
Genome-Wide Studies of FH Family Members in Soybean (Glycine max) and Their Responses under Abiotic Stresses


 ,
, 
Abstract
1. Background
2. Result
2.1. Identification and Characterization of FH Gene Family from Soybean
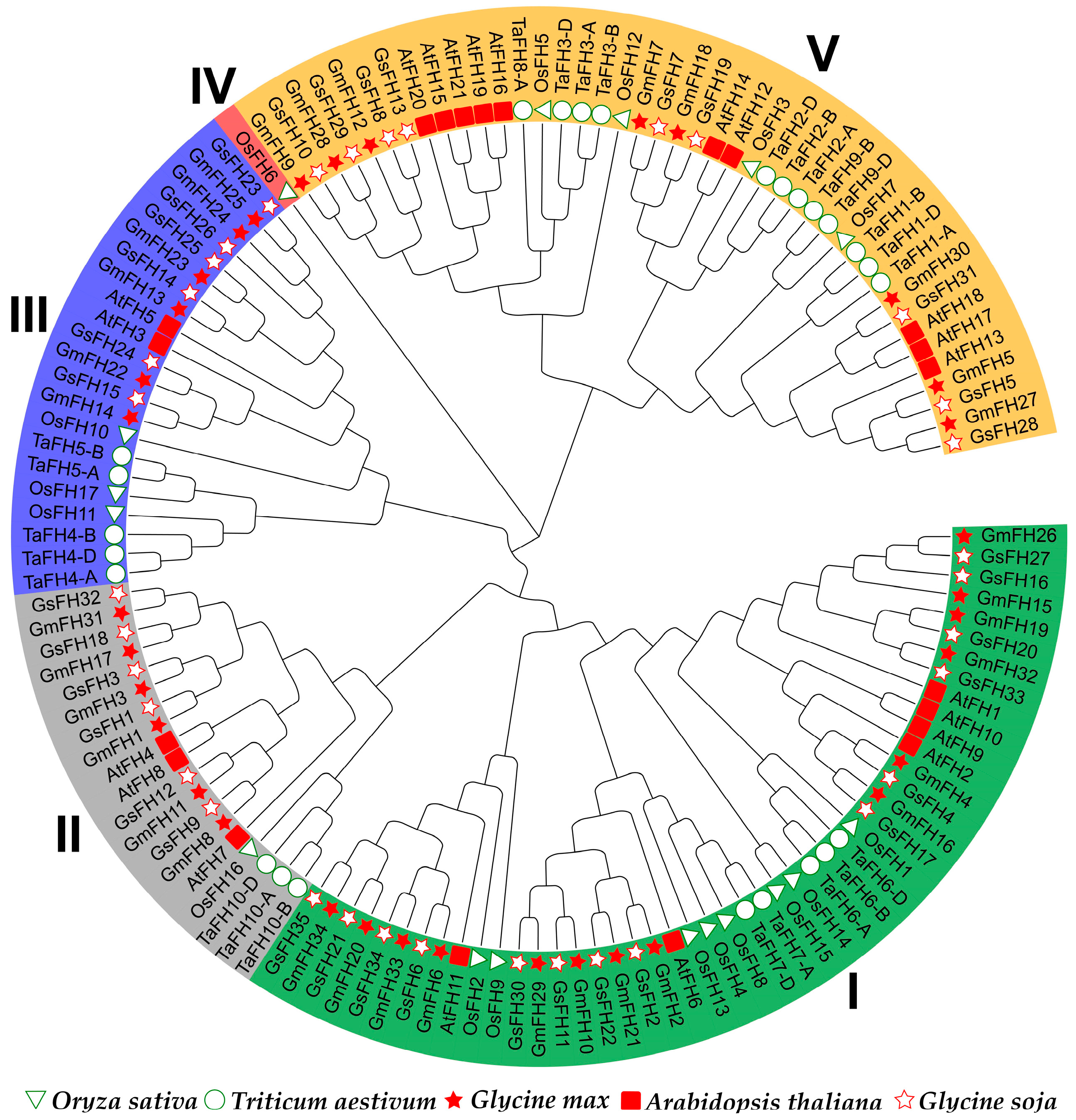
2.2. Evolutionary Comparison of the FH Gene Family from Diverse Plant Species
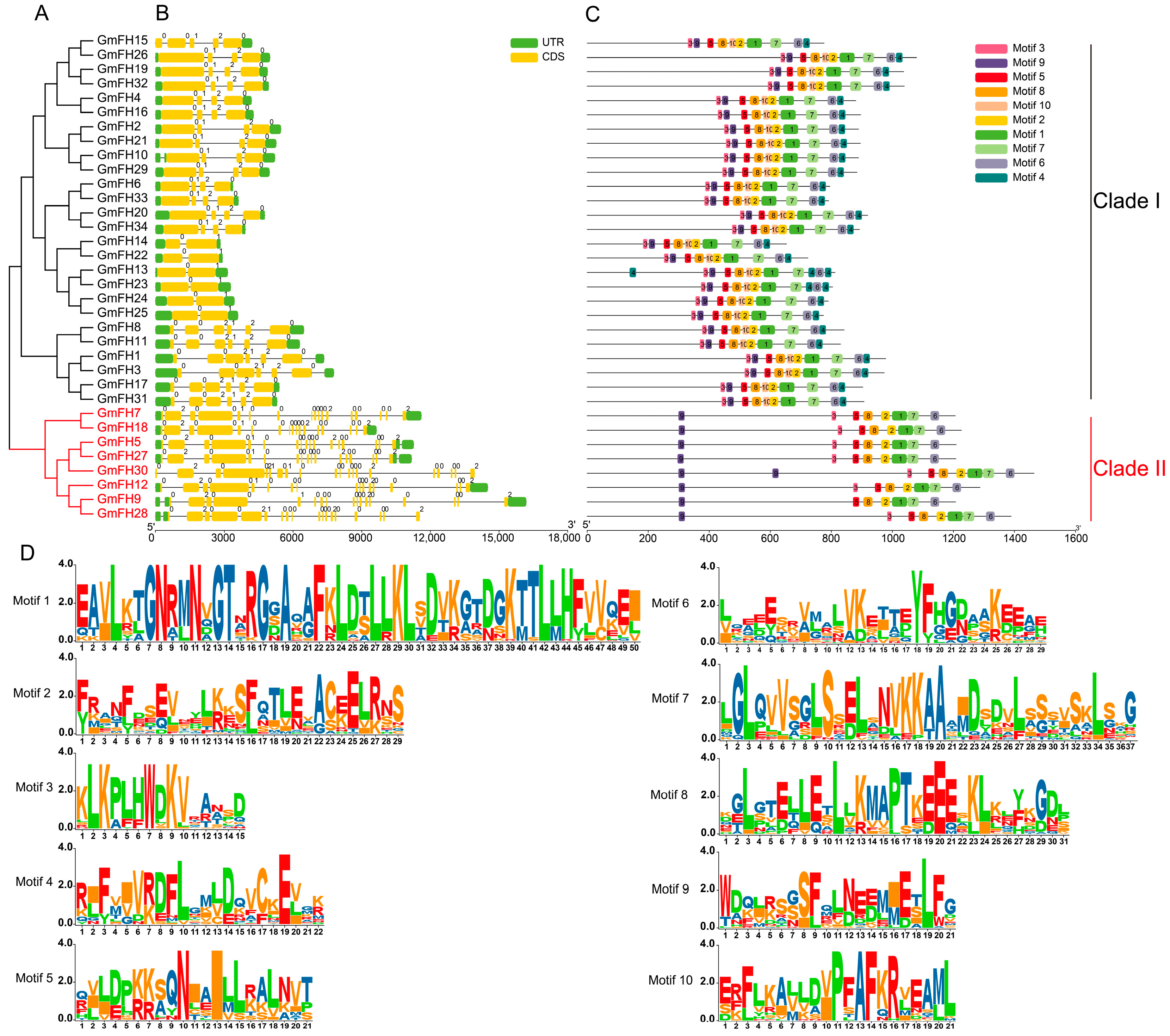
2.3. Analysis of Evolutionary Relationship, Gene Structure, and Conservative Motif of FH Family Members in Soybean
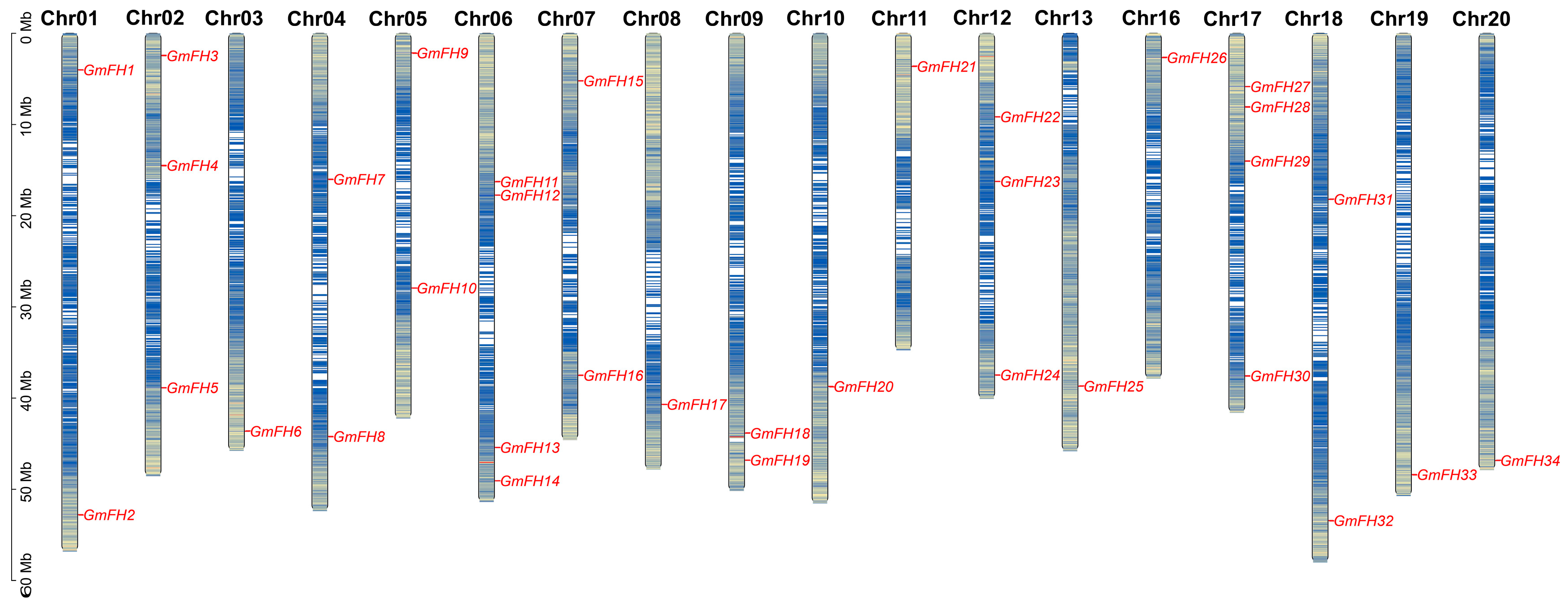
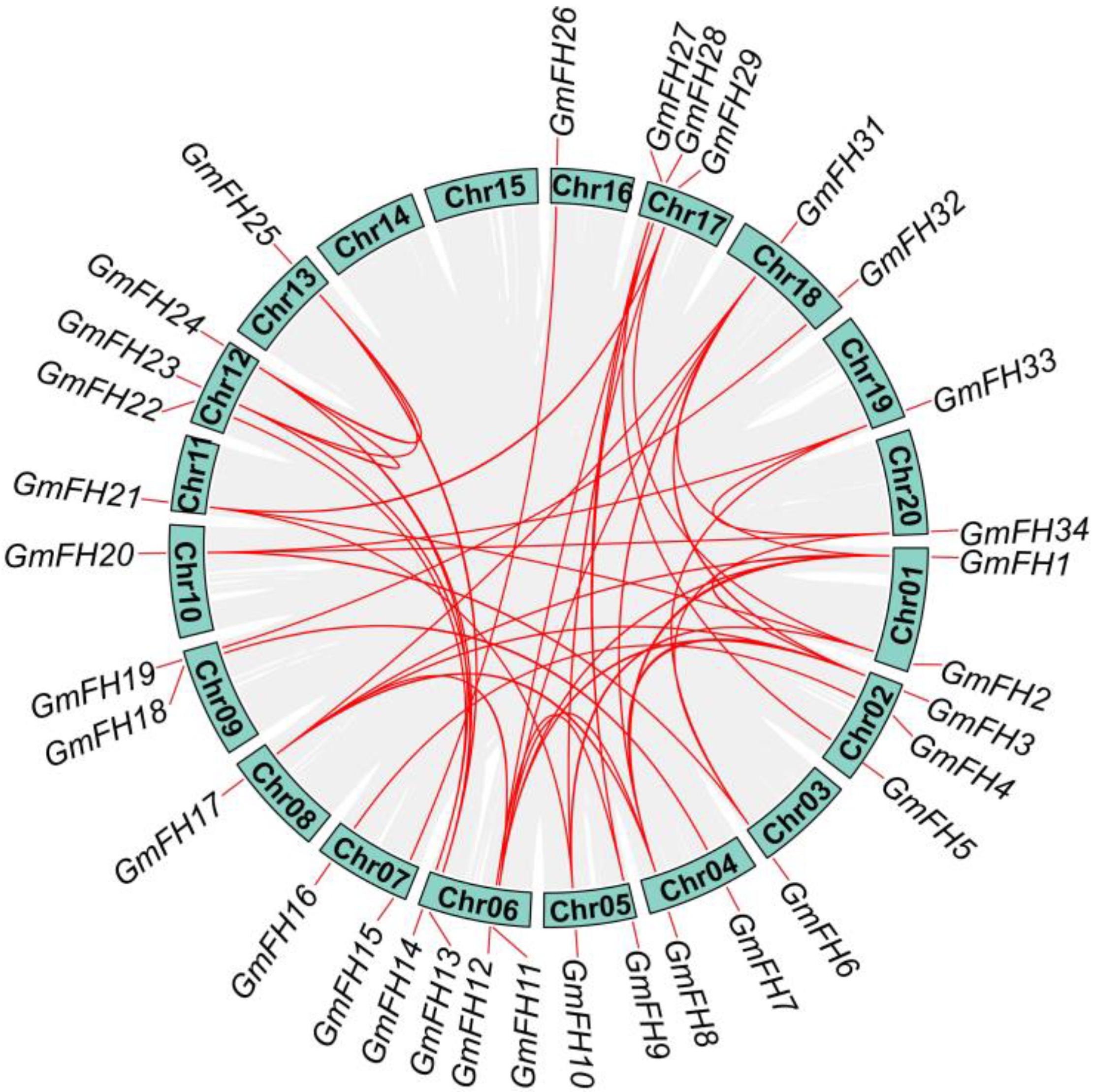
2.4. Chromosomal Distribution and Duplication Events
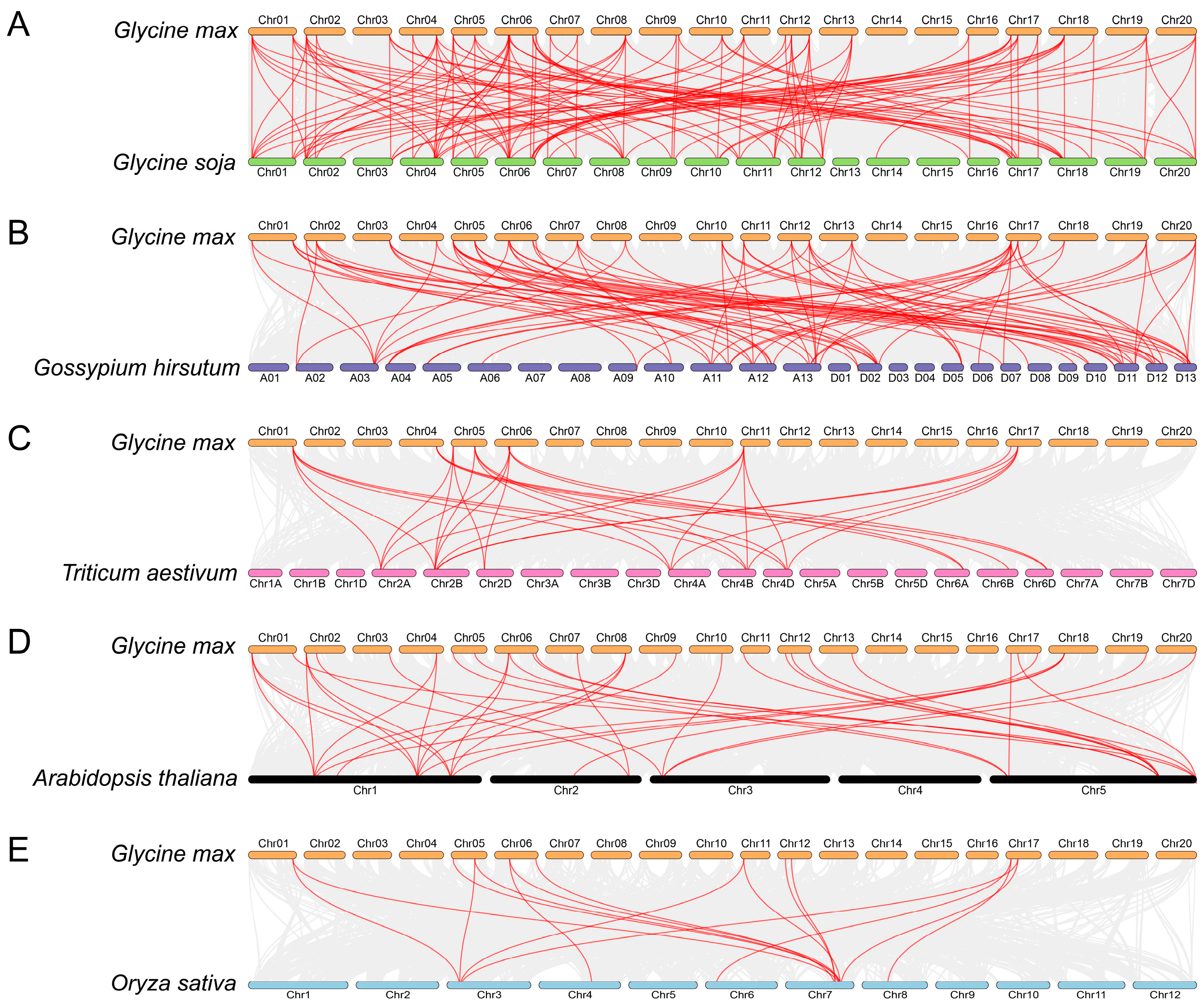
2.5. Investigation of Syntenic Relationships between Soybeans and Other Plant Species
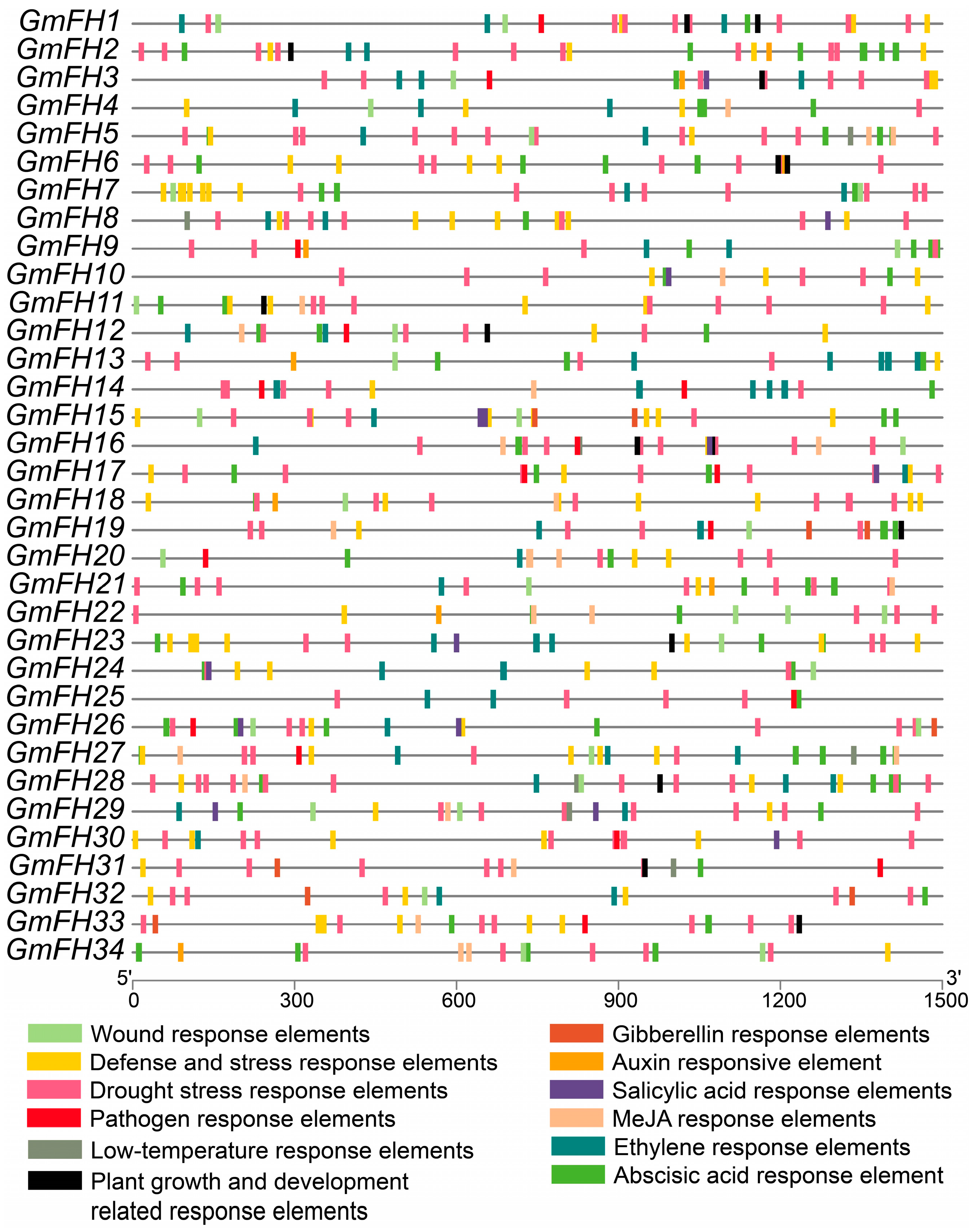
2.6. Analysis of Promoter Cis-Acting Elements in the FH Family Genes
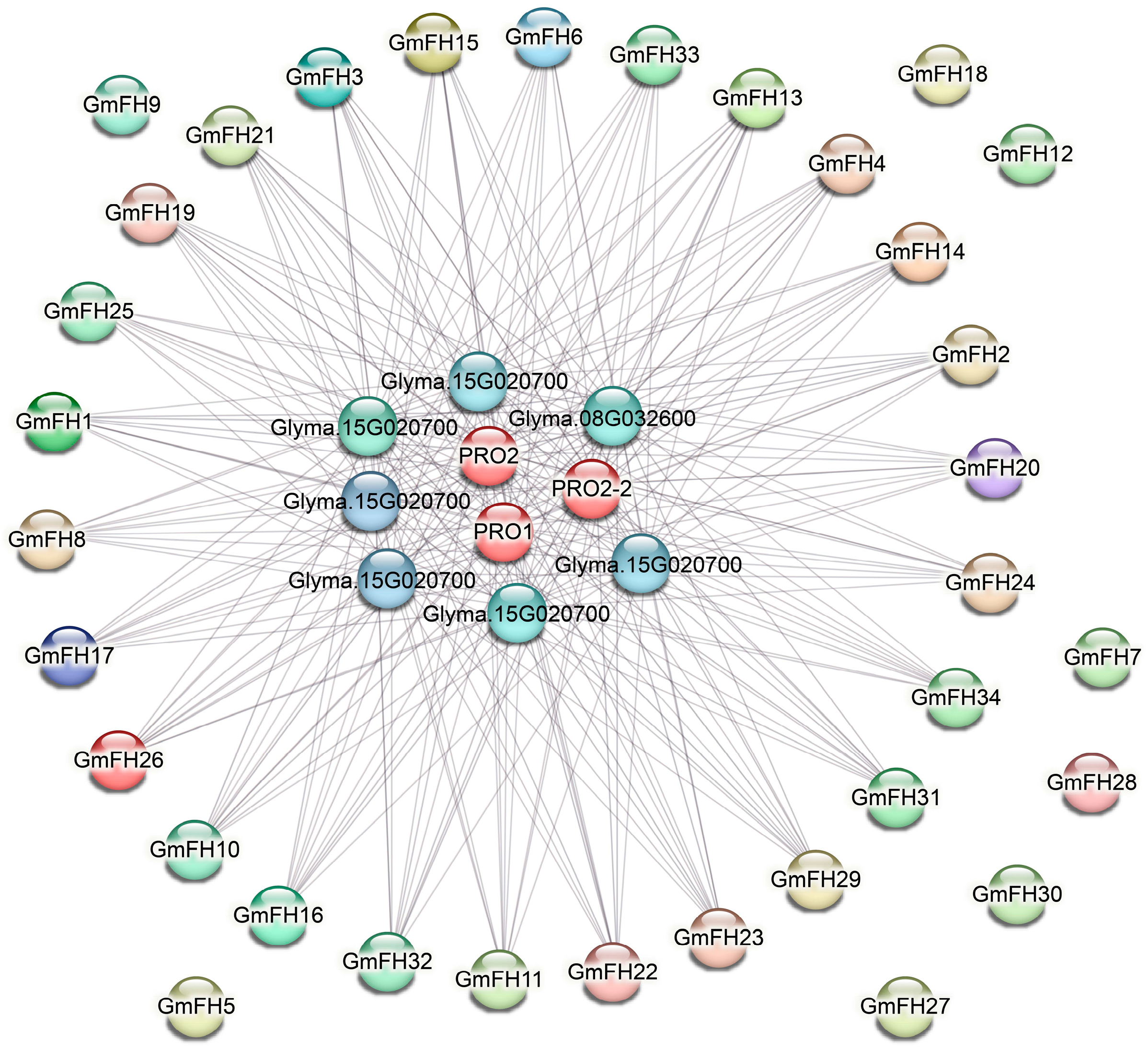
2.7. Investigation of Networks Depicting Protein-Protein Interactions
2.8. Expression Patterns of FH Genes in Soybean Specific to Different Tissues
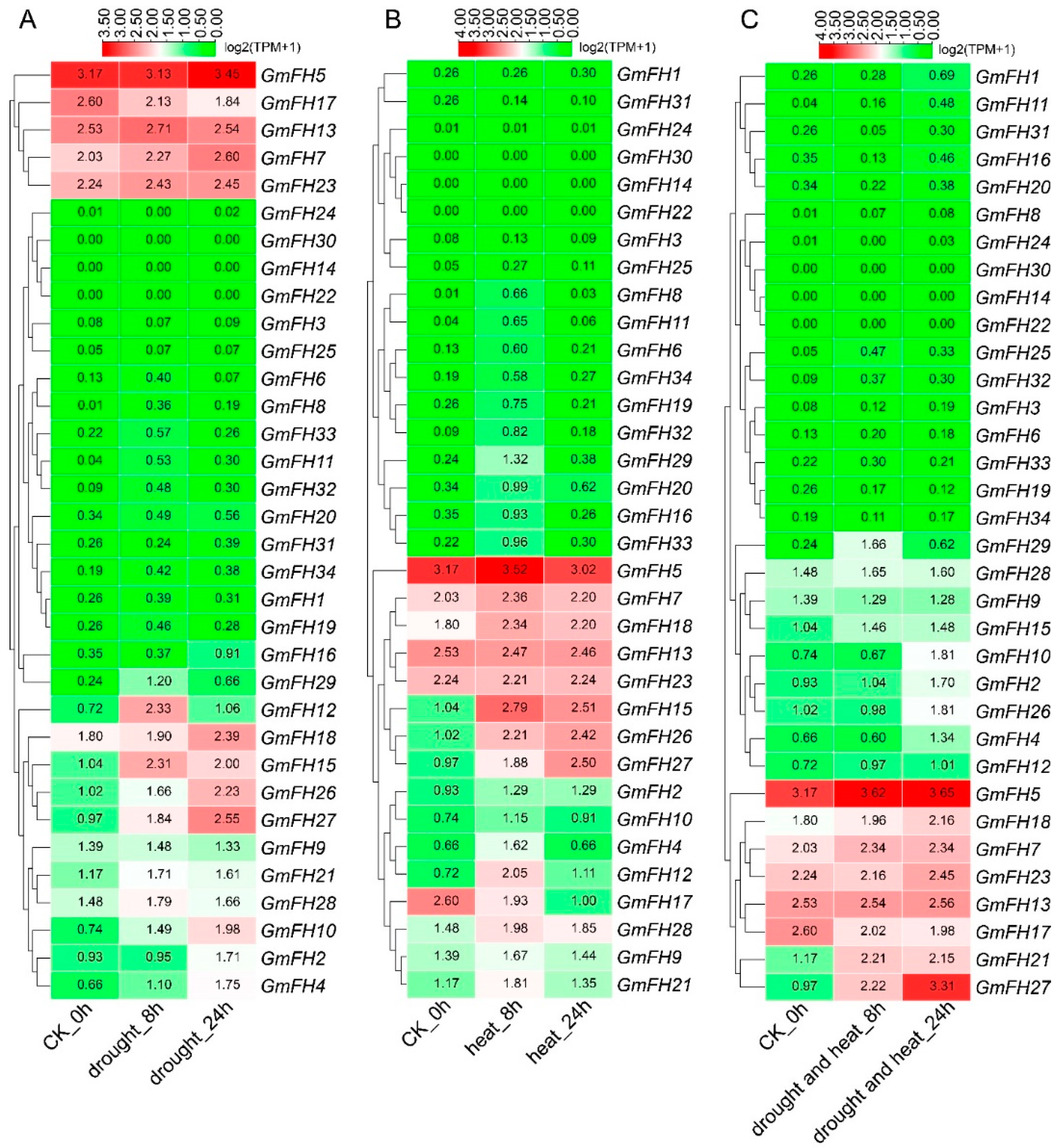
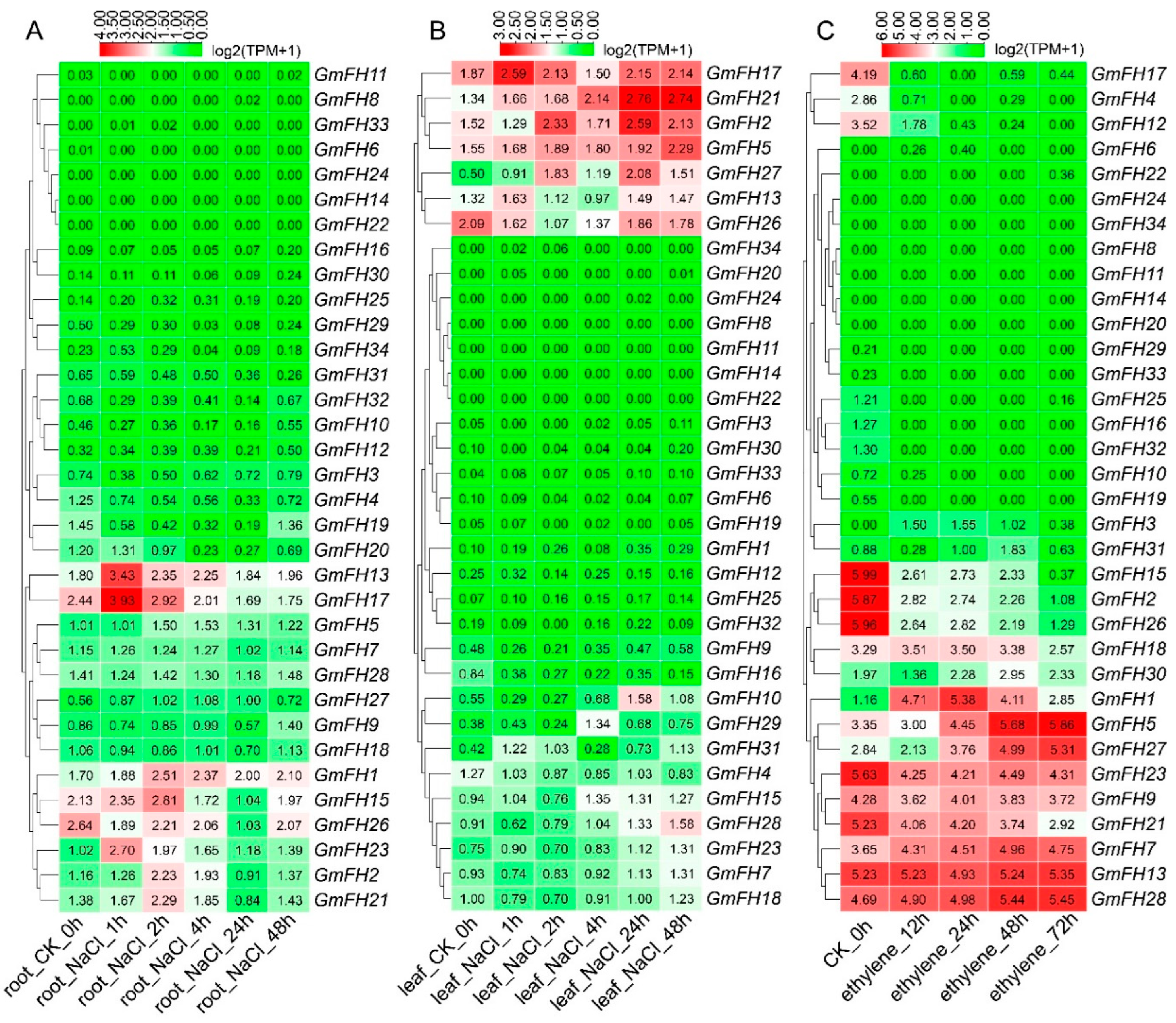
2.9. Expression Analysis of the FH Family Genes in Soybean in Response to Abiotic Stresses
2.10. Go Enrichment Analysis of GmFH Family Members
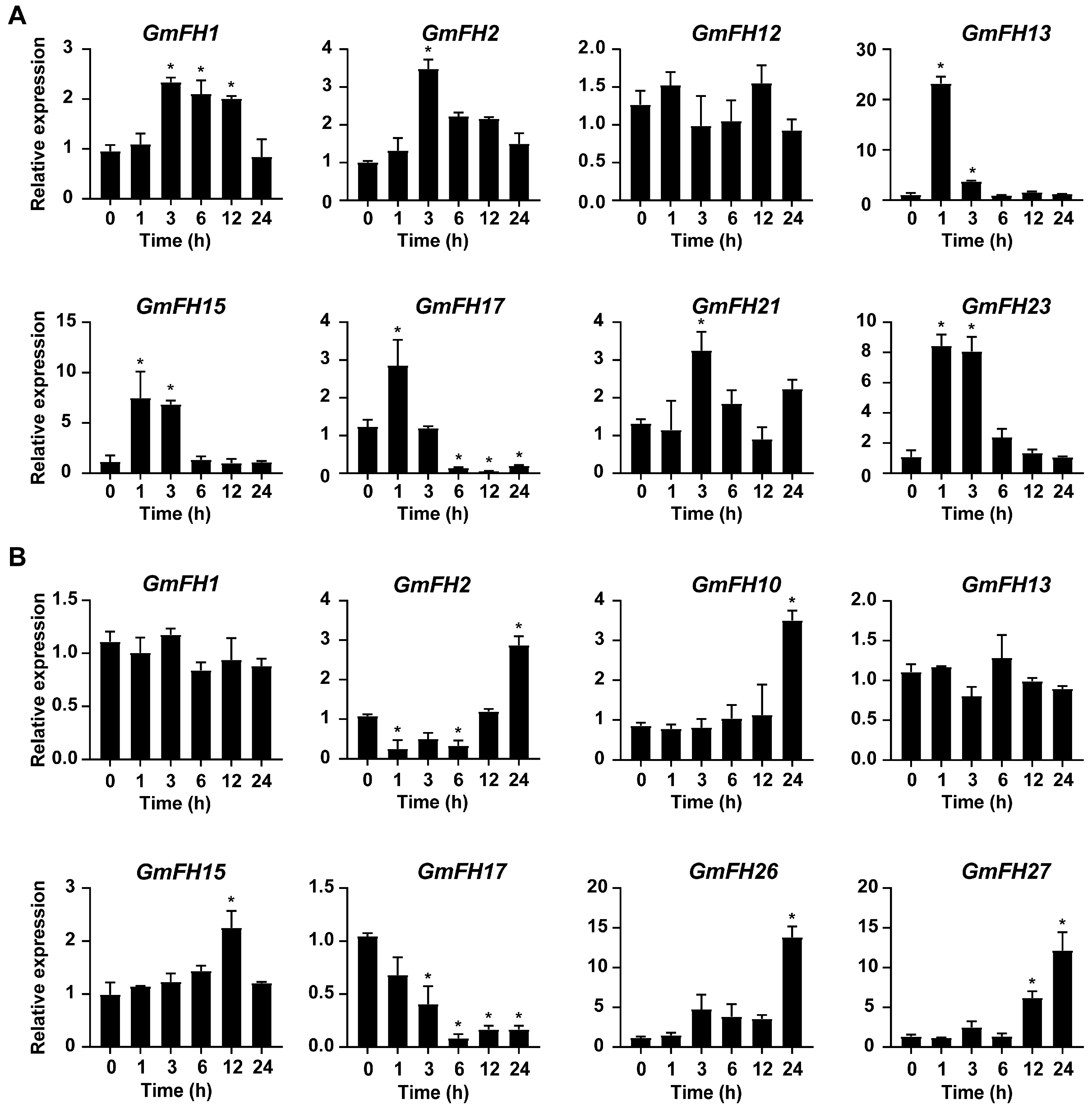
2.11. Verification of Expression Patterns of the GmFH Genes by qRT-PCR
3. Discussion
4. Methods and Materials
4.1. Plant Growth Conditions and Stress Treatment
4.2. Identification and Physicochemical Analysis of the GmFH Family Members
4.3. Evolutionary Analysis between Glycine Max and Other Plants
4.4. Evolutionary Relationships, Conserved Domain, and Gene Structure of GmFH Family Genes
4.5. Chromosomal Mapping and Gene Duplication
4.6. Promoter Elements, GO Enrichment Analysis, and Protein Interaction
4.7. qRT-PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, N.; Qi, X.; Li, M.; Xie, M.; Gao, Y.; Cheung, M.; Wong, F.; Lam, H. Improvement in nitrogen fixation capacity could be part of the domestication process in soybean. Heredity 2016, 117, 84–93. [Google Scholar] [CrossRef]
- De Borja Reis, A.F.; Moro Rosso, L.; Purcell, L.C.; Naeve, S.; Casteel, S.N.; Kovács, P.; Archontoulis, S.; Davidson, D.; Ciampitti, I.A. Environmental factors associated with nitrogen fixation prediction in soybean. Front. Plant Sci. 2021, 12, 27–39. [Google Scholar] [CrossRef] [PubMed]
- El Haddad, N.; En-Nahli, Y.; Choukri, H.; Aloui, K.; Mentag, R.; El-Baouchi, A.; Hejjaoui, K.; Rajendran, K.; Smouni, A.; Maalouf, F. Metabolic Mechanisms Underlying Heat and Drought Tolerance in Lentil Accessions: Implications for Stress Tolerance Breeding. Plants 2023, 12, 3962. [Google Scholar] [CrossRef]
- Tavakkoli, E.; Rengasamy, P.; McDonald, G.K. High concentrations of Na+ and Cl– ions in soil solution have simultaneous detrimental effects on growth of faba bean under salinity stress. J. Exp. Bot. 2010, 61, 4449–4459. [Google Scholar] [CrossRef]
- Isayenkov, S.V. Genetic sources for the development of salt tolerance in crops. Plant Growth Regul. 2019, 89, 1–17. [Google Scholar] [CrossRef]
- Tiwari, R.; Singh, A.K.; Rajam, M.V. GmPARPs differentially regulate the drought and heat stress tolerance in soybean. Plant Growth Regul. 2023, 101, 643–661. [Google Scholar] [CrossRef]
- Singh, A.K.; Raina, S.K.; Kumar, M.; Aher, L.; Ratnaparkhe, M.B.; Rane, J.; Kachroo, A. Modulation of GmFAD3 expression alters abiotic stress responses in soybean. Plant Mol. Biol. 2022, 110, 199–218. [Google Scholar] [CrossRef]
- Chen, L.; Yang, H.; Fang, Y.; Guo, W.; Chen, H.; Zhang, X.; Dai, W.; Chen, S.; Hao, Q.; Yuan, S. Overexpression of GmMYB14 improves high-density yield and drought tolerance of soybean through regulating plant architecture mediated by the brassinosteroid pathway. Plant Biotechnol. J. 2021, 19, 702–716. [Google Scholar] [CrossRef]
- Hao, Y.J.; Wei, W.; Song, Q.X.; Chen, H.W.; Zhang, Y.Q.; Wang, F.; Zou, H.F.; Lei, G.; Tian, A.G.; Zhang, W.K. Soybean NAC transcription factors promote abiotic stress tolerance and lateral root formation in transgenic plants. Plant J. 2011, 68, 302–313. [Google Scholar] [CrossRef]
- Wang, J.; Hossain, M.S.; Lyu, Z.; Schmutz, J.; Stacey, G.; Xu, D.; Joshi, T. SoyCSN: Soybean context-specific network analysis and prediction based on tissue-specific transcriptome data. Plant Direct 2019, 3, e00167. [Google Scholar] [CrossRef] [PubMed]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, 1178–1186. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xue, X.; Ren, H. New insights into the role of plant formins: Regulating the organization of the actin and microtubule cytoskeleton. Protoplasma 2012, 249, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Staiger, C.J. Signaling to the actin cytoskeleton in plants. Annu. Rev. Plant Biol. 2000, 51, 257–288. [Google Scholar] [CrossRef]
- Li, G.; Liang, W.; Zhang, X.; Ren, H.; Hu, J.; Bennett, M.J.; Zhang, D. Rice actin-binding protein RMD is a key link in the auxin–actin regulatory loop that controls cell growth. Proc. Natl. Acad. Sci. USA 2014, 111, 10377–10382. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, Y.; Tan, H.; Wang, Y.; Li, G.; Liang, W.; Yuan, Z.; Hu, J.; Ren, H.; Zhang, D. RICE MORPHOLOGY DETERMINANT encodes the type II formin FH5 and regulates rice morphogenesis. Plant Cell 2011, 23, 681–700. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, C.; Xie, M.; Liu, J.; Kong, Z.; Su, H. Actin bundles in the pollen tube. Int. J. Mol. Sci. 2018, 19, 3710. [Google Scholar] [CrossRef]
- Deeks, M.J.; Hussey, P.J.; Davies, B. Formins: Intermediates in signal-transduction cascades that affect cytoskeletal reorganization. Trends Plant Sci. 2002, 7, 492–498. [Google Scholar] [CrossRef]
- Cvrčková, F.; Novotný, M.; Pícková, D.; Žárský, V. Formin homology 2 domains occur in multiple contexts in angiosperms. BMC Genom. 2004, 5, 44. [Google Scholar] [CrossRef]
- Grunt, M.; Žárský, V.; Cvrčková, F. Roots of angiosperm formins: The evolutionary history of plant FH2 domain-containing proteins. BMC Evol. Biol. 2008, 8, 115. [Google Scholar] [CrossRef]
- Kovar, D.R. Molecular details of formin-mediated actin assembly. Curr. Opin. Cell Biol. 2006, 18, 11–17. [Google Scholar] [CrossRef]
- Rivero, F.; Muramoto, T.; Meyer, A.-K.; Urushihara, H.; Uyeda, T.Q.; Kitayama, C. A comparative sequence analysis reveals a common GBD/FH3-FH1-FH2-DAD architecture in formins from Dictyostelium, fungi and metazoa. BMC Genom. 2005, 6, 28. [Google Scholar] [CrossRef] [PubMed]
- Pruyne, D. Probing the origins of metazoan formin diversity: Evidence for evolutionary relationships between metazoan and non-metazoan formin subtypes. PLoS ONE 2017, 12, e0186081. [Google Scholar] [CrossRef] [PubMed]
- Cheung, A.Y.; Wu, H.-M. Overexpression of an Arabidopsis formin stimulates supernumerary actin cable formation from pollen tube cell membrane. Plant Cell 2004, 16, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Cheung, A.Y.; Niroomand, S.; Zou, Y.; Wu, H.-M. A transmembrane formin nucleates subapical actin assembly and controls tip-focused growth in pollen tubes. Proc. Natl. Acad. Sci. USA 2010, 107, 16390–16395. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Zheng, Y.; Yan, A.; Chen, N.; Wang, Z.; Huang, S.; Yang, Z. Arabidopsis formin3 directs the formation of actin cables and polarized growth in pollen tubes. Plant Cell 2009, 21, 3868–3884. [Google Scholar] [CrossRef]
- Li, Y.; Shen, Y.; Cai, C.; Zhong, C.; Zhu, L.; Yuan, M.; Ren, H. The type II Arabidopsis formin14 interacts with microtubules and microfilaments to regulate cell division. Plant Cell 2010, 22, 2710–2726. [Google Scholar] [CrossRef]
- Kollarova, E.; Baquero Forero, A.; Cvrčková, F. The Arabidopsis thaliana class II formin FH13 modulates pollen tube growth. Front. Plant Sci. 2021, 12, 599961. [Google Scholar] [CrossRef]
- Li, G.; Yang, X.; Zhang, X.; Song, Y.; Liang, W.; Zhang, D. Rice morphology determinant-mediated actin filament organization contributes to pollen tube growth. Plant Physiol. 2018, 177, 255–270. [Google Scholar] [CrossRef]
- Hussey, P.J.; Ketelaar, T.; Deeks, M.J. Control of the actin cytoskeleton in plant cell growth. Annu. Rev. Plant Biol. 2006, 57, 109–125. [Google Scholar] [CrossRef]
- Duan, W.-J.; Liu, Z.-H.; Bai, J.-F.; Yuan, S.-H.; Li, Y.-M.; Lu, F.-K.; Zhang, T.-B.; Sun, J.-H.; Zhang, F.-T.; Zhao, C.-P. Comprehensive analysis of formin gene family highlights candidate genes related to pollen cytoskeleton and male fertility in wheat (Triticum aestivum L.). BMC Genom. 2021, 22, 570. [Google Scholar] [CrossRef]
- Oulehlová, D.; Kollárova, E.; Cifrová, P.; Pejchar, P.; Žársky, V.; Cvrčková, F. Arabidopsis class I formin FH1 relocates between membrane compartments during root cell ontogeny and associates with plasmodesmata. Plant Cell Physiol. 2019, 60, 1855–1870. [Google Scholar] [CrossRef]
- Cvrčková, F. Formins and membranes: Anchoring cortical actin to the cell wall and beyond. Front. Plant Sci. 2013, 4, 436. [Google Scholar] [CrossRef] [PubMed]
- Diao, M.; Ren, S.; Wang, Q.; Qian, L.; Shen, J.; Liu, Y.; Huang, S. Arabidopsis formin 2 regulates cell-to-cell trafficking by capping and stabilizing actin filaments at plasmodesmata. eLife 2018, 7, e36316. [Google Scholar] [CrossRef]
- Van Gisbergen, P.A.; Bezanilla, M. Plant formins: Membrane anchors for actin polymerization. Trends Cell Biol. 2013, 23, 227–233. [Google Scholar] [CrossRef]
- Ha, S.; Vankova, R.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.-S.P. Cytokinins: Metabolism and function in plant adaptation to environmental stresses. Trends Plant Sci. 2012, 17, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.-J.; Zhou, Y.-H.; Shi, K.; Zhou, J.; Foyer, C.H.; Yu, J.-Q. Interplay between reactive oxygen species and hormones in the control of plant development and stress tolerance. J. Exp. Bot. 2015, 66, 2839–2856. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Smith, J.A.C.; Harberd, N.P.; Jiang, C. The regulatory roles of ethylene and reactive oxygen species (ROS) in plant salt stress responses. Plant Mol. Biol. 2016, 91, 651–659. [Google Scholar] [CrossRef]
- Xue, X.-H.; Guo, C.-Q.; Du, F.; Lu, Q.-L.; Zhang, C.-M.; Ren, H.-Y. AtFH8 is involved in root development under effect of low-dose latrunculin B in dividing cells. Mol. Plant 2011, 4, 264–278. [Google Scholar] [CrossRef]
- Sun, T.; Li, S.; Ren, H. OsFH15, a class I formin, interacts with microfilaments and microtubules to regulate grain size via affecting cell expansion in rice. Sci. Rep. 2017, 7, 6538. [Google Scholar] [CrossRef]
- Vidali, L.; van Gisbergen, P.A.; Guérin, C.; Franco, P.; Li, M.; Burkart, G.M.; Augustine, R.C.; Blanchoin, L.; Bezanilla, M. Rapid formin-mediated actin-filament elongation is essential for polarized plant cell growth. Proc. Natl. Acad. Sci. USA 2009, 106, 13341–13346. [Google Scholar] [CrossRef] [PubMed]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, 200–204. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, Z.; Li, W.; Cheng, H.; Zhao, W.; Li, T.; Sun, B.; You, Q.; Zhou, D. Genome-Wide Analysis Revealed NBS-LRR Gene Candidates Associated with Bacterial Wilt Resistance in Eggplant (Solanum melongena L.). Agronomy 2023, 13, 2583. [Google Scholar] [CrossRef]
- Chou, K.-C.; Shen, H.-B. Cell-PLoc: A package of Web servers for predicting subcellular localization of proteins in various organisms. Nat. Protoc. 2008, 3, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME suite. Nucleic Acids Res. 2015, 43, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Zhang, Z. KaKs_calculator 3.0: Calculating selective pressure on coding and non-coding sequences. Genom. Proteom. Bioinform. 2022, 20, 536–540. [Google Scholar] [CrossRef]
- Rombauts, S.; Déhais, P.; Van Montagu, M.; Rouzé, P. PlantCARE, a plant cis-acting regulatory element database. Nucleic Acids Res. 1999, 27, 295–296. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Luo, C.; Pei, X.; Wang, S.; Huang, Y.; Li, J.; Liu, B.; Kong, F.; Yang, Q.-Y.; Fang, C. SoyMD: A platform combining multi-omics data with various tools for soybean research and breeding. Nucleic Acids Res. 2024, 52, 1639–1650. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Transcript ID | ORF (bp) | PL (aa) | RMW (kDa) | TIP | Instability Index | Aliphatic Index | GRAVY | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|---|---|
| GmFH1 | Glyma.01G037600 | Glyma.01G037600.1 | 2439 | 812 | 88.77 | 9.32 | 57.22 | 82.25 | −0.433 | Nucleus |
| GmFH2 | Glyma.01G193200 | Glyma.01G193200.1 | 2670 | 889 | 97.5 | 9.22 | 54.85 | 72.63 | −0.59 | Nucleus |
| GmFH3 | Glyma.02G027200 | Glyma.02G027200.1 | 2415 | 804 | 88.16 | 9.43 | 54.34 | 82.7 | −0.423 | Nucleus |
| GmFH4 | Glyma.02G139500 | Glyma.02G139500.1 | 2643 | 880 | 97.24 | 6.36 | 61.3 | 74.9 | −0.504 | Nucleus |
| GmFH5 | Glyma.02G203200 | Glyma.02G203200.1 | 3630 | 1209 | 134.08 | 7.96 | 58.72 | 78.02 | −0.495 | Nucleus |
| GmFH6 | Glyma.03G236400 | Glyma.03G236400.1 | 2388 | 795 | 88.79 | 8.46 | 45.68 | 80.06 | −0.459 | Nucleus |
| GmFH7 | Glyma.04G123600 | Glyma.04G123600.1 | 3621 | 1206 | 130.82 | 8.26 | 59.26 | 79.18 | −0.407 | Nucleus |
| GmFH8 | Glyma.04G177800 | Glyma.04G177800.1 | 1962 | 653 | 72.5 | 7.14 | 54.4 | 80.9 | −0.477 | Nucleus |
| GmFH9 | Glyma.05G024500 | Glyma.05G024500.1 | 3624 | 1207 | 135.12 | 5.42 | 52.91 | 78.09 | −0.442 | Nucleus |
| GmFH10 | Glyma.05G105100 | Glyma.05G105100.1 | 2670 | 889 | 98.03 | 8.98 | 51.62 | 76.99 | −0.47 | Nucleus |
| GmFH11 | Glyma.06G187100 | Glyma.06G187100.1 | 2172 | 723 | 79.85 | 8.37 | 50.15 | 83.73 | −0.385 | Nucleus |
| GmFH12 | Glyma.06G196800 | Glyma.06G196800.1 | 3864 | 1287 | 143 | 6.32 | 54.37 | 76.37 | −0.477 | Nucleus |
| GmFH13 | Glyma.06G266000 | Glyma.06G266000.1 | 2937 | 978 | 105.73 | 7.93 | 55.26 | 80.7 | −0.378 | Nucleus |
| GmFH14 | Glyma.06G301700 | Glyma.06G301700.1 | 2529 | 842 | 91.55 | 6.18 | 52.57 | 78.21 | −0.516 | Nucleus |
| GmFH15 | Glyma.07G058400 | Glyma.07G058400.1 | 2331 | 776 | 85.3 | 5.83 | 61.38 | 78.25 | −0.546 | Nucleus |
| GmFH16 | Glyma.07G206000 | Glyma.07G206000.1 | 2691 | 896 | 99.13 | 6.57 | 62.96 | 74.01 | −0.537 | Nucleus |
| GmFH17 | Glyma.08G292600 | Glyma.08G292600.1 | 2373 | 790 | 85.75 | 9.33 | 52.68 | 81.47 | −0.454 | Nucleus |
| GmFH18 | Glyma.09G215100 | Glyma.09G215100.1 | 3681 | 1226 | 132.5 | 8.69 | 61.14 | 77.9 | −0.4 | Chloroplast |
| GmFH19 | Glyma.09G246200 | Glyma.09G246200.1 | 3114 | 1037 | 112.84 | 6.81 | 70.19 | 75.64 | −0.41 | Chloroplast |
| GmFH20 | Glyma.10G152300 | Glyma.10G152300.1 | 2760 | 919 | 101.62 | 6.73 | 59.57 | 80.39 | −0.502 | Nucleus |
| GmFH21 | Glyma.11G048600 | Glyma.11G048600.1 | 2688 | 895 | 98.36 | 9.33 | 52.58 | 72.79 | −0.61 | Nucleus |
| GmFH22 | Glyma.12G102700 | Glyma.12G102700.1 | 2493 | 830 | 90.54 | 6.57 | 54.96 | 77.07 | −0.499 | Nucleus |
| GmFH23 | Glyma.12G136800 | Glyma.12G136800.1 | 2922 | 973 | 105.28 | 8.78 | 56.68 | 79.32 | −0.388 | Nucleus |
| GmFH24 | Glyma.12G215400 | Glyma.12G215400.1 | 2712 | 903 | 99.99 | 8.55 | 48.17 | 79.47 | −0.503 | Chloroplast |
| GmFH25 | Glyma.13G286100 | Glyma.13G286100.1 | 2724 | 907 | 100.17 | 7.46 | 49.49 | 79.89 | −0.474 | Chloroplast |
| GmFH26 | Glyma.16G027400 | Glyma.16G027400.1 | 3240 | 1079 | 117.13 | 8.61 | 72.52 | 73.03 | −0.534 | Chloroplast |
| GmFH27 | Glyma.17G074500 | Glyma.17G074500.1 | 3627 | 1208 | 134.35 | 8.47 | 57.2 | 78.23 | −0.52 | Nucleus |
| GmFH28 | Glyma.17G102800 | Glyma.17G102800.1 | 4170 | 1389 | 152.57 | 6.39 | 63.01 | 71.48 | −0.532 | Nucleus |
| GmFH29 | Glyma.17G161400 | Glyma.17G161400.1 | 2655 | 884 | 97.56 | 9.21 | 54.7 | 73.35 | −0.533 | Nucleus |
| GmFH30 | Glyma.17G222600 | Glyma.17G222600.1 | 4395 | 1464 | 162.72 | 6.29 | 52.35 | 82.6 | −0.419 | Nucleus |
| GmFH31 | Glyma.18G130800 | Glyma.18G130800.1 | 2325 | 774 | 84.56 | 9.23 | 53.04 | 82.78 | −0.457 | Nucleus |
| GmFH32 | Glyma.18G246800 | Glyma.18G246800.1 | 3120 | 1039 | 113.58 | 6.69 | 70.01 | 76.25 | −0.413 | Chloroplast |
| GmFH33 | Glyma.19G234000 | Glyma.19G234000.1 | 2376 | 791 | 88.21 | 8.24 | 48.31 | 80.34 | −0.468 | Nucleus |
| GmFH34 | Glyma.20G236100 | Glyma.20G236100.1 | 2679 | 892 | 98.55 | 7.23 | 59.97 | 78.46 | −0.51 | Nucleus |
| Gene Pairs | Ka/Ks | Gene Pairs | Ka/Ks |
|---|---|---|---|
| GmFH1/GmFH3 | 0.31 | GmFH9/GmFH27 | 0.18 |
| GmFH1/GmFH8 | 0.19 | GmFH9/GmFH28 | 0.30 |
| GmFH1/GmFH11 | 0.22 | GmFH10/GmFH21 | 0.19 |
| GmFH1/GmFH17 | 0.36 | GmFH10/GmFH29 | 0.19 |
| GmFH1/GmFH31 | 0.19 | GmFH11/GmFH17 | 0.21 |
| GmFH2/GmFH10 | 0.22 | GmFH11/GmFH31 | 0.26 |
| GmFH2/GmFH21 | 0.17 | GmFH12/GmFH28 | 0.25 |
| GmFH2/GmFH29 | 0.20 | GmFH13/GmFH23 | 0.30 |
| GmFH3/GmFH8 | 0.20 | GmFH13/GmFH24 | 0.37 |
| GmFH3/GmFH11 | 0.25 | GmFH13/GmFH25 | 0.33 |
| GmFH3/GmFH17 | 0.36 | GmFH14/GmFH22 | 0.21 |
| GmFH3/GmFH31 | NaN | GmFH14/GmFH24 | 0.19 |
| GmFH4/GmFH16 | 0.20 | GmFH15/GmFH26 | 0.08 |
| GmFH5/GmFH27 | 0.26 | GmFH17/GmFH31 | NaN |
| GmFH6/GmFH20 | 0.26 | GmFH19/GmFH31 | 0.32 |
| GmFH6/GmFH33 | 0.33 | GmFH20/GmFH33 | 0.23 |
| GmFH6/GmFH34 | 0.28 | GmFH20/GmFH34 | 0.22 |
| GmFH7/GmFH18 | 0.17 | GmFH21/GmFH29 | 0.20 |
| GmFH8/GmFH11 | 0.16 | GmFH23/GmFH24 | 0.32 |
| GmFH8/GmFH17 | 0.21 | GmFH23/GmFH25 | 0.30 |
| GmFH8/GmFH31 | 0.24 | GmFH24/GmFH25 | 0.33 |
| GmFH9/GmFH12 | 0.24 | GmFH33/GmFH34 | 0.24 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Zhang, Z.; Shan, M.; Amjad, Z.; Xue, J.; Zhang, Z.; Wang, J.; Guo, Y. Genome-Wide Studies of FH Family Members in Soybean (Glycine max) and Their Responses under Abiotic Stresses. Plants 2024, 13, 276. https://doi.org/10.3390/plants13020276
Zhang Z, Zhang Z, Shan M, Amjad Z, Xue J, Zhang Z, Wang J, Guo Y. Genome-Wide Studies of FH Family Members in Soybean (Glycine max) and Their Responses under Abiotic Stresses. Plants. 2024; 13(2):276. https://doi.org/10.3390/plants13020276
Chicago/Turabian StyleZhang, Zhenbiao, Zhongqi Zhang, Muhammad Shan, Zarmeena Amjad, Jin Xue, Zenglin Zhang, Jie Wang, and Yongfeng Guo. 2024. "Genome-Wide Studies of FH Family Members in Soybean (Glycine max) and Their Responses under Abiotic Stresses" Plants 13, no. 2: 276. https://doi.org/10.3390/plants13020276
APA StyleZhang, Z., Zhang, Z., Shan, M., Amjad, Z., Xue, J., Zhang, Z., Wang, J., & Guo, Y. (2024). Genome-Wide Studies of FH Family Members in Soybean (Glycine max) and Their Responses under Abiotic Stresses. Plants, 13(2), 276. https://doi.org/10.3390/plants13020276