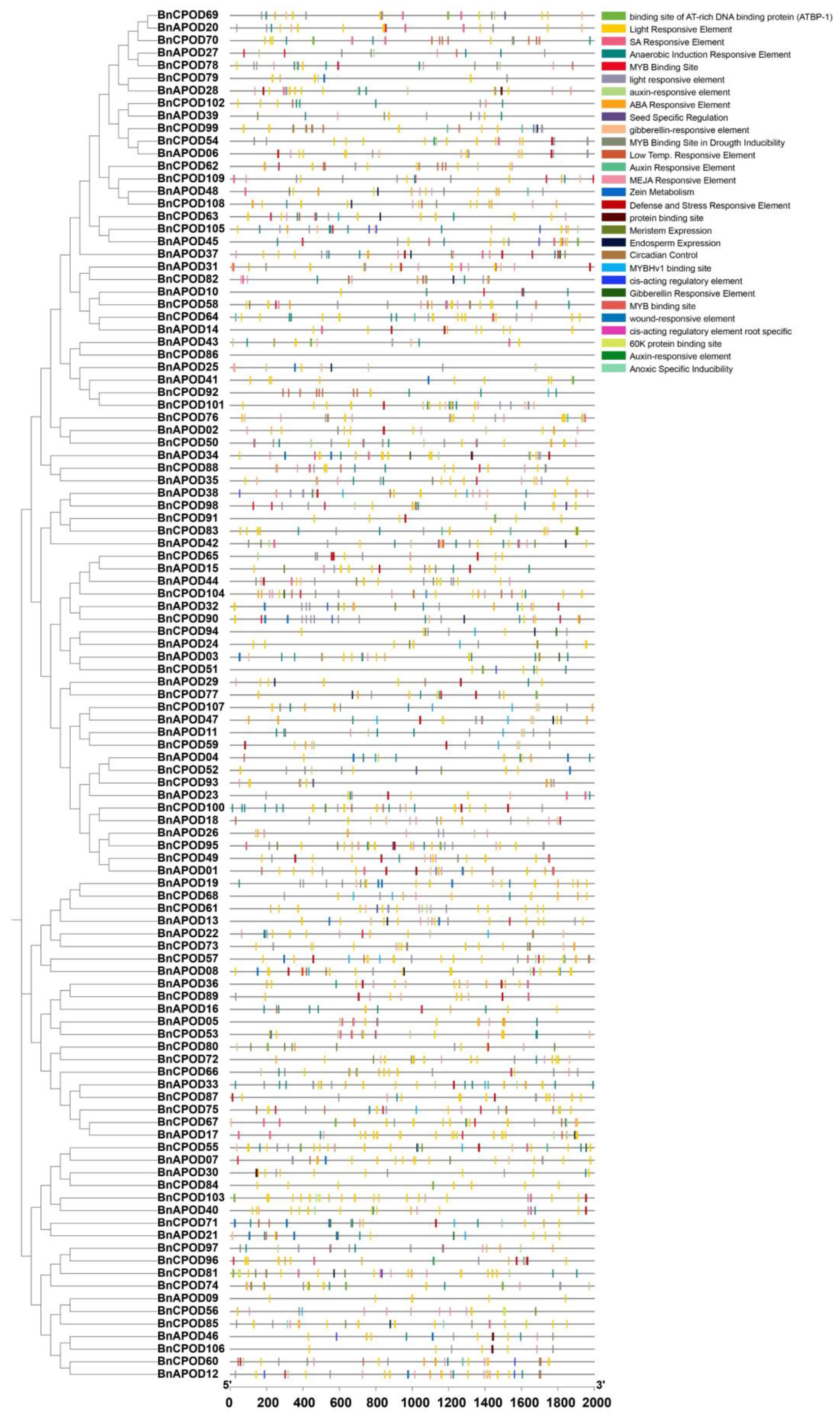
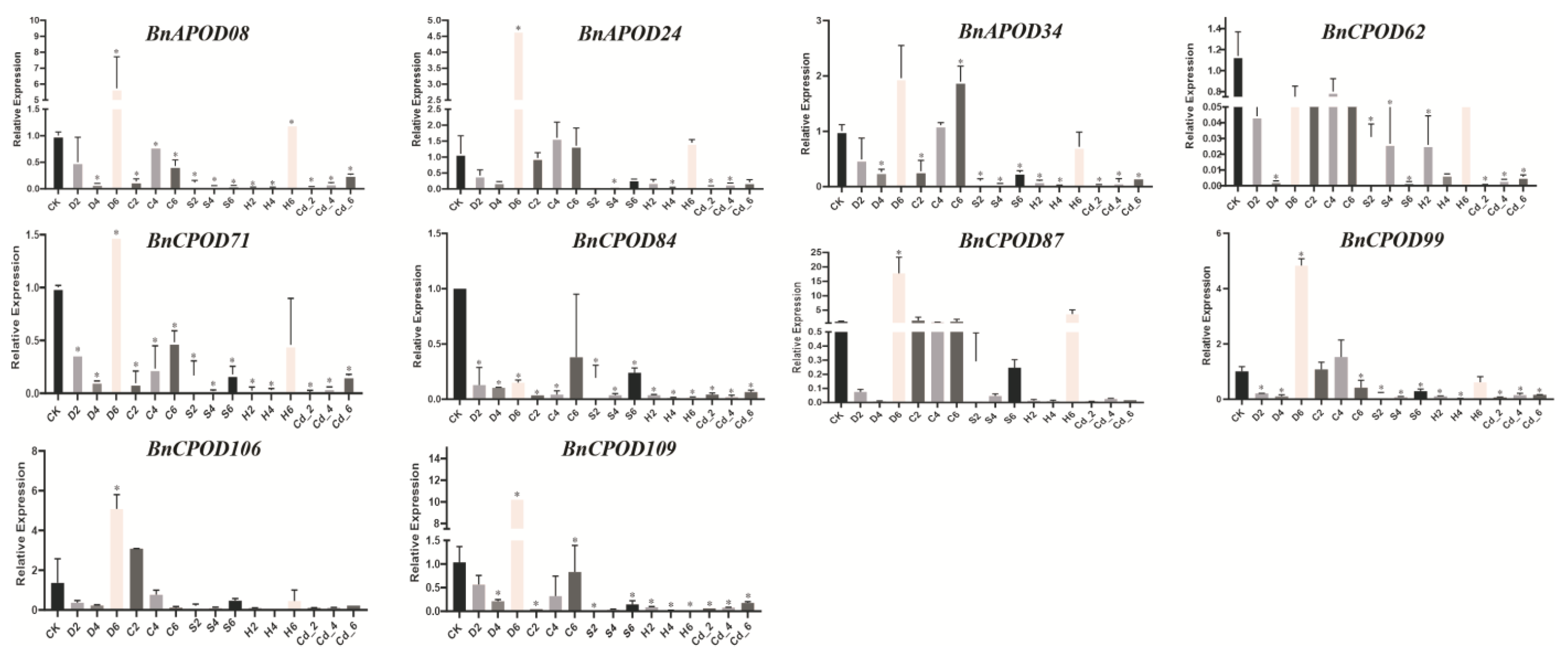
2.7. Expression Patterns of BnPODs during Abiotic Stresses
3. Discussion
4. Materials and Methods
4.1. Mining of BnPOD Family in B. napus
4.2. Phylogeny of B. napus POD Proteins
4.3. Gene Structural Analysis of B. napus POD Proteins
4.4. Gene Duplication and Evolutionary Analysis of BnPOD
4.5. Prediction of Putative miRNAs Targeting BnPOD Genes
4.6. Expression Analysis of BnPOD Genes in Different Tissues
4.7. Plant Materials and Abiotic Stresses
4.8. RNA Isolation and Real-Time Quantitative PCR Expression Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hiraga, S.; Sasaki, K.; Ito, H.; Ohashi, Y.; Matsui, H. A Large Family of Class III Plant Peroxidases. Plant Cell Physiol. 2001, 42, 462–468. [Google Scholar] [CrossRef]
- Hiraga, S.; Ichinose, C.; Onogi, T.; Niki, H.; Yamazoe, M. Bidirectional Migration of SeqA-Bound Hemimethylated DNA Clusters and Pairing of oriC Copies in Escherichia coli: Migration of SeqA-DNA Complex and oriC. Genes Cells 2000, 5, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Koua, D.; Cerutti, L.; Falquet, L.; Sigrist, C.J.A.; Theiler, G.; Hulo, N.; Dunand, C. PeroxiBase: A Database with New Tools for Peroxidase Family Classification. Nucleic Acids Res. 2009, 37, D261–D266. [Google Scholar] [CrossRef]
- Zámocký, M.; Furtmüller, P.G.; Obinger, C. Evolution of Structure and Function of Class I Peroxidases. Arch. Biochem. Biophys. 2010, 500, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Piontek, K.; Smith, A.T.; Blodig, W. Lignin Peroxidase Structure and Function. Biochem. Soc. Trans. 2001, 29, 111–116. [Google Scholar] [CrossRef]
- Mathé, C.; Barre, A.; Jourda, C.; Dunand, C. Evolution and Expression of Class III Peroxidases. Arch. Biochem. Biophys. 2010, 500, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Passardi, F.; Longet, D.; Penel, C.; Dunand, C. The Class III Peroxidase Multigenic Family in Rice and Its Evolution in Land Plants☆,☆☆. Phytochemistry 2004, 65, 1879–1893. [Google Scholar] [CrossRef] [PubMed]
- Herrero, J.; Fernández-Pérez, F.; Yebra, T.; Novo-Uzal, E.; Pomar, F.; Pedreño, M.Á.; Cuello, J.; Guéra, A.; Esteban-Carrasco, A.; Zapata, J.M. Bioinformatic and Functional Characterization of the Basic Peroxidase 72 from Arabidopsis Thaliana Involved in Lignin Biosynthesis. Planta 2013, 237, 1599–1612. [Google Scholar] [CrossRef]
- Pandey, V.P.; Awasthi, M.; Singh, S.; Tiwari, S.; Dwivedi, U.N. A Comprehensive Review on Function and Application of Plant Peroxidases. Biochem. Anal. Biochem. 2017, 6, 308. [Google Scholar] [CrossRef]
- Passardi, F.; Tognolli, M.; De Meyer, M.; Penel, C.; Dunand, C. Two Cell Wall Associated Peroxidases from Arabidopsis Influence Root Elongation. Planta 2006, 223, 965–974. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, Z.; How, J.; Xu, H.; Chen, L.; Li, K. Overexpression of a Peroxidase Gene (AtPrx64) of Arabidopsis Thaliana in Tobacco Improves Plant’s Tolerance to Aluminum Stress. Plant Mol. Biol. 2017, 95, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Llorente, F.; López-Cobollo, R.M.; Catalá, R.; Martínez-Zapater, J.M.; Salinas, J. A Novel Cold-inducible Gene from Arabidopsis, RCI3, Encodes a Peroxidase That Constitutes a Component for Stress Tolerance. Plant J. 2002, 32, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Jaggi, M.; Kumar, S.; Sinha, A.K. Overexpression of an Apoplastic Peroxidase Gene CrPrx in Transgenic Hairy Root Lines of Catharanthus Roseus. Appl. Microbiol. Biotechnol. 2011, 90, 1005–1016. [Google Scholar] [CrossRef]
- Su, P.; Yan, J.; Li, W.; Wang, L.; Zhao, J.; Ma, X.; Li, A.; Wang, H.; Kong, L. A Member of Wheat Class III Peroxidase Gene Family, TaPRX-2A, Enhanced the Tolerance of Salt Stress. BMC Plant Biol. 2020, 20, 392. [Google Scholar] [CrossRef]
- Sečenji, M.; Lendvai, Á.; Miskolczi, P.; Kocsy, G.; Gallé, Á.; Szűcs, A.; Hoffmann, B.; Sárvári, É.; Schweizer, P.; Stein, N.; et al. Differences in Root Functions during Long-Term Drought Adaptation: Comparison of Active Gene Sets of Two Wheat Genotypes: Adaptation of Wheat Root Genes to Drought. Plant Biol. 2010, 12, 871–882. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Q.; Zhao, Y.; Han, G.; Zhu, S. Systematic Analysis of Maize Class III Peroxidase Gene Family Reveals a Conserved Subfamily Involved in Abiotic Stress Response. Gene 2015, 566, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Tognolli, M.; Penel, C.; Greppin, H.; Simon, P. Analysis and Expression of the Class III Peroxidase Large Gene Family in Arabidopsis Thaliana. Gene 2002, 288, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Ding, X.; Ding, Z.; Tie, W.; Yan, Y.; Wang, Y.; Yang, H.; Hu, W. The Class III Peroxidase (POD) Gene Family in Cassava: Identification, Phylogeny, Duplication, and Expression. Int. J. Mol. Sci. 2019, 20, 2730. [Google Scholar] [CrossRef]
- Cao, Y.; Han, Y.; Meng, D.; Li, D.; Jin, Q.; Lin, Y.; Cai, Y. Structural, Evolutionary, and Functional Analysis of the Class III Peroxidase Gene Family in Chinese Pear (Pyrus bretschneideri). Front. Plant Sci. 2016, 7, 1874. [Google Scholar] [CrossRef]
- Li, Q.; Dou, W.; Qi, J.; Qin, X.; Chen, S.; He, Y. Genome-wide Analysis of the CIII Peroxidase Family in Sweet Orange (Citrus Sinensis) and Expression Profiles Induced by Xanthomonas Citri Subsp. Citri and Hormones. J. Genet. 2020, 99, 10. [Google Scholar] [CrossRef]
- Xiao, H.; Wang, C.; Khan, N.; Chen, M.; Fu, W.; Guan, L.; Leng, X. Genome-Wide Identification of the Class III POD Gene Family and Their Expression Profiling in Grapevine (Vitis vinifera L). BMC Genom. 2020, 21, 444. [Google Scholar] [CrossRef] [PubMed]
- Aleem, M.; Riaz, A.; Raza, Q.; Aleem, M.; Aslam, M.; Kong, K.; Atif, R.M.; Kashif, M.; Bhat, J.A.; Zhao, T. Genome-Wide Characterization and Functional Analysis of Class III Peroxidase Gene Family in Soybean Reveal Regulatory Roles of GsPOD40 in Drought Tolerance. Genomics 2022, 114, 45–60. [Google Scholar] [CrossRef] [PubMed]
- González-Gordo, S.; Muñoz-Vargas, M.A.; Palma, J.M.; Corpas, F.J. Class III Peroxidases (POD) in Pepper (Capsicum annuum L.): Genome-Wide Identification and Regulation during Nitric Oxide (NO)-Influenced Fruit Ripening. Antioxidants 2023, 12, 1013. [Google Scholar] [CrossRef] [PubMed]
- Almagro, L.; Gómez Ros, L.V.; Belchi-Navarro, S.; Bru, R.; Ros Barceló, A.; Pedreño, M.A. Class III Peroxidases in Plant Defence Reactions. J. Exp. Bot. 2009, 60, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Ayadi, J.; Debouba, M.; Rahmani, R.; Bouajila, J. Brassica Genus Seeds: A Review on Phytochemical Screening and Pharmacological Properties. Molecules 2022, 27, 6008. [Google Scholar] [CrossRef]
- Lohani, N.; Jain, D.; Singh, M.B.; Bhalla, P.L. Engineering Multiple Abiotic Stress Tolerance in Canola, Brassica napus. Front. Plant Sci. 2020, 11, 3. [Google Scholar] [CrossRef]
- Song, J.-M.; Guan, Z.; Hu, J.; Guo, C.; Yang, Z.; Wang, S.; Liu, D.; Wang, B.; Lu, S.; Zhou, R.; et al. Eight High-Quality Genomes Reveal Pan-Genome Architecture and Ecotype Differentiation of Brassica napus. Nat. Plants 2020, 6, 34–45. [Google Scholar] [CrossRef]
- Mehmood, S.S.; Lu, G.; Luo, D.; Hussain, M.A.; Raza, A.; Zafar, Z.; Zhang, X.; Cheng, Y.; Zou, X.; Lv, Y. Integrated Analysis of Transcriptomics and Proteomics Provides Insights into the Molecular Regulation of Cold Response in Brassica napus. Environ. Exp. Bot. 2021, 187, 104480. [Google Scholar] [CrossRef]
- Shang, H.; Fang, L.; Qin, L.; Jiang, H.; Duan, Z.; Zhang, H.; Yang, Z.; Cheng, G.; Bao, Y.; Xu, J.; et al. Genome-Wide Identification of the Class III Peroxidase Gene Family of Sugarcane and Its Expression Profiles under Stresses. Front. Plant Sci. 2023, 14, 1101665. [Google Scholar] [CrossRef]
- Cheng, L.; Ma, L.; Meng, L.; Shang, H.; Cao, P.; Jin, J. Genome-Wide Identification and Analysis of the Class III Peroxidase Gene Family in Tobacco (Nicotiana Tabacum). Front. Genet. 2022, 13, 916867. [Google Scholar] [CrossRef]
- Cai, K.; Liu, H.; Chen, S.; Liu, Y.; Zhao, X.; Chen, S. Genome-Wide Identification and Analysis of Class III Peroxidases in Betula Pendula. BMC Genom. 2021, 22, 314. [Google Scholar] [CrossRef]
- Ijaz, U. Plant Cis-Regulatory Elements: Methods of Identification and Applications. Asian J. Agric. Biol. 2020, 8, 207–222. [Google Scholar] [CrossRef]
- Khan, W.U.; Khan, L.U.; Chen, D.; Chen, F. Comparative Analyses of Superoxide Dismutase (SOD) Gene Family and Expression Profiling under Multiple Abiotic Stresses in Water Lilies. Horticulturae 2023, 9, 781. [Google Scholar] [CrossRef]
- Raza, A.; Su, W.; Gao, A.; Mehmood, S.S.; Hussain, M.A.; Nie, W.; Lv, Y.; Zou, X.; Zhang, X. Catalase (CAT) Gene Family in Rapeseed (Brassica napus L.): Genome-Wide Analysis, Identification, and Expression Pattern in Response to Multiple Hormones and Abiotic Stress Conditions. Int. J. Mol. Sci. 2021, 22, 4281. [Google Scholar] [CrossRef]
- Zaman, Q.U.; Hussain, M.A.; Khan, L.U.; Hui, L.; Khan, D.; Khokhar, A.A.; Cui, J.; Raza, A.; Wang, H.-F. Genome-Wide Identification and Expression Profiling of APX Gene Family under Multifactorial Stress Combinations and Melatonin-Mediated Tolerance in Pitaya. Sci. Hortic. 2023, 321, 112312. [Google Scholar] [CrossRef]
- Kumar, S.; Jaggi, M.; Sinha, A.K. Ectopic Overexpression of Vacuolar and Apoplastic Catharanthus Roseus Peroxidases Confers Differential Tolerance to Salt and Dehydration Stress in Transgenic Tobacco. Protoplasma 2012, 249, 423–432. [Google Scholar] [CrossRef]
- Kim, B.H.; Kim, S.Y.; Nam, K.H. Genes Encoding Plant-Specific Class III Peroxidases Are Responsible for Increased Cold Tolerance of the Brassinosteroid-Insensitive 1 Mutant. Mol. Cells 2012, 34, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.-L.; Liu, Y.-J.; Liu, H.-J.; Qian, T.-T.; Qi, L.-W.; Wang, X.-R.; Zeng, Q.-Y. Subcellular Relocalization and Positive Selection Play Key Roles in the Retention of Duplicate Genes of Populus Class III Peroxidase Family. Plant Cell 2014, 26, 2404–2419. [Google Scholar] [CrossRef]
- Li, X.; Ma, W.; Jiang, Y. Expression Patterns of Heat Shock Protein Genes and Antioxidase Genes in Apis cerana cerana (Hymenoptera: Apidae) under Heat Stress. J. Entomol. Sci. 2023, 58, 95–103. [Google Scholar] [CrossRef]
- Rhee, S.Y. The Arabidopsis Information Resource (TAIR): A Model Organism Database Providing a Centralized, Curated Gateway to Arabidopsis Biology, Research Materials and Community. Nucleic Acids Res. 2003, 31, 224–228. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein Localization Predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent Updates and New Developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for Motif Discovery and Searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Lescot, M. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Waseem, M.; Aslam, M.M.; Shaheen, I. The DUF221 Domain-Containing (DDP) Genes Identification and Expression Analysis in Tomato under Abiotic and Phytohormone Stress. GM Crops Food 2021, 12, 586–599. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.M.; Fritschi, F.B.; Di, Z.; Wang, G.; Li, H.; Lam, H.; Waseem, M.; Weifeng, X.; Zhang, J. Overexpres-sion of LaGRAS Enhances Phosphorus Acquisition via Increased Root Growth of Phosphorus-deficient White Lupin. Physiol. Plant. 2023, 175, e13962. [Google Scholar] [CrossRef]
- Koch, M.A.; Haubold, B.; Mitchell-Olds, T. Comparative Evolutionary Analysis of Chalcone Synthase and Al-cohol Dehydrogenase Loci in Arabidopsis, Arabis, and Related Genera (Brassicaceae). Mol. Biol. Evol. 2000, 17, 1483–1498. [Google Scholar] [CrossRef]
- Dai, X.; Zhao, P.X. psRNATarget: A Plant Small RNA Target Analysis Server. Nucleic Acids Res. 2011, 39, W155–W159. [Google Scholar] [CrossRef]
- Franz, M.; Lopes, C.T.; Huck, G.; Dong, Y.; Sumer, O.; Bader, G.D. Cytoscape.Js: A Graph Theory Library for Visualisation and Analysis. Bioinformatics 2016, 32, 309–311. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Liu, D.; Xie, W.; Yang, Z.; Guo, L.; Liu, K.; Yang, Q.; Chen, L. BnPIR: Brassica napus Pan-genome Information Resource for 1689 Accessions. Plant Biotechnol. J. 2021, 19, 412–414. [Google Scholar] [CrossRef] [PubMed]
- Hammer, P.A.; Tibbitts, T.W.; Langhans, R.W.; McFarlane, J.C. Base-Line Growth Studies of ‘Grand Rapids’ Lettuce in Controlled Environments1. J. Am. Soc. Hort. Sci. 1978, 103, 649–655. [Google Scholar] [CrossRef]
- Huang, Z.; Fasco, M.J.; Kaminsky, L.S. Optimization of DNase I Removal of Contaminating DNA from RNA for Use in Quantitative RNA-PCR. BioTechniques 1996, 20, 1012–1020. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Motif | Sequence | InterProScan Annotation |
|---|---|---|
| Motif-1 | DPRJAAALLRLHFHDCFVNGCDASVLLDS | Heme peroxidases |
| Motif-2 | AACPGVVSCADILALAARDSVVLAGGPSW | Heme peroxidases |
| Motif-3 | DPGTPNTFDNSYFKNLRQGKGLLQSDQAL | Heme peroxidases |
| Motif-4 | KDLVALSGAHTIGFAHCGSFTBRL | Heme peroxidases |
| Motif-5 | AQLSPGFYDKSCPNAESIVRN | - |
| Motif-6 | FFRAFAKAMVKMGNIGVLTGSQ | Heme peroxidases |
| Motif-7 | GDPDPTLBPTYAAQLRKKCPR | Heme peroxidases |
| Motif-8 | SLRGFEVIDDIKAALE | - |
| Motif-9 | PSPFDNVSQLITKFAAKGLNV | Heme peroxidases |
| Motif-10 | VPLGRRDGRVSNASE | Heme peroxidases |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, O.U.; Khan, L.U.; Basharat, S.; Zhou, L.; Ikram, M.; Peng, J.; Khan, W.U.; Liu, P.; Waseem, M. Genome-Wide Investigation of Class III Peroxidase Genes in Brassica napus Reveals Their Responsiveness to Abiotic Stresses. Plants 2024, 13, 942. https://doi.org/10.3390/plants13070942
Shah OU, Khan LU, Basharat S, Zhou L, Ikram M, Peng J, Khan WU, Liu P, Waseem M. Genome-Wide Investigation of Class III Peroxidase Genes in Brassica napus Reveals Their Responsiveness to Abiotic Stresses. Plants. 2024; 13(7):942. https://doi.org/10.3390/plants13070942
Chicago/Turabian StyleShah, Obaid Ullah, Latif Ullah Khan, Sana Basharat, Lingling Zhou, Muhammad Ikram, Jiantao Peng, Wasi Ullah Khan, Pingwu Liu, and Muhammad Waseem. 2024. "Genome-Wide Investigation of Class III Peroxidase Genes in Brassica napus Reveals Their Responsiveness to Abiotic Stresses" Plants 13, no. 7: 942. https://doi.org/10.3390/plants13070942
APA StyleShah, O. U., Khan, L. U., Basharat, S., Zhou, L., Ikram, M., Peng, J., Khan, W. U., Liu, P., & Waseem, M. (2024). Genome-Wide Investigation of Class III Peroxidase Genes in Brassica napus Reveals Their Responsiveness to Abiotic Stresses. Plants, 13(7), 942. https://doi.org/10.3390/plants13070942