

Anti-DENV-2 Activity of Ethanolic Extracts from Arachis hypogaea L.: Peanut Skin as a Relevant Resource of Bioactive Compounds against Dengue Virus

 , ,
, ,
Abstract

1. Introduction
2. Results and Discussion
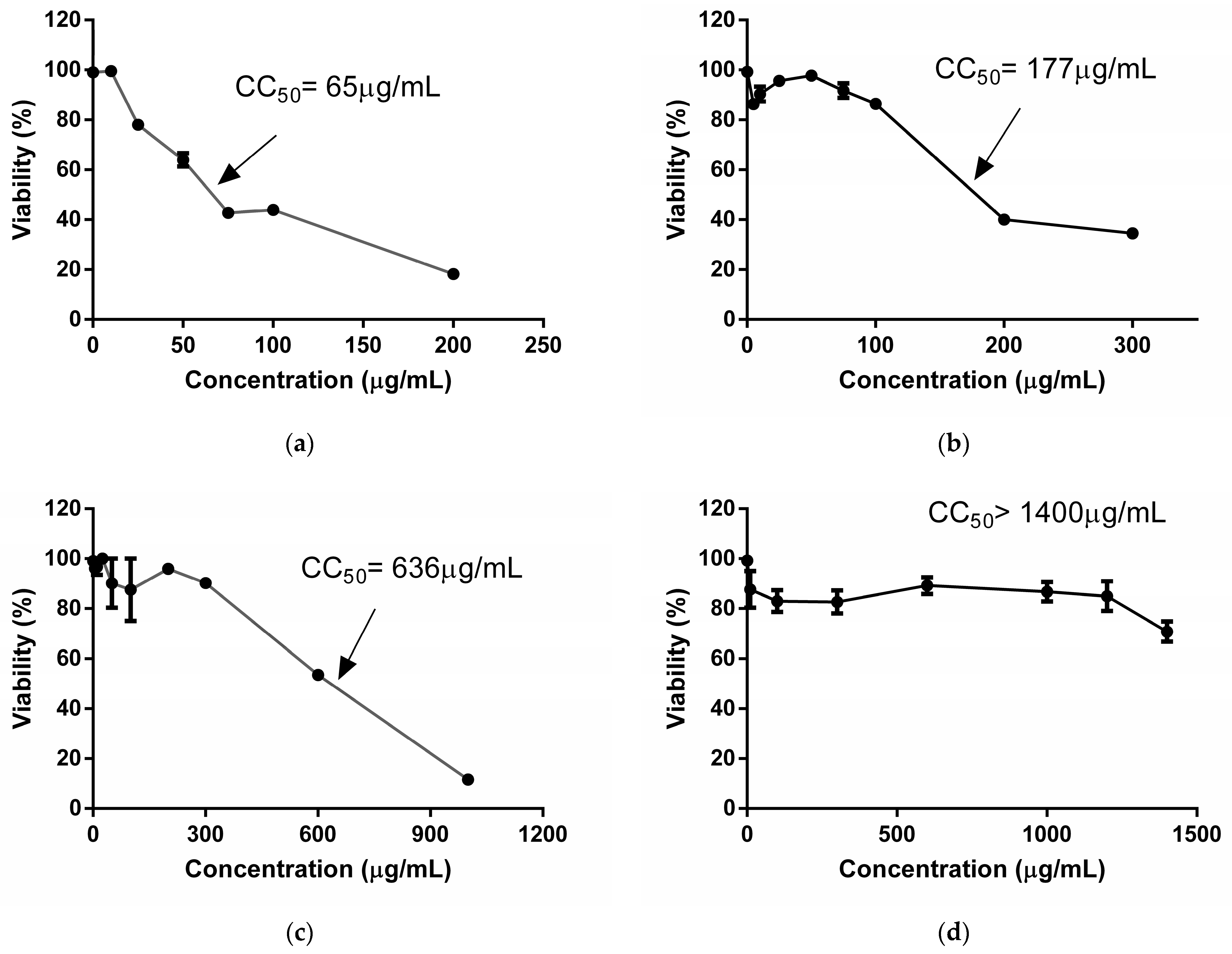
2.1. Cytotoxicity
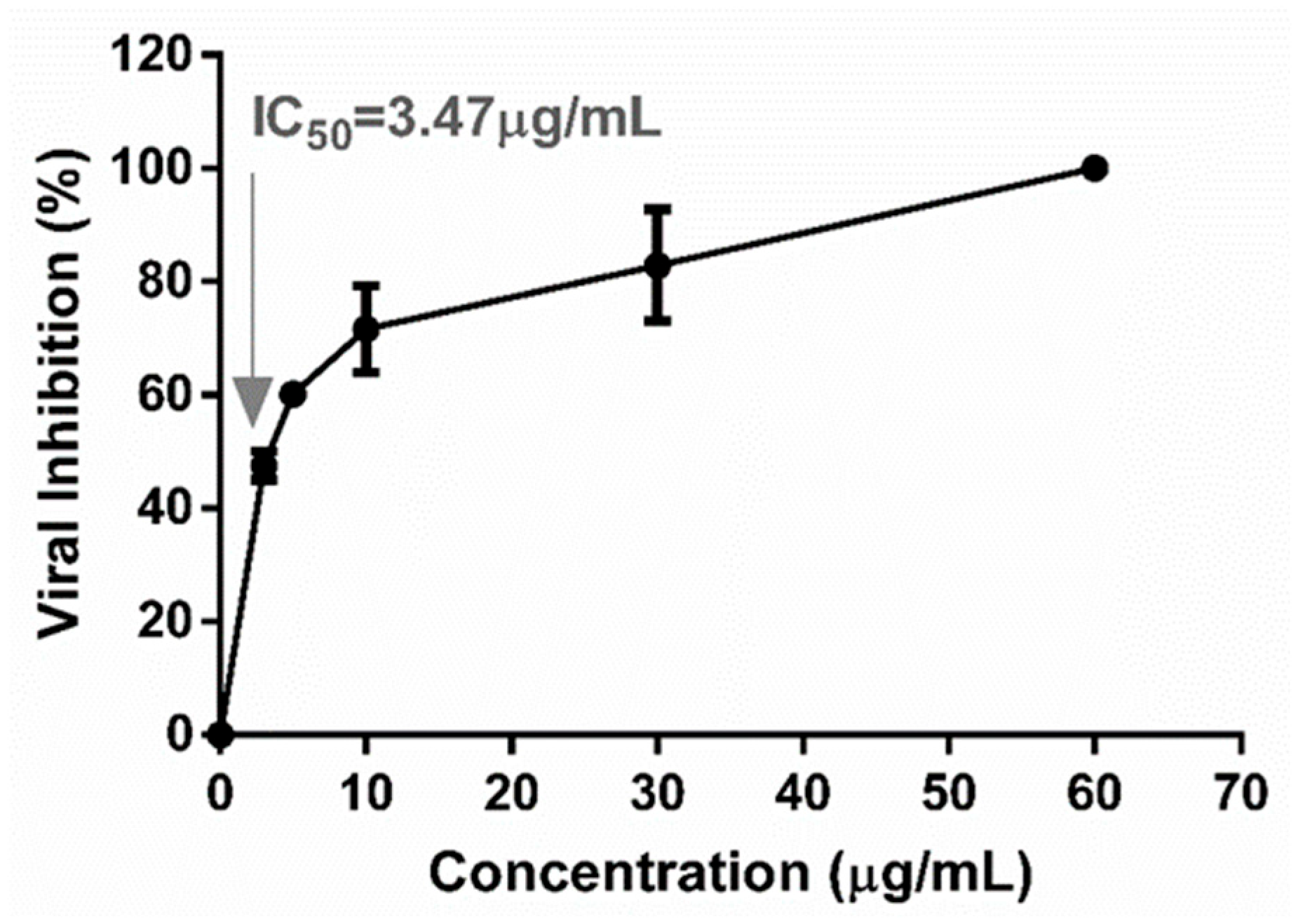
2.2. Antiviral Activity
2.3. Antiviral Activity throughout the Viral Replication Cycle
2.4. Quantitative Real-Time PCR Assay (qRT-PCR)
3. Materials and Methods
3.1. Ethanolic Extracts of Peanut
3.2. Cells and Virus
3.3. Cytotoxicity Assay In Vitro
3.3.1. Neutral Red Uptake (NRU)-Based Assay
3.3.2. 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium Bromide Metabolism (MTT)-Based Assay
3.4. Activity Antiviral of Ethanolic Extracts
3.4.1. Treatment in the Viral Adsorption and Penetration Stage
3.4.2. Treatment in Viral Post-Adsorption and Penetration Stage
3.4.3. Viral Pre-Treatment
3.4.4. Cell Pre-Treatment
3.5. Quantitative Real-Time PCR Assay (qRT-PCR)
3.6. Determination of 50% Inhibitory Concentration (IC50)
3.7. Selectivity Index
3.8. Data Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mustafa, M.S.; Rasotgi, V.; Jain, S.; Gupta, V. Discovery of fifth serotype of dengue virus (DENV-5): A new public health dilemma in dengue control. Med. J. Armed Forces India 2015, 71, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Velandia, M.; Castellanos, J. Virus del dengue: Estructura y ciclo viral. Infectio 2011, 15, 33–43. [Google Scholar] [CrossRef]
- Talarico, L. La entrada del virus dengue a la célula como un potencial blanco antiviral: Estudio de polisacáridos sulfatados como antivirales. Quím. Viva 2008, 7, 113–133. [Google Scholar]
- Teixeira, R.R.; Pereira, W.L.; Oliveira, A.F.; da Silva, A.M.; de Oliveira, A.S.; da Silva, M.L.; da Silva, C.C.; de Paula, S.O. Natural Products as Source of Potential Dengue Antivirals. Molecules 2014, 19, 8151–8176. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Castañeda, J.; Barreto dos Santos, F.; Martínez-Vega, R.; Galvão de Araujo, J.M.; Joint, G.; Sarti, E. Dengue in Latin America: Systematic Review of Molecular Epidemiological Trends. PLoS Negl. Trop. Dis. 2017, 11, e0005224. [Google Scholar] [CrossRef]
- Chew, M.F.; Tham, H.W.; Rajik, M.; Sharifah, S.H. Anti-dengue virus serotype 2 activity and mode of action of a novel peptide. J. Appl. Microbiol. 2015, 119, 1170–1180. [Google Scholar] [CrossRef]
- Kwon, H.J.; Kim, H.H.; Yoon, S.Y.; Ryu, Y.B.; Chang, J.S.; Cho, K.O.; Rho, M.C.; Park, S.J.; Lee, W.S. In vitro inhibitory activity of Alpinia katsumadai extracts against influenza virus infection and hemagglutination. Virol. J. 2010, 7, 307. [Google Scholar] [CrossRef]
- Zandi, K.; Taherzadeh, M.; Yaghoubi, R.; Tajbakhsh, S.; Rastian, Z.; Sartavi, K. Antiviral activity of Avicennia marina against herpes simplex virus type 1 and vaccine strain of poliovirus (An in vitro study). J. Med. Plants Res. 2009, 3, 771–775. [Google Scholar]
- Ocazionez, R.E.; Meneses, R.; Torres, F.A.; Stashenko, E. Virucidal activity of Colombian Lippia essential oils on dengue virus replication in vitro. Mem. Do Inst. Oswaldo Cruz 2010, 105, 3. [Google Scholar] [CrossRef]
- Zandi, K.; Lim, T.; Rahim, N.; Shu, M.-H.; Teoh, B.-T.; Sam, S.-S.; Danlami, M.-B.; Tan, K.-K.; Abubakar, S. Extract of Scutellaria baicalensis inhibits dengue virus replication. BMC Complement. Altern. Med. 2013, 13, 91. [Google Scholar] [CrossRef]
- Reinoso, E.; Oliva, M.M.; Pavicich, G.; Beoletto, V.; Carezzano, M.E.; Marioli, J.M.; Paletti Rovey, M.F.; Pimentel Betancourt, D.; Sabini, M.C.; Moliva, M.; et al. Una Farmacia en el Monte, 1st ed.; Ministerio de Ciencia y Tecnología de Córdoba: Córdoba, Argentina, 2019; ISBN 978-987-47203-0-6.
- Hishiki, T.; Kato, F.; Tajima, S.; Toume, K.; Umezaki, M.; Takasaki, T.; Miura, T. Hirsutine, an Indole Alkaloid of Uncaria rhynchophylla, Inhibits Late Step in Dengue Virus Lifecycle. Front. Microbiol. 2017, 8, 1674. [Google Scholar] [CrossRef] [PubMed]
- Ramalingam, S.; Karupannan, S.; Padmanaban, P.; Vijayan, S.; Sheriff, K.; Palani, G.; Krishnasamy, K. Anti-dengue activity of Andrographis paniculata extracts and quantification of dengue viral inhibition by SYBR green reverse transcription polymerase chain reaction. AYU J. 2018, 39, 87–91. [Google Scholar] [CrossRef]
- Jiang, S.; Ma, Y.; Yan, D. Antioxidant and antimicrobial properties of water soluble polysaccharide from Arachis hypogaea seeds. J. Food Sci. Technol. 2014, 51, 2839–2844. [Google Scholar] [CrossRef] [PubMed]
- AÑO XXXIX—N° Edición 2017. Available online: https://www.bcr.com.ar/es/mercados/investigacion-y-desarrollo/informativo-semanal/noticias-informativo-semanal/un-nuevo (accessed on 3 September 2023).
- Rossi, Y.E.; Bohl, L.P.; Vanden Braber, N.L.; Ballatore, M.B.; Escobar, F.M.; Bodoira, R.; Maestri, D.M.; Porporatto, C.; Cavaglieri, L.R.; Montenegro, M.A. Polyphenols of peanut (Arachis hypogaea L.) skin as bioprotectors of normal cells. Studies of cytotoxicity, cytoprotection and interaction with ROS. J. Funct. Foods 2020, 67, 103862. [Google Scholar] [CrossRef]
- Larrauri, M.; Zunino, M.P.; Zygadlo, J.A.; Grosso, N.R.; Nepote, V. Chemical characterization and antioxidant properties of fractions separated from extract of peanut skin derived from different industrial processes. Ind. Crops Prod. 2016, 95, 964. [Google Scholar] [CrossRef]
- Makau, J.N.; Watanabe, K.; Mohammed, M.M.D.; Nishida, N. Antiviral Activity of Peanut (Arachis hypogaea L.) Skin Extract Against Human Influenza Viruses. J. Med. Food 2018, 21, 777–784. [Google Scholar] [CrossRef]
- Toomer, O.T. A comprehensive review of the value-added uses of peanut (Arachis hypogaea) skins and by-products. Crit. Rev. Food Sci. Nutr. 2018, 60, 341–350. [Google Scholar] [CrossRef]
- Menis Candela, F.; Giordano, W.F.; Quiroga, P.L.; Escobar, F.M.; Mañas, F.; Roma, D.A.; Larrauri, M.; Comini, L.R.; Soria, E.A.; Sabini, M.C. Evaluation of cellular safety and the chemical composition of the peanut (Arachis hypogaea L.) ethanolic extracts. Heliyon 2020, 6, e05119. [Google Scholar] [CrossRef]
- Cossetin, J.F.; da Silva Brum, E.; Casoti, R.; Camponogara, C.; Castro, R.; Maziero, D.M.; de David, C.T.; Gaube, A.C.; Ramos, A.P.; Pintos, F.G.; et al. Peanut leaf extract has antioxidant and anti-inflammatory activity but no acute toxic effects. Regul. Toxicol. Pharmacol. 2019, 107, 104407. [Google Scholar] [CrossRef]
- Cordeiro-Massironi, K.; Soares-Freitas, R.A.M.; Sampaio, G.R.; Pinaffi-Langley, A.C.d.C.; Bridi, R.; de Camargo, A.C.; Torres, E.A.F.S. In Vitro Digestion of Peanut Skin Releases Bioactive Compounds and Increases Cancer Cell Toxicity. Antioxidants 2023, 12, 1356. [Google Scholar] [CrossRef]
- Tsai, C.-H.; Huang, H.-C.; Lin, K.-J.; Liu, J.-M.; Chen, G.-L.; Yeh, Y.-H.; Lu, T.-L.; Lin, H.-W.; Lu, M.-T.; Chu, P.-C. Inhibition of Autophagy Aggravates Arachis hypogaea L. Skin Extracts-Induced Apoptosis in Cancer Cells. Int. J. Mol. Sci. 2024, 25, 1345. [Google Scholar] [CrossRef] [PubMed]
- Angelina, M.; Hanafi, M.; Suyatna, F.; Mirawati, S.; Ratnasari, S.; Dewi, B. Antiviral Effect of Sub Fraction Cassia alata Leaves Extract to Dengue Virus Serotype-2 strain New Guinea C in Human Cell Line Huh-7 it-1. IOP Conf. Ser. Earth Environ. Sci. 2017, 101, 012004. [Google Scholar] [CrossRef]
- Clain, E.; Haddad, J.G.; Koishi, A.C.; Sinigaglia, L.; Rachidi, W.; Desprès, P.; Duarte dos Santos, C.N.; Guiraud, P.; Jouvenet, N.; Kalamouni, C. The Polyphenol-Rich Extract from Psiloxylon mauritianum, an Endemic Medicinal Plant from Reunion Island, Inhibits the Early Stages of Dengue and Zika Virus Infection. Int. J. Mol. Sci. 2019, 20, 1860. [Google Scholar] [CrossRef] [PubMed]
- Bodoira, R.; Cittadini, M.C.; Velez, A.; Rossi, Y.; Montenegro, M.; Martínez, M.; Maestri, D. An overview on extraction, composition, bioactivity and food applications of peanut phenolics. Food Chem. 2022, 381, 132250. [Google Scholar] [CrossRef]
- Terlizzi, M.E.; Occhipinti, A.; Luganini, A.; Maffei, M.E.; Gribaudo, G. Inhibition of herpes simplex type 1 and type 2 infections by Oximacro, a cranberry extract with a high content of A-type proanthocyanidins (PACs-A). Antivir. Res. 2016, 132, 154–164. [Google Scholar] [CrossRef]
- Joshi, S.S.; Howell, A.B.; D’Souza, D.H. Reduction of Enteric Viruses by Blueberry Juice and Blueberry Proanthocyanidins. Food Environ. Virol. 2016, 8, 235–243. [Google Scholar] [CrossRef]
- Sureram, S.; Chutiwitoonchai, N.; Pooprasert, T.; Sangsopha, W.; Limjiasahapong, S.; Jariyasopit, N.; Sirivatanauksorn, Y.; Khoomrung, S.; Mahidol, C.; Ruchirawat, S.; et al. Discovery of procyanidin condensed tannins of (-)-epicatechin from Kratom, Mitragyna speciosa, as virucidal agents against SARS-CoV-2. Int. J. Biol. Macromol. 2024, 273 Pt 1, 133059. [Google Scholar] [CrossRef]
- Sorita, G.D.; Leimann, F.V.; Salvador Ferreira, S.R. Phenolic Fraction from Peanut (Arachis hypogaea L.) By product: Innovative Extraction Techniques and New Encapsulation Trends for Its Valorization. Food Bioprocess Technol. 2023, 16, 726–748. [Google Scholar] [CrossRef]
- Alasalvar, C.; Bolling, B.W. Review of nut phytochemicals, fat-soluble bioactives, antioxidant components and health effects. Br. J. Nutr. 2015, 113, 68–78. [Google Scholar] [CrossRef]
- Flores-Ocelotl, M.R.; Rosas-Murrieta, N.H.; Moreno, D.A.; Vallejo-Ruiz, V.; Reyes-Leyva, J.; Domínguez, F.; Santos-López, G. Taraxacum officinale and Urtica dioica extracts inhibit dengue virus serotype 2 replication in vitro. BMC Complement. Altern. Med. 2018, 18, 95. [Google Scholar] [CrossRef]
- Rodríguez-Ortega, M.; Chumpitaz, Z.; Ríos, S.; Méndez, M.; Méndez, J.; Cabrera, G. Actividad antiviral contra el virus de la fiebre amarilla, cepa vacunal 17D, de extractos de hojas de Taraxacum officinale GH Weber ex Wiggers. Bol. Latinoam. Caribe Plantas Med. Aromát. 2013, 12, 346–355. Available online: http://saber.ucv.ve/jspui/handle/123456789/4837 (accessed on 2 November 2023).
- Wang, G.F.; Shi, L.; Ren, Y.; Liu, Q.; Liu, H.; Zhang, R.; Li, Z.; Zhu, F.; He, P.; Tang, W. Anti-hepatitis B virus activity of chlorogenic acid, quinic acid and caffeic acid in vivo and in vitro. Antivir. Res. 2009, 83, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Utsunomiya, H.; Ichinose, M.; Ikeda, K.; Uozaki, M.; Morishita, J.; Kuwahara, T.; Koyama, A.H.; Yamasaki, H. Inhibition by caffeic acid of the influenza a virus multiplication in vitro. Int. J. Mol. Med. 2014, 34, 1020–1024. [Google Scholar] [CrossRef]
- Ikeda, K.; Tsujimoto, K.; Uozaki, M.; Nishide, M.; Suzuki, Y.; Koyama, A.H.; Yamasaki, H. Inhibition of multiplication of herpes simplex virus by caffeic acid. Int. J. Mol. Med. 2011, 28, 595–598. [Google Scholar] [CrossRef]
- Mohammed, F.S.; Uysal, I.; Sevindik, M. A review on antiviral plants effective against different virus types prospectsin pharmaceutical sciences. Prospect. Pharm. Sci. 2023, 21, 1–21. Available online: https://prospects.wum.edu.pl (accessed on 7 October 2024).
- García, G.H.; Campos, R.; De Torres, R.; Broussalis, A.; Ferraro, G.; Martino, V.; Coussio, J. Antiherpetic Activity of some Argentine Medicinal Plants. Fitoterapia 1990, 41, 542–546. [Google Scholar]
- Dulbecco, R. Production of plaques in monolayer tissue culture by single particles of an animal virus. Proc. Natl. Acad. Sci. USA 1962, 38, 747–752. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid for cellular colorimetric assay grow and survival application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Sabini, M.C.; Escobar, F.; Tonn, C.; Zanon, S.; Contigiani, M.; Sabini, L. Evaluation of antiviral activity of aqueous extracts from Achyrocline satureioides against West. Equine Enceph. Virus Nat. Prod. Res. 2012, 26, 405–415. [Google Scholar] [CrossRef]
- Sabini, M.C.; Cariddi, L.N.; Escobar, F.; Mañas, F.; Comini, L.; Iglesias, D.; Larrauri, M.; Núñez Montoya, S.; Sereno, J.; Contigiani, M.; et al. Potent inhibition of Western Equine Encephalitis virus by a fraction rich in flavonoids and phenolic acids obtained from Achyrocline satureioides. Rev. Bras. Farmacogn. 2016, 26, 571–578. [Google Scholar] [CrossRef]
- Gurukumar, K.; Priyadarshini, D.; Patil, J.; Bhagat, A.; Singh, A.; Shah, P.S.; Cecilia, D. Development of real time PCR for detection and quantitation of Dengue Viruses. Virol. J. 2009, 6, 10. [Google Scholar] [CrossRef]
- Prada-Arismendy, J.; Castellanos, J. Real time PCR. Application in dengue studies. Colomb. Médica 2011, 42, 243–258. Available online: https://www.redalyc.org/pdf/283/28318450016.pdf (accessed on 10 November 2022). [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | Adsorption– Penetration (%) | Post-Adsorption and Penetration (%) | Viral Pre-Treatment (%) | Cell Pre-Treatment (%) |
|---|---|---|---|---|
| TEE (30 µg/mL) | 84.0 ± 1.4 | 99.9 ± 0.1 | 92.5 ± 10.6 | 95 ± 7 |
| SEE (300 µg/mL) | 13.7 ± 1.7 | 51.0 ± 8.6 | 33.2 ± 5.5 | 6.0 ± 2.8 |
| Method | Quotient CC50/IC50 | Selectivity Index |
|---|---|---|
| Neutral Red Uptake | 169/3.47 | 48.70 |
| MTT Reduction | 65/3.47 | 18.73 |
| Sample (µg/mL) | Ct | Viral cDNA Concentration (µg/µL) |
|---|---|---|
| VC | 19.44 | 2.36 |
| 10 | 27.79 | 1.42 |
| 20 | 34.83 | 0 |
| 30 | 35.65 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menis Candela, F.; Soria, E.A.; Moliva, M.V.; Suárez Perrone, A.; Reinoso, E.B.; Giordano, W.; Sabini, M.C. Anti-DENV-2 Activity of Ethanolic Extracts from Arachis hypogaea L.: Peanut Skin as a Relevant Resource of Bioactive Compounds against Dengue Virus. Plants 2024, 13, 2881. https://doi.org/10.3390/plants13202881
Menis Candela F, Soria EA, Moliva MV, Suárez Perrone A, Reinoso EB, Giordano W, Sabini MC. Anti-DENV-2 Activity of Ethanolic Extracts from Arachis hypogaea L.: Peanut Skin as a Relevant Resource of Bioactive Compounds against Dengue Virus. Plants. 2024; 13(20):2881. https://doi.org/10.3390/plants13202881
Chicago/Turabian StyleMenis Candela, Florencia, Elio Andrés Soria, Melina Vanesa Moliva, Agostina Suárez Perrone, Elina Beatríz Reinoso, Walter Giordano, and María Carola Sabini. 2024. "Anti-DENV-2 Activity of Ethanolic Extracts from Arachis hypogaea L.: Peanut Skin as a Relevant Resource of Bioactive Compounds against Dengue Virus" Plants 13, no. 20: 2881. https://doi.org/10.3390/plants13202881
APA StyleMenis Candela, F., Soria, E. A., Moliva, M. V., Suárez Perrone, A., Reinoso, E. B., Giordano, W., & Sabini, M. C. (2024). Anti-DENV-2 Activity of Ethanolic Extracts from Arachis hypogaea L.: Peanut Skin as a Relevant Resource of Bioactive Compounds against Dengue Virus. Plants, 13(20), 2881. https://doi.org/10.3390/plants13202881