





Assessment of Fertility Dynamics and Nutritional Quality of Potato Tubers in a Compost-Amended Mars Regolith Simulant

 , ,
, ,  ,
, 
 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Results
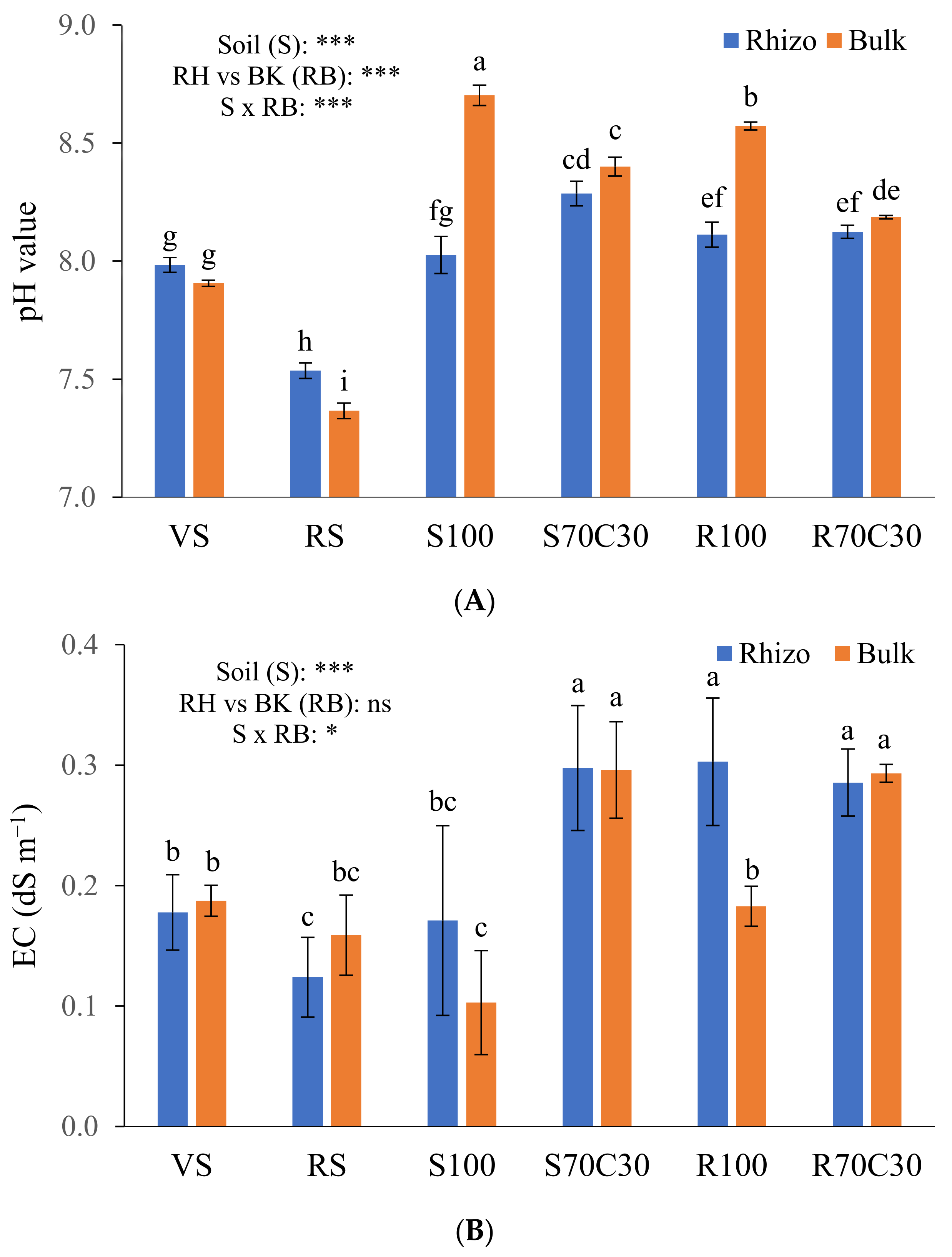
2.1. Physico-Hydraulic and Chemical Indicators of Substrate Fertility
2.2. Plant Growth
2.3. Tuber Quality
3. Discussion
4. Materials and Methods
4.1. Plant Cultivation
4.2. Sampling and Measurements
4.2.1. Characterization of the Substrates
Physical and Hydrological Properties
Chemical Properties
4.2.2. Plant Growth and Tuber Yield
4.2.3. Nutritional Quality Assessment of Potato Tubers
Elemental Analysis
Chemicals and Reagents
Glycoalkaloids Extraction
Liquid Chromatography Q-Orbitrap Mass Spectrometry Analysis
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Häder, D.-P.; Braun, M.; Hemmersbach, R. Bioregenerative Life Support Systems in Space Research. In Gravitational Biology I: Gravity Sensing and Graviorientation in Microorganisms and Plants; Braun, M., Böhmer, M., Häder, D.-P., Hemmersbach, R., Palme, K., Eds.; SpringerBriefs in Space Life Sciences; Springer International Publishing: Cham, Switzerland, 2018; pp. 113–122. [Google Scholar] [CrossRef]
- Wheeler, R.M. Plants for Human Life Support in Space: From Myers to Mars. Gravitational Space Biol. 2010, 23, 25–36. [Google Scholar]
- Hendrickx, L.; Mergeay, M. From the Deep Sea to the Stars: Human Life Support through Minimal Communities. Curr. Opin. Microbiol. 2007, 10, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Lasseur, C.; Brunet, J.; de Weever, H.; Dixon, M.; Dussap, G.; Godia, F.; Leys, N.; Mergeay, M.; Straeten, D.V.D. MELiSSA: The European project of closed life support system. Gravitational Space Res. 2010, 23, 3–12. [Google Scholar]
- Duri, L.; Caporale, A.; Rouphael, Y.; Vingiani, S.; Palladino, M.; Pascale, S.; Adamo, P. The Potential for Lunar and Martian Regolith Simulants to Sustain Plant Growth: A Multidisciplinary Overview. Front. Astron. Space Sci. 2022, 8, 747821. [Google Scholar] [CrossRef]
- Hoff, J.E.; Howe, J.M.; Mitchell, C.A. Nutritional and Cultural Aspects of Plant Species Selection for a Controlled Ecological Life Support System; No. NASA-CR-166324; National Aeronautics and Space Administration (NASA): Washington, DC, USA, 1982.
- Meinen, E.; Dueck, T.; Kempkes, F.; Stanghellini, C. Growing Fresh Food on Future Space Missions: Environmental Conditions and Crop Management. Sci. Hortic. 2018, 235, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, R.M.; Fitzpatrick, A.H.; Tibbitts, T.W. Potatoes as a Crop for Space Life Support: Effect of CO2, Irradiance, and Photoperiod on Leaf Photosynthesis and Stomatal Conductance. Front. Plant Sci. 2019, 10, 1632. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, R.M. Potato and Human Exploration of Space: Some Observations from NASA-Sponsored Controlled Environment Studies. Potato Res. 2006, 49, 67–90. [Google Scholar] [CrossRef]
- Molders, K.; Quinet, M.; Decat, J.; Secco, B.; Dulière, E.; Pieters, S.; van der Kooij, T.; Lutts, S.; Van Der Straeten, D. Selection and Hydroponic Growth of Potato Cultivars for Bioregenerative Life Support Systems. Adv. Space Res. 2012, 50, 156–165. [Google Scholar] [CrossRef]
- Paradiso, R.; Ceriello, A.; Pannico, A.; Sorrentino, S.; Palladino, M.; Giordano, M.; Fortezza, R.; De Pascale, S. Design of a Module for Cultivation of Tuberous Plants in Microgravity: The ESA Project “Precursor of Food Production Unit” (PFPU). Front. Plant Sci. 2020, 11, 417. [Google Scholar] [CrossRef]
- Westermann, D.T. Nutritional Requirements of Potatoes. Am. J. Potato Res. 2005, 82, 301–307. [Google Scholar] [CrossRef]
- Ming, D.W. Manufactured Soils for Plant Growth at a Lunar Base. In Lunar Base Agriculture: Soils for Plant Growth; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 1989; pp. 93–105. [Google Scholar] [CrossRef]
- Caporale, A.G.; Vingiani, S.; Palladino, M.; El-Nakhel, C.; Duri, L.G.; Pannico, A.; Rouphael, Y.; De Pascale, S.; Adamo, P. Geo-Mineralogical Characterisation of Mars Simulant MMS-1 and Appraisal of Substrate Physico-Chemical Properties and Crop Performance Obtained with Variable Green Compost Amendment Rates. Sci. Total Environ. 2020, 720, 137543. [Google Scholar] [CrossRef]
- Duri, L.G.; El-Nakhel, C.; Caporale, A.G.; Ciriello, M.; Graziani, G.; Pannico, A.; Palladino, M.; Ritieni, A.; De Pascale, S.; Vingiani, S.; et al. Mars Regolith Simulant Ameliorated by Compost as in Situ Cultivation Substrate Improves Lettuce Growth and Nutritional Aspects. Plants 2020, 9, 628. [Google Scholar] [CrossRef] [PubMed]
- Gilrain, M.R.; Hogan, J.A.; Cowan, R.M.; Finstein, M.S.; Logendra, L.S. Preliminary Study of Greenhouse Grown Swiss Chard in Mixtures of Compost and Mars Regolith Simulant; SAE International: Warrendale, PA, USA, 1999. [Google Scholar] [CrossRef]
- Mortley, D.G.; Aglan, H.A.; Bonsi, C.K.; Hill, W.A. Growth of Sweetpotato in Lunar and Mars Simulants; SAE International: Warrendale, PA, USA, 2000. [Google Scholar] [CrossRef]
- Wamelink, G.W.W.; Frissel, J.Y.; Krijnen, W.H.J.; Verwoert, M.R.; Goedhart, P.W. Can Plants Grow on Mars and the Moon: A Growth Experiment on Mars and Moon Soil Simulants. PLoS ONE 2014, 9, e103138. [Google Scholar] [CrossRef] [PubMed]
- Caporale, A.G.; Amato, M.; Duri, L.G.; Bochicchio, R.; De Pascale, S.; Simeone, G.D.R.; Palladino, M.; Pannico, A.; Rao, M.A.; Rouphael, Y.; et al. Can Lunar and Martian Soils Support Food Plant Production? Effects of Horse/Swine Monogastric Manure Fertilisation on Regolith Simulants Enzymatic Activity, Nutrient Bioavailability, and Lettuce Growth. Plants 2022, 11, 3345. [Google Scholar] [CrossRef] [PubMed]
- Caporale, A.G.; Palladino, M.; De Pascale, S.; Duri, L.G.; Rouphael, Y.; Adamo, P. How to Make the Lunar and Martian Soils Suitable for Food Production—Assessing the Changes after Manure Addition and Implications for Plant Growth. J. Environ. Manag. 2023, 325, 116455. [Google Scholar] [CrossRef] [PubMed]
- Duri, L.; Pannico, A.; Petropoulos, S.; Caporale, A.G.; Adamo, P.; Graziani, G.; Ritieni, A.; Pascale, S.; Rouphael, Y. Bioactive Compounds and Antioxidant Activity of Lettuce Grown in Different Mixtures of Monogastric-Based Manure with Lunar and Martian Soils. Front. Nutr. 2022, 9, 890786. [Google Scholar] [CrossRef] [PubMed]
- Caporale, A.G.; Paradiso, R.; Liuzzi, G.; Palladino, M.; Amitrano, C.; Arena, C.; Arouna, N.; Verrillo, M.; Cozzolino, V.; De Pascale, S.; et al. Green Compost Amendment Improves Potato Plant Performance on Mars Regolith Simulant as Substrate for Cultivation in Space. Plant Soil 2023, 486, 217–233. [Google Scholar] [CrossRef]
- Boujelben, N.; Bouzid, J.; Elouear, Z.; Feki, M.; Jamoussi, F.; Montiel, A. Phosphorus removal from aqueous solution using iron coated natural and engineered sorbents. J. Hazard. Mater. 2008, 151, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Gryschko, R.; Kuhnle, R.; Terytze, K.; Breuer, J.; Stahr, K. Soil Extraction of Readily Soluble Heavy Metals and As with 1 M NH4NO3-Solution—Evaluation of DIN 19730 (6 pp). J. Soils Sediments 2005, 5, 101–106. [Google Scholar] [CrossRef]
- Feng, M.-H.; Shan, X.-Q.; Zhang, S.-Z.; Wen, B. Comparison of a Rhizosphere-Based Method with Other One-Step Extraction Methods for Assessing the Bioavailability of Soil Metals to Wheat. Chemosphere 2005, 59, 939–949. [Google Scholar] [CrossRef]
- Zhu, Q.H.; Huang, D.Y.; Liu, S.L.; Luo, Z.C.; Zhu, H.H.; Zhou, B.; Lei, M.; Rao, Z.X.; Cao, X.L. Assessment of Single Extraction Methods for Evaluating the Immobilization Effect of Amendments on Cadmium in Contaminated Acidic Paddy Soil. Plant Soil Environ. 2012, 58, 98–103. [Google Scholar] [CrossRef]
- Maggi, F.; Pallud, C. Space Agriculture in Micro- and Hypo-Gravity: A Comparative Study of Soil Hydraulics and Biogeochemistry in a Cropping Unit on Earth, Mars, the Moon and the Space Station. Planet. Space Sci. 2010, 58, 1996–2007. [Google Scholar] [CrossRef]
- Kasiviswanathan, P.; Swanner, E.D.; Halverson, L.J.; Vijayapalani, P. Farming on Mars: Treatment of Basaltic Regolith Soil and Briny Water Simulants Sustains Plant Growth. PLoS ONE 2022, 17, e0272209. [Google Scholar] [CrossRef] [PubMed]
- Epstein, E. Effect of Soil Temperature at Different Growth Stages on Growth and Development of Potato Plants. Agron. J. 1966, 58, 169–171. [Google Scholar] [CrossRef]
- Paradiso, R.; Arena, C.; Rouphael, Y.; d’Aquino, L.; Makris, K.; Vitaglione, P.; De Pascale, S. Growth, Photosynthetic Activity and Tuber Quality of Two Potato Cultivars in Controlled Environment as Affected by Light Source. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2019, 153, 725–735. [Google Scholar] [CrossRef]
- Frusciante, L.; Roversi, G. La Patata—Coltura & Cultura; Bayer Crop Science: Bologna, Italy, 2011; p. 928. [Google Scholar]
- Barber, S.A. Soil Nutrient Bioavailability: A Mechanistic Approach; John Wiley & Sons: New York, NY, USA, 1995; ISBN 978-0-471-58747-7. [Google Scholar]
- Gowda, V.R.P.; Henry, A.; Yamauchi, A.; Shashidhar, H.E.; Serraj, R. Root Biology and Genetic Improvement for Drought Avoidance in Rice. Field Crops Res. 2011, 122, 1–13. [Google Scholar] [CrossRef]
- Roumet, C.; Birouste, M.; Picon-Cochard, C.; Ghestem, M.; Osman, N.; Vrignon-Brenas, S.; Cao, K.; Stokes, A. Root Structure–Function Relationships in 74 Species: Evidence of a Root Economics Spectrum Related to Carbon Economy. New Phytol. 2016, 210, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Rice, B.L.; Dlouhy, H.; Zwart, S.R. Assessment of Nutritional Intake during Space Flight and Space Flight Analogs. Procedia Food Sci. 2013, 2, 27–34. [Google Scholar] [CrossRef]
- Udensi, U.K.; Tchounwou, P.B. Potassium Homeostasis, Oxidative Stress, and Human Disease. Int. J. Clin. Exp. Physiol. 2017, 4, 111–122. [Google Scholar] [CrossRef]
- Bacchi, M.A.; De Nadai Fernandes, E.A.; Tsai, S.M.; Santos, L.G.C. Conventional and Organic Potatoes: Assessment of Elemental Composition Using K0-INAA. J. Radioanal. Nucl. Chem. 2004, 259, 421–424. [Google Scholar] [CrossRef]
- LeRiche, E.L.; Wang-Pruski, G.; Zheljazkov, V.D. Distribution of Elements in Potato (Solanum tuberosum L.) Tubers and Their Relationship to After-Cooking Darkening. HortScience 2009, 44, 1866–1873. [Google Scholar] [CrossRef]
- Zhou, L.; Mu, T.; Ma, M.; Zhang, R.; Sun, Q.; Xu, Y. Nutritional Evaluation of Different Cultivars of Potatoes (Solanum tuberosum L.) from China by Grey Relational Analysis (GRA) and Its Application in Potato Steamed Bread Making. J. Integr. Agric. 2019, 18, 231–245. [Google Scholar] [CrossRef]
- Bernard, E.; Larkin, R.P.; Tavantzis, S.; Erich, M.S.; Alyokhin, A.; Gross, S.D. Rapeseed Rotation, Compost and Biocontrol Amendments Reduce Soilborne Diseases and Increase Tuber Yield in Organic and Conventional Potato Production Systems. Plant Soil 2014, 374, 611–627. [Google Scholar] [CrossRef]
- Stark, J.C.; Porter, G.A. Potato Nutrient Management in Sustainable Cropping Systems. Am. J. Potato Res. 2005, 82, 329–338. [Google Scholar] [CrossRef]
- Wilson, C.; Zebarth, B.J.; Burton, D.L.; Goyer, C.; Moreau, G.; Dixon, T. Effect of Diverse Compost Products on Potato Yield and Nutrient Availability. Am. J. Potato Res. 2019, 96, 272–284. [Google Scholar] [CrossRef]
- Singh, B.; Dutt, S.; Raigond, P. Potato Glycoalkaloids. In Potato: Nutrition and Food Security; Raigond, P., Singh, B., Dutt, S., Chakrabarti, S.K., Eds.; Springer: Singapore, 2020; pp. 191–211. [Google Scholar] [CrossRef]
- Knuthsen, P.; Jensen, U.; Schmidt, B.; Larsen, I.K. Glycoalkaloids in Potatoes: Content of Glycoalkaloids in Potatoes for Consumption. J. Food Compos. Anal. 2009, 22, 577–581. [Google Scholar] [CrossRef]
- Jin, C.-Y.; Liu, H.; Xu, D.; Zeng, F.-K.; Zhao, Y.-C.; Zhang, H.; Liu, G. Glycoalkaloids and Phenolic Compounds in Three Commercial Potato Cultivars Grown in Hebei, China. Food Sci. Hum. Wellness 2018, 7, 156–162. [Google Scholar] [CrossRef]
- Benkeblia, N. Potato Glycoalkaloids: Occurrence, Biological Activities and Extraction for Biovalorisation—A Review. Int. J. Food Sci. Technol. 2020, 55, 2305–2313. [Google Scholar] [CrossRef]
- Kozukue, N.; Yoon, K.-S.; Byun, G.-I.; Misoo, S.; Levin, C.E.; Friedman, M. Distribution of Glycoalkaloids in Potato Tubers of 59 Accessions of Two Wild and Five Cultivated Solanum Species. J. Agric. Food Chem. 2008, 56, 11920–11928. [Google Scholar] [CrossRef] [PubMed]
- Dhalsamant, K.; Singh, C.B.; Lankapalli, R. A Review on Greening and Glycoalkaloids in Potato Tubers: Potential Solutions. J. Agric. Food Chem. 2022, 70, 13819–13831. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; He, M.; Feng, F.; Feng, X.; Zhang, Y.; Zhang, F. The Distribution and Changes of Glycoalkaloids in Potato Tubers under Different Storage Time Based on MALDI-TOF Mass Spectrometry Imaging. Talanta 2021, 221, 121453. [Google Scholar] [CrossRef] [PubMed]
- Omayio, D.G.; Abong, G.O.; Okoth, M.W. A Review of Occurrence of Glycoalkaloids in Potato and Potato Products. Curr. Res. Nutr. Food Sci. 2016, 4, 195–202. [Google Scholar] [CrossRef]
- Wesseling, J.G.; Elbers, J.A.; Kabat, P.; Van den Broek, B.J. SWATRE: Instructions for Input. Intern. Note Winand Star. Cent. Wagening. Neth. 1991, 1991, 700. [Google Scholar]
- van Genuchten, M.T. A Closed-Form Equation for Predicting the Hydraulic Conductivity of Unsaturated Soils. Soil Sci. Soc. Am. J. 1980, 44, 892–898. [Google Scholar] [CrossRef]
- ISO 19730:2008; Soil Quality—Extraction of Trace Elements from Soil Using Ammonium Nitrate Solution. ISO: Geneva, Switzerland, 2008.
- Rauret, G.; López-Sánchez, J.F. New Sediment and Soil CRMs for Extractable Trace Metal Content. Int. J. Environ. Anal. Chem. 2001, 79, 81–95. [Google Scholar] [CrossRef]
- Latimer, G.W., Jr. (Ed.) Official Methods of Analysis of AOAC International, 20th ed.; AOAC International: Rockville, MD, USA, 2016. [Google Scholar]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Total Dietary Fiber Assay Kit. Available online: https://www.megazyme.com/total-dietary-fiber-assay-kit (accessed on 22 January 2024).
- Maldonado, A.F.; Mudge, E.; Gänzle, M.G.; Schieber, A. Extraction and Fractionation of Phenolic Acids and Glycoalkaloids from Potato Peels Using Acidified Water/Ethanol-Based Solvents. Food Res. Int. 2014, 65, 27–34. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| C | N | C/N | S | |
|---|---|---|---|---|
| g kg−1 DW | g kg−1 DW | g kg−1 DW | ||
| VS | 18.2 ± 0.9 b | 1.53 ± 0.05 b | 11.9 ± 0.29 b | 0.83 ± 0.04 c |
| RS | 15.2 ± 0.6 b | 1.43 ± 0.06 b | 10.6 ± 0.10 c | 0.66 ± 0.04 c |
| S100 | 2.2 ± 0.2 c | 0.18 ± 0.01 c | 12.2 ± 0.34 b | 0.09 ± 0.01 d |
| S70C30 | 72.8 ± 2.4 a | 5.63 ± 0.26 a | 13.0 ± 0.19 a | 2.02 ± 0.09 b |
| R100 | 2.1 ± 0.2 c | 0.30 ± 0.02 c | 7.0 ± 0.31 d | 0.13 ± 0.01 d |
| R70C30 | 75.4 ± 2.9 a | 5.76 ± 0.22 a | 13.1 ± 0.11 a | 2.23 ± 0.10 a |
| Soil (S) | *** | *** | ** | *** |
| RH | 31.6 ± 14.5 | 2.51 ± 1.10 | 11.6 ± 0.91 | 1.02 ± 0.40 |
| BK | 30.3 ± 14.3 | 2.43 ± 1.07 | 11.0 ± 1.04 | 0.96 ± 0.38 |
| RH vs. BK (RB) | ns | ns | ns | ns |
| S × RB | ns | ns | ns | ns |
| Ca | K | Mg | Na | Fe | Mn | Cu | Zn | B | |
|---|---|---|---|---|---|---|---|---|---|
| mg kg−1 DW | |||||||||
| VS | 2895 b | 1391 a | 185 d | 119 b | 1.1 cd | 0.40 c | 0.27 a | 0.51 | 0.08 c |
| RS | 6425 a | 260 e | 282 b | 108 b | 1.8 b | 0.66 b | 0.05 d | 0.24 | 0.01 c |
| S100 | 1060 d | 65 f | 72 e | 28 c | 1.7 bc | 1.4 a | 0.13 c | 0.11 | 0.03 c |
| S70C30 | 1522 c | 653 d | 236 c | 39 c | 2.7 a | 0.67 b | 0.22 b | 0.14 | 0.45 c |
| R100 | 2944 b | 1069 b | 510 a | 286 a | 0.8 d | 0.30 c | 0.13 c | 0.08 | 6.7 b |
| R70C30 | 3140 b | 947 c | 505 a | 308 a | 0.7 d | 0.28 c | 0.11 c | 0.01 | 8.6 a |
| Soil (S) | *** | *** | *** | *** | *** | *** | *** | ns | *** |
| RH | 2979 | 801 | 318 | 137 | 1.5 | 0.59 | 0.17 | 0.18 | 2.0 |
| BK | 3016 | 661 | 279 | 159 | 1.4 | 0.65 | 0.14 | 0.17 | 3.3 |
| RH vs. BK (RB) | ns | *** | ** | ns | ns | ns | ns | ns | *** |
| S × RB | ns | *** | ** | * | ns | *** | ns | ns | *** |
| Ca | K | Mg | Na | Fe | Mn | Cu | Zn | B | |
|---|---|---|---|---|---|---|---|---|---|
| mg kg−1 DW | |||||||||
| VS | 11,723 c | 1046 a | 437 b | 98 b | 109 d | 43 c | 19 a | 14 a | 0.8 b |
| RS | 5204 e | 215 e | 286 c | 85 b | 291 a | 610 a | 14 b | 6.3 b | 0.7 b |
| S100 | 15,440 a | 60 f | 152 d | 36 c | 184 b | 89 b | 2.3 cd | 1.8 c | 0.5 b |
| S70C30 | 14,269 b | 512 c | 323 c | 41 c | 275 a | 82 b | 4.1 c | 5.4 b | 1.4 b |
| R100 | 5220 e | 364 d | 401 b | 177 a | 9.6 e | 16 d | 0.8 d | 0.9 c | 8.1 a |
| R70C30 | 8117 d | 788 b | 616 a | 194 a | 138 c | 31 c | 3.8 c | 5.2 b | 9.4 a |
| Soil (S) | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| RH | 10,220 | 534 | 382 | 99 | 174 | 148 | 7.8 | 6.2 | 2.8 |
| BK | 9771 | 461 | 356 | 111 | 162 | 142 | 7.0 | 5.1 | 4.2 |
| RH vs. BK (RB) | ns | *** | ns | ns | ns | ns | ns | * | ** |
| S × RB | ns | ** | ns | * | ** | ns | ns | ns | *** |
| VS | RS | S100 | S70C30 | R100 | R70C30 | Sig. | |
|---|---|---|---|---|---|---|---|
| g kg−1 DW | |||||||
| C | 419 | 399 | 419 | 417 | 419 | 420 | ns |
| K | 18.3 d | 16.0 e | 20.8 c | 25.8 a | 23.0 b | 25.8 a | *** |
| N | 11.7 c | 11.9 c | 10.8 c | 14.0 b | 17.4 a | 15.0 b | *** |
| S | 2.6 bc | 2.4 c | 3.1 ab | 3.2 ab | 2.9 abc | 3.7 a | ** |
| P | 2.5 a | 1.3 c | 1.9 b | 1.9 b | 2.1 b | 2.0 b | *** |
| Mg | 1.0 e | 1.2 d | 1.3 c | 1.5 c | 1.8 a | 1.7 b | *** |
| mg kg−1 DW | |||||||
| Ca | 197 | 419 | 179 | 366 | 265 | 357 | ns |
| Na | 37.8 c | 66.4 bc | 78.3 bc | 88.2 abc | 134 ab | 144 a | ** |
| Fe | 23.2 ab | 13.5 b | 16.6 b | 14.2 b | 33.5 a | 32.9 a | ** |
| Zn | 16.0 b | 9.2 c | 6.7 c | 21.7 a | 9.2 c | 17.8 b | *** |
| B | 2.8 c | 5.3 c | 4.5 c | 3.0 c | 25.0 a | 9.3 b | *** |
| Mn | 4.3 d | 5.0 cd | 5.6 bc | 7.4 a | 6.2 b | 8.3 a | *** |
| Cu | 5.4 b | 3.4 c | 6.0 b | 5.5 b | 7.6 a | 4.9 b | *** |
| VS | RS | S100 | S70C30 | R100 | R70C30 | Sig. | |
|---|---|---|---|---|---|---|---|
| g plant−1 DW | |||||||
| C | 7.4 ab | 5.6 c | 7.6 a | 7.0 ab | 3.4 d | 6.5 bc | *** |
| mg plant−1 DW | |||||||
| K | 322 c | 227 d | 376 b | 436 a | 189 d | 401 ab | *** |
| N | 208 bc | 169 d | 198 c | 237 a | 141 e | 233 ab | *** |
| S | 45.9 a | 33.2 b | 57.3 a | 54.7 a | 24.2 b | 54.5 a | *** |
| P | 43.4 a | 18.6 c | 35.1 b | 32.1 b | 17.5 c | 31.4 b | *** |
| Mg | 18.1 b | 16.7 bc | 24.4 a | 24.5 a | 14.7 c | 25.8 a | *** |
| Ca | 3.4 | 5.8 | 3.3 | 6.1 | 2.2 | 5.6 | ns |
| Na | 0.7 b | 0.9 b | 1.4 ab | 1.5 ab | 1.1 b | 2.2 a | * |
| µg plant−1 DW | |||||||
| Fe | 404 ab | 188 c | 302 bc | 241 bc | 277 bc | 501 a | ** |
| Zn | 281 b | 129 c | 122 c | 367 a | 73 c | 275 b | *** |
| B | 43.0 c | 75.2 c | 81.5 c | 52.1 c | 209 a | 145 b | *** |
| Mn | 74.5 c | 70.9 c | 102 b | 125 a | 50.6 d | 128 a | *** |
| Cu | 94.8 ab | 48.8 d | 108 a | 92.5 ab | 62.1 cd | 75.4 bc | *** |
| Proteins | Starch | Total Dietary Fiber | Ascorbic Acid | α-Solanine | α-Chaconine | Total Glicoalkaloids | |
|---|---|---|---|---|---|---|---|
| (mg/g DM) | (%) | (g/100g dw) | (mg/100g fw) | (mg/kg dw) | (mg/kg dw) | (mg/kg dw) | |
| VS | 63.00 ± 0.46 d | 54.52 ± 0.52 a | 10.63 ± 0.05 b | 18.77 ± 1.64 | 269.52 ± 15.39 ab | 600.12 ± 39.59 a | 869.64 ± 53.02 a |
| RS | 45.64 ± 0.47 e | 54.78 ± 0.18 a | 9.34 ± 0.08 e | 15.83 ± 0.58 | 214.76 ± 18.95 bc | 427.69 ± 37.69 b | 642.44 ± 50.13 b |
| S100 | 45.73 ± 0.26 e | 48.43 ± 0.14 b | 10.37 ± 0.09 d | 17.24 ± 0.70 | 191.07 ± 22.93 c | 395.25 ± 28.40 b | 586.32 ± 44.44 b |
| S70C30 | 82.82 ± 0.61 a | 47.49 ± 0.07 c | 10.58 ± 0.07 bc | 18.12 ± 0.56 | 234.80 ± 17.01 bc | 484.49 ± 23.71 b | 719.29 ± 40.40 b |
| R100 | 70.57 ± 0.26 c | 48.17 ± 0.28 bc | 11.17 ± 0.02 a | 15.77 ± 0.95 | 320.57 ± 22.86 a | 646.73 ± 27.38 a | 967.29 ± 49.80 a |
| R70C30 | 75.68 ± 0.29 b | 45.87 ± 0.36 d | 10.42 ± 0.06 cd | 16.86 ± 1.12 | 217.82 ± 16.63 bc | 465.34 ± 30.35 b | 683.15 ± 46.02 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caporale, A.G.; Paradiso, R.; Palladino, M.; Arouna, N.; Izzo, L.; Ritieni, A.; De Pascale, S.; Adamo, P. Assessment of Fertility Dynamics and Nutritional Quality of Potato Tubers in a Compost-Amended Mars Regolith Simulant. Plants 2024, 13, 747. https://doi.org/10.3390/plants13050747
Caporale AG, Paradiso R, Palladino M, Arouna N, Izzo L, Ritieni A, De Pascale S, Adamo P. Assessment of Fertility Dynamics and Nutritional Quality of Potato Tubers in a Compost-Amended Mars Regolith Simulant. Plants. 2024; 13(5):747. https://doi.org/10.3390/plants13050747
Chicago/Turabian StyleCaporale, Antonio Giandonato, Roberta Paradiso, Mario Palladino, Nafiou Arouna, Luana Izzo, Alberto Ritieni, Stefania De Pascale, and Paola Adamo. 2024. "Assessment of Fertility Dynamics and Nutritional Quality of Potato Tubers in a Compost-Amended Mars Regolith Simulant" Plants 13, no. 5: 747. https://doi.org/10.3390/plants13050747
APA StyleCaporale, A. G., Paradiso, R., Palladino, M., Arouna, N., Izzo, L., Ritieni, A., De Pascale, S., & Adamo, P. (2024). Assessment of Fertility Dynamics and Nutritional Quality of Potato Tubers in a Compost-Amended Mars Regolith Simulant. Plants, 13(5), 747. https://doi.org/10.3390/plants13050747