
Long-Term Benefits of Cenchrus fungigraminus Residual Roots Improved the Quality and Microbial Diversity of Rhizosphere Sandy Soil through Cellulose Degradation in the Ulan Buh Desert, Northwest China


 ,
,
Abstract
1. Introduction
2. Material and Methods
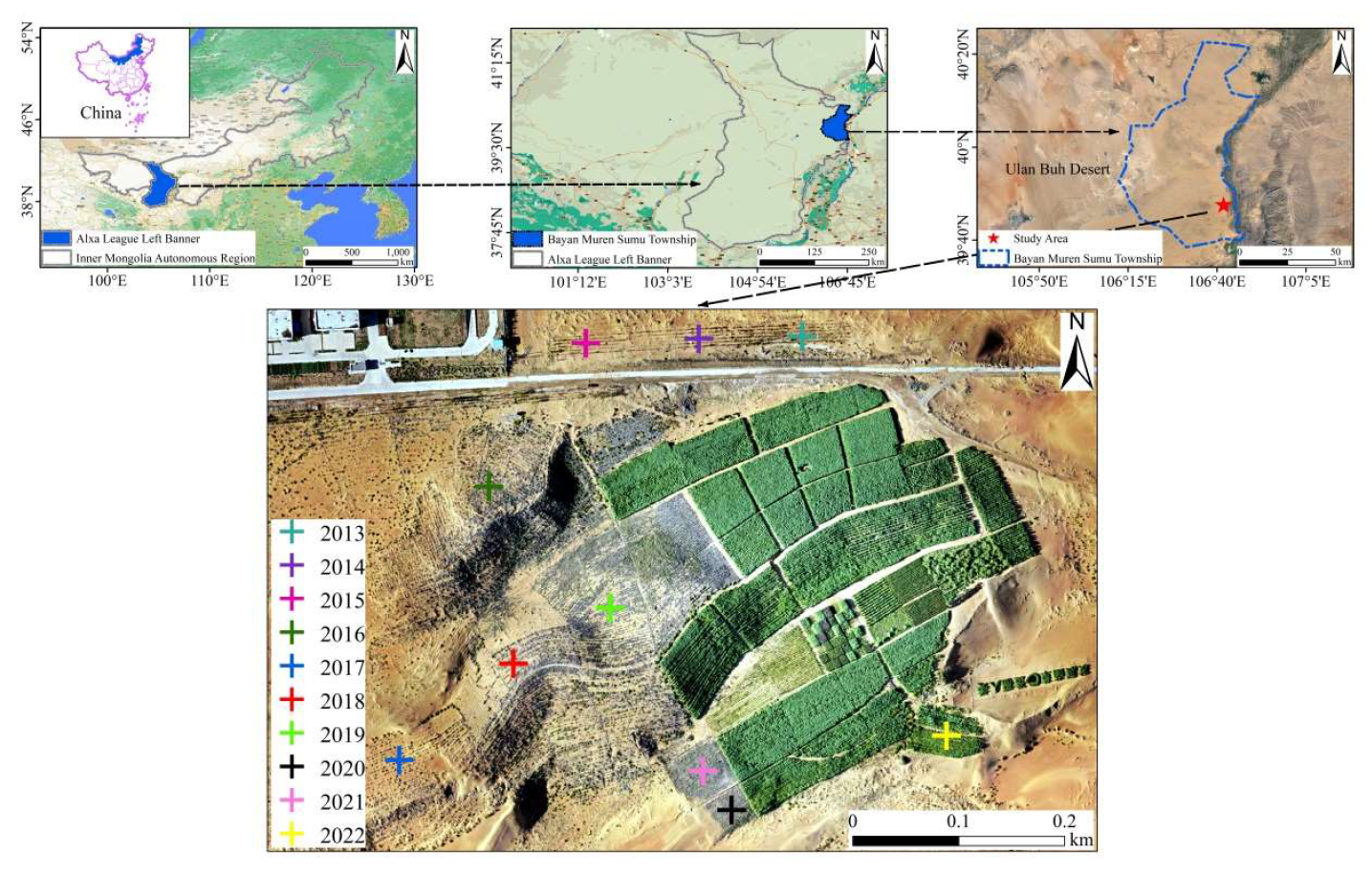
2.1. Study Area Description
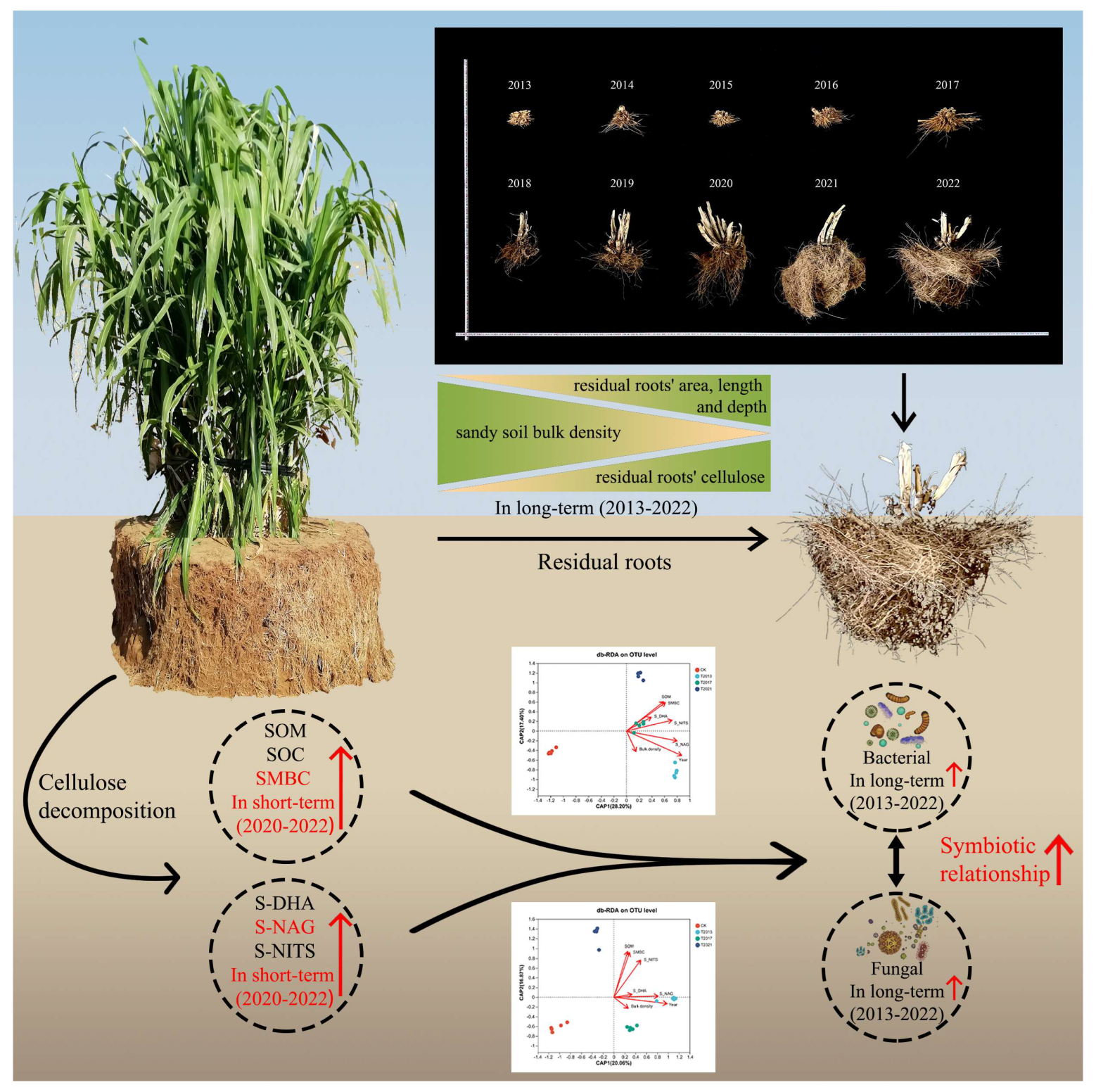
2.2. Experimental Design and Field Sampling
2.3. Determination of the Physical–Chemical Properties of the Sand Soil
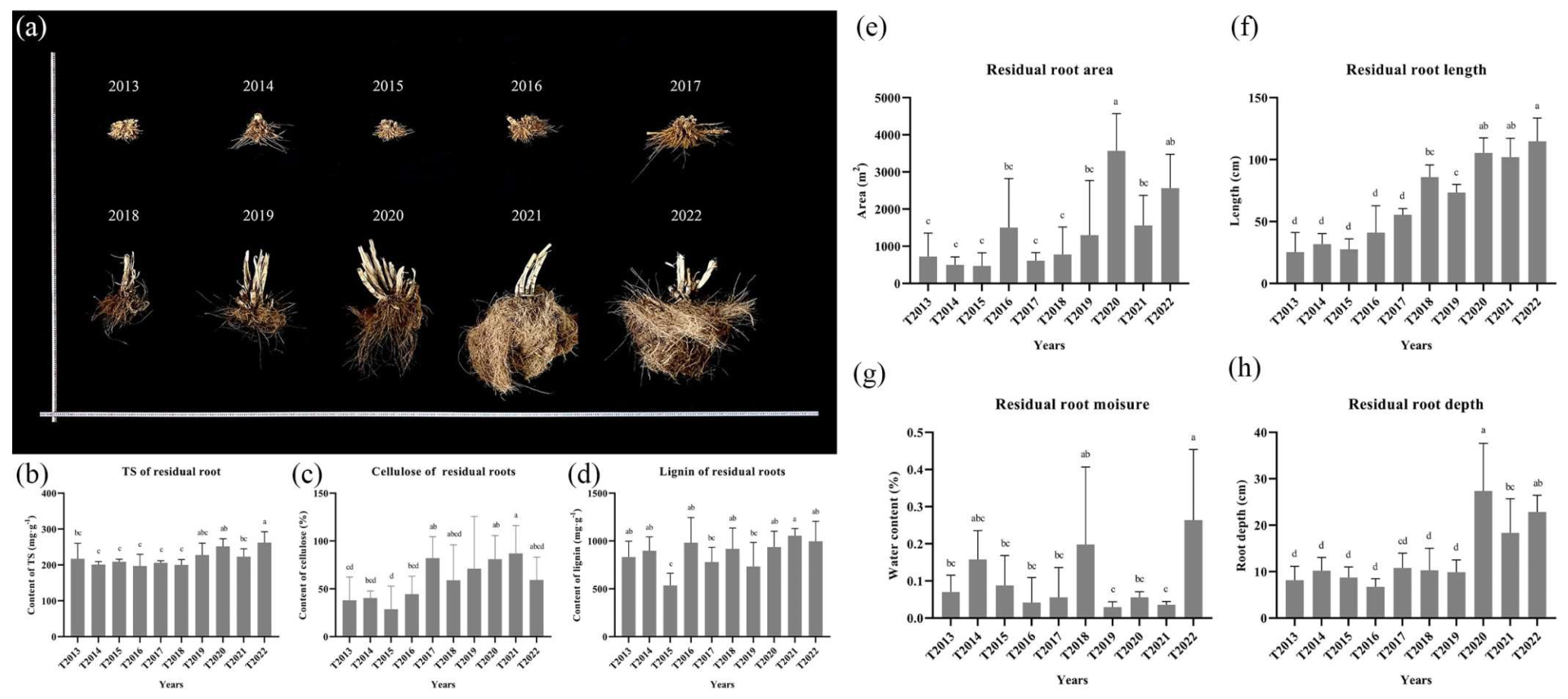
2.4. Residual Root Measurement
2.5. Total DNA Extraction and Sequencing
2.6. Bioinformatic Processing and Statistical Analysis
3. Results
3.1. Sandy Soil and C. fungigraminus Residual Root Measurements
3.2. Basic Sequencing Data of the Four Sandy Soils
3.3. Taxonomic Profile of Sandy Soil Bacterial and Fungal Communities
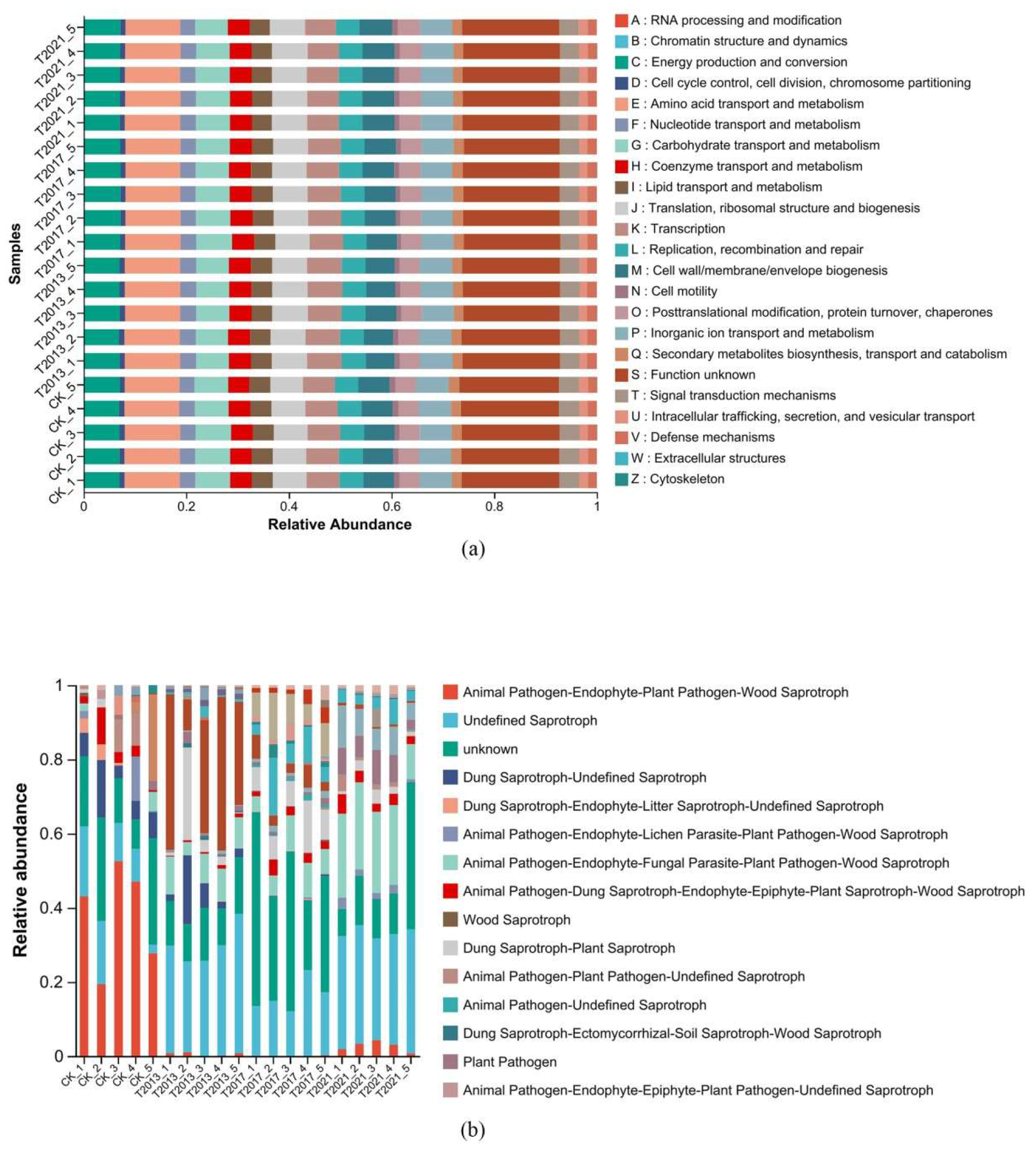
3.4. Microbial Communities, Functional, and Co-Occurrence Network Analysis
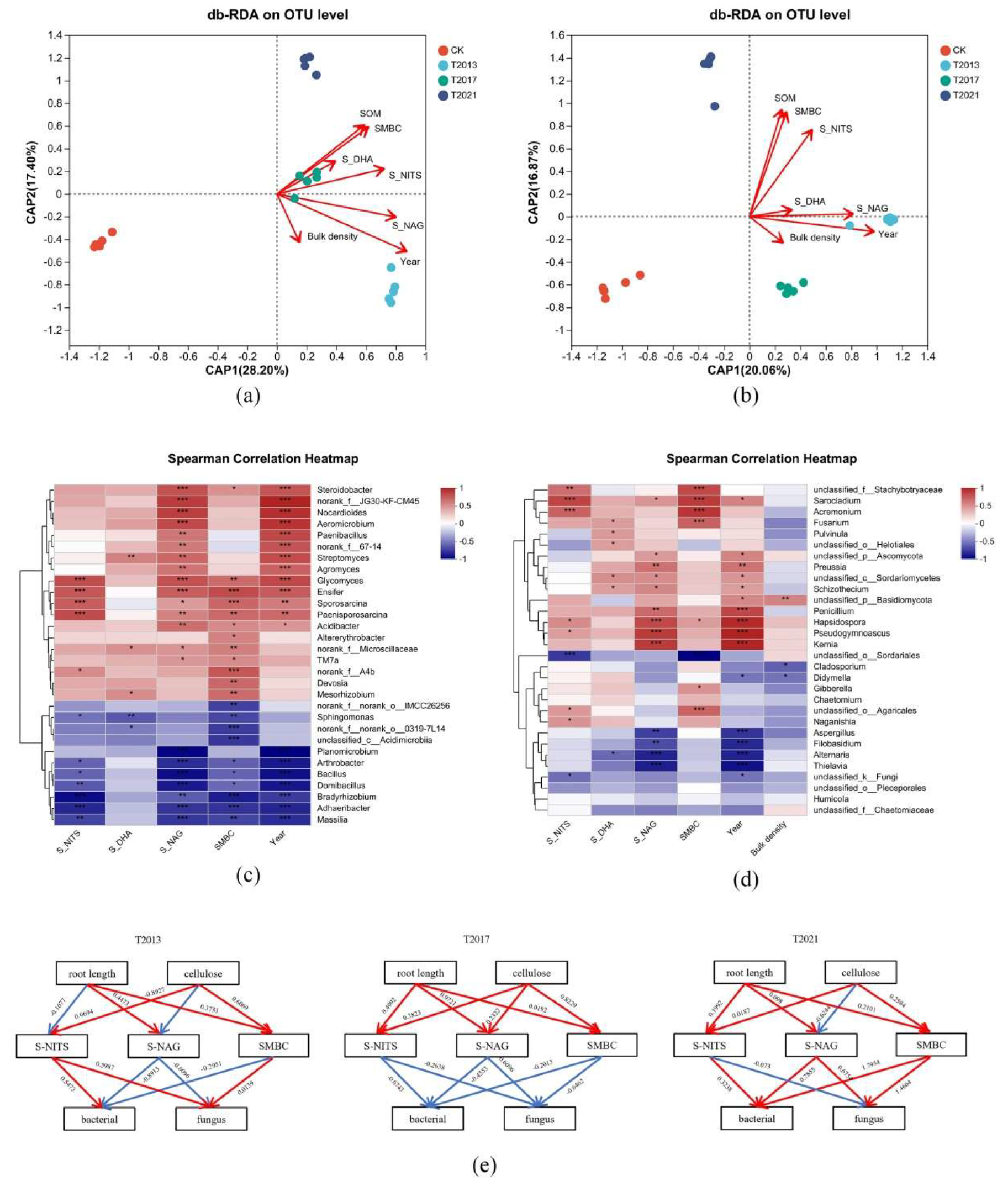
3.5. Correlations between Bacterial and Fungal Structures and Environmental Variables
4. Discussion
4.1. Effects of Plant Residue Retention on Soil Physicochemical Properties and Carbon Stock
4.2. Effect of Plant Residues on Soil Microbial Structure and Diversity
4.3. Inspiration of Planting Annual Plants as “Pioneer Plants” in Sandy Land in the Long Term
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, C.; Park, T.; Wang, X.H.; Piao, S.L.; Xu, B.D.; Chaturvedi, R.K.; Fuchs, R.; Brovkin, V.; Cias, P.; Fensholt, R.; et al. China and India lead in greening of the world through land-use management. Nat. Sustain. 2019, 2, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Han, X.Y.; Jia, G.P.; Yang, G.; Wang, N.; Liu, F.; Chen, H.Y.; Guo, X.Y.; Yang, W.B.; Liu, J. Spatiotemporal dynamic evolution and driving factors of desertification in the Mu Us Sandy Land in 30 years. Sci. Rep. 2020, 10, 21734. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.S.; Li, X.R.; Liu, L.C.; Jia, R.L.; Zhang, J.G.; Wang, T. Distribution, biomass, and dynamics of roots in a revegetated stand of Caragana korshinskii in the Tengger Desert, northwestern China. J. Plant Res. 2009, 122, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.Z.; Zhao, H.L. Soil properties and plant species in an age sequence of Caragana microphylla plantations in the Horqin Sandy Land, North China. Ecol. Eng. 2003, 20, 223–235. [Google Scholar] [CrossRef]
- Li, X.J.; Li, X.R.; Song, W.M.; Gao, Y.P.; Zheng, J.G.; Jia, R.L. Effects of crust and shrub patches on runoff, sedimentation, and related nutrient (C, N) redistribution in the desertified steppe zone of the Tengger Desert, Northern China. Geomorphology 2008, 96, 221–232. [Google Scholar] [CrossRef]
- Li, Y.Q.; Zhao, X.Y.; Zhang, F.X.; Awada, T.; Wang, S.K.; Zhao, H.L.; Zhang, T.H.; Li, Y.L. Accumulation of soil organic carbon during natural restoration of desertified grassland in China’s Horqin Sandy Land. J. Arid Land 2015, 7, 328–340. [Google Scholar] [CrossRef]
- Zuo, X.A.; Zhao, X.Y.; Wang, S.K.; Li, Y.L.; Lian, J.; Zhou, X. Influence of dune stabilization on relationship between plant diversity and productivity in Horqin Sand Land, Northern China. Environ. Earth Sci. 2012, 67, 1547–1556. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Tang, Z.X.; You, Y.M.; Guo, X.W.; Wu, C.J.; Liu, S.R.; Osbert Sun, J.X. Differential effects of forest-floor litter and roots on soil organic carbon formation in a temperate oak forest. Soil Biol. Biochem. 2023, 180, 109017. [Google Scholar] [CrossRef]
- Zhang, W.P.; Fornara, D.; Yang, H.; Yu, R.P.; Callaway, R.M.; Li, L. Plant litter strengthens positive biodiversity-ecosystem functioning relationships over time. Trends Ecol. Evol. 2023, 38, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Sandeep, S.; Pritpal, S. Tillage intensity and straw retention impacts on soil organic carbon, phosphorus and biological pools in soil aggregates under rice-wheat cropping system in Punjab, North-western India. Eur. J. Agron. 2023, 149, 126913. [Google Scholar] [CrossRef]
- Zhou, J.; Chen, S.Q.; Shi, W.J.; Rakefet, D.S.; Li, S.T.; Yang, F.L.; Lin, Z.X. Transcriptome profiling reveals the effects of drought tolerance in Giant Juncao. BMC Plant Biol. 2021, 21, 2–21. [Google Scholar] [CrossRef]
- Zheng, H.K.; Wang, B.Y.; Hua, X.T.; Gao, R.T.; Wang, Y.H.; Zhang, Z.X.; Zhang, Y.X.; Mei, J.; Huang, Y.J.; Huang, Y.M.; et al. A near-complete genome assembly of the allotetrapolyploid Cenchrus fungigraminus (JUJUNCAO) provides insights into its evolution and C4 photosynthesis. Plant Commun. 2023, 3, 100633. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.D.; Dang, X.H.; Gao, Y.; Yang, X.; Liang, Y.M.; Zhao, C.; Duan, X.T. Natural deterioration processes of Salix psammophila sand barriers in atmospheric exposure section. Front. Plant Sci. 2022, 13, 850391. [Google Scholar] [CrossRef] [PubMed]
- He, L.Z.; Jia, Z.Q.; Li, Q.X.; Zhang, Y.Y.; Wu, R.N.; Dai, J.; Gao, Y. Fine root dynamic characteristics and effect on plantation’s carbon sequestration of three Salix shrub plantations in Tibetan Plateau alpine sandy land. Ecol. Evol. 2021, 11, 2645–2659. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.M.; Gao, Y.; Wang, R.D.; Yang, X. Fungal community characteristics and driving factors during the decaying process of Salix psammophila sand barriers in the desert. PLoS ONE 2021, 16, e0258159. [Google Scholar] [CrossRef] [PubMed]
- Li, C.H.; Li, Y.; Tang, L.S.; Ikenaga, M.; Liu, R.; Xu, G.Q. Soil microbial community shifts explain habitat heterogeneity in two Haloxylon species from a nutrient perspective. Ecol. Evol. 2023, 13, e9727. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wang, B.R.; Fakher, A.; An, S.S.; Kuzyakov, Y. Contribution of roots to soil organic carbon: From growth to decomposition experiment. Catena 2023, 231, 107317. [Google Scholar] [CrossRef]
- Zhang, J.; Guo, Z. Response of soil structure and crop yield to soft rock in Mu Us sandy land, China. Sci. Rep. 2022, 12, 876. [Google Scholar] [CrossRef]
- Post, W.M.; Emanuel, W.R.; Zinke, P.J.; Stangenberger, A.G. Soil carbon pools and world life zone. Nature 1982, 298, 156–159. [Google Scholar] [CrossRef]
- Salehi Hikouei, I.; Kim, S.S.; Mishra, D.R. Machine-Learning Classification of soil bulk density in salt marsh environments. Sensors 2021, 21, 4408. [Google Scholar] [CrossRef]
- Yeomans, J.C.; Bremner, J.M. A rapid and precise method for routine determination of organic carbon in soil. Commun. Soil Sci. Plan. 1988, 19, 1467–1476. [Google Scholar] [CrossRef]
- Pere, R.; Teresa, S.Y.; Joan, R. Equivalent-mass versus fixed-depth as criteria for quantifying soil carbon sequestration: How relevant is the difference? Catena 2022, 214, 106283. [Google Scholar] [CrossRef]
- Macchado, R.R.; Conceicao, S.V.; Leite, H.G.; De Souza, A.L.; Wolff, E. Evaluation of forest growth and carbon stock in forestry projects by system dynamics. J. Clean. Prod. 2015, 96, 520–530. [Google Scholar] [CrossRef]
- Liu, M.H.; Gan, B.P.; Li, Q.; Xiao, W.F.; Song, X.Z. Effects of nitrogen and phosphorus addition on soil extracellular enzyme activity and stoichiometry in Chinese Fir (Cunninghamia lanceolata) Forests. Front. Plant Sci. 2022, 13, 834184. [Google Scholar] [CrossRef] [PubMed]
- Jenkinson, D.S. Microbial biomass in soil: Measurement and turnover. Soil Biol. Biochem. 1981, 5, 415–471. [Google Scholar]
- Niu, Y.Y.; Duan, Y.L.; Li, Y.Q.; Wang, X.Y.; Chen, Y.; Wang, L.L. Soil microbial community responses to short-term nitrogen addition in China’s Horqin Sandy Land. PLoS ONE 2021, 16, e0242643. [Google Scholar] [CrossRef]
- Stark, S.; Hilli, S.; Willfor, S.; Smeds, A.I.; Reundnen, M.; Penttinen, M.; Hautajrvi, R. Composition of lipophilic compounds carbohydrates in the accumulated plant litter and soil organic matter in boreal forests. Eur. J. Soil Sci. 2012, 63, 65–74. [Google Scholar] [CrossRef]
- Edgar Robert, C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Goebel, B.M. Taxonomic Note: A place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. Int. J. Syst. Bacteriol. 1994, 44, 846–849. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microb. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-Independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microb. 2009, 75, 7537. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Barberan, A.; Bates, S.T.; Casamayor, E.O.; Fierer, N. Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 2012, 6, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Kögel-Knabner, I. The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter: Fourteen years on. Soil Biol. Biochem. 2017, 105, A3–A8. [Google Scholar] [CrossRef]
- Lavallee, J.M.; Soong, J.L.; Cotrufo, M.F. Conceptualizing soil organic matter into particulate and mineral-associated forms to address global change in the 21st century. Glob. Chang. Biol. 2020, 26, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Schimel, J.P.; Jastrow, J.D. The importance of anabolism in microbial control over soil carbon storage. Nat. Microbiol. 2017, 2, 17105. [Google Scholar] [CrossRef]
- Andrew, J.; Midwood, K.D.; Hannam, T.G.; David, E.; Melanie, D.J. Storage of soil carbon as particulate and mineral associated organic matter in irrigated woody perennial crops. Geoderma 2021, 403, 115185. [Google Scholar] [CrossRef]
- Das, B.; Chakraborty, D.; Singh, V.K.; Aggarwal, P.; Singh, R.; Dwivedi, B.S.; Mishra, R.P. Effect of integrated nutrient management practice on soil aggregate properties, its stability and aggregate-associated carbon content in an intensive rice-wheat system. Soil Till. Res. 2014, 136, 9–18. [Google Scholar] [CrossRef]
- Hao, X.Y.; He, W.; Lam, S.K.; Li, P.; Zong, Y.Z.; Zhang, D.S.; Frank Li, H.Y. Enhancement of no-tillage, crop straw return and manure application on field organic matter content overweight the adverse effects of climate change in the arid and semi-arid Northwest China. Agric. For. Meteorol. 2020, 295, 108199. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, Y.D.; Pei, J.B.; Li, M.; Shan, T.; Zhang, W.J.; Wang, J.K. Below ground residues were more conducive to soil organic carbon accumulation than above ground ones. Appl. Soil Ecol. 2020, 148, 103509. [Google Scholar] [CrossRef]
- Xu, Y.D.; Sun, L.J.; Lal, R.; Bol, R.; Wang, Y.; Gao, X.D.; Ding, F.; Liang, S.W.; Li, S.Y.; Wang, J.K. Microbial assimilation dynamics differs but total mineralization from added root and shoot residues is similar in agricultural Alfisols. Soil Biol. Biochem. 2020, 148, 107901. [Google Scholar] [CrossRef]
- Tagliavini, M.; Tonon, G.; Scandellari, F.; Quiñones, A.; Palmieri, S.; Menarbin, G.; Gioacchini, P.; Masia, A. Nutrient recycling during the decomposition of apple leaves (Malus domestica) and mowed grasses in an orchard. Agric. Ecosyst. Environ. 2007, 118, 191–200. [Google Scholar] [CrossRef]
- Li, J.Y.; Yuan, X.L.; Ge, L.; Li, Q.; Li, Z.G.; Wang, L.; Liu, Y. Rhizosphere effects promote soil aggregate stability and associated organic carbon sequestration in rocky areas of desertification. Agric. Ecosyst. Environ. 2020, 304, 107126. [Google Scholar] [CrossRef]
- Zhou, Y.; Bomfim, B.; Bond, W.J.; Boutton, T.W.; Case, M.F.; Coetsee, C.; Davies, A.B.; February, E.C.; Gray, E.F.; Silva, L.C.R.; et al. Soil carbon in tropical savannas mostly derived from grasses. Nat. Geosci. 2023, 16, 710–716. [Google Scholar] [CrossRef]
- Piao, S.L.; Fang, J.Y.; Zhou, L.M.; Tan, K.; Tao, S. Changes in biomass carbon stocks in China’s grasslands between 1982 and 1999. Glob. Biogeochem. Cycles 2007, 21, 1–10. [Google Scholar] [CrossRef]
- Harris, R.B. Rangeland degradation on the Qinghai-Tibetan plateau: A review of the evidence of its magnitude and causes. J. Arid Environ. 2010, 74, 1–12. [Google Scholar] [CrossRef]
- Batjes, N.H. Total carbon and nitrogen in the soils of the world. Eur. J. Soil Sci. 1996, 47, 151–163. [Google Scholar] [CrossRef]
- Xin, X.P.; Jin, D.Y.; Ge, Y.; Wang, J.H.; Chen, J.Q.; Qi, J.G.; Chu, H.S.; Shao, C.L.; Murray, P.J.; Zhao, R.X.; et al. Climate change dominated long-term soil carbon losses of inner Mongolian garasslands. Glob. Biogeochem. Cycles 2020, 34, 2020GB006559. [Google Scholar] [CrossRef]
- Cai, M.K.; Zhao, G.; Zhao, B.; Cong, N.; Zheng, Z.T.; Zhu, J.T.; Duan, X.Q.; Zhang, Y.J. Climate warming alters the relative importance of plant root and microbial community in regulating the accumulation of soil microbial necromass carbon in a Tibetan alpine meadow. Glob. Chang. Biol. 2023, 29, 3193–3204. [Google Scholar] [CrossRef]
- He, M.; Fang, K.; Chen, L.Y.; Feng, X.H.; Qin, S.Q.; Kou, D.; He, H.B.; Liang, C.; Yang, Y.H. Depth-dependent drivers of soil microbial necromass carbon across Tibetan alpine grasslands. Glob. Chang. Biol. 2022, 28, 936–949. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Dou, Y.X.; Wang, B.R.; Wang, Y.Q.; Liang, C.; An, S.S.; Soromotin, A.; Kuzyakov, Y. Increasing contribution of microbial residues to soil organic carbon in grassland restoration chronosequence. Soil Biol. Biochem. 2022, 170, 108688. [Google Scholar] [CrossRef]
- Buckeridge, K.M.; Mason, K.E.; McNamara, N.P.; Ostle, N.; Puissant, J.; Tim, G.; Griffiths, R.I.; Stott, A.W.; Whitaker, J. Environmental and microbial controls on microbial necromass recycling, an important precursor for soil carbon stabilization. Commun. Earth Environ. 2020, 36, 36. [Google Scholar] [CrossRef]
- Chen, H.; Liu, X.W.; Xue, D.; Zhu, D.; Zhan, W.; Li, W.; Wu, N.; Yang, G. Methane emissions during different freezing-thawing periods from a fen on the Qinghai-Tibetan Plateau: Four years of measurements. Agric. For. Meteorol. 2021, 297, 108279. [Google Scholar] [CrossRef]
- Luo, Y.; Xiao, M.L.; Yuan, H.Z.; Liang, C.; Zhu, Z.K.; Xu, J.M.; Kuzyakov, Y.; Wu, J.S.; Ge, T.D.; Tang, C.X. Rice rhizodeposition promotes the build-up of organic carbon in soil via fungal necromass. Soil Biol. Biochem. 2021, 160, 108345. [Google Scholar] [CrossRef]
- Li, M.; Zhang, L.W.; Liu, G. Estimation of thermal properties of soil and backfilling material from thermal response tests (TRTs) for exploiting shallow geothermal energy: Sensitivity, identifiability, and uncertainty. Renew. Energy 2019, 132, 1263–1270. [Google Scholar] [CrossRef]
- Jiao, S.; Peng, Z.H.; Qi, J.J.; Gao, J.M.; Wei, G.H. Linking Bacterial-Fungal relationships to microbial diversity and soil nutrient cycling. mSystems 2021, 6, e01052-20. [Google Scholar] [CrossRef]
- Marasco, R.; Fusi, M.; Mosqueira, M.; Booth, J.M.; Rossi, F.; Cardinale, M.; Michoud, G.; Rolli, E.; Mugnai, G.; Vergani, L.; et al. Rhizosheath-root system changes exopolysaccharide content but stabilizes bacterial community across contrasting seasons in a desert environment. Environ. Microb. 2022, 17, 14. [Google Scholar] [CrossRef]
- Keilhofer, N.; Nachtigall, J.; Kulik, A.; Ecke, M.; Hampp, R.; Süssmuth, R.D.; Fiedler, H.P.; Schrey, S.D. Streptomyces AcH 505 triggers production of a salicylic acid analogue in the fungal pathogen Heterobasidion abietinum that enhances infection of Norway spruce seedlings. Antonie van Leeuwenhoek. Int. J. Gener. Mol. Microbiol. 2018, 111, 691–704. [Google Scholar] [CrossRef]
- Wu, Q.; Ni, M.; Liu, W.C.; Ren, J.H.; Rao, Y.H.; Chen, J.; Lu, C.G. Omics for understanding the mechanisms of Streptomyces lydicus a01 promoting the growth of tomato seedlings. Plant Soil 2018, 431, 129–141. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Srinivas, V.; Alekhya, G.; Prakash, B. Effect of plant growth-promoting Streptomyces sp. on growth promotion and grain yield in chickpea (Cicer arietinum L). 3 Biotech 2015, 5, 799–806. [Google Scholar] [CrossRef] [PubMed]
- Ham, Y.; Kim, T.J. Anthranilamide from Streptomyces spp. inhibited Xanthomonas oryzae biofilm formation without affecting cell growth. Appl. Biol. Chem. 2018, 61, 673–680. [Google Scholar] [CrossRef]
- Vijayabharathi, R.; Sathya, A.; Gopalakrishnan, S. Extracellular biosynthesis of silver nanoparticles using Streptomyces griseoplanus sai-25 and its antifungal activity against Macrophomina phaseolina, the charcoal rot pathogen of sorghum. Biocatal. Agric. Biotechnol. 2018, 14, 166–171. [Google Scholar] [CrossRef]
- Yaish, M.W. Draft genome sequence of the endophytic Bacillus aryabhattai strain SQU-R12, identified from Phoenix dactylifera L. roots. Genome Announc. 2017, 5, e00718. [Google Scholar] [CrossRef] [PubMed]
- Kavamura, V.N.; Santos, S.N.; Taketani, R.G.; Vasconcellos, R.L.F.; Melo, I.S. Draft genome se-quence of plant growth-promoting drought-tolerant Bacillus sp. strain CMAA 1363 isolated from the Brazilian Caatinga Biome. Genome Announc. 2017, 5, e01534. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.G.; Mun, B.G.; Kang, S.M.; Hussain, A.; Shahzad, R.; Seo, C.W.; Kim, A.Y.; Lee, S.U.; Oh, K.Y.; Lee, D.Y.; et al. Bacillus aryabhattai SRB02 tolerates oxidative and nitrosative stress and promotes the growth of soybean by modulating the production of phytohormones. PLoS ONE 2017, 12, e0173203. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.G.; Raddhakrishnan, R.; Al, E. Application of phytohormones producing Bacillus arybhattai SRB02 on soil can stimulate soy-bean plant growth during high temperature. In Proceedings of the KMB Meeting and International Symposium, Gyeongju, Republic of Korea, 24–26 June 2015. [Google Scholar]
- Paredes-Páliz, K.I.; Pajuelo, E.; Doukkali, B.; Ángel Caviedes, M.; Rodríguez-Llorentea, I.D.; Mateos-Naranjo, E. Bacterial inoculants for enhanced seed germination of Spartina densiflora: Implications for restoration of metal polluted areas. Mar. Pollut. Bull. 2016, 110, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Gupta, S.; Marwa, N.; Pandey, V.; Verma, P.C.; Rathaur, S.; Singh, N. Arsenic mediated modifications in Bacillus aryabhattai and their biotechnological applications for arsenic bioremediation. Chemosphere 2016, 164, 524–534. [Google Scholar] [CrossRef]
- Bending, G.D.; Lincoln, S.D.; Sørensen, S.R.; Morgan, J.A.; Aamand, J.; Walker, A. In-field spatial variability in the degradation of the phenyl-urea herbicide isoproturon is the result of interactions between degradative Sphingomonas spp. and soil pH. Appl. Environ. Microb. 2003, 69, 827–834. [Google Scholar] [CrossRef]
- Adhikari, T.B.; Joseph, C.M.; Yang, G.; Phillips, D.A.; Nelson, L.M. Evaluation of bacteria isolated from rice for plant growth promotion and biological control of seedling disease of rice. Can. J. Microbiol. 2001, 47, 916–924. [Google Scholar] [CrossRef]
- Hsueh, P.R.; Teng, L.J.; Yang, P.C.; Chen, Y.C.; Pan, H.J.; Ho, S.W.; Luh, K.T. Nosocomial infections caused by Sphingomonas paucimobilis: Clinical features and microbiological characteristic. Clin. Infect. Dis. 1998, 26, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.L.; He, H.Y.; Liu, Y.Y.; Yang, H.F.; Huang, J.Q. Screening and identification of a wild β-glucosidase producing fungal and its enzymatic property. China Brew. 2012, 31, 53–57. (In Chinese) [Google Scholar]
- Guo, Z.; Han, J.C.; Li, J.; Xu, Y.; Wang, X.L. Effects of long-term fertilization on soil organic carbon mineralization and microbial community structure. PLoS ONE 2019, 14, e0211163. [Google Scholar] [CrossRef]
- Tang, H.M.; Cheng, K.K.; Shi, L.H.; Li, C.; Wen, L.; Li, W.Y.; Sun, M.; Sun, G.; Long, Z.D. Effects of long-term organic matter application on soil carbon accumulation and nitrogen use efficiency in a double-cropping rice field. Environ. Res. 2022, 213, 113700. [Google Scholar] [CrossRef]
- Wang, S.K.; Zuo, X.A.; Zhao, X.Y.; Awada, T.; Luo, Y.Q.; Li, Y.Q.; Qu, H. Dominant plant species shape soil bacterial community in semiarid sandy land of northern China. Ecol. Evol. 2018, 8, 1693–1704. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.S.; Liu, J.M.; Zhang, X.; Weng, C.R.; Lin, Z.X. The flora compositions of nitrogen-fixing bacteria and the differential expression of nifH gene in Pennisetum giganteum z.x.lin Roots. BioMed. Res. Int. 2021, 2021, 5568845. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhou, B.X.; Li, T.T.; Lin, H.; Lin, Z.X.; Lu, G.D.; Liu, Y.L.; Lin, B.S.; Lin, D.M. Isolation of rhizobacteria from the Cenchrus fungigraminus rhizosphere and characterization of their nitrogen-fixing performance and potential role in plant growth promotion. Plant Soil 2023, 486, 405–421. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | CK | T2013 | T2014 | T2015 | T2016 | T2017 | T2018 | T2019 | T2020 | T2021 | T2022 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Bulk density (g·cm−3) | 1.81 abcd ±0.09 | 1.86 ab ±0.06 | 1.87 a ±0.03 | 1.81 abcd ±0.05 | 1.84 abc ±0.07 | 1.80 abcd ±0.06 | 1.79 abcd ±0.04 | 1.75 cd ±0.03 | 1.81 bcd ±0.03 | 1.77 abcd ±0.08 | 1.73 d ±0.04 |
| SOM (g·kg−1) | 1.79 e ±0.28 | 3.15 bc ±0.43 | 2.83 cd ±0.13 | 2.60 cd ±0.37 | 2.54 cde ±0.28 | 2.11 de ±0.23 | 2.22 de ±0.30 | 2.29 de ±0.27 | 4.33 a ±0.81 | 4.20 a ±0.48 | 3.88 ab ±1.03 |
| SOC (%) | 0.10 e ±0.02 | 0.18 bc ±0.02 | 0.16 cd ±0.01 | 0.15 cd ±0.02 | 0.12 de ±0.01c | 0.12 de ±0.01 | 0.13 de ±0.02 | 0.13 de ±0.02 | 0.25 a ±0.05 | 0.24 a ±0.03 | 0.22 ab ±0.06 |
| SMBC (mg·kg−1) | 89.65 e ±3.28 | 107.23 bc ±5.53 | 102.68 cd ±1.91 | 100.51 cd ±5.11 | 99.43 cde ±4.14 | 94.68 de ±3.76 | 95.37 ce ±3.86 | 96.83 cde ±3.74 | 121.55 a ±11.21 | 119.16 a ±6.34 | 117.91 ab ±14.01 |
| Ct (t CO2·hm−2) | 20.66 d ±3.48 | 32.79 cd ±25.3 | 35.55 bcd ±8.75 | 28.06 cd ±10.7 | 41.75 bcd ±23.06 | 44.91 bcd ±6.26 | 73.07 b ±17.48 | 63.25 bc ±12.55 | 175.53 a ±20.79 | 161.01 a ±27.03 | 160.00 a ±28.17 |
| S-DHA(U·g−1) | 1.88 c ±1.64 | 3.18 bc ±2.51 | 3.67 bc ±2.32 | 4.40 bc ±1.85 | 4.37 b ±2.68c | 5.70 b ±1.61a | 8.43 a ±3.32 | 5.78 ab ±3.89 | 4.75 bc ±2.35 | 4.12 bc ±2.23 | 5.95 ab ±2.04 |
| S-NAG (U·g−1) | 0.53 e ±0.09 | 1.52 abcd ±0.47 | 1.81 ab ±0.63 | 1.38 abcd ±0.38 | 1.27 bcd ±10.7 | 1.16 bcde ±0.32 | 1.28 bcd ±0.18 | 1.73 abc ±0.67 | 2.09 a ±0.41 | 0.89 de ±0.26 | 1.01 cde ±0.31 |
| S-NITS (IU·L−1) | 170.58 e ±9.06 | 229.85 a ±10.79 | 168.27 e ±12.32 | 221.45 ab ±3.41 | 205.66 bc ±10.81 | 175.77 de ±11.48 | 218.95 ab ±8.15 | 190.36 cd ±9.08 | 226.84 a ±13.37 | 206.10 bc ±8.21 | 229.10 a ±8.81 |
| Treatments | ACE | Chao | Sobs | Coverage | Shannon | Shannoneven | |
|---|---|---|---|---|---|---|---|
| CK | Bacterial | 1315.448 c ±175.998 | 1335.925 c ±175.163 | 1259.200 c ±161.788 | 0.997 a ±0.001 | 5.205 d ±0.057 | 0.731 c ±0.011 |
| T2013 | 3072.304 b ±318.617 | 2976.522 b ±290.213 | 2400.600 b ±189.942 | 0.983 b ±0.003 | 5.744 c ±0.084 | 0.738 c ±0.014 | |
| T2017 | 3202.074 b ±204.347 | 3152.048 b ±193.117 | 2570.600 b ±136.045 | 0.983 b ±0.002 | 6.092 b ±0.002 | 0.776 b ±0.134 | |
| T2021 | 4368.254 a ±254.529 | 4200.932 a ±248.425 | 3388.400 a ±116.999 | 0.975 c ±0.003 | 6.534 a ±0.081 | 0.804 a ±0.008 | |
| CK | Fungal | 32.366 d ±4.440 | 32.000 d ±4.360 | 32.000 d ±4.360 | 1.000 a ±0.0001 | 2.316 b ±0.230 | 0.671 a ±0.070 |
| T2013 | 116.247 c ±18.990 | 119.400 c ±20.590 | 108.000 c ±14.370 | 0.999 b ±0.0001 | 2.537 b ±0.184 | 0.544 b ±0.051 | |
| T2017 | 200.775 b ±27.470 | 201.364 b ±26.900 | 193.600 b ±26.300 | 0.999 b ±0.0001 | 3.211 a ±0.364 | 0.611 ab ±0.075 | |
| T2021 | 387.931 a ±53.300 | 382.700 a ±46.896 | 329.800 a ±48.122 | 0.998 c ±0.001 | 3.305 a ±0.222 | 0.570 b ±0.027 | |
| Treatments | Degree | Degree Centrality | Closeness Centrality | Betweenness Centrality | |
|---|---|---|---|---|---|
| CK | Bacterial | 691 | 0.596 | 0.554 | 0.259 |
| T2013 | 756 | 0.652 | 0.590 | 0.202 | |
| T2017 | 826 | 0.712 | 0.636 | 0.270 | |
| T2021 | 909 | 0.784 | 0.699 | 0.379 | |
| CK | Fungal | 44 | 0.196 | 0.386 | 0.099 |
| T2013 | 90 | 0.402 | 0.458 | 0.159 | |
| T2017 | 123 | 0.549 | 0.530 | 0.310 | |
| T2021 | 172 | 0.768 | 0.689 | 0.631 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Zhang, L.; Yu, S.; Luo, Z.; Su, D.; Zheng, D.; Zhou, H.; Zhu, J.; Lin, X.; Luo, H.; et al. Long-Term Benefits of Cenchrus fungigraminus Residual Roots Improved the Quality and Microbial Diversity of Rhizosphere Sandy Soil through Cellulose Degradation in the Ulan Buh Desert, Northwest China. Plants 2024, 13, 708. https://doi.org/10.3390/plants13050708
Li J, Zhang L, Yu S, Luo Z, Su D, Zheng D, Zhou H, Zhu J, Lin X, Luo H, et al. Long-Term Benefits of Cenchrus fungigraminus Residual Roots Improved the Quality and Microbial Diversity of Rhizosphere Sandy Soil through Cellulose Degradation in the Ulan Buh Desert, Northwest China. Plants. 2024; 13(5):708. https://doi.org/10.3390/plants13050708
Chicago/Turabian StyleLi, Jing, Lili Zhang, Shikui Yu, Zongzhi Luo, Dewei Su, Dan Zheng, Hengyu Zhou, Jieyi Zhu, Xingsheng Lin, Hailing Luo, and et al. 2024. "Long-Term Benefits of Cenchrus fungigraminus Residual Roots Improved the Quality and Microbial Diversity of Rhizosphere Sandy Soil through Cellulose Degradation in the Ulan Buh Desert, Northwest China" Plants 13, no. 5: 708. https://doi.org/10.3390/plants13050708
APA StyleLi, J., Zhang, L., Yu, S., Luo, Z., Su, D., Zheng, D., Zhou, H., Zhu, J., Lin, X., Luo, H., Rensing, C., Lin, Z., & Lin, D. (2024). Long-Term Benefits of Cenchrus fungigraminus Residual Roots Improved the Quality and Microbial Diversity of Rhizosphere Sandy Soil through Cellulose Degradation in the Ulan Buh Desert, Northwest China. Plants, 13(5), 708. https://doi.org/10.3390/plants13050708