
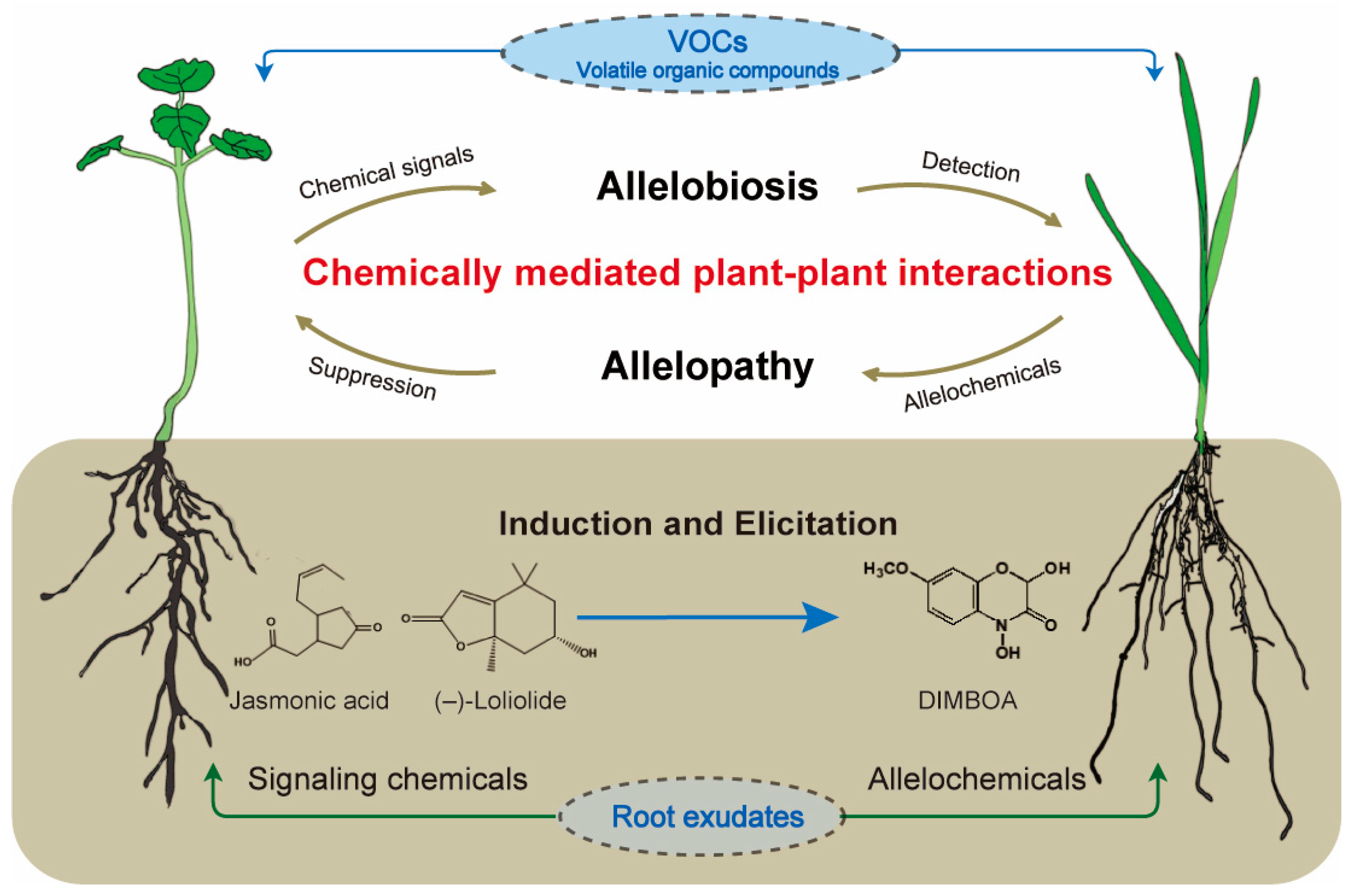
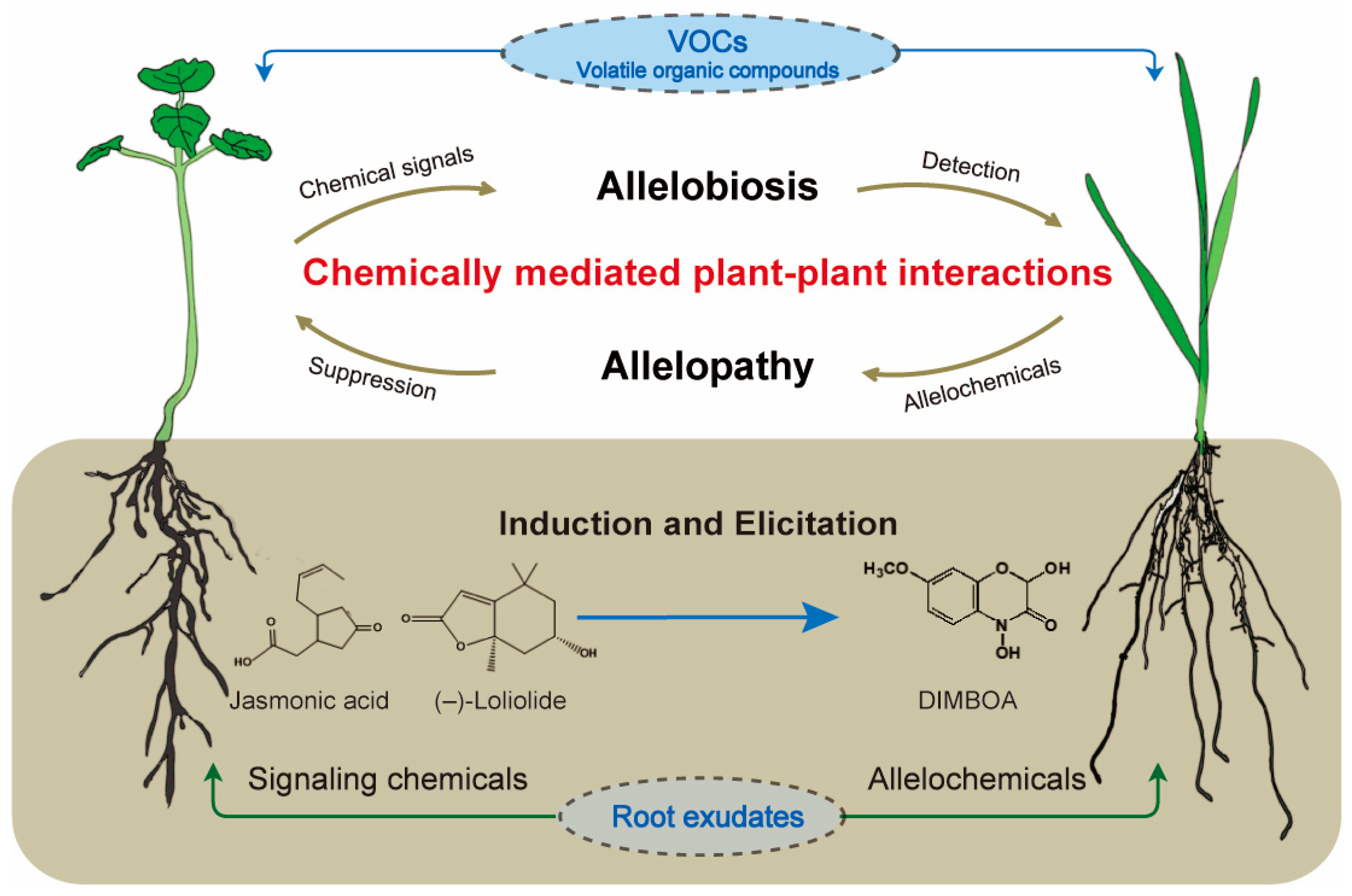
Chemically Mediated Plant–Plant Interactions: Allelopathy and Allelobiosis




Abstract
1. Introduction
2. Allelopathy, Allelochemicals, and Their Application for Sustainable Agriculture and Forestry
2.1. Allelopathy, Competition, and Plant Invasion
2.2. Allelochemical Responses and Interactions
2.3. Allelopathy for Sustainable Agriculture and Forestry
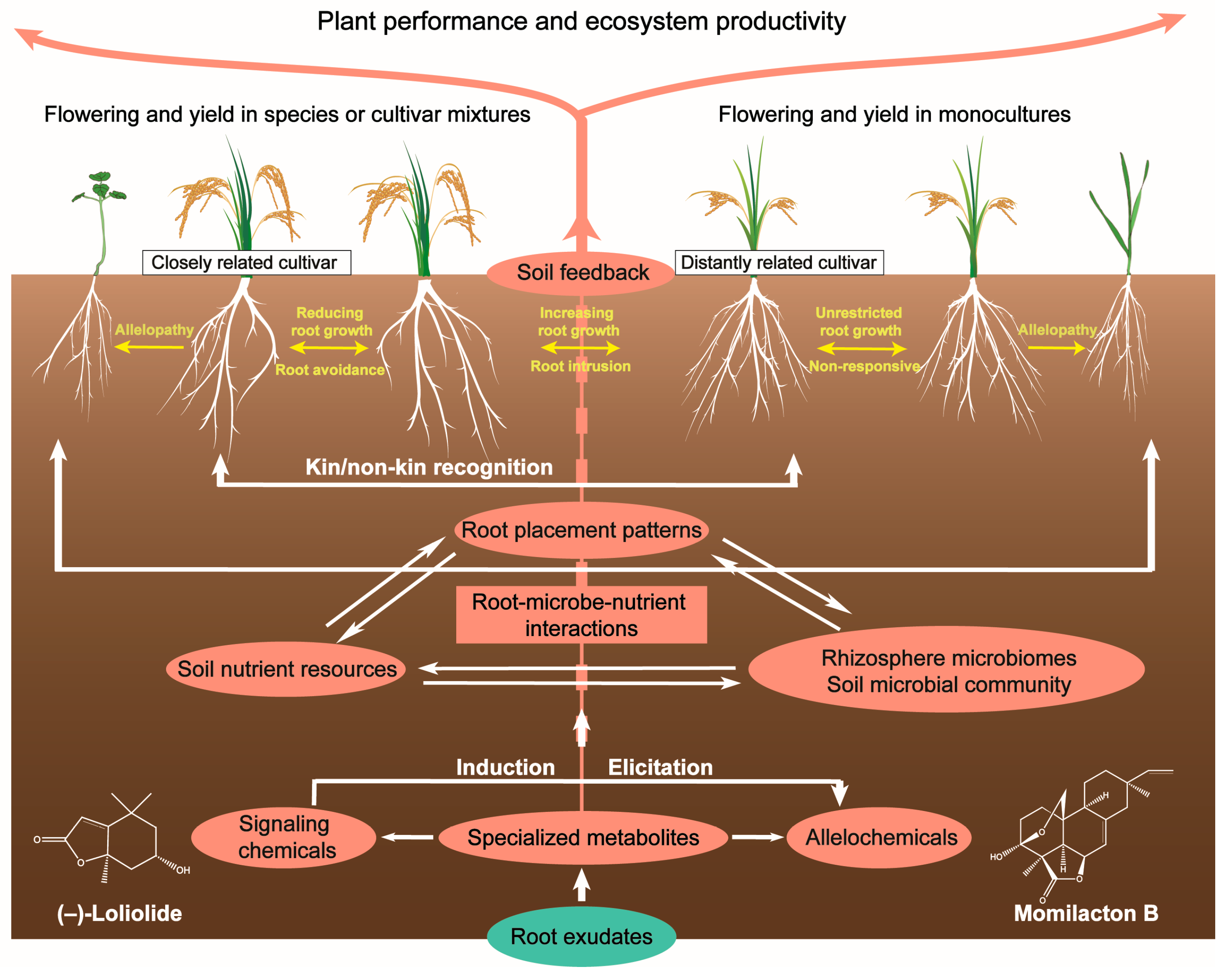
3. Allelobiosis and Plant Identity Recognition
3.1. Allelobiosis and Plant–Plant Signaling Interactions
3.2. Plant Neighbor Detection and Identity Recognition
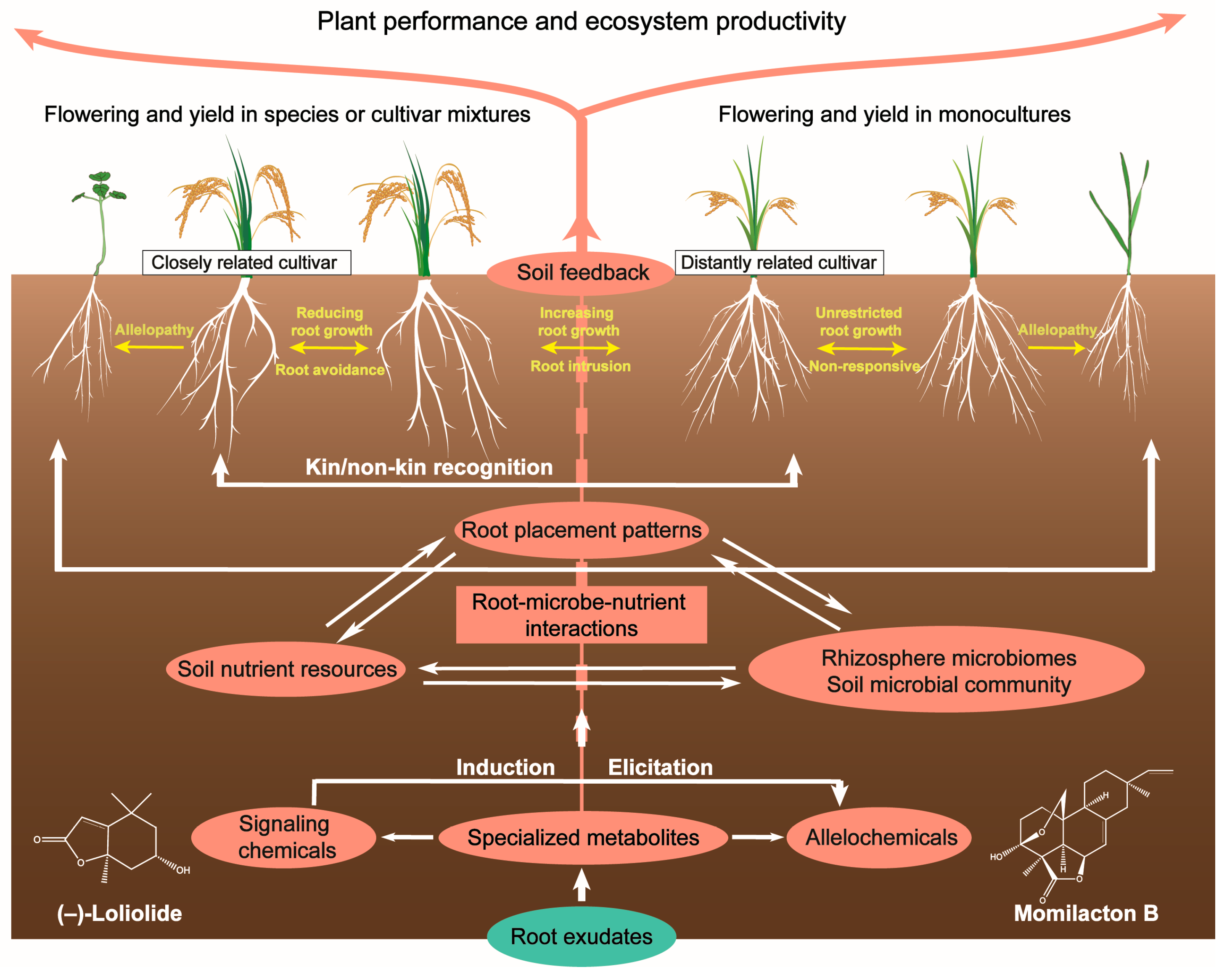
3.3. Relatedness-Mediated Neighbor Recognition
4. Chemically Mediated Root–Soil Interactions and Plant–Soil Feedback
4.1. Root–Root Interactions and Root Placement Patterns
4.2. Soil Mobility and Microbial Interactions
4.3. Belowground and Aboveground Interactions
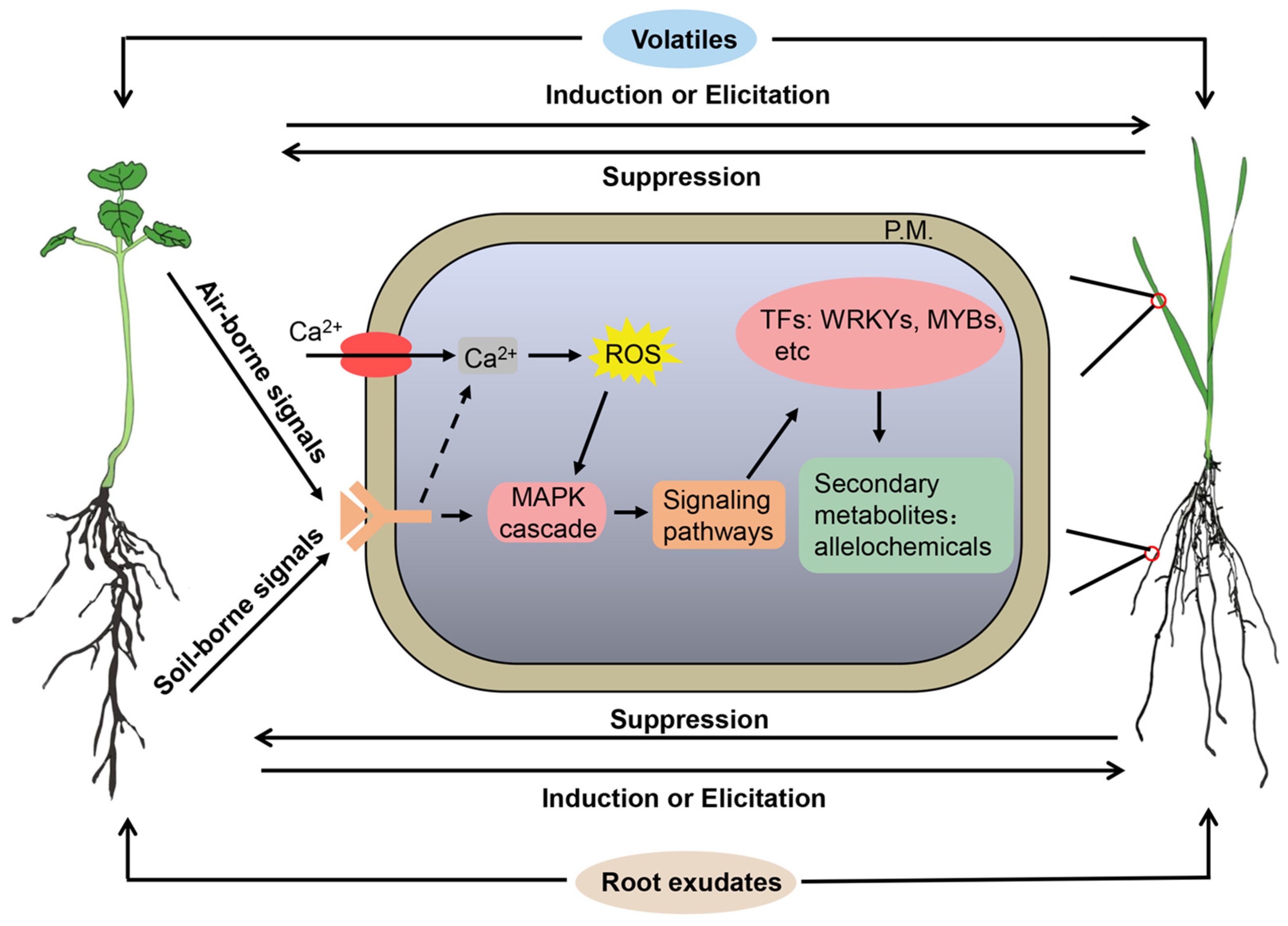
5. Biosynthesis and Molecular Mechanisms of Allelochemicals and Signaling Chemicals
5.1. Biosynthesis and Mechanisms of Allelochemicals
5.2. Biosynthesis and Mechanisms of Signaling Chemicals
5.3. Molecular Mechanisms of Signaling-Induced Allelochemical Responses
6. Perspectives and Outlooks
- Identifying chemical mediation before a plant–plant interaction is considered to be allelopathy or allelobiosis.
- Developing an ecological context for allelopathy and allelobiosis, particularly for a linkage between allelopathy and allelobiosis.
- Deciphering field-based evidence for allelopathy and allelobiosis in plant coexistence and community assembly.
- Detecting and determining spatiotemporal dynamics of allelochemicals and signaling chemicals from the living plants and their environments in situ.
- Interpreting the concept of plant neighbor detection and identity recognition, particularly for relatedness-mediated neighbor detection and identity recognition and their altering of the consequences of intra-specific and inter-specific interactions in species or cultivar mixtures.
- Deciphering root–soil interactions, particularly for chemical mechanisms that structure competitive rhizosphere interactions as well as the rhizosphere microbiome.
- Evaluating allelopathy and allelobiosis under environmental variability and global changes.
- Elucidating molecular mechanisms of allelopathy and allelobiosis in plant–plant interactions from gene expressions, protein receptors, and their transportation.
- Applying ecological approaches to agriculture and forestry by allelopathy with allelochemicals and allelobiosis with signaling chemicals.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zepeda, V.; Martorell, C. Seed mass equalises the strength of positive and negative plant-plant interactions in a semi-arid grassland. Oecologia 2019, 190, 287–296. [Google Scholar] [CrossRef]
- Wuest, S.E.; Peter, R.; Niklaus, P.A. Ecological and evolutionary approaches to improving crop variety mixtures. Nat. Ecol. Evol. 2021, 5, 1068–1077. [Google Scholar] [CrossRef] [PubMed]
- Chesson, P. Mechanisms of maintenance of species diversity. Ann. Rev. Ecol. Evol. Syst. 2000, 31, 343–366. [Google Scholar] [CrossRef]
- Zepeda, V.; Martorell, C. Fluctuation-independent niche differentiation and relative non-linearity drive coexistence in a species-rich grassland. Ecology 2019, 100, e02726. [Google Scholar] [CrossRef] [PubMed]
- Fréville, H.; Roumet, P.; Rode, N.O.; Rocher, A.; Latreille, M.; Muller, M.H. Preferential helping to relatives: A potential mechanism responsible for lower yield of crop variety mixtures? Evol. Appl. 2019, 12, 1837–1849. [Google Scholar] [CrossRef]
- Mazal, L.; Fajardo, A.; Till-Bottraud, I.T.; Corenblit, D.; Fumanal, B. Kin selection, kin recognition and kin discrimination in plants revisited: A claim for considering environmental and genetic variability. Plant Cell Environ. 2023, 46, 2007–2016. [Google Scholar] [CrossRef] [PubMed]
- Baldwin, I.T.; Halitschke, R.; Paschold, A.; von Dahl, C.C.; Preston, C.A. Volatile signaling in plant-plant interactions: “Talking Trees” in the genomics era. Science 2006, 311, 812–815. [Google Scholar] [CrossRef]
- Pierik, R.; Mommer, L.; Voesenek, L.A.C.J. Molecular mechanisms of plant competition: Neighbour detection and response strategies. Funct. Ecol. 2013, 27, 841–853. [Google Scholar] [CrossRef]
- Karban, R. Plant communication. Ann. Rev. Ecol. Evol. Syst. 2021, 52, 1–24. [Google Scholar] [CrossRef]
- Meiners, S.J.; Kong, C.; Ladwig, L.M.; Pisula, N.L.; Lang, K.A. Developing an ecological context for allelopathy. Plant Ecol. 2012, 213, 1221–1227. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathy; Academic Press: New York, NY, USA, 1974. [Google Scholar]
- Pettersson, J.; Ninkovic, V.; Glinwood, R. Plant activation of barley by intercropped conspecifics and weeds: Allelobiosis. Proc. BCPC Int. Congr. Crops Sci. Technol. 2003, 2, 1135–1144. [Google Scholar]
- Ninkovic, V.; Glinwood, R.; Pettersson, J. Communication between undamaged plants by volatiles: The role of allelobiosis. In Communication in Plants; Baluška, F., Mancuso, S., Volkmann, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; pp. 421–434. [Google Scholar]
- Ninkovic, V. Volatile interaction between undamaged plants: A short cut to coexistence. In Plant Communication from an Ecological Perspective; Baluska, F., Ninkovic, V., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 75–86. [Google Scholar]
- Glinwood, R.; Ninkovic, V.; Pettersson, J. Chemical interaction between undamaged plants—Effects on herbivores and natural enemies. Phytochemistry 2011, 72, 1683–1689. [Google Scholar] [CrossRef]
- Li, Y.H.; Xia, Z.C.; Kong, C.H. Allelobiosis in the interference of allelopathic wheat with weeds. Pest Manag. Sci. 2016, 72, 2146–2153. [Google Scholar] [CrossRef] [PubMed]
- Inderjit, S.; Wardle, D.A.; Karban, R.; Callaway, R.M. The ecosystem and evolutionary contexts of allelopathy. Trends Ecol. Evol. 2011, 26, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.H.; Xuan, T.D.; Khanh, T.D.; Tran, H.; Trung, N.T. Allelochemicals and signaling chemicals in plants. Molecules 2019, 24, 2737. [Google Scholar] [CrossRef] [PubMed]
- Macias, F.A.; Mecias, F.J.; Molinillo, J.M. Recent advances in allelopathy for weed control: From knowledge to applications. Pest Manag. Sci. 2019, 75, 2413–2436. [Google Scholar] [CrossRef] [PubMed]
- Hierro, J.L.; Callaway, R.M. The ecological importance of allelopathy. Ann. Rev. Ecol. Evol. Syst. 2021, 52, 25–45. [Google Scholar] [CrossRef]
- Han, M.; Yang, H.; Huang, H.; Du, J.; Zhang, S.; Fu, Y. Allelopathy and allelobiosis: Efficient and economical alternatives in agroecosystems. Plant Biol. 2024, 26, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Chen, X.; Ding, L.; Kong, C.H. Allelopathy and allelochemicals in grasslands and forests. Forests 2023, 14, 562. [Google Scholar] [CrossRef]
- Kong, C.H.; Zhang, S.Z.; Li, Y.H.; Xia, Z.C.; Yang, X.F.; Meiners, S.J.; Wang, P. Plant neighbor detection and allelochemical response are driven by root-secreted signaling chemicals. Nat. Commun. 2018, 9, 3867. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Kohli, R.K. Autotoxicity: Concept, organisms and ecological significance. Crit. Rev. Plant Sci. 1999, 18, 757–772. [Google Scholar] [CrossRef]
- Inderjit; del Moral, R. Is separating resource competition from allelopathy realistic? Bot. Rev. 1997, 63, 221–230. [Google Scholar] [CrossRef]
- Mahé, I.; Chauvel, B.; Colbach, N.; Cordeau, S.; Gfeller, A.; Reiss, A.; Moreau, D. Deciphering field-based evidences for crop allelopathy in weed regulation. A review. Agron. Sustain. Dev. 2022, 42, 50. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Liu, Y.J.; Yuan, L.; Weber, E.; van Kleunen, M. Effect of allelopathy on plant performance: A meta-analysis. Ecol. Lett. 2021, 24, 348–362. [Google Scholar] [CrossRef] [PubMed]
- Soderquist, C.J. Juglone and allelopathy. J. Chem. Educ. 1973, 50, 782. [Google Scholar] [CrossRef]
- Muller, C.H.; Muller, W.H.; Haines, B.L. Volatile growth inhibitors produced by aromatic shrubs. Science 1964, 143, 471–473. [Google Scholar] [CrossRef]
- Fitter, A. Making allelopathy respectable. Science 2003, 301, 1337–1338. [Google Scholar] [CrossRef]
- Callaway, A.; Aschehoug, E.T. Invasive plants versus their new and old neighbors: A mechanism for exotic invasion. Science 2000, 290, 521–523. [Google Scholar] [CrossRef]
- Bais, H.P.; Vepachedu, R.; Gilroy, S.; Callaway, R.M.; Vivanco, J.M. Allelopathy and exotic plant invasion: From molecules and genes to species interactions. Science 2003, 301, 1377–1380. [Google Scholar] [CrossRef]
- Callaway, R.M.; Ridenour, W.M. Novel weapons: Invasive success and the evolution of increased competitive ability. Front. Ecol. Environ. 2004, 2, 436–443. [Google Scholar] [CrossRef]
- Thorpe, A.S.; Thelen, G.C.; Diaconu, A.; Callaway, R.M. Root exudate is allelopathic in invaded community but not in native community: Field evidence for the novel weapons hypothesis. J. Ecol. 2009, 97, 641–645. [Google Scholar] [CrossRef]
- Huang, F.; Lankau, R.; Peng, S. Coexistence via coevolution driven by reduced allelochemical effects and increased tolerance to competition between invasive and native plants. New Phytol. 2018, 218, 357–369. [Google Scholar] [CrossRef]
- Uddin, M.N.; Asaeda, T.; Shampa, S.H.; Robinson, R.W. Allelopathy and its coevolutionary implications between native and non-native neighbors of invasive Cynara cardunculus L. Ecol. Evol. 2020, 10, 7463–7475. [Google Scholar] [CrossRef] [PubMed]
- Jeddi, K.; Fakhfakh, L.M.; Siddique, K.H.M.; Hessini, K.; Chaieb, M. Effect of Acacia saligna (Labill.) Wendl. extracts on seed germination and seedling performance of three native Mediterranean shrubs. Bot. Lett. 2022, 169, 51–60. [Google Scholar] [CrossRef]
- Kim, Y.O.; Lee, E.J. Comparison of phenolic compounds and the effects of invasive and native species in East Asia: Support for the novel weapons hypothesis. Ecol. Res. 2011, 26, 87–94. [Google Scholar] [CrossRef]
- Reinhart, K.O.; Rinella, M. Comparing susceptibility of eastern and western US grasslands to competition and allelopathy from spotted knapweed [Centaurea stoebe L. subsp. micranthos (Gugler) Hayek]. Plant Ecol. 2011, 212, 821–828. [Google Scholar] [CrossRef]
- Levacic, D.; Perkovic, L.; Vukovic, N.; Jelaska, S.D. Bohemian Knotweed Reynoutria x bohemica Chrtek et Chrtkova seems not to rely heavily on allelopathy for its persistence in invaded sites in the Southwest Part of the Zagreb, Croatia. Plants 2023, 12, 2222. [Google Scholar] [CrossRef] [PubMed]
- Lankau, R.A.; Strauss, S.Y. Mutual feedbacks maintain both genetic and species diversity in a plant community. Science 2007, 317, 1561–1563. [Google Scholar] [CrossRef] [PubMed]
- Ehlers, B.K.; Thompson, J. Do co-occurring plant species adapt to one another? The response of Bromus erectus to the presence of different Thymus vulgaris chemotypes. Oecologia 2004, 141, 511–518. [Google Scholar] [CrossRef]
- Metlen, K.L.; Aschehoug, E.T.; Callaway, R.M. Plant behavioural ecology: Dynamic plasticity in secondary metabolites. Plant Cell Environ. 2009, 32, 641–653. [Google Scholar] [CrossRef]
- Broz, A.K.; Broeckling, C.D.; De-La-Peña, C.; Lewis, M.R.; Greene, E.; Callaway, R.M.; Vivanco, S.J.M. Plant neighbour identity influences plant biochemistry and physiology related to defence. BMC Plant Biol. 2010, 10, 115–119. [Google Scholar] [CrossRef]
- Suzuki, R.O.; Suzuki, S.N. Morphological adaptation of a palatable plant to long-term grazing can shift interactions with an unpalatable plant from facilitative to competitive. Plant Ecol. 2012, 213, 175–183. [Google Scholar] [CrossRef]
- Kong, C.H.; Hu, F.; Xu, X.H. Allelopathic potential and chemical constituents of volatiles from Ageratum conyzoides under stress. J. Chem. Ecol. 2002, 28, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Dayan, F.E. Factors modulating the levels of the allelochemical sorgolene in Sorghum bicolor. Planta 2006, 224, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Uesugi, A.; Kessler, A. Herbivore exclusion drives the evolution of plant competitiveness via increased allelopathy. New Phytol. 2013, 198, 916–924. [Google Scholar] [CrossRef]
- Shan, Z.X.; Zhou, S.X.; Shah, A.S.; Arafat, Y.; Rizvi, S.A.H.; Shao, H. Plant allelopathy in response to biotic and abiotic factors. Agronomy 2023, 13, 2358. [Google Scholar] [CrossRef]
- Bilas, R.D.; Bretman, A.; Bennett, T. Friends, neighbours and enemies: An overview of the communal and social biology of plants. Plant Cell Environ. 2021, 44, 997–1013. [Google Scholar] [CrossRef]
- Kato-Noguchi, H. Barnyard grass-induced rice allelopathy and momilactone B. J. Plant Physiol. 2011, 168, 1016–1020. [Google Scholar] [CrossRef]
- Rasher, D.B.; Hay, M.E. Competition induces allelopathy but suppresses growth and anti-herbivore defence in a chemically rich seaweed. Proc. R. Soc. B 2014, 281, 20132615. [Google Scholar] [CrossRef]
- Lankau, R.A.; Kliebenstein, D.J. Competition, herbivory and genetics interact to determine the accumulation and fitness consequences of a defence metabolite. J. Ecol. 2009, 97, 78–88. [Google Scholar] [CrossRef]
- Rivoal, A.; Fernandez, C.; Greff, S.; Montes, N.; Vila, B. Does competition stress decrease allelopathic potential? Biochem. Syst. Ecol. 2011, 39, 401–407. [Google Scholar] [CrossRef]
- Wasternack, C. A plant’s balance of growth and defense—Revisited. New Phytol. 2017, 215, 1291–1294. [Google Scholar] [CrossRef]
- Niemeyer, H.M. Hydroxamic acids derived from 2-hydroxy-2H-1,4-benzoxazin-3(4H)-one: Key defense chemicals of cereals. J. Agric. Food Chem. 2009, 57, 1677–1696. [Google Scholar] [CrossRef]
- Zhang, S.Z.; Li, Y.H.; Kong, C.H.; Xu, X.H. Interference of allelopathic wheat with different weeds. Pest Manag. Sci. 2016, 72, 172–178. [Google Scholar] [CrossRef]
- Uesugi, A.; Johnson, R.; Kessler, A. Context-dependent induction of allelopathy in plants under competition. Oikos 2019, 128, 1492–1502. [Google Scholar] [CrossRef]
- Xia, Z.C.; Kong, C.H.; Chen, L.C.; Wang, P.; Wang, S.L. A broadleaf species enhances an autotoxic conifers growth through belowground chemical interactions. Ecology 2016, 97, 2283–2292. [Google Scholar] [CrossRef]
- Yang, X.F.; Kong, C.H. Interference of allelopathic rice with paddy weeds at the root level. Plant Biol. 2017, 19, 584–591. [Google Scholar] [CrossRef]
- Asaduzzaman, M.; An, M.; Pratley, J.E.; Luckett, D.J.; Lemerle, D.; Coombes, N. The seedling root response of annual ryegrass (Lolium rigidum) to neighbouring seedlings of a highly-allelopathic canola (Brassica napus). Flora 2016, 219, 18–24. [Google Scholar] [CrossRef]
- Yan, S.B.; Wang, P. Effects of alleolchemicals on morphological traits of roots: A meta-analysis. Chin. J. Appl. Ecol. 2020, 31, 2168–2174. [Google Scholar]
- Wang, C.Y.; Li, L.L.; Meiners, S.J.; Kong, C.H. Root placement patterns in allelopathic plant-plant interactions. New Phytol. 2023, 237, 563–575. [Google Scholar] [CrossRef]
- Kato-Noguchi, H.; Ino, T. The chemical-mediated allelopathic interaction between rice and barnyard grass. Plant Soil 2013, 370, 267–275. [Google Scholar] [CrossRef]
- Tabaglio, V.; Gavazzi, C.; Schulz, M.; Marocco, A. Alternative weed control using the allelopathic effect of natural benzoxazinoids from rye mulch. Agron. Sustain. Dev. 2008, 28, 397–401. [Google Scholar] [CrossRef]
- Kong, C.H.; Hu, F.; Wang, P.; Wu, J.L. Effect of allelopathic rice varieties combined with cultural management options on paddy field weeds. Pest Manag. Sci. 2008, 64, 276–282. [Google Scholar] [CrossRef]
- Macias, F.A.; Chinchilla, N.; Varela, R.M.; Molinillo, J.M.G.; Marin, D.; de Siqueira, J.M. Aromatic-ring-functionalised benzoxazinones in the system Oryza sativa–Echinochloa crus-galli as biorational herbicide models. Pest Manag. Sci. 2009, 65, 1104–1113. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.H.; Chen, X.H.; Hu, F.; Zhang, S.Z. Breeding of commercially acceptable allelopathic rice cultivars in China. Pest Manag. Sci. 2011, 67, 1100–1106. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.H.; Wang, P.; Xu, X.H. Allelopathic interference of Ambrosia trifida with wheat (Triticum aestivum). Agric. Ecosyst. Environ. 2007, 119, 416–420. [Google Scholar] [CrossRef]
- Williams, M.M.; Masiunas, J.B. Functional relationships between giant ragweed (Ambrosia trifida) interference and sweet corn yield and ear traits. Weed Sci. 2006, 54, 948–953. [Google Scholar] [CrossRef]
- Kong, C.H.; Hu, F.; Xu, X.H.; Zhang, M.X.; Liang, W.J. Volatile allelochemicals in the Ageratum conyzoides intercropped citrus orchard and their effects on mites Amblyseius newsami and Panonychus citri. J. Chem. Ecol. 2005, 31, 2193–2203. [Google Scholar] [CrossRef]
- Kong, C.H. Ecological pest management and control by using allelopathic weeds (Ageratum conyzoides, Ambrosia trifida and Lantana camara) and their allelochemicals in China. Weed Biol. Manag. 2010, 10, 73–80. [Google Scholar] [CrossRef]
- Zhao, H.H.; Kong, C.H.; Xu, X.H. Herbicidal efficacy and ecological safety of an allelochemical-based benzothiazine derivative. Pest Manag. Sci. 2019, 75, 2690–2697. [Google Scholar] [CrossRef]
- Fernandez, C.; Voiriot, S.; Mevy, J.P.; Vila, B.; Ormeno, E.; Dupouyet, S.; Bousquet-Melou, A. Regeneration failure of Pinus halepensis Mill.: The role of autotoxicity and some abiotic environmental parameters. For. Ecol. Manag. 2008, 255, 2928–2936. [Google Scholar] [CrossRef]
- Kong, C.H.; Chen, L.C.; Xu, X.H.; Wang, P.; Wang, S.L. Allelochemicals and activities in replanted Chinese fir (Cunninghamia lanceolata (Lamb.) Hook.) tree ecosystem. J. Agric. Food Chem. 2008, 56, 11734–11739. [Google Scholar] [CrossRef]
- Chen, L.C.; Wang, S.L.; Wang, P.; Kong, C.H. Autoinhibition and soil allelochemical (cyclic dipeptide) levels in replanted Chinese fir (Cunninghamia lanceolata) plantations. Plant Soil 2014, 374, 793–801. [Google Scholar] [CrossRef]
- Xia, Z.C.; Kong, C.H.; Chen, L.C.; Wang, S.L. Allelochemical-mediated soil microbial community in long-term monospecific Chinese fir forest plantations. Appl. Soil Ecol. 2015, 96, 52–59. [Google Scholar] [CrossRef]
- Williams, R.A. Mitigating biodiversity concerns in Eucalyptus plantations located in South China. J. Biosci. Med. 2015, 3, 1. [Google Scholar] [CrossRef]
- Zhang, C.; Li, X.; Chen, Y.; Zhao, J.; Wan, S.; Lin, Y.; Fu, S. Effects of Eucalyptus litter and roots on the establishment of native tree species in Eucalyptus plantations in South China. For. Ecol. Manag. 2016, 375, 76–83. [Google Scholar] [CrossRef]
- Qin, F.; Liu, S.; Yu, S. Effects of allelopathy and competition for water and nutrients on survival and growth of tree species in Eucalyptus urophylla plantations. For. Ecol. Manag. 2018, 424, 387–395. [Google Scholar] [CrossRef]
- Yang, L.X.; Wang, P.; Kong, C.H. Effect of larch (Larix gmelini Rupr.) root exudates on Manchurian walnut (Juglans mandshurica Maxim.) growth and soil juglone in mixed-species plantation. Plant Soil 2010, 329, 249–258. [Google Scholar] [CrossRef]
- Forrester, D.I.; Bauhus, J.; Cowie, A.L.; Vanclay, J.K. Mixed-species plantations of Eucalyptus with nitrogen-fixing trees: A review. For. Ecol. Manag. 2006, 233, 211–230. [Google Scholar] [CrossRef]
- Bever, J.D.; Dickie, I.A.; Facelli, E.; Facelli, J.M.; Klironomos, J.; Moora, M.; Rillig, M.C.; Stock, W.D.; Tibbett, M.; Zobel, M. Rooting theories of plant community ecology in microbial interactions. Trends Ecol. Evol. 2010, 25, 468–478. [Google Scholar] [CrossRef]
- Wang, N.Q.; Kong, C.H.; Wang, P.; Meiners, S.J. Root exudate signals in plant-plant interactions. Plant Cell Environ. 2021, 44, 1044–1058. [Google Scholar] [CrossRef]
- Loreto, F.; D’Auria, S. How do plants sense volatiles sent by other plants? Trends Plant Sci. 2022, 27, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Berens, M.L.; Berry, H.M.; Mine, A.; Argueso, C.T.; Tsuda, K. Evolution of hormone signaling networks in plant defense. Annu. Rev. Phytopathol. 2017, 55, 401–425. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.F.; Li, L.L.; Xu, Y.; Kong, C.H. Kin recognition in rice (Oryza sativa L.) lines. New Phytol. 2018, 220, 567–578. [Google Scholar] [CrossRef]
- Karban, R.; Yang, L.H.; Edwards, K.F. Volatile communication between plants that affects herbivory: A meta-analysis. Ecol. Lett. 2014, 17, 44–52. [Google Scholar] [CrossRef]
- Ninkovic, V.; Markovic, D.; Rensing, M. Plant volatiles as cues and signals in plant communication. Plant Cell Environ. 2021, 44, 1030–1043. [Google Scholar] [CrossRef] [PubMed]
- Brosset, A.; Blande, J.D. Volatile-mediated plant-plant interactions: Volatile organic compounds as modulators of receiver plant defence, growth, and reproduction. J. Exp. Bot. 2022, 73, 511–528. [Google Scholar] [CrossRef]
- Chen, B.J.W.; During, H.J.; Anten, N.P.R. Detect thy neighbor: Identity recognition at the root level in plants. Plant Sci. 2012, 195, 157–167. [Google Scholar] [CrossRef]
- Yoneyama, K.; Bennett, T. Whispers in the dark: Signals regulating underground plant–plant interactions. Curr. Opin. Plant Biol. 2024, 77, 102456. [Google Scholar] [CrossRef]
- Maag, D.; Erb, M.; Koellner, T.G.; Gershenzon, J. Defensive weapons and defense signals in plants: Some metabolites serve both roles. Bioassays 2015, 37, 167–174. [Google Scholar] [CrossRef]
- Rasmann, S.; Turlings, T.C.J. Root signals that mediate mutualistic interactions in the rhizosphere. Curr. Opin. Plant Biol. 2016, 32, 62–68. [Google Scholar] [CrossRef]
- Huber, M.; Nieuwendijk, N.M.; Pantazopoulou, C.K.; Pierik, R. Light signalling shapes plant–plant interactions in dense canopies. Plant Cell Environ. 2021, 44, 1014–1029. [Google Scholar] [CrossRef]
- Berger, K.; Machwitz, M.; Kycko, M.; Kefauver, S.C.; Van Wittenberghe, S.; Gerhards, M.; Verrelst, J.; Atzberger, C.; van der Tol, C.; Damm, A. Multi-sensor spectral synergies for crop stress detection and monitoring in the optical domain: A review. Remote Sens. Environ. 2022, 280, 113198. [Google Scholar] [CrossRef]
- Zeng, Y.; Chen, M.; Hao, D.; Damm, A.; Badgley, G.; Rascher, U.; Johnson, J.E.; Dechant, B.; Siegmann, B.; Ryu, Y. Combining near-infrared radiance of vegetation and fluorescence spectroscopy to detect effects of abiotic changes and stresses. Remote Sens. Environ. 2022, 270, 112856. [Google Scholar] [CrossRef]
- Eskelinen, A.; Harpole, W.S.; Jessen, M.; Virtanen, R.; Hautier, Y. Light competition drives herbivore and nutrient effects on plant diversity. Nature 2022, 611, 301–305. [Google Scholar] [CrossRef]
- Li, H.; Burgie, E.S.; Gannam, Z.T.; Li, H.; Vierstra, R.D. Plant phytochrome B is an asymmetric dimer with unique signalling potential. Nature 2022, 604, 127–133. [Google Scholar] [CrossRef]
- Casal, J.J. Photoreceptor signaling networks in plant responses to shade. Ann. Rev. Plant Biol. 2013, 64, 403–427. [Google Scholar] [CrossRef]
- Keller, M.M.; Jaillais, Y.; Pedmale, U.V.; Moreno, J.E.; Chory, J.; Ballaré, C.L. Cryptochrome 1 and phytochrome B control shade-avoidance responses in Arabidopsis via partially independent hormonal cascades. Plant J. 2011, 67, 195–207. [Google Scholar] [CrossRef]
- Farmer, E.E. Surface-to-air signals. Nature 2001, 411, 854–856. [Google Scholar] [CrossRef]
- Farmer, E.E.; Ryan, C.A. Interplant communication: Airborne methyl jasmonate induces synthesis of proteinase inhibitors in plant leaves. Proc. Natl. Acad. Sci. USA 1990, 87, 7713–7716. [Google Scholar] [CrossRef]
- Kessler, A.; Baldwin, I.T. Defensive function of herbivore-induced plant volatile emissions in nature. Science 2001, 291, 2141–2144. [Google Scholar] [CrossRef]
- Shulaev, V.; Silverman, P.; Raskin, I. Airborne signalling by methyl salicylate in plant pathogen resistance. Nature 1997, 385, 718–721. [Google Scholar] [CrossRef]
- Ameye, M.; Van Meulebroek, L.; Meuninck, B.; Vanhaecke, L.; Smagghe, G.; Haesaert, G.; Audenaert, K. Metabolomics reveal induction of ROS production and glycosylation events in wheat upon exposure to the green leaf volatile Z-3-hexenyl acetate. Front. Plant Sci. 2020, 11, 596271. [Google Scholar] [CrossRef]
- Hirao, T.; Okazawa, A.; Harada, K.; Kobayashi, A.; Muranaka, T.; Hirata, K. Green leaf volatiles enhance methyl jasmonate response in Arabidopsis. J. Biosci. Bioeng. 2012, 114, 540–545. [Google Scholar] [CrossRef]
- Engelberth, J.; Contreras, C.F.; Dalvi, C.; Li, T.; Engelberth, M. Early transcriptome analyses of Z-3-hexenol-treated Zea mays revealed distinct transcriptional networks and anti-herbivore defense potential of green leaf volatiles. PLoS ONE 2013, 8, e77465. [Google Scholar] [CrossRef]
- Frank, L.; Wenig, M.; Ghirardo, A.; van der Krol, A.; Vlot, A.C.; Schnitzler, J.P.; Rosenkranz, M. Isoprene and β-caryophyllene confer plant resistance via different plant internal signalling pathways. Plant Cell Environ. 2021, 44, 1151–1164. [Google Scholar] [CrossRef]
- Godard, K.A.; White, R.; Bohlmann, J. Monoterpene-induced molecular responses in Arabidopsis thaliana. Phytochemistry 2008, 69, 1838–1849. [Google Scholar] [CrossRef]
- Jing, T.T.; Du, W.K.; Gao, T.; Wu, Y.; Zhang, N.; Zhao, M.Y.; Jin, J.Y.; Wang, J.M.; Schwab, W.; Wan, X.C. Herbivore-induced DMNT catalyzed by CYP82D47 plays an important role in the induction of JA-dependent herbivore resistance of neighboring tea plants. Plant Cell Environ. 2020, 44, 1178–1191. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.L.; Zhang, L.P.; Cai, X.M.; Li, X.; Bian, L.; Luo, Z.X.; Li, Z.Q.; Chen, Z.M.; Xin, Z.J. (E)-Nerolidol is a volatile signal that induces defenses against insects and pathogens in tea plants. Hortic. Res. 2020, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- Arimura, G.; Ozawa, R.; Shimoda, T.; Nishioka, T.; Boland, W.; Takabayashi, J. Herbivory-induced volatiles elicit defence genes in lima bean leaves. Nature 2000, 406, 512–515. [Google Scholar] [CrossRef]
- Horiuchi, J.; Badri, D.V.; Kimball, B.A.; Negre, F.; Dudareva, N.; Paschke, M.W.; Vivanco, J.M. The floral volatile, methyl benzoate, from snapdragon (Antirrhinum majus) triggers phytotoxic effects in Arabidopsis thaliana. Planta 2007, 226, 1–10. [Google Scholar] [CrossRef]
- Erb, M.; Veyrat, N.; Robert, C.A.; Xu, H.; Frey, M.; Ton, J.; Turlings, T.C. Indole is an essential herbivore-induced volatile priming signal in maize. Nat. Commun. 2015, 6, 6273. [Google Scholar] [CrossRef]
- Pierik, R.; Visser, E.J.W.; De Kroon, H.; Voesenek, L.A.C.J. Ethylene is required in tobacco to successfully compete with proximate neighbours. Plant Cell Environ. 2003, 26, 1229–1234. [Google Scholar] [CrossRef]
- Pierik, R.; Whitelam, G.C.; Voesenek, L.A.C.J.; De Kroon, H.; Visser, E.J.W. Canopy studies on ethylene-insensitive tobacco identify ethylene as a novel element in blue light and plant–plant signalling. Plant J. 2004, 38, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Pandey, B.K.; Huang, G.Q.; Bhosale, R.; Sjon, H.; Sturrock, C.J.; Jose, L.; Martin, O.C.; Karady, M.; Voesenek, L.A.C.J.; Ljung, K.; et al. Plant roots sense soil compaction through restricted ethylene diffusion. Science 2021, 371, 276–280. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Wang, P.; Kong, C.H. The levels of jasmonic acid and salicylic acid in a rice-barnyardgrass coexistence system and their relation to rice allelochemicals. Biochem. Syst. Ecol. 2011, 39, 491–497. [Google Scholar] [CrossRef]
- Zhang, P.F.; Wang, X.R.; Lu, Q.; Zhang, H.W.; Chen, J.T.; Zhang, H.; Wang, Y.B.; Li, C.F. Allantoin, a purine metabolite, confers saline–alkaline tolerance to sugar beet by triggering a self-amplifying feedback loop comprising jasmonic acid and nitric oxide. Environ. Exp. Bot. 2023, 206, 105172. [Google Scholar] [CrossRef]
- Murata, M.; Nakai, Y.; Kawazu, K.; Ishizaka, M.; Kajiwara, H.; Abe, H.; Takeuchi, K.; Ichinose, Y.; Mitsuhara, I.; Mochizuki, A.; et al. Loliolide, a carotenoid metabolite, is a potential endogenous inducer of herbivore resistance. Plant Physiol. 2019, 179, 1822–1833. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Zhao, H.H.; Li, H.Y.; Yang, X.F.; Kong, C.H. Kin recognition in an herbicide-resistant barnyardgrass (Echinochloa crus-galli L.) biotype. Plants 2023, 12, 1498. [Google Scholar] [CrossRef]
- Li, F.L.; Chen, X.; Luo, H.M.; Meiners, S.J.; Kong, C.H. Root-secreted (–)-loliolide modulates both belowground defense and aboveground flowering in Arabidopsis and tobacco. J. Exp. Bot. 2023, 74, 964–975. [Google Scholar] [CrossRef]
- Chen, X.; Li, F.L.; Kong, C.H. Rhizosphere bacteria mediate flowering time of two genotypes of Arabidopsis with and without root-secreted signaling (–)-loliolide. Rhizosphere 2023, 27, 100774. [Google Scholar] [CrossRef]
- Akiyama, K.; Ogasawara, S.; Ito, S.; Hayashi, H. Structural requirements of strigolactones for hyphal branching in AM fungi. Plant Cell Physiol. 2010, 51, 1104–1117. [Google Scholar] [CrossRef]
- Akiyama, K.; Hayashi, H. Strigolactones: Chemical signals for fungal symbionts and parasitic weeds in plant roots. Ann. Bot. 2006, 97, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Peters, N.K.; Frost, J.W.; Long, S.R. A plant flavone, luteolin, induces expression of Rhizobium meliloti nodulation genes. Science 1986, 233, 977–980. [Google Scholar] [CrossRef]
- Semchenko, M.; Saar, S.; Lepik, A. Plant root exudates mediate neighbour recognition and trigger complex behavioural changes. New Phytol. 2014, 204, 631–637. [Google Scholar] [CrossRef]
- Pickles, B.J.; Wilhelm, R.; Asay, A.K.; Hahn, A.S.; Simard, S.W.; Mohn, W.W. Transfer of 13C between paired Douglas-fir seedlings reveals plant kinship effects and uptake of exudates by ectomycorrhizas. New Phytol. 2017, 214, 400–411. [Google Scholar] [CrossRef] [PubMed]
- Li, L.L.; Zhao, H.H.; Kong, C.H. (–)-Loliolide, the most ubiquitous lactone, is involved in barnyardgrass-induced rice allelopathy. J. Exp. Bot. 2020, 71, 1540–1550. [Google Scholar] [CrossRef] [PubMed]
- Schreiner, M.; Krumbein, A.; Knorr, D.; Smetanska, I. Enhanced glucosinolates in root exudates of Brassica rapa ssp. rapa mediated by salicylic acid and methyl jasmonate. J. Agric. Food Chem. 2011, 59, 1400–1405. [Google Scholar] [CrossRef]
- Li, L.L.; Li, Z.; Lou, Y.G.; Meiners, S.J.; Kong, C.H. (–)-Loliolide is a general signal of plant stress that activates jasmonate-related responses. New Phytol. 2023, 238, 2099–2112. [Google Scholar] [CrossRef] [PubMed]
- Frost, C.J. Information potential of an ubiquitous phytochemical cue. New Phytol. 2023, 238, 1749–1751. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Y.; Guo, Y.; Jin, B.Y.; Yang, X.F.; Kong, C.H. Phytochemical cue for the fitness cost of herbicide-resistant weeds. Plants 2023, 12, 3158. [Google Scholar] [CrossRef] [PubMed]
- Shen, G.J.; Zhang, J.X.; Lei, Y.T.; Xu, Y.X.; Wu, J.Q. Between-plant signaling. Annu. Rev. Plant Biol. 2023, 74, 367–386. [Google Scholar] [CrossRef] [PubMed]
- Lanfranco, L.; Fiorilli, V.; Gutjahr, C. Partner communication and role of nutrients in the arbuscular mycorrhizal symbiosis. New Phytol. 2018, 220, 1031–1046. [Google Scholar] [CrossRef] [PubMed]
- Ninkovic, V.; Rensing, M.; Dahlin, I.; Markovic, D. Who is my neighbor? Volatile cues in plant interactions. Plant Signal. Behav. 2019, 14, 1634993. [Google Scholar] [CrossRef]
- Patni, B.; Bhattacharyya, M.; Pokhriyal, A. The role of signaling compounds in enhancing rice allelochemicals for sustainable agriculture: An overview. Planta 2023, 258, 90. [Google Scholar] [CrossRef] [PubMed]
- McCormick, A.C.; Effah, E.; Najar-Rodriguez, A. Ecological aspects of volatile organic compounds emitted by exotic invasive plants. Front. Ecol. Evol. 2023, 11, 1059125. [Google Scholar] [CrossRef]
- Chen, K.J.; Zheng, Y.Q.; Kong, C.H.; Zhang, S.Z.; Li, J.; Liu, X.G. 2, 4-Dihydroxy-7- methoxy-1,4-benzoxazin-3-one (DIMBOA) and 6-methoxy-benzoxazolin-2-one (MBOA) levels in the wheat rhizosphere and their effect on soil microbial community structure. J. Agric. Food Chem. 2010, 58, 12710–12716. [Google Scholar] [CrossRef]
- Li, B.; Li, Y.Y.; Wu, H.M.; Zhang, F.F.; Li, C.J.; Li, X.X.; Lambers, H.; Li, L. Root exudates drive interspecific facilitation by enhancing nodulation and N2 fixation. Proc. Natl. Acad. Sci. USA 2016, 113, 6496–6501. [Google Scholar] [CrossRef]
- Xu, Y.; Cheng, H.F.; Kong, C.H.; Meiners, S.J. Intra-specific kin recognition contributes to inter-specific allelopathy: A case study of allelopathic rice interference with paddy weeds. Plant Cell Environ. 2021, 44, 3709–3721. [Google Scholar] [CrossRef]
- Falik, O.; Reides, P.; Gersani, M.; Novoplansky, A. Self/non-self discrimination in roots. J. Ecol. 2003, 91, 525–531. [Google Scholar] [CrossRef]
- Gruntman, M.; Novoplansky, A. Physiologically mediated self/non-self discrimination in roots. Proc. Natl. Acad. Sci. USA 2004, 101, 3863–3867. [Google Scholar] [CrossRef]
- Mahall, B.E.; Callaway, R.M. Root communication mechanisms and intracommunity distributions of 2 Mojave desert shrubs. Ecology 1992, 73, 2145–2151. [Google Scholar] [CrossRef]
- Gersani, M.; Brown, J.S.; O’Brien, E.E.; Maina, G.M.; Abramsky, Z. Tragedy of the commons as a result of root competition. J. Ecol. 2001, 89, 660–669. [Google Scholar] [CrossRef]
- Maina, G.G.; Brown, J.S.; Gersani, M. Intra-plant versus inter-plant root competition in beans: Avoidance, resource matching or tragedy of the commons. Plant Ecol. 2002, 160, 235–247. [Google Scholar] [CrossRef]
- O’Brien, E.E.; Gersani, M.; Brown, J.S. Root proliferation and seed yield in response to spatial heterogeneity of below-ground competition. New Phytol. 2005, 168, 401–412. [Google Scholar] [CrossRef]
- Falik, O.; de Kroon, H.; Novoplansky, A. Physiologically-mediated self/non-self root discrimination in Trifolium repens has mixed effects on plant performance. Plant Signal. Behav. 2006, 1, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Anten, N.P.R.; Chen, B.J.W. Detect thy family: Mechanisms, ecology, and agricultural aspects of kin recognition in plants. Plant Cell Environ. 2021, 44, 1059–1071. [Google Scholar] [CrossRef] [PubMed]
- Dudley, S.A.; File, A.L. Kin recognition in an annual plant. Biol. Lett. 2007, 3, 435–438. [Google Scholar] [CrossRef]
- Torices, R.; DeSoto, L.; Narbona, E.; Gómez, J.M.; Pannell, J.R. Effects of the relatedness of neighbours on floral colour. Front. Ecol. Evol. 2021, 9, 589781. [Google Scholar] [CrossRef]
- Lehmann, L.; Perrin, N. Altruism, dispersal, and phenotype-matching kin recognition. Am. Nat. 2002, 159, 451–468. [Google Scholar] [CrossRef]
- Nakamura, R.R. Plant kin selection. Evol. Theory 1980, 5, 113–117. [Google Scholar]
- Lepik, A.; Abakumova, M.; Zobel, K.; Semchenko, M. Kin recognition is density-dependent and uncommon among temperate grassland plants. Funct. Ecol. 2012, 26, 1214–1220. [Google Scholar] [CrossRef]
- Pezzola, E.; Pandolfi, C.; Mancuso, S. Resource availability affects kin selection in two cultivars of Pisum sativum. Plant Growth Regul. 2020, 90, 321–329. [Google Scholar] [CrossRef]
- Dudley, S.A.; Murphy, G.P.; File, A.L. Kin recognition and competition in plants. Funct. Ecol. 2013, 27, 898–906. [Google Scholar] [CrossRef]
- Murphy, G.P.; Swanton, C.J.; van Acker, R.C.; Dudley, S.A. Kin recognition, multilevel selection and altruism in crop sustainability. J. Ecol. 2017, 105, 930–934. [Google Scholar] [CrossRef]
- Bhatt, V.M.; Khandelwal, A.; Dudley, S.A. Kin recognition, not competitive interactions predicts root allocation in young Cakile edentula seeding pairs. New Phytol. 2011, 189, 1135–1142. [Google Scholar] [CrossRef]
- Belter, P.R.; Cahill, J.F., Jr. Disentangling root system responses to neighbours: Identification of novel root behavioural strategies. AoB Plants 2015, 7, plv059. [Google Scholar] [CrossRef]
- Biedrzycki, M.L.; Bais, H.P. Kin recognition in plants: Did we learn anything from roots? Front. Ecol. Evol. 2022, 9, 785019. [Google Scholar] [CrossRef]
- Pereira, L.; Sadras, V.O.; Batista, W.; Casal, J.J.; Hall, A.J. Light mediated self-organization of sunflower stands increases oil yield in the field. Proc. Natl. Acad. Sci. USA 2017, 114, 7975–7980. [Google Scholar] [CrossRef] [PubMed]
- Simonsen, A.K.; Chow, T.; Stinchcombe, J.R. Reduced plant competition among kin can be explained by Jensen’s inequality. Ecol. Evol. 2014, 4, 4454–4466. [Google Scholar] [CrossRef] [PubMed]
- Goddard, E.L.; Varga, S.; John, E.A.; Soulsbury, C.D. Evidence for root kin recognition in the clonal plant species Glechoma hederacea. Front. Ecol. Evol. 2020, 8, 578141. [Google Scholar] [CrossRef]
- Biedrzycki, M.L.; Jilany, T.A.; Dudley, S.A.; Bais, H.P. Root exudates mediate kin recognition in plants. Commun. Integr. Biol. 2010, 3, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.N.; Pan, Z.C.; Zhu, W.; Wu, E.J.; He, D.C.; Yuan, X.; Qin, Y.Y.; Wang, Y.; Chen, R.S.; Thrall, P.H.; et al. Enhanced agricultural sustainability through within-species diversification. Nat. Sustain. 2019, 2, 46–52. [Google Scholar] [CrossRef]
- Ryan, M.R. Crops better when grown together. Nat. Sustain. 2021, 4, 926–927. [Google Scholar] [CrossRef]
- Yu, R.P.; Lambers, H.; Callaway, R.M.; Wright, A.J.; Li, L. Belowground facilitation and trait matching: Two or three to tango? Trends Plant Sci. 2021, 26, 1227–1235. [Google Scholar] [CrossRef]
- Bawa, K.S. Kin selection and the evolution of plant reproductive traits. Proc. R. Soc. B 2016, 283, 20160789. [Google Scholar] [CrossRef]
- Callaway, R.M.; Li, L. Decisions, decisions, decisions: Plant roots detect and respond to complex environmental cues. New Phytol. 2020, 226, 11–12. [Google Scholar] [CrossRef]
- Hodge, A. Root decisions. Plant Cell Environ. 2009, 32, 628–640. [Google Scholar] [CrossRef]
- Falik, O.; Reides, P.; Gersani, M.; Novoplansky, A. Root navigation by self inhibition. Plant Cell Environ. 2005, 28, 562–569. [Google Scholar] [CrossRef]
- Novoplansky, A. What plant roots know? Semin. Cell Dev. Biol. 2019, 92, 126–133. [Google Scholar] [CrossRef]
- Semchenko, M.; John, E.A.; Hutchings, M.J. Effects of physical connection and genetic identity of neighbouring ramets on root placement patterns in two clonal species. New Phytol. 2007, 176, 644–654. [Google Scholar] [CrossRef]
- Korenblum, E.; Dong, Y.H.; Szymanski, J.; Panda, S.; Jozwiak, A.; Massalha, H.; Meir, S.; Rogachev, I.; Aharoni, A. Rhizosphere microbiome mediates systemic root metabolite exudation by root-to-root signaling. Proc. Natl. Acad. Sci. USA 2020, 117, 3874–3883. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.H.; Wang, P.; Gu, Y.; Xu, X.H.; Wang, M.L. The fate and impact on microorganisms of rice allelochemicals in paddy soil. J. Agric. Food Chem. 2008, 56, 5043–5049. [Google Scholar] [CrossRef] [PubMed]
- Li, X.J.; Xia, Z.C.; Kong, C.H.; Xu, X.H. Mobility and microbial activity of allelochemicals in soil. J. Agric. Food Chem. 2013, 61, 5072–5079. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.H.; Zhao, H.; Xu, X.H.; Wang, P.; Gu, Y. Activity and allelopathy of soil of flavone O-glycosides from rice. J. Agric. Food Chem. 2007, 55, 6007–6012. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.H.; Wang, P.; Zhao, H.; Xu, X.H.; Zhu, Y.D. Impact of allelochemical exuded from allelopathic rice on soil microbial community. Soil Biol. Biochem. 2008, 40, 1862–1869. [Google Scholar] [CrossRef]
- Maver, M.; Trevisan, F.; Miras-Moreno, B.; Lucini, L.; Trevisan, M.; Cesco, S.; Mimmo, T. The interplay between nitrogenated allelochemicals, mineral nutrition and metabolic profile in barley roots. Plant Soil 2022, 479, 715–730. [Google Scholar] [CrossRef]
- Schütz, V.; Frindte, K.; Cui, J.X.; Zhang, P.F.; Hacquard, S.; Schulze-Lefert, P.; Knief, C.; Schulz, M.; Dörmann, P. Differential impact of plant secondary metabolites on the soil microbiota. Front. Microbiol. 2021, 12, 666010. [Google Scholar] [CrossRef]
- Dong, H.Y.; Kong, C.H.; Wang, P.; Huang, Q.L. Temporal variation of soil friedelin and microbial community under different land uses in a long-term agroecosystem. Soil Biol. Biochem. 2014, 69, 275–281. [Google Scholar] [CrossRef]
- Guo, Z.Y.; Kong, C.H.; Wang, J.G.; Wang, Y.F. Rhizosphere isoflavones (daidzein and genistein) levels and their relation to the microbial community structure of mono-cropped soybean soil in field and controlled conditions. Soil Biol. Biochem. 2011, 43, 2257–2264. [Google Scholar] [CrossRef]
- Shi, K.; Shao, H. Changes in the soil fungal community mediated by a Peganum harmala allelochemical. Front. Microbiol. 2022, 13, 911836. [Google Scholar] [CrossRef]
- Inderjit. Soil microorganisms: An important determinant of allelopathic activity. Plant Soil 2005, 274, 227–236. [Google Scholar] [CrossRef]
- Revillini, D.; David, A.S.; Reyes, A.L.; Knecht, L.D.; Vigo, C.; Allen, P.; Searcy, C.A.; Afkhami, M.E. Allelopathy-selected microbiomes mitigate chemical inhibition of plant performance. New Phytol. 2023, 240, 2007–2019. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, H.; Qian, X.; Gu, J.; Chen, W.M.; Shen, X.H.; Tao, S.H.; Jiao, S.; Wei, G.H. Metagenomics insights into responses of rhizobacteria and their alleviation role in licorice allelopathy. Microbiome 2023, 11, 109. [Google Scholar] [CrossRef] [PubMed]
- Korenblum, E.; Massalha, H.; Aharoni, A. Plant–microbe interactions in the rhizosphere via a circular metabolic economy. Plant Cell 2022, 34, 3168–3182. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Kong, C.H.; Hu, F.; Xu, X.H. Allantoin involved in species interactions with rice and other organisms in paddy soil. Plant Soil 2007, 296, 43–51. [Google Scholar] [CrossRef]
- Wang, P.; Kong, C.H.; Sun, B.; Xu, X.H. Allantoin-induced change of microbial diversity and community in rice soil. Plant Soil 2010, 332, 357–368. [Google Scholar] [CrossRef]
- Zhou, B.; Kong, C.H.; Li, Y.H.; Wang, P.; Xu, X.H. Crabgrass (Digitaria sanguinalis) allelochemicals that interfere with crop growth and the soil microbial community. J. Agric. Food Chem. 2013, 61, 5310–5317. [Google Scholar] [CrossRef]
- Zhao, H.H.; Li, H.Y.; Kong, C.H. Penoxsulam-resistant barnyardgrass-mediated root microbial communities affect the growth of rice. Pest Manag. Sci. 2023, 79, 2664–2674. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Bonkowski, M.; Shen, Y.; Griffiths, B.S.; Jiang, Y.J.; Wang, X.Y.; Sun, B. Root ethylene mediates rhizosphere microbial community reconstruction when chemically detecting cyanide produced by neighbouring plants. Microbiome 2020, 8, 4. [Google Scholar] [CrossRef]
- Yin, C.T.; Vargas, J.M.C.; Schlatter, D.C.; Hagerty, C.H.; Hulbert, S.H.; Paulitz, T.C. Rhizosphere community selection reveals bacteria associated with reduced root disease. Microbiome 2021, 9, 86. [Google Scholar] [CrossRef]
- Elhaissoufi, W.; Ghoulam, C.; Barakat, A.; Zeroual, Y.; Bargaz, A. Phosphate bacterial solubilization: A key rhizosphere driving force enabling higher P use efficiency and crop productivity. J. Adv. Res. 2022, 38, 13–28. [Google Scholar] [CrossRef]
- Hao, C.K.; Dungait, J.A.J.; Wei, X.M.; Ge, T.D.; Kuzyakov, Y.; Cui, Z.L.; Tian, J.; Zhang, F.S. Maize root exudate composition alters rhizosphere bacterial community to control hotspots of hydrolase activity in response to nitrogen supply. Soil Biol. Biochem. 2022, 170, 108717. [Google Scholar] [CrossRef]
- Ulbrich, T.C.; Rivas-Ubach, A.; Tiemann, L.K.; Friesen, M.L.; Evans, S.E. Plant root exudates and rhizosphere bacterial communities shift with neighbor context. Soil Biol. Biochem. 2022, 172, 108753. [Google Scholar] [CrossRef]
- Singh, K.; Chandra, R.; Purchase, D. Unraveling the secrets of rhizobacteria signaling in rhizosphere. Rhizosphere 2022, 21, 100484. [Google Scholar] [CrossRef]
- Chaney, L.; Baucom, R.S. The soil microbial community alters patterns of selection on flowering time and fitness-related traits in ipomoea purpurea. Am. J. Bot. 2020, 107, 186–194. [Google Scholar] [CrossRef]
- Hu, L.; Robert, C.A.M.; Cadot, S.; Zhang, X.; Ye, M.; Li, B.; Manzo, D.; Chervet, N.; Steinger, T.; van der Heijden, M.G.A. Root exudate metabolites drive plant–soil feedbacks on growth and defense by shaping the rhizosphere microbiota. Nat. Commun. 2018, 9, 2738. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Ke, M.J.; Lavoie, M.; Jin, Y.J.; Fan, X.J.; Zhang, Z.Y.; Fu, Z.W.; Sun, L.W.; Gillings, M.; Penuelas, J.; et al. Rhizosphere microorganisms can influence the timing of plant flowering. Microbiome 2018, 6, 231. [Google Scholar] [CrossRef]
- Panke-Buisse, K.; Poole, A.C.; Goodrich, J.K.; Ley, R.E.; Kao-Kniffin, J. Selection on soil microbiomes reveals reproducible impacts on plant function. ISME J. 2015, 9, 980–989. [Google Scholar] [CrossRef]
- Wagner, M.R.; Lundberg, D.S.; Coleman-Derr, D.; Tringe, S.G.; Dangl, J.L.; Mitchell-Olds, T. Natural soil microbes alter flowering phenology and the intensity of selection on flowering time in a wild Arabidopsis relative. Ecol. Lett. 2014, 17, 717–726. [Google Scholar] [CrossRef]
- Ramandi, A.; Nourashrafeddin, S.M.; Marashi, S.H.; Seifi, A. Microbiome contributes to phenotypic plasticity in saffron crocus. World J. Microbiol. Biotechol. 2023, 39, 9. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.A.; Klironomos, J. Mechanisms of plant-soil feedback: Interactions among biotic and abiotic drivers. New Phytol. 2019, 222, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Doornbos, R.F.; van Loon, L.C.; Bakker, P.A.H.M. Impact of root exudates and plant defense signaling on bacterial communities in the rhizosphere. A review. Agron. Sustain. Dev. 2012, 32, 227–243. [Google Scholar] [CrossRef]
- Sun, B.; Wang, P.; Kong, C.H. Plant-soil feedback in the interference of allelopathic rice with barnyardgrass. Plant Soil 2014, 377, 309–321. [Google Scholar] [CrossRef]
- Massalha, H.; Korenblum, E.; Tholl, D.; Aharoni, A. Small molecules below-ground: The role of specialized metabolites in the rhizosphere. Plant J. 2017, 90, 788–807. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.X.; Yang, L.K.; Chen, W.S.; Li, L.L.; Zhang, P.L.; Li, Y.Z.; He, H.B.; Lin, W.X. MYB57 transcriptionally regulates MAPK11 to interact with PAL2;3 and modulate rice allelopathy. J. Exp. Bot. 2020, 71, 2127–2141. [Google Scholar] [CrossRef]
- Fraser, C.M.; Chapple, C. The phenylpropanoid pathway in arabidopsis. Arab. Book 2011, 9, e0152. [Google Scholar] [CrossRef] [PubMed]
- Muro-Villanueva, F.; Mao, X.Y.; Chapple, C. Linking phenylpropanoid metabolism, lignin deposition, and plant growth inhibition. Curr. Opin. Biotech. 2019, 56, 202–208. [Google Scholar] [CrossRef]
- Li, C.N.; Ng, C.; Fan, L.M. MYB transcription factors, active players in abiotic stress signaling. Environ. Exp. Bot. 2015, 114, 80–91. [Google Scholar] [CrossRef]
- Dias, A.P.; Grotewold, E. Manipulating the accumulation of phenolics in maize cultured cells using transcription factors. Biochem. Eng. J. 2003, 14, 207–216. [Google Scholar] [CrossRef]
- Rommens, C.M.; Richael, C.M.; Yan, H.; Navarre, D.A.; Ye, J.S.; Krucker, M.; Swords, K. Engineered native pathways for high kaempferol and caffeoylquinate production in potato. Plant Biotech. J. 2008, 6, 870–886. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Butelli, E.; Hill, L.; Parr, A.; Niggeweg, R.; Bailey, P.; Weisshaar, B.; Martin, C. AtMYB12 regulates caffeoyl quinic acid and flavonol synthesis in tomato: Expression in fruit results in very high levels of both types of polyphenol. Plant J. 2008, 56, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.P.; Wang, Y.; Huang, F.F.; Lu, S.J.; Zhao, L.M.; Ma, X.Y.; Kai, G.Y. SmMYB2 promotes salvianolic acid biosynthesis in the medicinal herb Salvia miltiorrhiza. J. Integr. Plant Biol. 2020, 62, 1688–1702. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.; Misra, P.; Khan, M.P.; Swarnkar, G.; Tewari, M.C.; Bhambhani, S.; Trivedi, R.; Chattopadhyay, N.; Trivedi, P.K. Co-expression of Arabidopsis transcription factor, AtMYB12, and soybean isoflavone synthase, GmIFS1, genes in tobacco leads to enhanced biosynthesis of isoflavones and flavonols resulting in osteoprotective activity. Plant Biotech. J. 2014, 12, 69–80. [Google Scholar] [CrossRef] [PubMed]
- Fornalé, S.; Lopez, E.; Salazar-Henao, J.E.; Fernández-Nohales, P.; Rigau, J.; Caparros-Ruiz, D. AtMYB7, a new player in the regulation of uv-sunscreens in Arabidopsis thaliana. Plant Cell Physiol. 2014, 55, 507–516. [Google Scholar] [CrossRef]
- Nakatsuka, T.; Saito, M.; Yamada, E.; Fujita, K.; Kakizaki, Y.; Nishihara, M. Isolation and characterization of GtMYBP3 and GtMYBP4, orthologues of R2R3-MYB transcription factors that regulate early flavonoid biosynthesis, in gentian flowers. J. Exp. Bot. 2012, 63, 6505–6517. [Google Scholar] [CrossRef]
- Vimolmangkang, S.; Han, Y.P.; Wei, G.C.; Korban, S.S. An apple MYB transcription factor, MdMYB3, is involved in regulation of anthocyanin biosynthesis and flower development. BMC Plant Biol. 2013, 13, 176. [Google Scholar] [CrossRef]
- Xu, F.; Ning, Y.J.; Zhang, W.W.; Liao, Y.L.; Li, L.L.; Cheng, H.; Cheng, S.Y. An R2R3-MYB transcription factor as a negative regulator of the flavonoid biosynthesis pathway in Ginkgo biloba. Funct. Integr. Genom. 2014, 14, 177–189. [Google Scholar] [CrossRef]
- Yan, J.H.; Wang, B.; Zhong, Y.P.; Yao, L.M.; Cheng, L.J.; Wu, T.L. The soybean R2R3 MYB transcription factor GmMYB100 negatively regulates plant flavonoid biosynthesis. Plant Mol. Biol. 2015, 89, 35–48. [Google Scholar] [CrossRef]
- Zhao, M.Z.; Wang, T.L.; Wu, P.; Guo, W.Y.; Su, L.T.; Wang, Y.; Liu, Y.J.; Yan, F.; Wang, Q.Y. Isolation and characterization of GmMYBJ3, an R2R3-MYB transcription factor that affects isoflavonoids biosynthesis in soybean. PLoS ONE 2017, 12, e0179990. [Google Scholar] [CrossRef]
- Huang, D.; Yu, L.C.; Ming, R.H.; Tan, X.M.; Li, L.B.; Huang, R.S.; Tan, Y.; Yao, S.C. A chromosome-level genome assembly of Callerya speciosa sheds new light on the biosynthesis of root-specific isoflavonoids. Ind. Crop. Prod. 2023, 200, 116877. [Google Scholar] [CrossRef]
- Saxena, S.; Pal, L.; Naik, J.; Singh, Y.; Verma, P.K.; Chattopadhyay, D.; Pandey, A. The R2R3-MYB-SG7 transcription factor CaMYB39 orchestrates surface phenylpropanoid metabolism and pathogen resistance in chickpea. New Phytol. 2023, 238, 798–816. [Google Scholar] [CrossRef] [PubMed]
- Widhalm, J.R.; Rhodes, D. Biosynthesis and molecular actions of specialized 1,4-naphthoquinone natural products produced by horticultural plants. Hortic. Res. 2016, 3, 16046. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.L.; Pu, Z.Q.; Cao, G.; You, D.W.; Zhou, Y.; Deng, C.P.; Shi, M.; Nile, S.H.; Wang, Y.; Zhou, W.; et al. Tanshinone and salvianolic acid biosynthesis are regulated by SmMYB98 in Salvia miltiorrhiza hairy roots. J. Adv. Res. 2020, 23, 1–12. [Google Scholar] [CrossRef]
- McCoy, R.M.; Utturkar, S.M.; Crook, J.W.; Thimmapuram, J.; Widhalm, J.R. The origin and biosynthesis of the naphthalenoid moiety of juglone in black walnut. Hortic. Res. 2018, 5, 67. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.C.; Yang, Y.; Peng, Y.; Wang, S.Y.; Zhang, J.; Liu, X.B.; Liu, J.J.; Wen, B.B.; Li, M.F. Biosynthesis and the transcriptional regulation of terpenoids in tea plants (Camellia sinensis). Int. J. Mol. Sci. 2023, 24, 6937. [Google Scholar] [CrossRef] [PubMed]
- Karunanithi, P.S.; Zerbe, P. Terpene synthases as metabolic gatekeepers in the evolution of plant terpenoid chemical diversity. Front. Plant Sci. 2019, 10, 1166. [Google Scholar] [CrossRef]
- Chen, S.; Xie, P.F.; Li, Y.Y.; Wang, X.X.; Liu, H.H.; Wang, S.S.; Han, W.B.; Wu, R.M.; Li, X.L.; Guan, Y.F.; et al. New insights into stress-induced β-Ocimene biosynthesis in tea (Camellia sinensis) leaves during oolong tea processing. J. Agric. Food Chem. 2021, 69, 11656–11664. [Google Scholar] [CrossRef]
- Xu, Q.S.; He, Y.X.; Yan, X.M.; Zhao, S.Q.; Zhu, J.Y.; Wei, C.L. Unraveling a crosstalk regulatory network of temporal aroma accumulation in tea plant (Camellia sinensis) leaves by integration of metabolomics and transcriptomics. Environ. Exp. Bot. 2018, 149, 81–94. [Google Scholar] [CrossRef]
- Kitaoka, N.; Zhang, J.; Oyagbenro, R.K.; Brown, B.; Wu, Y.S.; Yang, B.; Li, Z.H.; Peters, R.J. Interdependent evolution of biosynthetic gene clusters for momilactone production in rice. Plant Cell 2021, 33, 290–305. [Google Scholar] [CrossRef]
- Cárdenas, P.D.; Sonawane, P.D.; Heinig, U.; Bocobza, S.E.; Burdman, S.; Aharoni, A. The bitter side of the nightshades: Genomics drives discovery in Solanaceae steroidal alkaloid metabolism. Phytochemistry 2015, 113, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Cambier, V.; Hance, T.; de Hoffmann, E. Variation of DIMBOA and related compounds content in relation to the age and plant organ in maize. Phytochemistry 2000, 53, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Richter, A.; Powell, A.F.; Mirzaei, M.; Wang, L.J.; Movahed, N.; Miller, J.K.; Piñeros, M.A.; Jander, G. Indole-3-glycerolphosphate synthase, a branchpoint for the biosynthesis of tryptophan, indole, and benzoxazinoids in maize. Plant J. 2021, 106, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Czjzek, M.; Cicek, M.; Zamboni, V.; Bevan, D.R.; Henrissat, B.; Esen, A. The mechanism of substrate (aglycone) specificity in β-glucosidases is revealed by crystal structures of mutant maize β-glucosidase-DIMBOA, -DIMBOAGlc, and -dhurrin complexes. Proc. Natl. Acad. Sci. USA 2000, 97, 13555–13560. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Liu, H.; Zhuang, H.F.; Zhao, C.X.; Xu, Y.X.; Wu, S.B.; Qi, J.F.; Li, J.; Hettenhausen, C.; Wu, J.Q. Transcriptomics and alternative splicing analyses reveal large differences between maize lines b73 and mo17 in response to aphid Rhopalosiphum padi infestation. Front. Plant Sci. 2017, 8, 1738. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Li, Z.; Magnusson, E.; Cano, F.G.; Crisp, P.A.; Noshay, J.M.; Grotewold, E.; Hirsch, C.N.; Briggs, S.P.; Springer, N.M. Meta gene regulatory networks in maize highlight functionally relevant regulatory interactions. Plant Cell 2020, 32, 1377–1396. [Google Scholar] [CrossRef]
- Batyrshina, Z.S.; Shavit, R.; Yaakov, B.; Bocobza, S.; Tzin, V. The transcription factor TaMYB31 regulates the benzoxazinoid biosynthetic pathway in wheat. J. Exp. Bot. 2022, 73, 5634–5649. [Google Scholar] [CrossRef]
- Dudareva, N.; Pichersky, E.; Gershenzon, J. Biochemistry of plant volatiles. Plant Physiol. 2004, 135, 1893–1902. [Google Scholar] [CrossRef]
- Rodríguez-Concepción, M.; Boronat, A. Elucidation of the methylerythritol phosphate pathway for isoprenoid biosynthesis in bacteria and plastids.: A metabolic milestone achieved through genomics. Plant Physiol. 2002, 130, 1079–1089. [Google Scholar] [CrossRef]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef]
- Yu, Z.X.; Li, J.X.; Yang, C.Q.; Hu, W.L.; Wang, L.J.; Chen, X.Y. The jasmonate-responsive AP2/ERF transcription factors AaERF1 and AaERF2 positively regulate artemisinin biosynthesis in Artemisia annua L. Mol. Plant 2012, 5, 353–365. [Google Scholar] [CrossRef]
- Ji, Y.P.; Xiao, J.W.; Shen, Y.L.; Ma, D.M.; Li, Z.Q.; Pu, G.B.; Li, X.; Huang, L.L.; Liu, B.Y.; Ye, H.C.; et al. Cloning and characterization of AabHLH1, a bHLH transcription factor that positively regulates artemisinin biosynthesis in Artemisia annua. Plant Cell Physiol. 2014, 55, 1592–1604. [Google Scholar] [CrossRef]
- Mertens, J.; Pollier, J.; Vanden Bossche, R.; Lopez-Vidriero, I.; Franco-Zorrilla, J.M.; Goossens, A. The bHLH transcription factors TSAR1 and TSAR2 regulate triterpene saponin biosynthesis in Medicago truncatula. Plant Physiol. 2016, 170, 194–210. [Google Scholar] [CrossRef]
- Sun, W.J.; Zhan, J.Y.; Zheng, T.R.; Sun, R.; Wang, T.; Tang, Z.Z.; Bu, T.L.; Li, C.L.; Wu, Q.; Chen, H. The jasmonate-responsive transcription factor CbWRKY24 regulates terpenoid biosynthetic genes to promote saponin biosynthesis in Conyza blinii H. Lév. J. Genet. 2018, 97, 1379–1388. [Google Scholar] [CrossRef]
- Hernández-Aparicio, F.; Lisón, P.; Rodrigo, I.; Bellés, J.M.; López-Gresa, M.P. Signaling in the tomato immunity against Fusarium oxysporum. Molecules 2021, 26, 1818. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, J.L.; Lin, J.H.; Lin, J.T.; Wu, Z.Z. Odorant-binding proteins and chemosensory proteins potentially involved in host plant recognition in the Asian citrus psyllid, Diaphorina citri. Pest Manag. Sci. 2020, 76, 2609–2618. [Google Scholar] [CrossRef]
- Clark, J.; Bennett, T. Cracking the enigma: Understanding strigolactone signalling in the rhizosphere. J. Exp. Bot. 2024, 75, 1159–1173. [Google Scholar] [CrossRef]
- Jia, K.P.; Baz, L.; Al-Babili, S. From carotenoids to strigolactones. J. Exp. Bot. 2018, 69, 2189–2204. [Google Scholar] [CrossRef]
- Lahari, Z.; van Boerdonk, S.; Omoboye, O.O.; Reichelt, M.; Höfte, M.; Gershenzon, J.; Gheysen, G.; Ullah, C. Strigolactone deficiency induces jasmonate, sugar and flavonoid phytoalexin accumulation enhancing rice defense against the blast fungus Pyricularia oryzae. New Phytol. 2024, 241, 827–844. [Google Scholar] [CrossRef]
- Han, Z.; Zhang, C.; Zhang, H.; Duan, Y.; Zou, Z.; Zhou, L.; Zhu, X.; Fang, W.; Ma, Y. CsMYB transcription factors participate in jasmonic acid signal transduction in response to cold stress in tea plant (Camellia sinensis). Plants 2022, 11, 2869. [Google Scholar] [CrossRef]
- Qi, P.F.; Zhang, Y.Z.; Liu, C.H.; Chen, Q.; Guo, Z.R.; Wang, Y.; Xu, B.J.; Jiang, Y.F.; Zheng, T.; Gong, X.; et al. Functional analysis of FgNahG clarifies the contribution of salicylic acid to wheat (Triticum aestivum) resistance against fusarium head blight. Toxins 2019, 11, 59. [Google Scholar] [CrossRef]
- Du, L.Q.; Ali, G.S.; Simons, K.A.; Hou, J.G.; Yang, T.B.; Reddy, A.S.N.; Poovaiah, B.W. Ca2+/calmodulin regulates salicylic-acid-mediated plant immunity. Nature 2009, 457, 1154–1158. [Google Scholar] [CrossRef]
- Sarkar, S.; Das, A.; Khandagale, P.; Maiti, I.B.; Chattopadhyay, S.; Dey, N. Interaction of Arabidopsis TGA3 and WRKY53 transcription factors on Cestrum yellow leaf curling virus (CmYLCV) promoter mediates salicylic acid-dependent gene expression in plants. Planta 2018, 247, 181–199. [Google Scholar] [CrossRef]
- Song, G.C.; Ryu, C.M. Evidence for volatile memory in plants: Boosting defence priming through the recurrent application of plant volatiles. Mol. Cells 2018, 41, 724–732. [Google Scholar]
- Ye, M.; Liu, M.M.; Erb, M.; Glauser, G.; Zhang, J.; Li, X.W.; Sun, X.L. Indole primes defence signalling and increases herbivore resistance in tea plants. Plant Cell Environ. 2021, 44, 1165–1177. [Google Scholar] [CrossRef]
- Ye, M.; Glauser, G.; Lou, Y.G.; Erb, M.; Hu, L.F. Molecular dissection of early defense signaling underlying volatile-mediated defense regulation and herbivore resistance in rice. Plant Cell 2019, 31, 687–698. [Google Scholar] [CrossRef]
- Stenzel, I.; Hause, B.; Miersch, O.; Kurz, T.; Maucher, H.; Weichert, H.; Ziegler, J.; Feussner, I.; Wasternack, C. Jasmonate biosynthesis and the allene oxide cyclase family of Arabidopsis thaliana. Plant Mol. Biol. 2003, 51, 895–911. [Google Scholar] [CrossRef]
- Matsui, K. Green leaf volatiles: Hydroperoxide lyase pathway of oxylipin metabolism. Curr. Opin. Plant Biol. 2006, 9, 274–280. [Google Scholar] [CrossRef]
- Werner, A.K.; Witte, C.P. The biochemistry of nitrogen mobilization: Purine ring catabolism. Trends Plant Sci. 2011, 16, 381–387. [Google Scholar] [CrossRef]
- Watanabe, S.; Matsumoto, M.; Hakomori, Y.; Takagi, H.; Shimada, H.; Sakamoto, A. The purine metabolite allantoin enhances abiotic stress tolerance through synergistic activation of abscisic acid metabolism. Plant Cell Environ. 2014, 37, 1022–1036. [Google Scholar] [CrossRef]
- Takagi, H.; Ishiga, Y.; Watanabe, S.; Konishi, T.; Egusa, M.; Akiyoshi, N.; Matsuura, T.; Mori, I.C.; Hirayama, T.; Kaminaka, H.; et al. Allantoin, a stress-related purine metabolite, can activate jasmonate signaling in a MYC2-regulated and abscisic acid-dependent manner. J. Exp. Bot. 2016, 67, 2519–2532. [Google Scholar] [CrossRef]
- Liao, P.; Ray, S.; Boachon, B.; Lynch, J.H.; Deshpande, A.; McAdam, S.; Morgan, J.A.; Dudareva, N. Cuticle thickness affects dynamics of volatile emission from petunia flowers. Nat. Chem. Biol. 2021, 17, 138–145. [Google Scholar] [CrossRef]
- Wang, L.; Erb, M. Volatile uptake, transport, perception, and signaling shape a plant’s nose? Essays Biochem. 2022, 66, 695–702. [Google Scholar]
- Chang, C.; Kwok, S.F.; Bleecker, A.B.; Meyerowitz, E.M. Arabidopsis ethylene-response gene etr1—Similarity of product to 2-component regulators. Science 1993, 262, 539–544. [Google Scholar] [CrossRef]
- Nagashima, A.; Higaki, T.; Koeduka, T.; Ishigami, K.; Hosokawa, S.; Watanabe, H.; Matsui, K.; Hasezawa, S.; Touhara, K. Transcriptional regulators involved in responses to volatile organic compounds in plants. J. Biol. Chem. 2019, 294, 2256–2266. [Google Scholar] [CrossRef]
- Adebesin, F.; Widhalm, J.R.; Boachon, B.; Lefèvre, F.; Pierman, B.; Lynch, J.H.; Alam, I.; Junqueira, B.; Benke, R.; Ray, S.; et al. Emission of volatile organic compounds from petunia flowers is facilitated by an ABC transporter. Science 2017, 356, 1386–1388. [Google Scholar] [CrossRef]
- Meents, A.K.; Mithöfer, A. Plant-plant communication: Is there a role for volatile damage-associated molecular patterns? Front. Plant Sci. 2020, 11, 583275. [Google Scholar] [CrossRef]
- Asai, N.; Nishioka, T.; Takabayashi, J.; Furuichi, T. Plant volatiles regulate the activities of Ca2+-permeable channels and promote cytoplasmic calcium transients in Arabidopsis leaf cells. Plant Signal. Behav. 2009, 4, 294–300. [Google Scholar] [CrossRef]
- Yin, W.C.; Wang, X.H.; Liu, H.; Wang, Y.; van Nocker, S.; Tu, M.X.; Fang, J.H.; Guo, J.Q.; Li, Z.; Wang, X.P. Overexpression of VqWRKY31 enhances powdery mildew resistance in grapevine by promoting salicylic acid signaling and specific metabolite synthesis. Hortic. Res. 2022, 9, uhab064. [Google Scholar] [CrossRef]
- Boudsocq, M.; Willmann, M.R.; McCormack, M.; Lee, H.; Shan, L.B.; He, P.; Bush, J.; Cheng, S.H.; Sheen, J. Differential innate immune signalling via Ca2+ sensor protein kinases. Nature 2010, 464, 418–422. [Google Scholar] [CrossRef]
- Manna, M.; Rengasamy, B.; Sinha, A.K. Revisiting the role of MAPK signalling pathway in plants and its manipulation for crop improvement. Plant Cell Environ. 2023, 46, 2277–2295. [Google Scholar] [CrossRef]
- Dombrowski, J.E.; Kronmiller, B.A.; Hollenbeck, V.G.; Rhodes, A.C.; Henning, J.A.; Martin, R.C. Transcriptome analysis of the model grass Lolium temulentum exposed to green leaf volatiles. BMC Plant Biol. 2019, 19, 222. [Google Scholar] [CrossRef]
- Zhang, M.M.; Su, J.B.; Zhang, Y.; Xu, J.; Zhang, S.Q. Conveying endogenous and exogenous signals: MAPK cascades in plant growth and defense. Curr. Opin. Plant Biol. 2018, 45, 1–10. [Google Scholar] [CrossRef]
- Kandoth, P.K.; Ranf, S.; Pancholi, S.S.; Jayanty, S.; Walla, M.D.; Miller, W.; Howe, G.A.; Lincoln, D.E.; Stratmann, J.W. Tomato MAPKs LeMPK1, LeMPK2, and LeMPK3 function in the systemin-mediated defense response against herbivorous insects. Proc. Natl. Acad. Sci. USA 2007, 104, 12205–12210. [Google Scholar] [CrossRef]
- Song, N.; Wu, J.S. Synergistic induction of phytoalexins in Nicotiana attenuata by jasmonate and ethylene signaling mediated by NaWRKY70. J. Exp. Bot. 2024, 75, 1063–1080. [Google Scholar] [CrossRef]
- Ding, M.L.; Xie, Y.F.; Zhang, Y.H.; Cai, X.N.; Zhang, B.; Ma, P.D.; Dong, J.E. Salicylic acid regulates phenolic acid biosynthesis via SmNPR1-SmTGA2/SmNPR4 modules in Salvia miltiorrhiza. J. Exp. Bot. 2023, 74, 5736–5751. [Google Scholar] [CrossRef]
- Jiang, W.M.; Fu, X.Q.; Pan, Q.F.; Tang, Y.L.; Shen, Q.; Lv, Z.Y.; Yan, T.X.; Shi, P.; Li, L.; Zhang, L.D.; et al. Overexpression of AaWRKY1 leads to an enhanced content of artemisinin in Artemisia annua. Biomed. Res. Int. 2016, 971, 7314971. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Signaling Chemicals | Function/Response | References | |
|---|---|---|---|
| Air-borne signals | Ethylene | Alarm and induce defense | [102] |
| Participate in plant neighbor detection | [116] | ||
| Induce shade avoidance response | [117] | ||
| Alarm for roots to avoid compacted soils | [118] | ||
| Methyl jasmonate | Alarm and induce defense | [103] | |
| Attract natural enemies of the herbivores | [104] | ||
| (Z)-3-Hexenyl acetate | Induce glycosylation reaction | [106] | |
| (E)-2-Hexenal | Induce defense responses | [107] | |
| (Z)-3-Hexenol | Induce defense responses | [108] | |
| β-caryophyllene | Induce defense responses | [109] | |
| Myrcene | Induce the accumulation of MeJA | [110] | |
| Ocimene | Induce the accumulation of MeJA | [110] | |
| 4,8-Dimethylnona-1,3,7-triene | Induce defense responses | [111] | |
| (E)-Nerolidol | Induce defense responses | [112] | |
| 4,8,12-Trimethyltrideca-1,3,7,11-tetraene | Induce defense responses | [113] | |
| Methyl salicylate | Induce defense responses | [105] | |
| Indole | Induce defense responses | [115] | |
| Methyl benzoate | Trigger phytotoxic effects | [114] | |
| Soil-borne signals | Jasmonic acid | Induce allelopathy | [119] |
| Participate in plant neighbor detection | [23] | ||
| Salicylic acid | Participate in plant neighbor detection and allelopathy | [16] | |
| Allantoin | Participate in kin recognition in rice | [87] | |
| Activate stress response | [120] | ||
| (–)-Loliolide | Participate in plant neighbor detection | [23] | |
| Induce defense responses | [121] | ||
| Mediate plant kin recognition | [122] | ||
| Modulate flowering time | [123] | ||
| Modify the rhizosphere microbial community | [124] | ||
| Root–microbe signals | Strigolactones | The symbiosis between roots and arbuscular mycorrhizal fungi | [125] |
| Induce parasitic weeds to search for potential hosts | [126] | ||
| Luteolin Quercetin-3-O-rhamnoside | Attract rhizobia; induce the early formation of root nodules | [127] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, C.-H.; Li, Z.; Li, F.-L.; Xia, X.-X.; Wang, P. Chemically Mediated Plant–Plant Interactions: Allelopathy and Allelobiosis. Plants 2024, 13, 626. https://doi.org/10.3390/plants13050626
Kong C-H, Li Z, Li F-L, Xia X-X, Wang P. Chemically Mediated Plant–Plant Interactions: Allelopathy and Allelobiosis. Plants. 2024; 13(5):626. https://doi.org/10.3390/plants13050626
Chicago/Turabian StyleKong, Chui-Hua, Zheng Li, Feng-Li Li, Xin-Xin Xia, and Peng Wang. 2024. "Chemically Mediated Plant–Plant Interactions: Allelopathy and Allelobiosis" Plants 13, no. 5: 626. https://doi.org/10.3390/plants13050626
APA StyleKong, C.-H., Li, Z., Li, F.-L., Xia, X.-X., & Wang, P. (2024). Chemically Mediated Plant–Plant Interactions: Allelopathy and Allelobiosis. Plants, 13(5), 626. https://doi.org/10.3390/plants13050626