
Strong Invasive Mechanism of Wedelia trilobata via Growth and Physiological Traits under Nitrogen Stress Condition

 , ,
, ,  ,
, 

Abstract
1. Introduction
2. Results
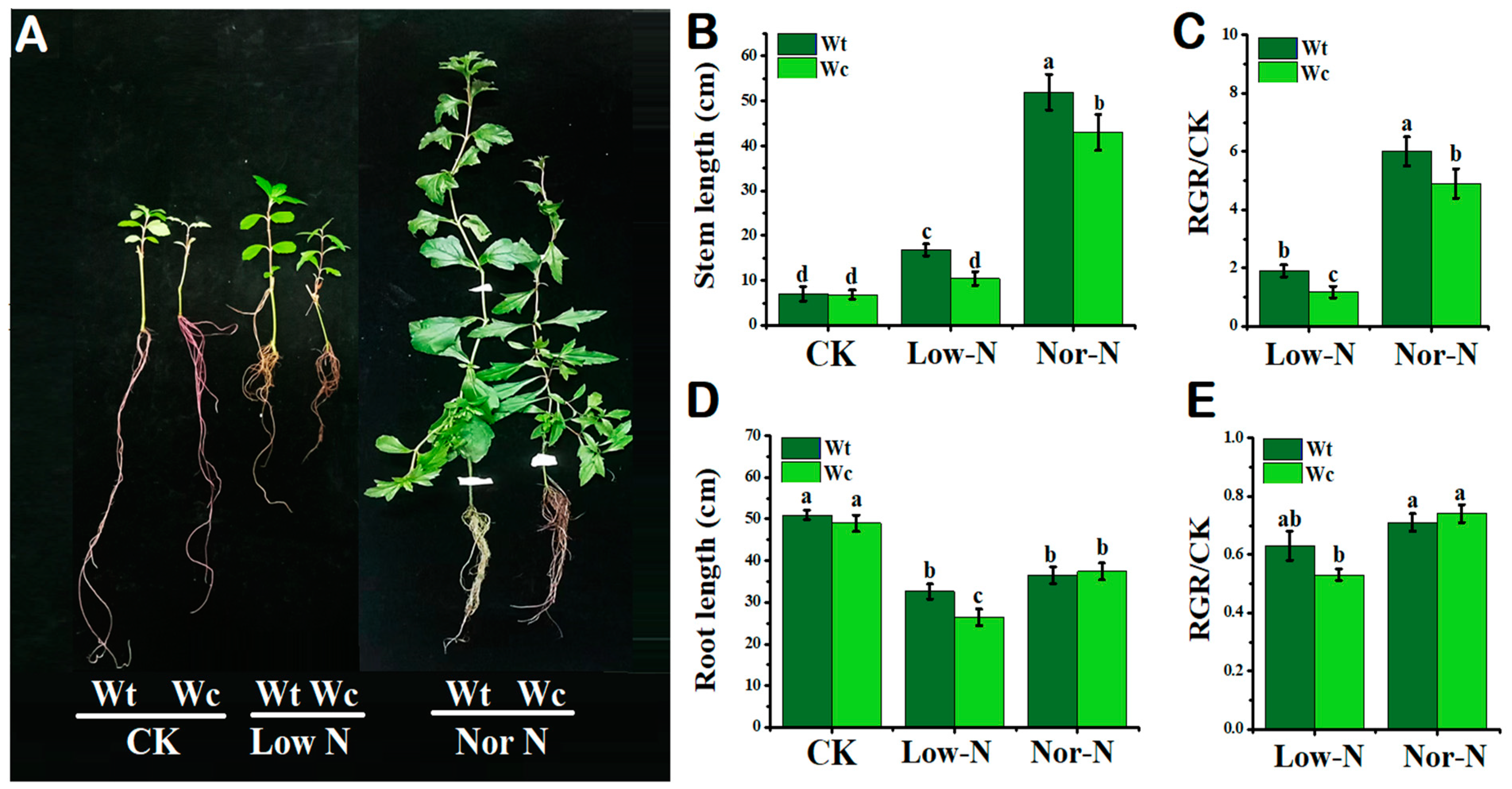
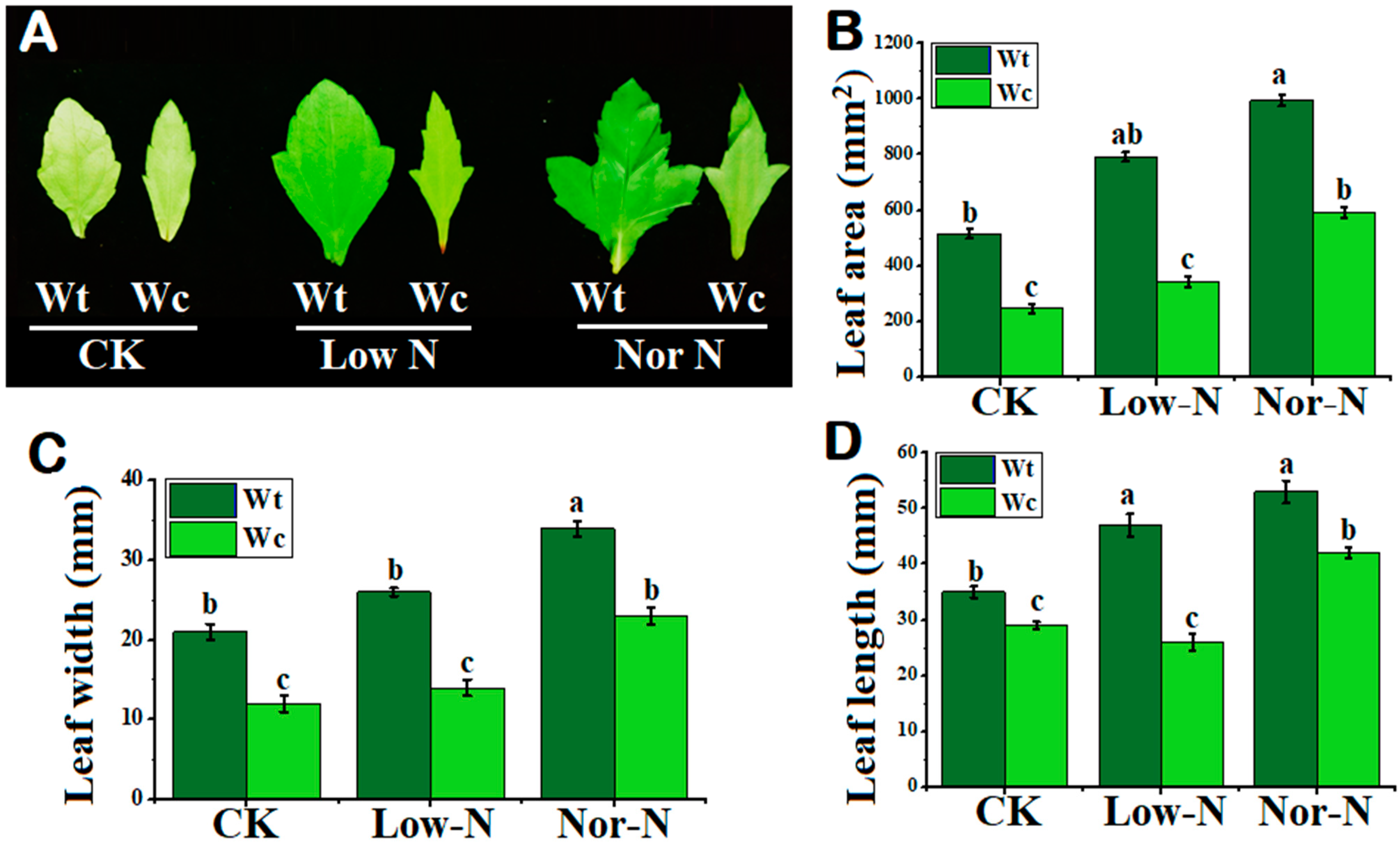
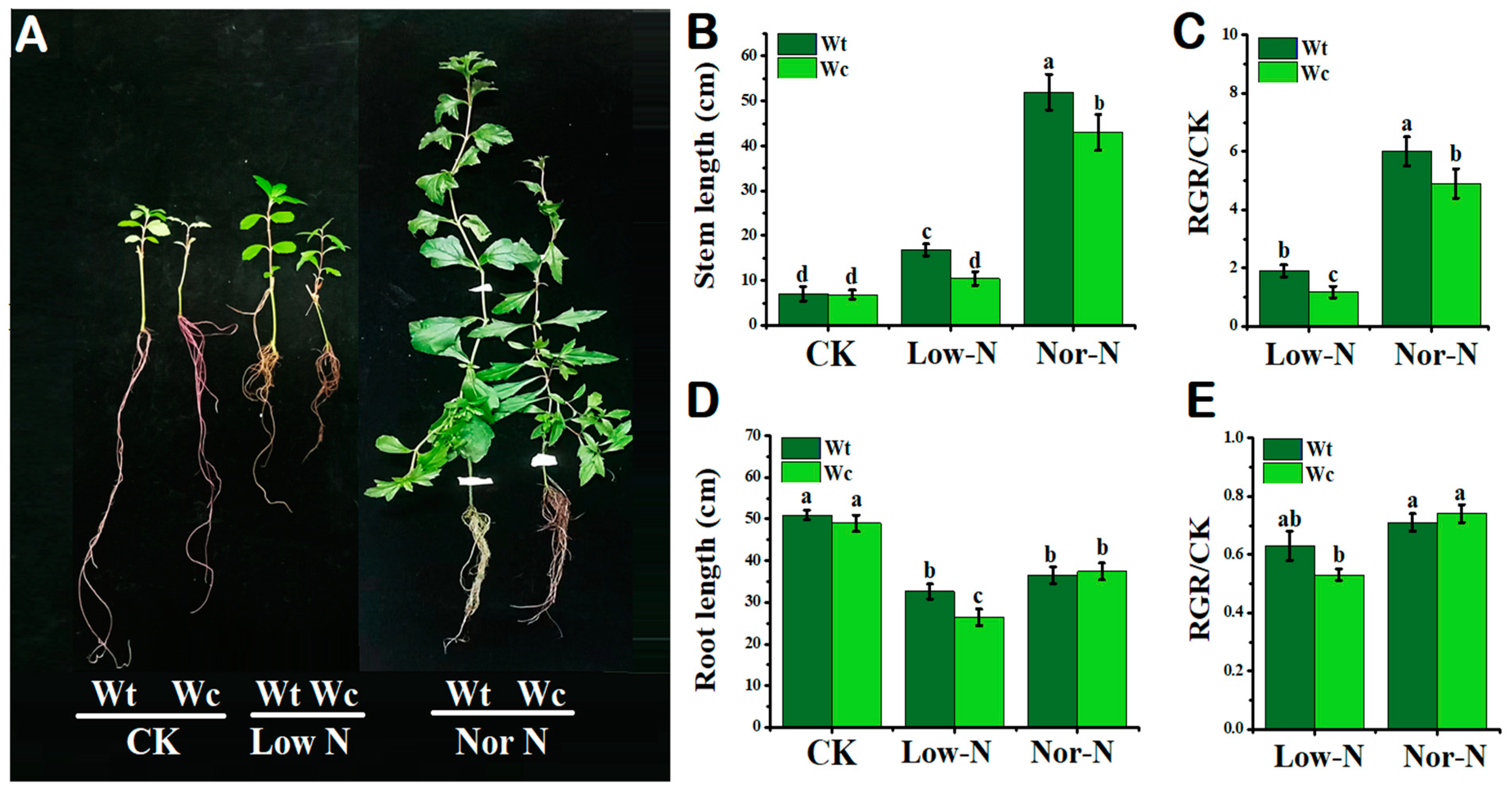
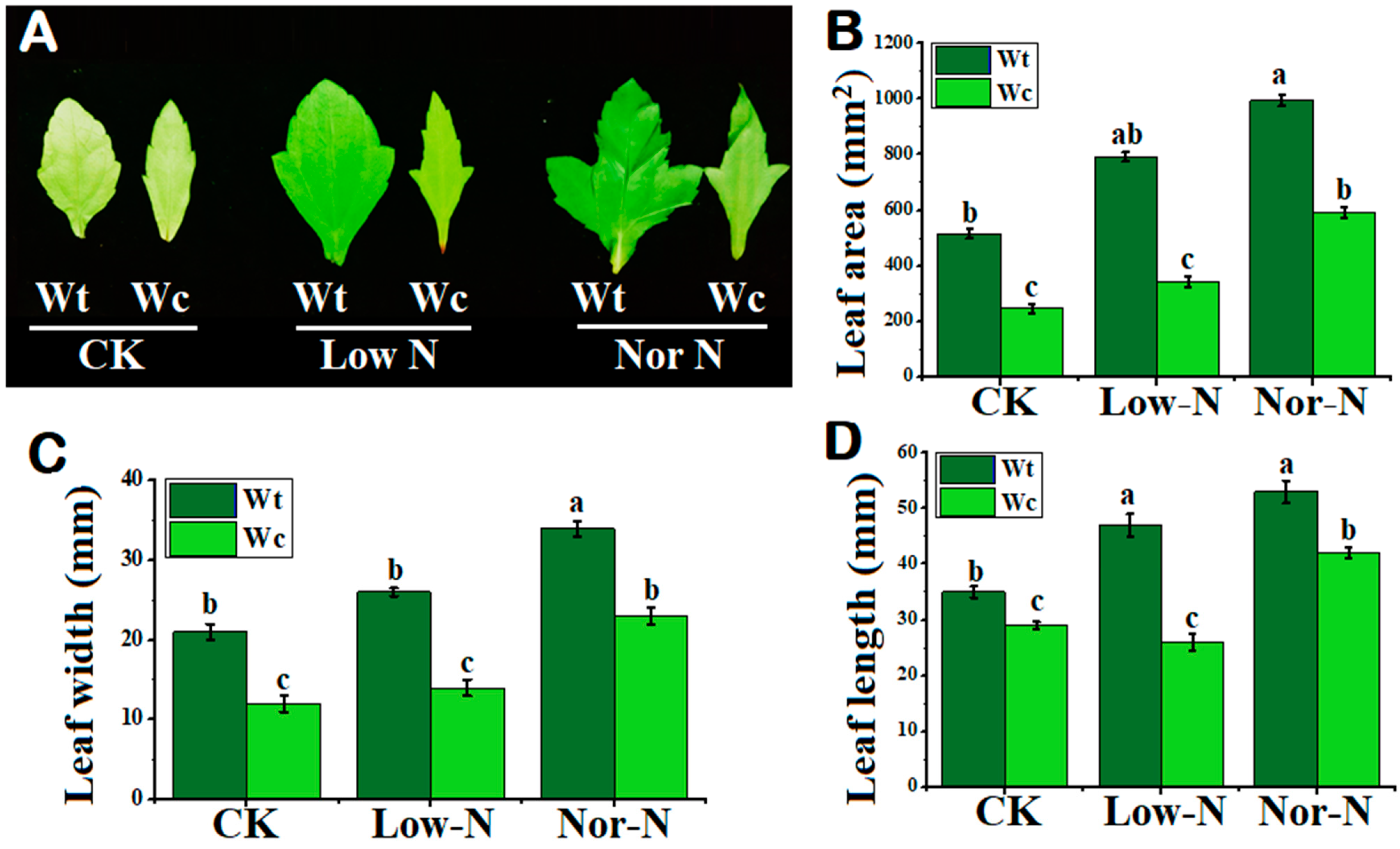
2.1. Comparison of Plants by Phenotypical and Growth Traits
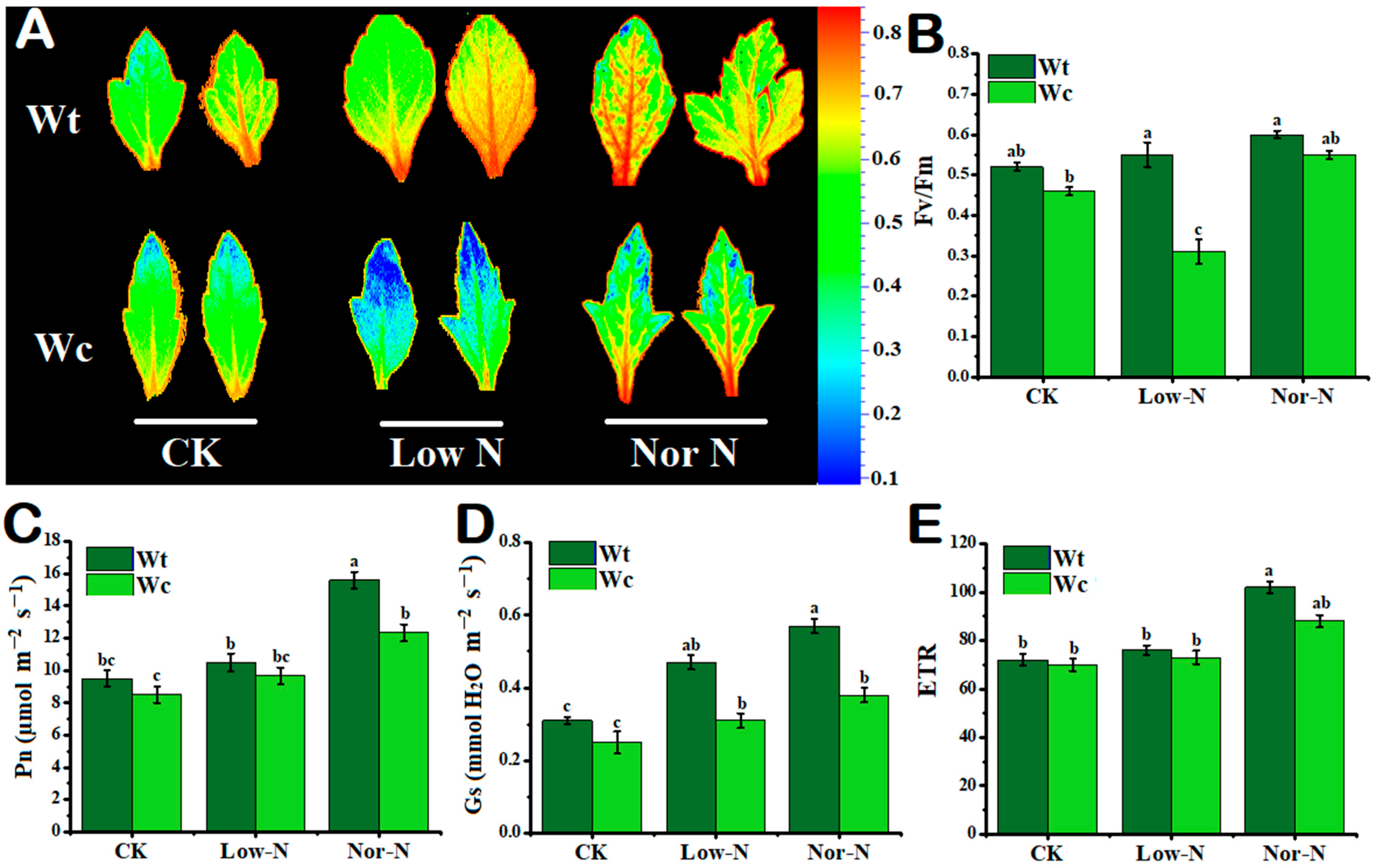
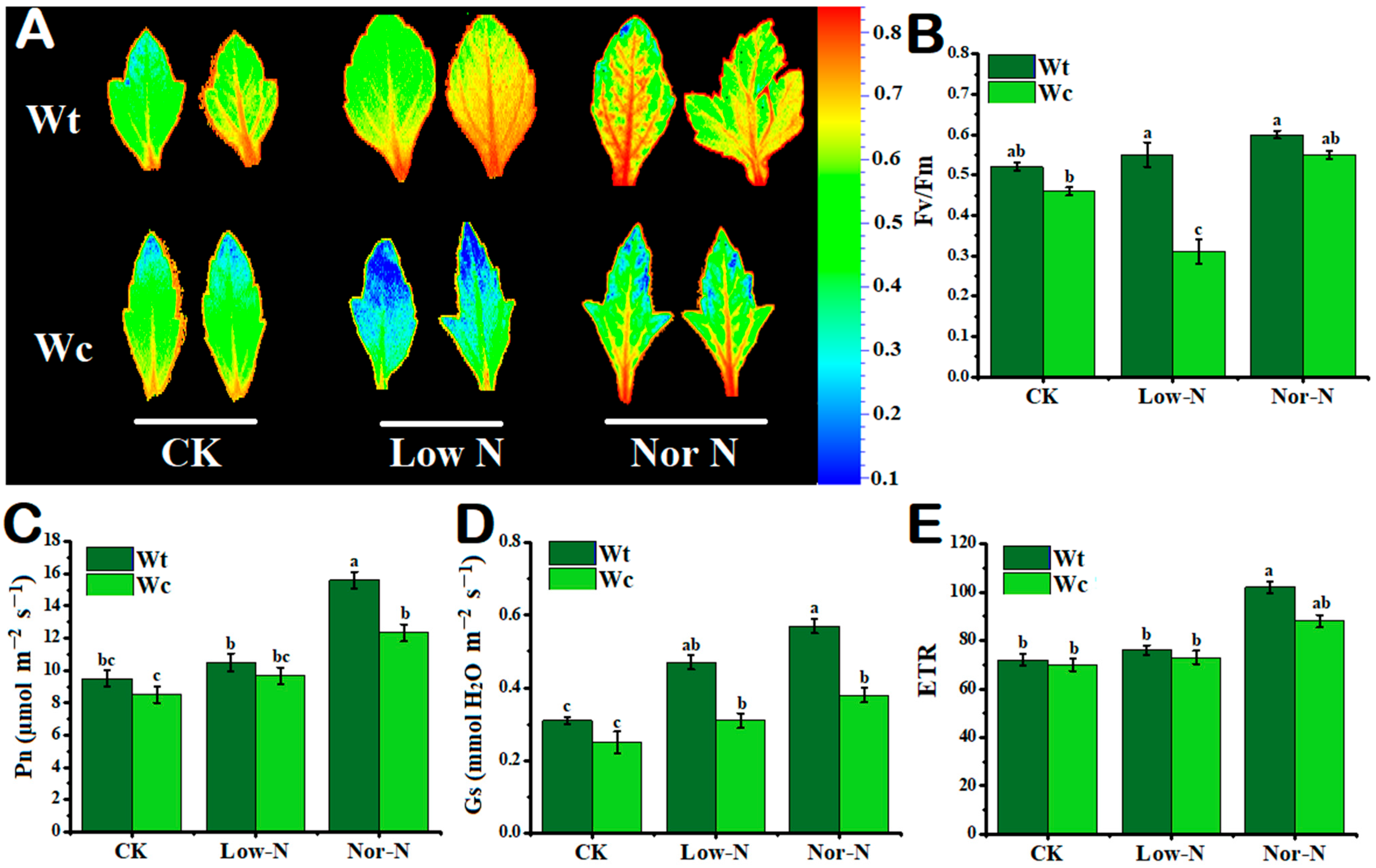
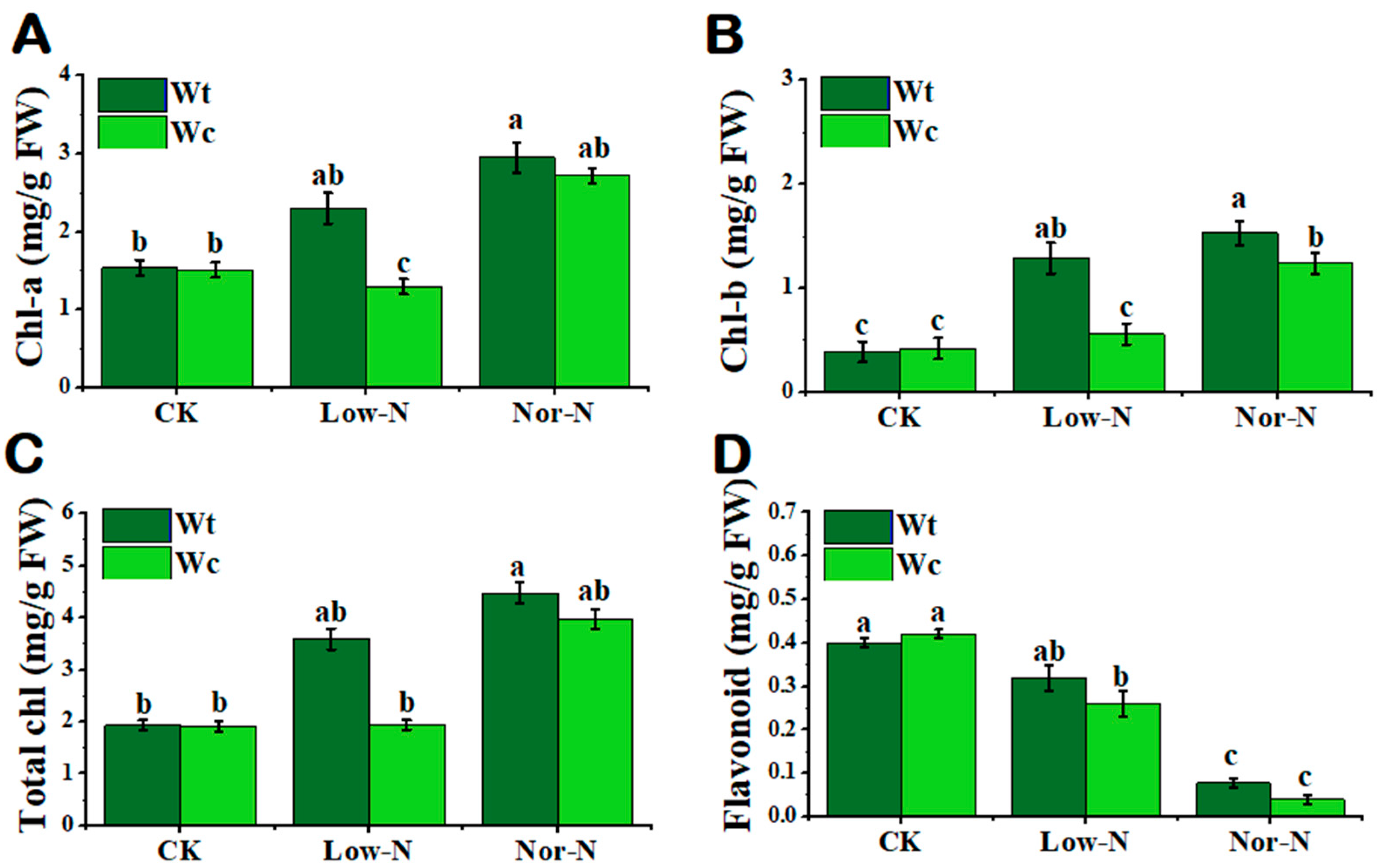
2.2. Tolerance of Leaves under Different N Conditions
2.3. Leaf Photosynthetic Response to N Conditions
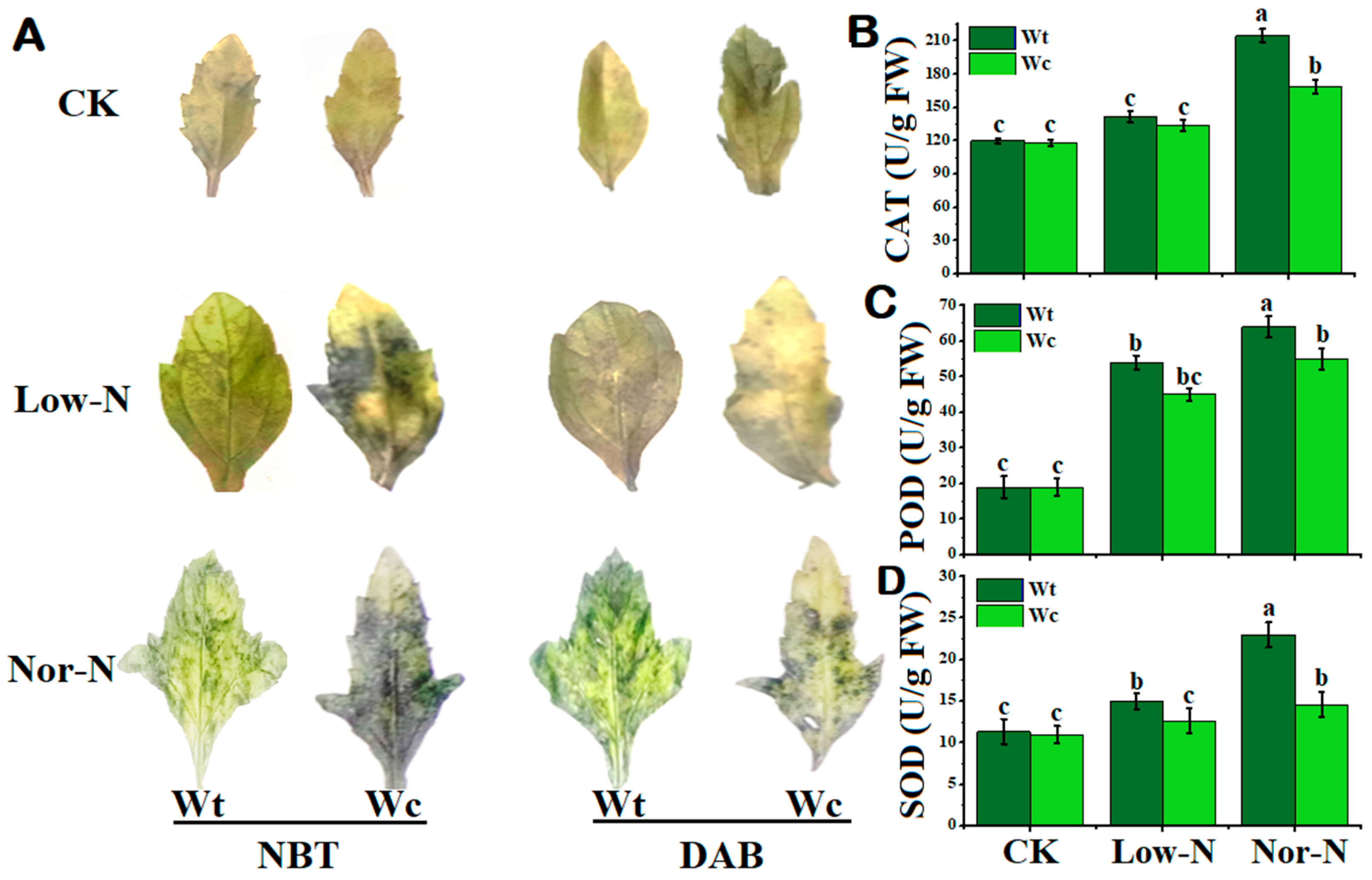
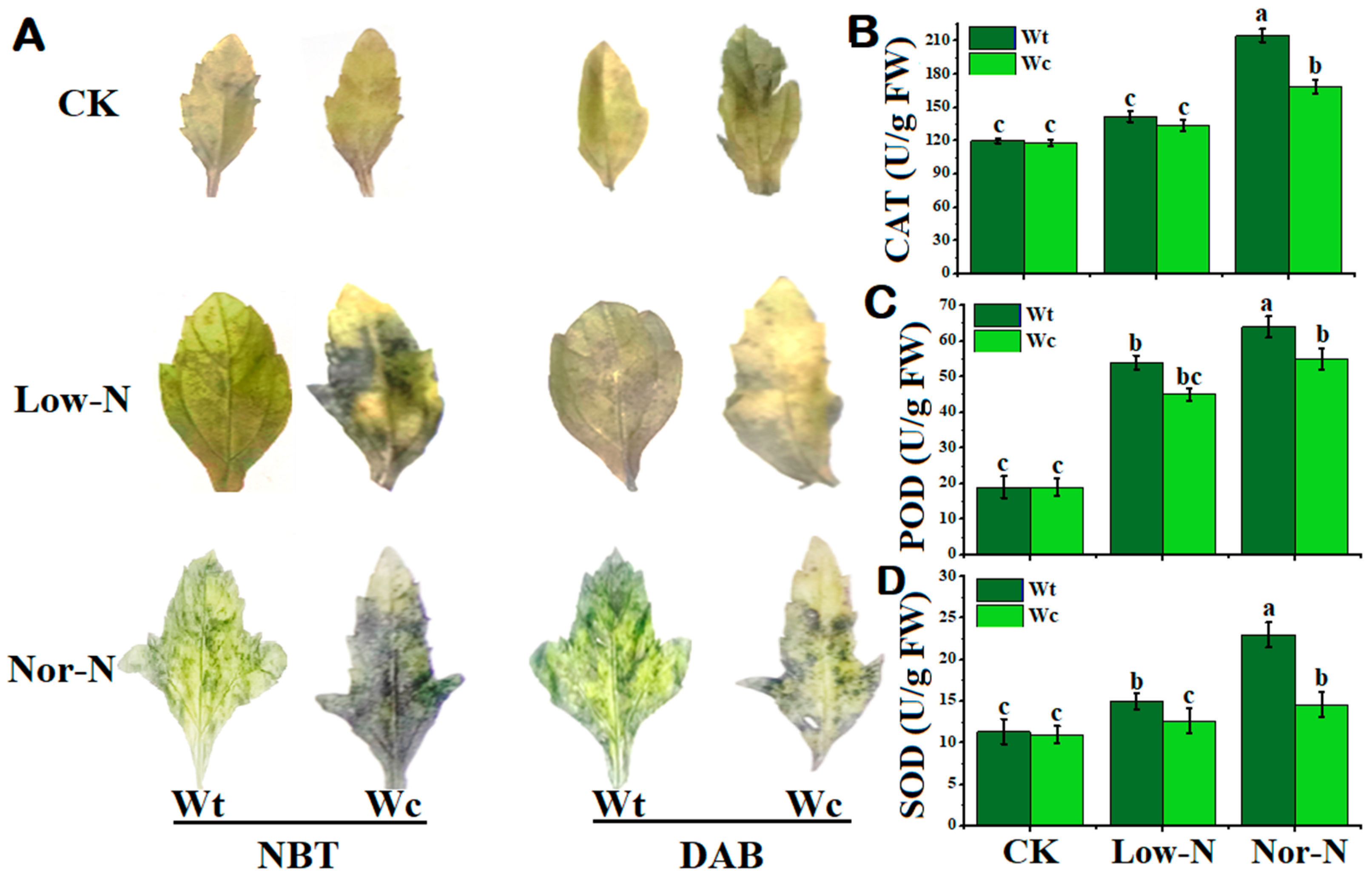
2.4. Detection of Reactive Oxygen Species and Antioxidant Enzyme Activities of Plants in N Conditions
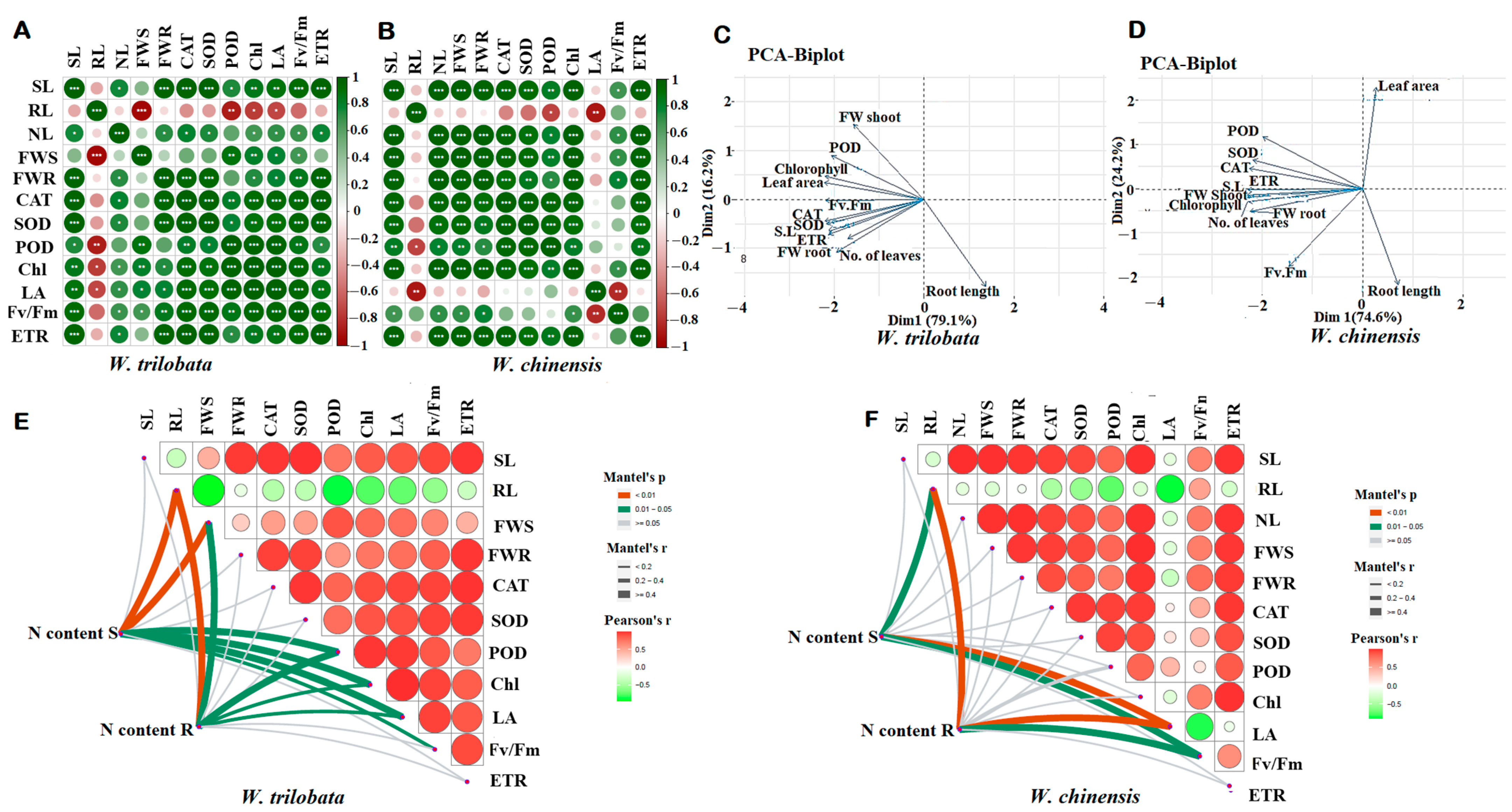
2.5. Correlation of Growth Traits and Physiological Activity
3. Discussion
3.1. The Growth Response of W. trilobata Is Better Than W. chinensis under Low-N Conditions
3.2. Physiological Response of W. trilobata to Low-N Conditions
3.3. Alleviation of ROS by Defensive Antioxidant Enzyme Activities
4. Materials and Methods
4.1. Experimental Design
4.2. Phenotypes and Growth Measurement
4.3. Analysis of Leaf Morphology
4.4. Analysis of Photosynthetic Parameters
4.5. Measurement of Nonenzymatic Flavonoid Content
4.6. Localization of ROS with NBT and DAB Solution
4.7. Analysis of Antioxidant Enzyme Activity
4.8. Statistical Analysis of the Study
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Asma Bano, S. Biological Nitrogen Fixation to Improve Plant Growth and Productivity Vaccine Designing View Project Antibiotic Producing Bacteria View Project. Int. J. Agric. Innov. Res. 2016, 4, 597–599. [Google Scholar]
- Bhuvaneswari, G.; Sivaranjani, R.; Reetha, S.; Ramakrishnan, K. Application of Nitrogen Fertilizer on Plant Density, Growth, Yield and Fruit of Bell Peppers (Capsicum annuum L.). Int. Lett. Nat. Sci. 2014, 13, 81–90. [Google Scholar] [CrossRef]
- Carranca, C.; Brunetto, G.; Tagliavini, M. Nitrogen Nutrition of Fruit Trees to Reconcile Productivity and Environmental Concerns. Plants 2018, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Yue, S.; Hou, P.; Cui, Z.; Chen, X. Improving Yield and Nitrogen Use Efficiency Simultaneously for Maize and Wheat in China: A Review. Pedosphere 2016, 26, 137–147. [Google Scholar] [CrossRef]
- Yotsova, E.; Dobrikova, A.; Stefanov, M.; Misheva, S.; Bardáčová, M.; Matušíková, I.; Žideková, L.; Blehová, A.; Apostolova, E. Effects of Cadmium on Two Wheat Cultivars Depending on Different Nitrogen Supply. Plant Physiol. Biochem. 2020, 155, 789–799. [Google Scholar] [CrossRef]
- Xuan, W.; Beeckman, T.; Xu, G. Plant Nitrogen Nutrition: Sensing and Signaling. Curr. Opin. Plant Biol. 2017, 39, 57–65. [Google Scholar] [CrossRef]
- Wang, W.; Zhu, Q.; Dai, S.; Meng, L.; He, M.; Chen, S.; Zhao, C.; Dan, X.; Cai, Z.; Zhang, J.; et al. Effects of Solidago Canadensis L. on Mineralization-Immobilization Turnover Enhance Its Nitrogen Competitiveness and Invasiveness. Sci. Total Environ. 2023, 882, 163641. [Google Scholar] [CrossRef]
- Xin, W.; Zhang, L.; Gao, J.; Zhang, W.; Yi, J.; Zhen, X.; Bi, C.; He, D.; Liu, S.; Zhao, X. Adaptation Mechanism of Roots to Low and High Nitrogen Revealed by Proteomic Analysis. Rice 2021, 14, 5. [Google Scholar] [CrossRef]
- Nadeem, F.; Mahmood, R.; Sabir, M.; Khan, W.u.D.; Haider, M.S.; Wang, R.; Zhong, Y.; Ishfaq, M.; Li, X. Foxtail Millet [Setaria italica (L.) Beauv.] over-Accumulates Ammonium under Low Nitrogen Supply. Plant Physiol. Biochem. 2022, 185, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Jia, X.; Huo, L.; Che, R.; Gong, X.; Wang, P.; Ma, F. MdATG18a Overexpression Improves Tolerance to Nitrogen Deficiency and Regulates Anthocyanin Accumulation through Increased Autophagy in Transgenic Apple. Plant Cell Environ. 2018, 41, 469–480. [Google Scholar] [CrossRef]
- Sousa, H.C.; Sousa, G.G.d.; Lessa, C.I.N.; Lima, A.F.d.S.; Ribeiro, R.M.R.; Rodrigues, F.H.d.C. Growth and Gas Exchange of Corn under Salt Stress and Nitrogen Doses TT—Crescimento e Trocas Gasosas Do Milho Sob Estresse Salino e Doses de Nitrogênio. Rev. Bras. Eng. Agrícola Ambient. 2021, 25, 174–181. [Google Scholar] [CrossRef]
- Gao, Y.; Sun, Y.; Ou, Y.; Zheng, X.; Feng, Q.; Zhang, H.; Fei, Y.; Luo, J.; Resco de Dios, V.; Yao, Y. Pretreating Poplar Cuttings with Low Nitrogen Ameliorates Salt Stress Responses by Increasing Stored Carbohydrates and Priming Stress Signaling Pathways. Ecotoxicol. Environ. Saf. 2021, 225, 112801. [Google Scholar] [CrossRef]
- Ullah, I.; Mao, H.; Rasool, G.; Gao, H.; Javed, Q.; Sarwar, A.; Khan, M.I. Effect of Deficit Irrigation and Reduced n Fertilization on Plant Growth, Root Morphology and Water Use Efficiency of Tomato Grown in Soilless Culture. Agronomy 2021, 11, 228. [Google Scholar] [CrossRef]
- Cruz-Alvarez, O.; Hernández-Rodríguez, O.A.; Jacobo-Cuellar, J.L.; Ávila-Quezada, G.; Morales-Maldonado, E.; Parra-Quezada, R.Á.; Robles-Hernandez, L.; Ojeda-Barrios, D.L. Nitrogen fertilization in pecan and its effect on leaf nutrient concentration, yield and nut quality. Rev. Chapingo Ser. Hort. 2020, 26, 163–173. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; Ali, D.F.I.; Xiong, Y.C.; Brestic, M.; Skalicky, M.; Hamoud, Y.A.; Ulhassan, Z.; Shaghaleh, H.; AbdElgawad, H.; Farooq, M.; et al. Physiological and Biochemical Responses of Soybean Plants Inoculated with Arbuscular Mycorrhizal Fungi and Bradyrhizobium under Drought Stress. BMC Plant Biol. 2021, 21, 195. [Google Scholar] [CrossRef] [PubMed]
- Vance, C.P. Symbiotic Nitrogen Fixation and Phosphorus Acquisition. Plant Nutrition in a World of Declining Renewable Resources. Plant Physiol. 2001, 127, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.E.; Barea, J.M.; McNeill, A.M.; Prigent-Combaret, C. Acquisition of Phosphorus and Nitrogen in the Rhizosphere and Plant Growth Promotion by Microorganisms. Plant Soil 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Kabbadj, A.; Makoudi, B.; Mouradi, M.; Pauly, N.; Frendo, P.; Ghoulam, C. Physiological and Biochemical Responses Involved in Water Deficit Tolerance of Nitrogen-Fixing Vicia Faba. PLoS ONE 2017, 12, e0190284. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Hu, Y.; Ma, J.Y.; Wang, H.; Wang, K.L.; Wang, T.; Jiang, S.Y.; Jiao, J.B.; Sun, Y.K.; Jiang, X.L.; et al. Nitrogen Deposition Effects on Invasive and Native Plant Competition: Implications for Future Invasions. Ecotoxicol. Environ. Saf. 2023, 259, 115029. [Google Scholar] [CrossRef]
- Luo, J.; Zhou, J.J.; Masclaux-Daubresse, C.; Wang, N.; Wang, H.; Zheng, B. Morphological and Physiological Responses to Contrasting Nitrogen Regimes in Populus Cathayana Is Linked to Resources Allocation and Carbon/Nitrogen Partition. Environ. Exp. Bot. 2019, 162, 247–255. [Google Scholar] [CrossRef]
- Guo, Q.; Song, H.; Kang, J.; Korpelainen, H.; Li, C. Different Responses in Leaf-Level Physiology to Competition and Facilitation under Different Soil Types and N Fertilization. Environ. Exp. Bot. 2018, 150, 69–78. [Google Scholar] [CrossRef]
- Wei, S.; Wang, X.; Shi, D.; Li, Y.; Zhang, J.; Liu, P.; Zhao, B.; Dong, S. The Mechanisms of Low Nitrogen Induced Weakened Photosynthesis in Summer Maize (Zea mays L.) under Field Conditions. Plant Physiol. Biochem. 2016, 105, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Qin, S.; Tang, J.; Chen, G.; Xie, J.; Chen, L.; Han, S.; Wang, X.; Zhu, T.; Liu, Y.; et al. Exogenous Nitrogen Enhances Poplar Resistance to Leaf Herbivory and Pathogen Infection after Exposure to Soil Cadmium Stress. Ecotoxicol. Environ. Saf. 2021, 208, 111688. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Xie, X.Y.; Qiu, J.; Fang, W.H.; Liang, R.; Ren, X.; Ji, X.; Cui, G.; Asiri, A.M.; Cui, G.; et al. High-Performance Artificial Nitrogen Fixation at Ambient Conditions Using a Metal-Free Electrocatalyst. Nat. Commun. 2018, 9, 3485. [Google Scholar] [CrossRef] [PubMed]
- Zou, N.; Huang, L.; Chen, H.; Huang, X.; Song, Q.; Yang, Q.; Wang, T. Nitrogen Form Plays an Important Role in the Growth of Moso Bamboo (Phyllostachys edulis) Seedlings. PeerJ 2020, 8, e9938. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chu, Y.; Huang, Q.; Ding, C.; Zhang, W.; Li, B.; Zhang, J.; Su, X. Morphological and Physiological Plasticity Response to Low Nitrogen Stress in Black Cottonwood (Populus deltoides Marsh.). J. For. Res. 2022, 33, 51–62. [Google Scholar] [CrossRef]
- Huang, P.; Shen, F.; Abbas, A.; Wang, H.; Du, Y.; Du, D.; Hussain, S.; Javed, T.; Alamri, S. Effects of Different Nitrogen Forms and Competitive Treatments on the Growth and Antioxidant System of Wedelia Trilobata and Wedelia Chinensis Under High Nitrogen Concentrations. Front. Plant Sci. 2022, 13, 851099. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Yang, J.X.; Zhou, X.H.; Peng, P.H.; Li, J.J.; Zhang, S.M.; He, W.M. An Invasive Population of Solidago Canadensis Is Less Sensitive to Warming and Nitrogen-Addition than Its Native Population in an Invaded Range. Biol. Invasions 2019, 21, 151–162. [Google Scholar] [CrossRef]
- Qi, S.S.; Dai, Z.C.; Miao, S.L.; Zhai, D.L.; Si, C.C.; Huang, P.; Wang, R.P.; Du, D.L. Light Limitation and Litter of an Invasive Clonal Plant, Wedelia Trilobata, Inhibit Its Seedling Recruitment. Ann. Bot. 2014, 114, 425–433. [Google Scholar] [CrossRef]
- Cai, M.L.; Zhang, Q.L.; Zhang, J.J.; Ding, W.Q.; Huang, H.Y. Comparative Physiological and Transcriptomic Analyses of Photosynthesis in Sphagneticola calendulacea (L.) Pruski and Sphagneticola trilobata (L.) Pruski. Sci. Rep. 2020, 10, 17810. [Google Scholar] [CrossRef]
- Sun, J.; Rutherford, S.; Saif Ullah, M.; Ullah, I.; Javed, Q.; Rasool, G.; Ajmal, M.; Azeem, A.; Nazir, M.J.; Du, D. Plant-Soil Feedback during Biological Invasions: Effect of Litter Decomposition from an Invasive Plant (Sphagneticola trilobata) on Its Native Congener (S. calendulacea). J. Plant Ecol. 2022, 15, 610–624. [Google Scholar] [CrossRef]
- Zhang, L.D.; Song, L.Y.; Dai, M.J.; Guo, Z.J.; Wei, M.Y.; Li, J.; Xu, C.Q.; Zhu, X.Y.; Zheng, H.L. Cadmium Promotes the Absorption of Ammonium in Hyperaccumulator Solanum nigrum L. Mediated by Ammonium Transporters and Aquaporins. Chemosphere 2022, 307, 136031. [Google Scholar] [CrossRef]
- Zhang, Q.; Huang, J.; Ke, W.; Cai, M.; Chen, G.; Peng, C. Responses of Sphagneticola trilobata, Sphagneticola calendulacea and Their Hybrid to Drought Stress. Int. J. Mol. Sci. 2021, 22, 1288. [Google Scholar] [CrossRef]
- Azeem, A.; Wenxuan, M.; Changyan, T.; Javed, Q.; Abbas, A. Competition and plant trait plasticity of invasive (Wedelia trilobata) and native species (Wedelia chinensis, WC) under nitrogen enrichment and flooding condition. Water 2021, 13, 3472. [Google Scholar] [CrossRef]
- Ke, W.Q.; Pan, Y.R.; Chen, L.H.; Huang, J.D.; Zhang, J.J.; Long, X.Y.; Cai, M.L.; Peng, C.L. Adaptive Photosynthetic Strategies of the Invasive Plant Sphagneticola Trilobata and Its Hybrid to a Low-Light Environment. Photosynthetica 2022, 60, 549–561. [Google Scholar] [CrossRef]
- Khan, I.U.; Rono, J.K.; Liu, X.S.; Feng, S.J.; Li, H.; Chen, X.; Yang, Z.M. Functional Characterization of a New Metallochaperone for Reducing Cadmium Concentration in Rice Crop. J. Clean. Prod. 2020, 272, 123152. [Google Scholar] [CrossRef]
- Lin, Z.H.; Chen, C.S.; Zhao, S.Q.; Liu, Y.; Zhong, Q.S.; Ruan, Q.C.; Chen, Z.H.; You, X.M.; Shan, R.Y.; Li, X.L.; et al. Molecular and Physiological Mechanisms of Tea (Camellia sinensis (L.) O. Kuntze) Leaf and Root in Response to Nitrogen Deficiency. BMC Genom. 2023, 24, 27. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hu, D.; Shi, X.; Luo, J.; Ren, G.; Dai, Z.; Qi, S.; Du, D. Different Responses of Invasive Weed Alternanthera Philoxeroides and Oryza Sativa to Plant Growth Regulators. Life 2022, 12, 1069. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Gong, X.; Wang, H.; Dang, K.; Deng, X.; Feng, B. Low-Nitrogen Tolerance Comprehensive Evaluation and Physiological Response to Nitrogen Stress in Broomcorn Millet (Panicum miliaceum L.) Seedling. Plant Physiol. Biochem. 2020, 151, 233–242. [Google Scholar] [CrossRef]
- Sun, X.; Chen, H.; Wang, P.; Chen, F.; Yuan, L.; Mi, G. Low Nitrogen Induces Root Elongation via Auxin-Induced Acid Growth and Auxin-Regulated Target of Rapamycin (TOR) Pathway in Maize. J. Plant Physiol. 2020, 254, 153281. [Google Scholar] [CrossRef]
- Luo, J.S.; Zhang, Z. Proteomic Changes in the Xylem Sap of Brassica Napus under Cadmium Stress and Functional Validation. BMC Plant Biol. 2019, 19, 280. [Google Scholar] [CrossRef]
- Gul, F.; Khan, I.U.; Rutherford, S.; Dai, Z.C.; Li, G.; Du, D.L. Plant growth promoting rhizobacteria and biochar production from Parthenium hysterophorus enhance seed germination and productivity in barley under drought stress. Front. Plant Sci. 2023, 14, 1175097. [Google Scholar] [CrossRef]
- Khan, I.U.; Qi, S.-S.; Gul, F.; Manan, S.; Rono, J.K.; Naz, M.; Shi, X.-N.; Zhang, H.; Dai, Z.-C.; Du, D.-L. A Green Approach Used for Heavy Metals ‘Phytoremediation’ Via Invasive Plant Species to Mitigate Environmental Pollution: A Review. Plants 2023, 12, 725. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, C.; Xiong, Y.; Wang, Y.; Li, Q. Combination Effects of Heavy Metal and Inter-Specific Competition on the Invasiveness of Alternanthera Philoxeroides. Environ. Exp. Bot. 2021, 189, 104532. [Google Scholar] [CrossRef]
- Tahir, M.; Ahmad, I.; Shahid, M.; Shah, G.M.; Farooq, A.B.U.; Akram, M.; Tabassum, S.A.; Naeem, M.A.; Khalid, U.; Ahmad, S.; et al. Regulation of Antioxidant Production, Ion Uptake and Productivity in Potato (Solanum tuberosum L.) Plant Inoculated with Growth Promoting Salt Tolerant Bacillus Strains. Ecotoxicol. Environ. Saf. 2019, 178, 33–42. [Google Scholar] [CrossRef]
- Javed, Q.; Sun, J.; Rutherford, S.; Li, J.; Iqbal, B.; Xiang, Y.; Ren, G.; He, F.; Pan, L.; Bo, Y.; et al. Soil Pollution and the Invasion of Congener Sphagneticola in Crop Lands. J. Environ. Manag. 2023, 340, 118013. [Google Scholar] [CrossRef]
- Cai, M.L.; Ding, W.Q.; Zhai, J.J.; Zheng, X.T.; Yu, Z.C.; Zhang, Q.L.; Lin, X.H.; Chow, W.S.; Peng, C.L. Photosynthetic Compensation of Non-Leaf Organ Stems of the Invasive Species Sphagneticola trilobata (L.) Pruski at Low Temperature. Photosynth. Res. 2021, 149, 121–134. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Bassi, D.; Menossi, M.; Mattiello, L. Nitrogen Supply Influences Photosynthesis Establishment along the Sugarcane Leaf. Sci. Rep. 2018, 8, 2327. [Google Scholar] [CrossRef] [PubMed]
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Møller, I.S.; White, P. Functions of Macronutrients; Elsevier Ltd.: Amsterdam, The Netherlands, 2011; ISBN 9780123849052. [Google Scholar]
- Zhu, Y.; Fan, X.; Hou, X.; Wu, J.; Wang, T. Effect of Different Levels of Nitrogen Deficiency on Switchgrass Seedling Growth. Crop J. 2014, 2, 223–234. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive Oxygen Species and Antioxidant Machinery in Abiotic Stress Tolerance in Crop Plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Tomar, N.S.; Tittal, M.; Argal, S.; Agarwal, R.M. Plant Growth under Water/Salt Stress: ROS Production; Antioxidants and Significance of Added Potassium under Such Conditions. Physiol. Mol. Biol. Plants 2017, 23, 731–744. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A. Modulation of Phytoremediation and Plant Growth by the Treatment with PGPR, Ag Nanoparticle and Untreated Municipal Wastewater. Int. J. Phytoremediation 2016, 18, 1258–1269. [Google Scholar] [CrossRef]
- Fu, W.; Huang, K.; Cai, H.H.; Li, J.; Zhai, D.L.; Dai, Z.C.; Du, D.L. Exploring the Potential of Naturalized Plants for Phytoremediation of Heavy Metal Contamination. Int. J. Environ. Res. 2017, 11, 515–521. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Modulation of Antioxidant Defense System Is Associated with Combined Drought and Heat Stress Tolerance in Citrus. Front. Plant Sci. 2017, 8, 953. [Google Scholar] [CrossRef] [PubMed]
- Farid, M.; Sajjad, A.; Asam, Z.U.Z.; Zubair, M.; Rizwan, M.; Abbas, M.; Farid, S.; Ali, S.; Alharby, H.F.; Alzahrani, Y.M.; et al. Phytoremediation of Contaminated Industrial Wastewater by Duckweed (Lemna minor L.): Growth and Physiological Response under Acetic Acid Application. Chemosphere 2022, 304, 135262. [Google Scholar] [CrossRef] [PubMed]
- Chu, X.; Wang, J.G.; Li, M.; Zhang, S.; Gao, Y.; Fan, M.; Han, C.; Xiang, F.; Li, G.; Wang, Y.; et al. HBI Transcription Factor-Mediated ROS Homeostasis Regulates Nitrate Signal Transduction. Plant Cell 2021, 33, 3004–3021. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.D.; Liu, X.; Wei, M.Y.; Guo, Z.J.; Zhao, Z.Z.; Gao, C.H.; Li, J.; Xu, J.X.; Shen, Z.J.; Zheng, H.L. Ammonium Has Stronger Cd Detoxification Ability than Nitrate by Reducing Cd Influx and Increasing Cd Fixation in Solanum nigrum L. J. Hazard. Mater. 2022, 425, 127947. [Google Scholar] [CrossRef]
- Iqbal, B.; Khan, I.; Javed, Q.; Alabbosh, K.; Inamullah, I.; Zhou, Z.; Rehman, A. The High Phosphorus Incorporation Promotes the Soil Enzymatic Activity, Nutritional Status, and Biomass of the Crop. Polish J. Environ. Stud. 2023, 32, 2125–2139. [Google Scholar] [CrossRef]
- Qi, S.; Wang, J.; Wan, L.; Dai, Z.; da Silva Matos, D.M.; Du, D.; Egan, S.; Bonser, S.P.; Thomas, T.; Moles, A.T. Arbuscular Mycorrhizal Fungi Contribute to Phosphorous Uptake and Allocation Strategies of Solidago Canadensis in a Phosphorous-Deficient Environment. Front. Plant Sci. 2022, 13, 831654. [Google Scholar] [CrossRef]
- Khan, I.U.; Zhang, Y.F.; Shi, X.N.; Qi, S.S.; Zhang, H.Y.; Du, D.L.; Gul, F.; Wang, J.H.; Naz, M.; Shah, S.W.A.; et al. Dose dependent effect of nitrogen on the phyto extractability of Cd in metal contaminated soil using Wedelia trilobata. Ecotoxicol. Environ. Saf. 2023, 264, 115419. [Google Scholar] [CrossRef] [PubMed]
- Khattak, W.A.; He, J.; Abdalmegeed, D.; Hu, W.; Wang, Y.; Zhou, Z. Foliar Melatonin Stimulates Cotton Boll Distribution Characteristics by Modifying Leaf Sugar Metabolism and Antioxidant Activities during Drought Conditions. Physiol. Plant. 2022, 174, e13526. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, H.; Chen, J.; Zhang, S.; Xu, J.; Han, X.; Feng, Y.; Chen, Y.; Zhang, X.; Dong, G.; et al. Xylem Development, Cadmium Bioconcentration, and Antioxidant Defense in Populus × Euramericana Stems under Combined Conditions of Nitrogen and Cadmium. Environ. Exp. Bot. 2019, 164, 1–9. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | CK | Low-N | Nor-N | |||
|---|---|---|---|---|---|---|
| Plant Species | Wt | Wc | Wt | Wc | Wt | Wc |
| No of leaves | 8 d | 8 d | 12 b | 10 c | 53 a | 48 ab |
| FW of leaves (g) | 0.28 e | 0.26 e | 0.63 c | 0.56 d | 8.8 a | 6.9 b |
| DW of leaves (g) | 0.07 e | 0.025 e | 0.085 c | 0.06 d | 0.87 a | 0.6 b |
| No. of roots | 7 c | 8 c | 13 a | 10 b | 15 a | 14 a |
| FW of roots (g) | 0.34 b | 0.33 b | 0.27 b | 0.2 c | 1.7 a | 1.36 ab |
| DW of roots (g) | 0.031 b | 0.03 b | 0.031 b | 0.025 c | 0.1 a | 0.09 a |
| Root surface area (cm2) | 16 d | 15 d | 25 c | 15 d | 42 a | 35 b |
| N content in leaves (mg/g) | 4.66 b | 4 b | 7.33 ab | 5.33 b | 11.33 a | 10.67 a |
| N content in roots (mg/g) | 6 d | 5 d | 10 c | 7 d | 13.6 a | 11 b |
| Serial No. | Treatments | CaNO3 (mg kg−1) |
|---|---|---|
| 1 | Wt-CK | 0 (only water) |
| 2 | Wc-CK | 0 (only water) |
| 3 | Wt-Nor-N | 91.05 (with Hoagland) |
| 4 | Wc-Nor-N | 91.05 (with Hoagland) |
| 5 | Wt-Low-N | 0.9105 (with Hoagland) |
| 6 | Wc-Low-N | 0.9105 (with Hoagland) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, Z.-C.; Kong, F.-L.; Li, Y.-F.; Ullah, R.; Ali, E.A.; Gul, F.; Du, D.-L.; Zhang, Y.-F.; Jia, H.; Qi, S.-S.; et al. Strong Invasive Mechanism of Wedelia trilobata via Growth and Physiological Traits under Nitrogen Stress Condition. Plants 2024, 13, 355. https://doi.org/10.3390/plants13030355
Dai Z-C, Kong F-L, Li Y-F, Ullah R, Ali EA, Gul F, Du D-L, Zhang Y-F, Jia H, Qi S-S, et al. Strong Invasive Mechanism of Wedelia trilobata via Growth and Physiological Traits under Nitrogen Stress Condition. Plants. 2024; 13(3):355. https://doi.org/10.3390/plants13030355
Chicago/Turabian StyleDai, Zhi-Cong, Fang-Li Kong, Yi-Fan Li, Riaz Ullah, Essam A. Ali, Farrukh Gul, Dao-Lin Du, Yi-Fan Zhang, Hui Jia, Shan-Shan Qi, and et al. 2024. "Strong Invasive Mechanism of Wedelia trilobata via Growth and Physiological Traits under Nitrogen Stress Condition" Plants 13, no. 3: 355. https://doi.org/10.3390/plants13030355
APA StyleDai, Z.-C., Kong, F.-L., Li, Y.-F., Ullah, R., Ali, E. A., Gul, F., Du, D.-L., Zhang, Y.-F., Jia, H., Qi, S.-S., Uddin, N., & Khan, I. U. (2024). Strong Invasive Mechanism of Wedelia trilobata via Growth and Physiological Traits under Nitrogen Stress Condition. Plants, 13(3), 355. https://doi.org/10.3390/plants13030355