Abstract
Moreyra and co-authors, in their recent paper published in Plants, presented a desperately needed Hyb-Seq phylogeny of the genus Cirsium sensu lato. Through their findings, they provided a critical assessment of an earlier proposal of ours to separate Cirsium sect. Eriolepis and Cirsium italicum into the genera Lophiolepis and Epitrachys, respectively. While we fully respect their right not to accept our proposal, we cannot remain silent to their arguments that not only misinterpret our findings and statements but are often in direct conflict with their own results that actually support our proposal to segregate Lophiolepis. In addition, as Moreyra and co-authors did not include Epitrachys italica (formerly Cirsium italicum) in their analyses; they have no basis for arguing in favour or against our proposal to separate it as a monotypic genus. Finally, we conclude that the genus Cirsium, as circumscribed by the above-mentioned authors (i.e., including Lophiolepis), is definitively not monophyletic.
1. Introduction
Recent phylogenetic investigations [1,2,3,4] have unveiled the polyphyletic nature of a broadly circumscribed genus Cirsium Mill. This is especially evident when other traditionally recognized genera—Carduus L., Galactites Moench, Lamyropsis (Kharadze) Dittrich, Nothobasis (Cass.) Cass., Picnomon Adans., Silybum Adans., and Tyrimnus (Cass.) Bosc—are to be preserved. In the light of these findings, we proposed [3] the separation of two genera from the species-rich Cirsium genus, namely Lophiolepis Cass. (encompassing 104 Eurasian and North African species) and Epitrachys (DC. ex Duby) K.Koch, consisting of a single Mediterranean species. Later, Moreyra et al. [1] proposed the separation of two Central African genera, namely Afrocirsium Calleja, Garcia-Jacas, Moreyra & Susanna (with three species) and Nuriaea Susanna, Calleja & Moreyra (with two).
Problems with a clear morphological distinction between Lophiolepis and Cirsium are mainly related to the intermediate features displayed by Cirsium vulgare (Savi) Ten. and have been addressed by Bureš et al. [5]. These authors identified C. vulgare as an established intergeneric allopolyploid species and segregated it into the monotypic Ascalea Hill [5,6]. It is important to note that the segregation of Lophiolepis is supported by both phylogenetic evidence and morphological data [1,3,4], as well as by genomic features [4].
Surprisingly, Moreyra et al. [1] strongly dissented with this choice and argued, somehow polemically, against the generic distinctness of Lophiolepis. As we are convinced that, on some occasions, they object to our proposal by using inconsistent or unsupported arguments, we are forced to rebut their objections.
2. Results and Discussion
2.1. Scopes and Limits of Del Guacchio et al. [3]
Moreyra and co-authors [1] (p. 13) stated that “[…] the published phylogeny [by [3]] fails to support the classification proposed [by [3]] since Cirsium s. str. is not recovered as monophyletic as the entire genus Carduus was nested within Cirsium s. str. In addition, two species of African Carduus, now Afrocarduus, are recovered within Cirsium s. str. [as intended by [3]], Figure S1, whereas our study reveals that African species of Carduus, now Afrocarduus, are a group evolutionary [sic!] independent from Cirsium”. This is a misleading claim, because we [3] neither proposed a stand-alone Cirsium including Carduus, nor regarded Cirsium (after the exclusion of Lophiolepis and Epitrachys) as a monophyletic genus; above all, we never proposed any “Cirsium s. str.”. In [3], we only proposed a generic recognition of Lophiolepis and Epitrachys and did not infer any conclusion regarding the monophyly of the group, including the remaining Cirsium species. This is obvious throughout our entire contribution, from the title itself [3] (“Towards a monophyletic classification of Cardueae”); from the first sentence of the abstract (“Using molecular data and representative species coverage, we confirmed the monophyly of Cirsium sect. Eriolepis and, therefore, we propose to treat it as a separate genus (Lophiolepis)“); and even from the concluding paragraph of the introduction, which states that “[...] this paper principally aims at the nomenclatural revision of the species belonging to Cirsium sect. Eriolepis [...]”. Therefore, our work [3] was clearly conceived only as a first step towards a modern classification of Cirsium s. lat. We would also like to note that according to our phylogeny [3] (Suppl. fig. S2), the two Afrocarduus sequences (retrieved from NCBI) were robustly grouped together but in a poorly supported and resolved clade with a few other Cirsium species; this clade, in turn, was nested in a very large and unresolved group including not only Cirsium (excluded Lophiolepis) but also Carduus subg. Carduus and Tyrimnus (see also Figure 1d). Therefore, on the one hand, our study [3] was doubtlessly not sufficient for any inference about Carduus subg. Afrocarduus Kazmi and even less for any reassessment of Cirsium s. lat. (issues which were never among our aims); on the other hand, the two Afrocarduus were confirmed to be not monophyletic with Carduus (cf. also in [2]).
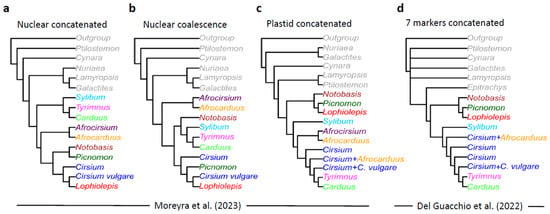
Figure 1.
Comparison of phylogenetic trees (simplified and relabeled) from Moreyra et al. (2023) [1] (a–c) and Del Guacchio et al. (2022) [3] (d). In (d), each Cirsium tip (blue) represents several polytomic branches that we merged into one for clarity’s sake. Only topologies are shown, while branch lengths are arbitrary.
In conclusion, we do not understand how our study [3] would support or propose a monophyletic Cirsium s. str., even excluding Epitrachys and Lophiolepis, or how it would contradict an independent evolution of Afrocarduus.
2.2. The Separation of Lophiolepis Is Not Based on “Weakly Supported Molecular Results”
Moreyra et al. [1] (p. 2) wrote that “The recent taxonomic proposal for the Carduus-Cirsium group [by [3]] split Cirsium into four genera, accepting Cassini’s subgenus Lophiolepis at the generic level, reinstating the genus Epitrachys (DC. Ex [!] Duby) K.Koch and describing the hybrid genus xLophiocirsium Del Guacchio, Bureš, Iamonico & P. Caputo, mainly based on the weakly supported molecular results from Ackerfield et al. (2020)” and again [1] (p. 13) “Recently, the authors of [[3]] split Cirsium into four genera (Cirsium s. str., Lophiolepis, Epitrachys and Lophiocirsium). This work encompasses a remarkable number of species (n = 225), yet it is based only on two nuclear and five plastid markers that have proven to be insufficient for phylogenetic resolution in this genus [[2]].”
First of all, we take issue with the claim that we split Cirsium “into four genera”. In fact, xLophiocirsium is a nothogenus and cannot be included in this count; in addition, we simply proposed [3] the separation of Cirsium sect. Eriolepis and Cirsium italicum DC. at a generic rank from a polyphyletic Cirsium s. lat. The proposal was not even new, because Lophiolepis (and in a certain sense, Epitrachys) had already been recognized at a generic level in the past, e.g., [7,8,9].
Second, five authors [2], who are also co-authors of [1], used only six markers (two nuclear and four plastid); therefore, [2] simply could not prove the insufficiency of the seven markers we used in [3,4], despite what stated by Moreyra et al. [1].
Third, one of the key results of [2] (p. 722, Figures 2 and 3) was a clear phylogenetic delimitation of the so-called “clade one”, comprising a monophyletic “Eriolepis” (= Lophiolepis) and the two monotypic genera Nothobasis and Picnomon. The unexpected placement of Cirsium vulgare and C. cephalotes in “clade two” in [2] (i.e., Carduus + the Cirsium species not included in Lophiolepis) and not in “clade one” [2] actually does not challenge the monophyly of Lophiolepis in any way. In fact, C. vulgare has been suggested [1,4] and later proven [5] to be of hybridogenous origin, deriving from a cross between Lophiolepis and Cirsium, so its incongruent position in the phylogenetic trees is fully justified. Regarding the “C. cephalotes” specimen employed by [2], we suggest that it was likely misidentified, because in subsequent studies, the newly sequenced individuals of C. cephalotes were nested within Lophiolepis [3,4], which also happened in the Hyb-Seq phylogeny by Moreyra et al. [1] themselves.
Fourth, our phylogeny [3] was based on 905 sequences, of which 95 (10.50%) were generated by [2], 107 (11.82%) by the group of the same leading authors as [1,2], and 289 (31.93%) from other sources (all were downloaded from NCBI), while 414 (45.75%) were newly generated by our research groups. The tree in [3] includes 255 tips covering 236 taxa and 225 species, while that in [2] includes 173 tips covering 164 taxa and 154 species. Taken together, the claim of Moreyra et al. [1] that our results [3] are “mainly based on the weakly supported molecular results from Ackerfield et al. (2020)” is therefore inaccurate and misleading.
Fifth, Moreyra et al. [1] (pp. 13) further argue that “[...] according to the Supplementary Material [by us [3]], Table S2, the matrix lacks almost 50% of sequences (880 out of 1785)”. This observation, correct per se, may seem to captiously insinuate that our phylogeny has been heavily affected by missing sequences. Actually, using datasets with even far more than 50% of missing data is a common practice [10]. More importantly, the IQ-tree version v2.1.3, which was used in [3] for tree construction, systematically accounts for missing data by implementing terrace-aware computational approaches, and if a problem is found, users are advised to gather more data or filter out gappy taxa/loci [11].
Moreover, despite the statements by [1], our topology [3] largely concurs with their plastid phylogeny (Figure 1c,d), which would be obvious if the Lophiolepis clade is labelled as such in the trees, something that Moreyra et al. [1] decided not to carry out in their figures. This similarity seems final to us in order to reject any alleged inadequateness of our results to reach the proposed aim of our previous study, i.e., to characterize the Lophiolepis clade.
In conclusion, the separation of Lophiolepis from Cirsium has been clearly supported in an independent fashion by [2,3], as well as by Moreyra et al. [1] themselves (Figure 1).
2.3. There Is No Evidence of an Extremely Frequent Hybridization Between Cirsium and Lophiolepis
Moreyra et al. [1] (p. 13) stated that according to [3,4] “hybrids between the two largest new genera [i.e., Lophiolepis and Cirsium] proposed in [[3]] are extremely frequent” and employed this argument against the separation of Lophiolepis from Cirsium.
In [3], we made the following comment on intergeneric hybridization: “Numerous intergeneric hybrids have been described between Lophiolepis and Cirsium (especially C. vulgare)”. Although we admit that the word “numerous” is ambiguous, we intended to refer to the effectively frequent records of presumed intergeneric hybrids (partly listed in [3]). In fact, we did recombine only one name, because most of the remaining ones were misapplied to taxa which are not of hybrid origin or which require further study. Indeed, later [5], we recognized only four intergeneric hybrids. In [4], hybridization is only mentioned as a process detectable by flow cytometry. It is to be noted, however, that most of the presumed or ascertained hybrids actually involve Cirsium vulgare as a parent, which is now recognized as an established hybrid between Lophiolepis and Cirsium lineages (see discussion in [5]). Furthermore, we have demonstrated [5] that Cirsium vulgare, i.e., Ascalea lanceolata (L.) Hill, hybridizes with species of the parental lineages (i.e., Lophiolepis and Cirsium) more frequently than these genera do with each other, as is expected as a consequence of its intergeneric hybridogenous nature. In addition, the propensity to form intergeneric hybrids between Lophiolepis taxa and the remaining Cirsium species is significantly lower than the propensity to form interspecific hybrids within these genera ([5], Table 1). Although the results of [5] were not available to Moreyra et al. [1], the claim of [1] that “hybrids between the two largest new genera [Lophiolepis and Cirsium] [...] are extremely frequent” while citing [3,4] is an overinterpretation of what those studies actually say. Moreover, Moreyra et al. [1] do not take into account that these intergeneric hybrids are overall rare and ephemeral. Moreover, although intergeneric hybrids are also reported, for example, between Cirsium and Carduus (see [12] (p. 206), [13,14,15,16]), this was never used as an argument against the taxonomic distinctness of these genera.
2.4. The Carduineae Treatment by Moreyra et al. [1] Is Inconsistent and Contradictory
A relevant inconsistency of the treatment by Moreyra et al. [1] is their choice of keeping together Lophiolepis and Cirsium as subgenera but segregating the small genera Afrocarduus and Afrocirsium at the same time. Incidentally, this latter choice is acceptable in our opinion but falls in contradiction with the former. In fact, Afrocarduus and Afrocirsium (a) are sister groups in each phylogenetic reconstruction published by [1], and (b) can be morphologically separated chiefly (or exclusively!) on account of their phyllary appendages (absent vs. fimbriate) and, less clearly, by characters of the pappus bristles (scabrid-short barbellate vs. plumose) and stems (unwinged or interruptedly winged vs. winged). On the contrary, Moreyra et al. [1] reject Lophiolepis as a distinct genus, but the latter (a) is not the sister group of Cirsium in two out of the three topologies of theirs (one nuclear, the other plastidial!), and (b) has been proven to differ morphologically from Cirsium by more numerous and relevant characters [3,4,5]. The presence/absence of setae on leaves, for example, is universally recognized as diagnostic between Cirsium and Lophiolepis (e.g., [17,18,19,20,21,22,23]), and several other morphological, anatomical, phenological, karyological, and genomic differences are known (see [5] for details), even if we agree that a wider sampling is necessary to verify character distribution within the genera.
We also add that according to Moreyra et al. [1], the three species segregated by them into Afrocirsium “show distinctive characters such as phyllaries with well-developed pectinate appendages, which are absent in Cirsium and in all other genera in the Carduus-Cirsium group […]. However, we consider the presence of a unique diagnostic morphological character combined with strong evidence of being an independent evolutionary lineage to be sufficient to propose a new genus, Afrocirsium”. Actually, such “unique” autapomorphy, i.e., pectinate appendages, occurs, for example, in Cirsium echinus (M.Bieb.) Hand.-Mazz., C. sieversii (Fisch. & C.A.Mey.) Petr., C. griffithii Boiss., C. swaticum Petr., and C. wallichii DC. (the first two species were also included in the dataset of [1]).
Another inconsistency in Moreyra et al. [1] concerns the treatment of intergeneric hybridogenous species. As already discussed, these authors incorrectly reject Lophiolepis also based on the presumed “extremely frequent hybrids” and the occurrence of the established hybrid C. vulgare itself [1]. However, their own analyses strongly suggest that hybridization also occurred between Cirsium and Afrocirsium, namely in the origin of Afrocirsium straminispinum (C.Jeffrey) Calleja, Garcia-Jacas, Moreyra & Susanna, i.e., one of the only three species of Afrocirsium. This presumed hybrid origin was admitted by them, who, however, did not show any doubt in segregating Afrocirsium despite this!
As a further inconsistency, it is to be noted that the phylogenetic reconstructions by Moreyra et al. [1] do not include Cirsium italicum (i.e., the only representative of the crucial genus Epitrachys in our treatment [3]), which has been proven to clearly differ from the taxa involved in this discussion [3,4].
On the contrary, Lophiolepis is a rather homogeneous, even speciose group, which likely originated in mountain environments of western Asia–eastern Mediterranean, where it is most diversified [3] (Supplementary Figure S7) and adapted to seasonal drought and grazing, also under human influence. Its reduction to a subgenus of Cirsium does not do justice to its natural history and would not help in understanding the reticulate evolution of Carduineae.
2.5. The Genus Cirsium, as Circumscribed by Moreyra et al. [1], Is Definitively Polyphyletic as a Consequence of Their Own Results
As said, Moreyra et al. [1] fiercely refuse the recognition of Lophiolepis at a generic rank; in doing so, however, they completely disregard their own results, especially those obtained by the plastid dataset, where Lophiolepis clearly shows a different origin as compared to other Cirsium (even after excluding Nuriaea, Afrocirsium, and Afrocarduus) (Figure 1c). Curiously, this is not apparent from the main body of the text, where Moreyra et al. [1] (Figure 3) showed a concatenated chloroplast tree exhibiting a polyphyletic Cirsium s. lat. However, if we examine their Supplementary Figure S2, we find that the Cirsium clade most distant from the others in their Figure 3 includes all Lophiolepis taxa and only them (!). In that reconstruction, Lophiolepis resulted as sister to Picnomon, and, in turn, both to Notobasis. All the remaining Cirsium taxa (including C. vulgare), represented in their figure by three groups, are paraphyletic, as one of the Cirsium clades is a sister to Carduus and Tyrimnus (Figure 1c).
From what is said above, we may suppose that the maternal lineage of Lophiolepis has a last common ancestor with Picnomon and Notobasis, which is not shared by the paternal line. The latter is largely shared with Cirsium s. str., but evidently diverged over time, as the two clades (i.e., Cirsium s.str. and Lophiolepis) in nuclear reconstructions are never admixed. This implies that the evolution of Lophiolepis has been different and largely independent from that of Cirsium s. str.
Therefore, based on a comparison of the incongruent concatenated phylogenies (nuclear and plastidial) in [1], we are forced to conclude that a genus Cirsium as conceived by them (i.e., including Lophiolepis) cannot be accepted simply because it would not be monophyletic.
2.6. Phyletic Classification and Name Inflation
Finally, quoting Moreyra et al. [1]: “...our conservative classification maintaining Cirsium as a single genus is also the most robust and operational one, because it avoids the inflation of hundreds of new nomenclatural combinations that would increase the already voluminous synonymy of Cirsium”. Nevertheless, this does not prevent these authors from providing several new combinations as well [1]. The dreaded “hundreds of new nomenclatural combinations” (actually 130) had already been provided [3] and are now easily available [24].
In this regard, it might be useful to note that according to the previous contribution of the same group [2], “An alternative solution [i.e., alternative to combine all genera in a single genus] is to recognize each major clade of the phylogeny as a genus. This would result in recognition of seven genera: Carduus, Cirsium, Eriolepis [= Lophiolepis in our treatment [3]!], Notobasis, Picnomon, Silybum, and Tyrimnus”. The same authors also provided a table with additional morphological differences between Eriolepis (= Lophiolepis) and Cirsium and foresaw a future (at the time) segregation of African mountain thistles. However, from a mere nomenclatural point of view, it is a fortunate case that they refrained from segregating Cirsium sect. Eriolepis on that occasion, considering that otherwise, they might have provided numerous erroneous combinations under Eriolepis [2], entirely neglecting the priority of Lophiolepis [25].
3. Material and Methods
This response is based on recent achievements in Carduineae taxonomy and evolution [2,3,4,5] and a thorough examination of [1], including their Supplementary Materials. For the sake of clarity, we refer in the text to Cirsium sensu lato, the genus traditionally conceived [17,18,19,20,21,22], i.e., including Lophiolepis, Epitrachys, Nuriaea, and Afrocirsium; and to Cirsium sensu stricto, intending to use the same genus not including those taxa (this, however, would not necessarily imply any assessment about its monophyly). In order to facilitate discussion, in Figure 1, we compared the phylogenetic trees proposed by [1,3], merging monophyletic groups under single labels.
4. Conclusions
The Carduus-Cirsium group presents a substantial phylogenetic challenge. The path to a taxonomic solution to this vast problem necessarily requires successive steps, some of which have already been taken. A full taxonomic resolution of Cirsium s. lat. may only arise from greater coverage and the application of phylogenomic techniques, as proposed by Moreyra et al. [1]. We believe this team of colleagues, whose experience and great results we sincerely appreciate, will succeed.
Their concept of keeping a wide concept genus Cirsium [1] is altogether respectable. Still, the only reliable alternative to the recognition of the genus Lophiolepis is merging it into Picnomon, a possibility already discussed but discouraged by us [3]. This option, if adopted, would inevitably lead, for coherence, to merging several small but well-recognized genera, including Afrocirsium and Afrocarduus.
In addition, to justify a widely circumscribed Cirsium, we would be forced to adopt several infrageneric taxa, presumably at different ranks, the meaning of which would be difficult to explain in a modern phylogenetic taxonomy. Moreover, any rejection of taxonomic novelties in favour of nomenclatural stability cannot disregard the idea that genera must be monophyletic. Indeed, this casts doubt on whether, for stability and simplicity purposes, the best choice would be merging again all the above-cited genera in the Linnaean Carduus (cf. “solution one” in [2]).
Author Contributions
Conceptualization and original draft preparation, E.D.G. and P.B.; figure preparation, F.Z.; data curation F.Z: and P.B.; writing, review and editing, E.D.G., F.Z., P.C., D.I., and P.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
This paper is a rebuttal to a contribution including critical remarks against a previous work by us. In this respect, we have conflicting opinions with the scientists who authored that contribution and a legitimate interest in defending our scientific choices. Under a legal profile, however, we declare no conflict of interest.
References
- Moreyra, L.D.; Garcia-Jacas, N.; Roquet, C.; Ackerfield, J.R.; Arabacı, T.; Blanco-Gavaldà, C.; Brochmann, C.; Calleja, J.A.; Dirmenci, T.; Fujikawa, K.; et al. African mountain thistles: Three new genera in the Carduus-Cirsium Group. Plants 2023, 12, 3083. [Google Scholar] [CrossRef] [PubMed]
- Ackerfield, J.; Susanna, A.; Funk, V.; Kelch, D.; Park, D.S.; Thornhill, A.H.; Yildiz, B.; Arabaci, T.; Dirmenci, T. A prickly puzzle: Generic delimitations in the Carduus-Cirsium group (Compositae: Cardueae: Carduinae). Taxon 2020, 69, 715–738. [Google Scholar] [CrossRef]
- Del Guacchio, E.; Bureš, P.; Iamonico, D.; Carucci, F.; De Luca, D.; Zedek, F.; Caputo, P. Towards a monophyletic classification of Cardueae: Restoration of the genus Lophiolepis (= Cirsium pp) and new circumscription of Epitrachys. Plant. Biosyst. 2022, 56, 1269–1290. [Google Scholar] [CrossRef]
- Bureš, P.; Özcan, M.; Šmerda, J.; Michálková, E.; Horová, L.; Plačková, K.; Šmarda, P.; Elliott, T.L.; Veselý, P.; Ćato, S.; et al. Evolution of genome size and GC content in the tribe Carduinae (Asteraceae): Rare descending dysploidy and polyploidy, limited environmental control and strong phylogenetic signal. Preslia 2023, 95, 185–213. [Google Scholar] [CrossRef]
- Bureš, P.; Del Guacchio, E.; Šmerda, J.; Özcan, M.; Blizňáková, P.; Vavrinec, M.; Michálková, E.; Veselý, P.; Veselá, K.; Zedek, F. Intergeneric hybrid origin of the serious invasive tetraploid Cirsium Vulgare. Plant Biol. 2024, 26, 749–763. [Google Scholar] [CrossRef] [PubMed]
- Del Guacchio, E. (Department of Biology, University of Naples Federico II, Co. Botanical Garden, Naples, Italy); Zedek, F. (Department of Botany and Zoology, Faculty of Science, Masaryk University, Brno, Czech Republic); Caputo, P. (Department of Biology, University of Naples Federico II, Co. Botanical Garden, Naples, Italy); Bureš, P. (Department of Botany and Zoology, Faculty of Science, Masaryk University, Brno, Czech Republic). manuscript in preparation, 2024.
- Cassini, H. Lophiolèpe, Lophiolepis. In Dictionnaire des Sciences Naturelles; Cuvier, F., Ed.; F. G. Levrault & Le Normant: Paris & Strasbourg: Paris, France, 1823; Volume 27, pp. 180–192. [Google Scholar]
- Koch, K. Beitrage zu einer Flora des Orientes. Linnaea 1851, 24, 305–475. [Google Scholar]
- Fourreau, J. Catalogue des Plantes du cours du Rhône. Ann. Soc. Linn. Lyon Sér. 2 1869, 17, 89–200. [Google Scholar]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace Aware Data Structure for Phylogenomic Inference from Supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Vukotinović, L.F. Nove biline i razjašnjenja o nekih dvojbenih. Rad. Jugoslav. Akad. Znan. 1877, 39, 193–216. [Google Scholar]
- Guétrot, M. Plantes Hybrides de France; Imprimerie O. Marquant: Lille, France, 1927; Volume 1–2, pp. 29–30. [Google Scholar]
- Fournier, P.-V. Flore Complétive De la Plaine Française; Paul Lechavalier Éditeur: Paris, France, 1928; p. 277. [Google Scholar]
- Fournier, P.-V. Les Quatre Flores de la France, 1st ed.; Paul Lechavalier Éditeur: Paris, France, 1940; pp. 1001–1004. [Google Scholar]
- Sennen, F. Session de la Société botanique de France en 1931. Monde. Pl. 1931, 32, 18–19. [Google Scholar]
- Kharadze, A. Cirsium Mill. emend. Scop. In Flora of the USSR; Bobrov, E.G., Czerepanov, S.K., Eds.; Editio Academiae Scientiarum URSS: Moskow, Russia; Saint Petersburg, Russia, 1963; Volume 28, pp. 51–215. [Google Scholar]
- Talavera, S.; Valdés, B. Revisión del género Cirsium (Compositae) en la Península Ibérica. Lagascalia 1976, 5, 127–223. [Google Scholar]
- Davis, P.H.; Parris, S.B. Cirsium Mill. In Flora of Turkey and the East Aegean Islands; Davis, P.H., Ed.; Edinburgh University Press: Edinburgh, UK, 1975; Volume 5, pp. 370–412. [Google Scholar]
- Werner, K. Cirsium Miller. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1976; Volume 4, pp. 232–242. [Google Scholar]
- Petrak, F. Cirsium Mill. In Flora Iranica; Rechinger, K.H., Ed.; Akademische Druck- und Verlagsanstalt: Graz, Austria, 1979; Volume 139a, pp. 231–280. [Google Scholar]
- Shih, C. Notulae de Plantis Tribus Cynarearum Familiae Compositarum Sinicae (II). Acta. Phytotax. Sin. 1984, 22, 386–396. [Google Scholar]
- Dirmenci, T.; Özcan, T.; Arabacı, T.; Çelenk, S.; İsmailova, G.; Yıldız, B. Hybridization among three Cirsium (Asteraceae) species and important evidence for three new hybrids from Turkey. Turk. J. Bot. 2019, 43, 366–385. [Google Scholar] [CrossRef]
- Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Available online: http://www.plantsoftheworldonline.org/ (accessed on 12 April 2024).
- Flann, C.; Greuter, W.; Hind, D.J.N. Cassini’s Compositae genera: A nomenclatural and taxonomic assessment. Taxon 2010, 59, 1206–1244. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).