

In the Qaidam Basin, Soil Nutrients Directly or Indirectly Affect Desert Ecosystem Stability under Drought Stress through Plant Nutrients


Abstract
1. Introduction
2. Materials and Methods
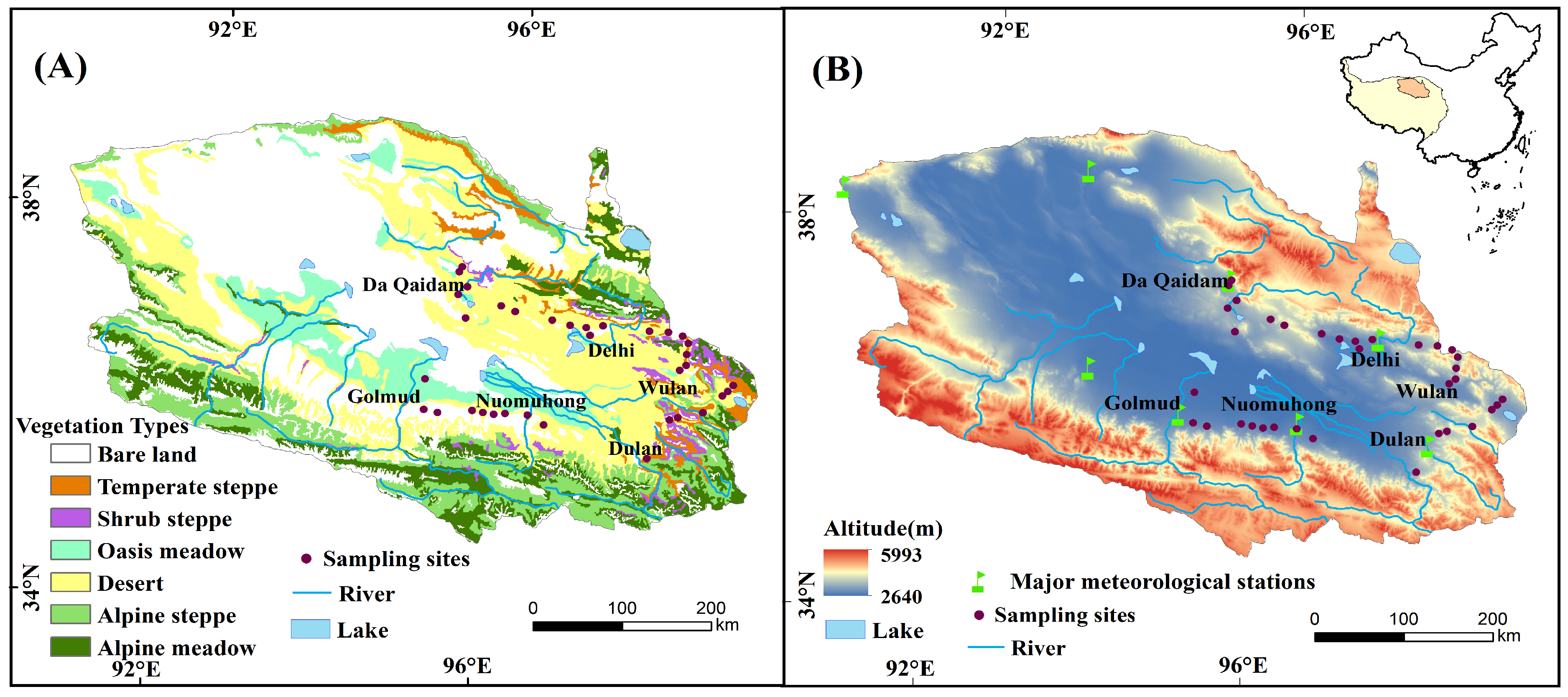
2.1. Study Area
2.2. Field Survey and Sampling
2.3. Experimental Methods and Index Calculation
2.4. Climate Data Collection and Processing
2.5. Climate Data Collection and Processing
2.6. Data Analysis
3. Results
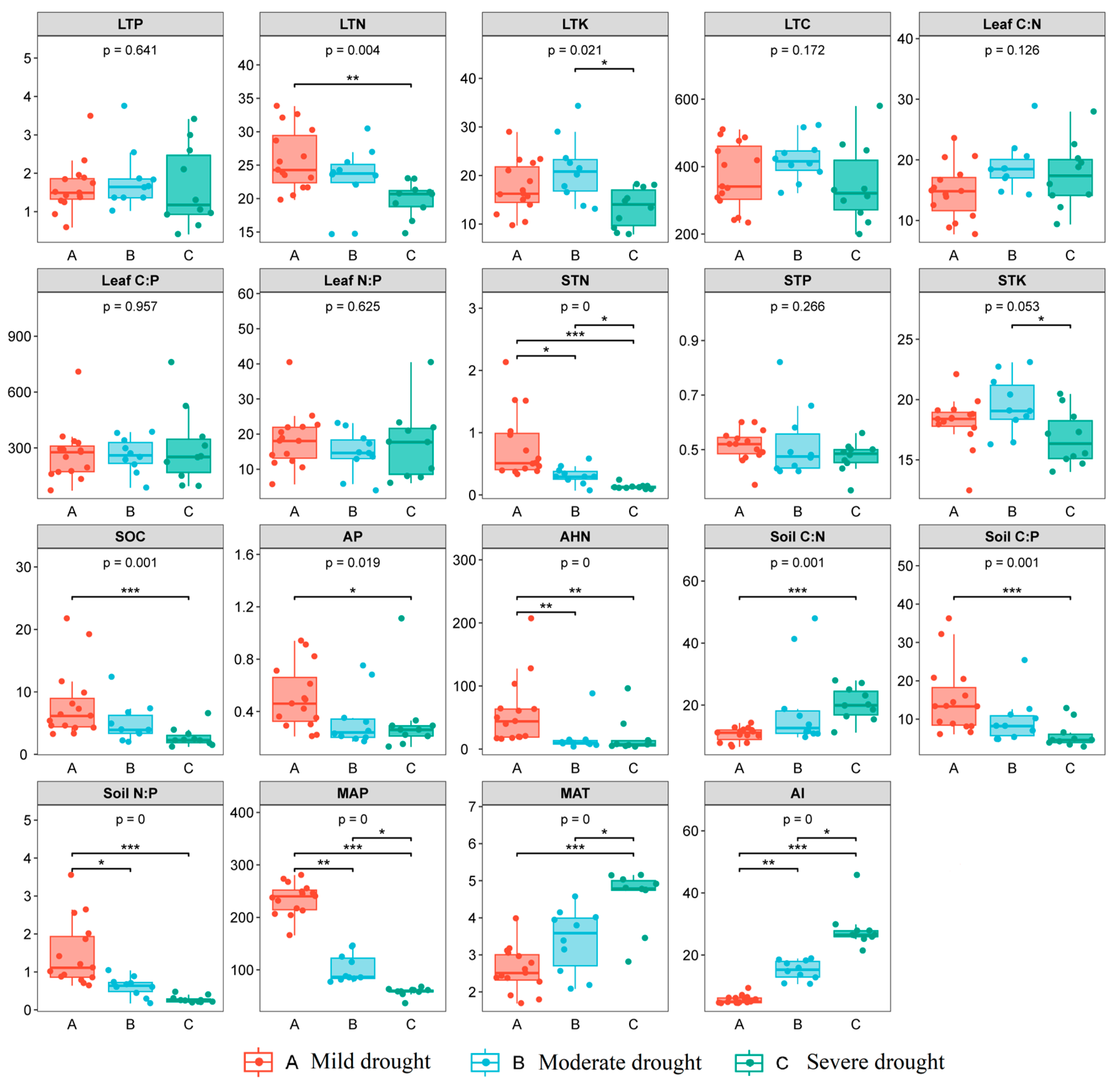
3.1. Variation Characteristics of Soil and Plant Nutrient Elements along Drought Gradient
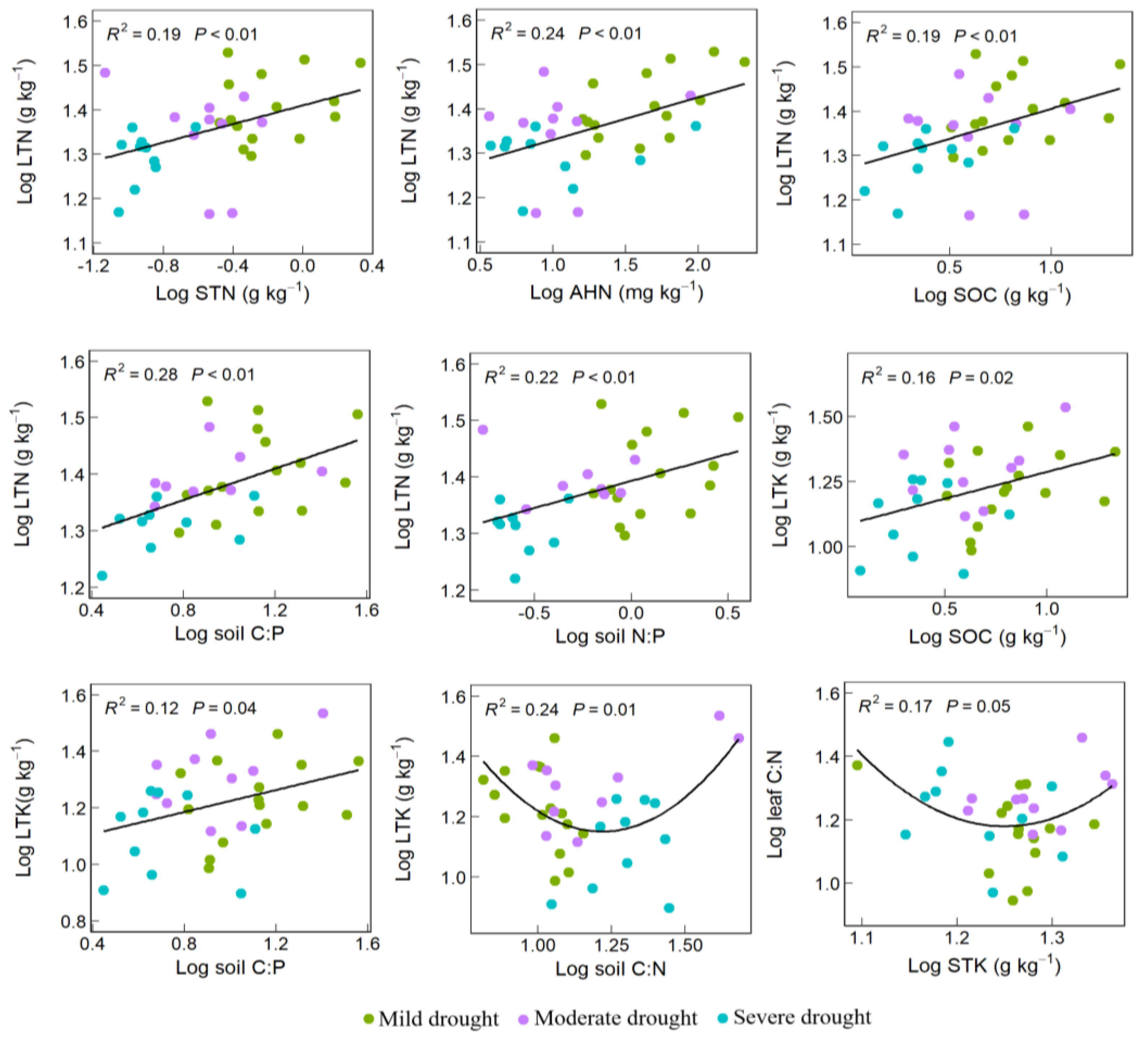
3.2. Soil–Plant Nutrient Element Relationship of Desert Vegetation
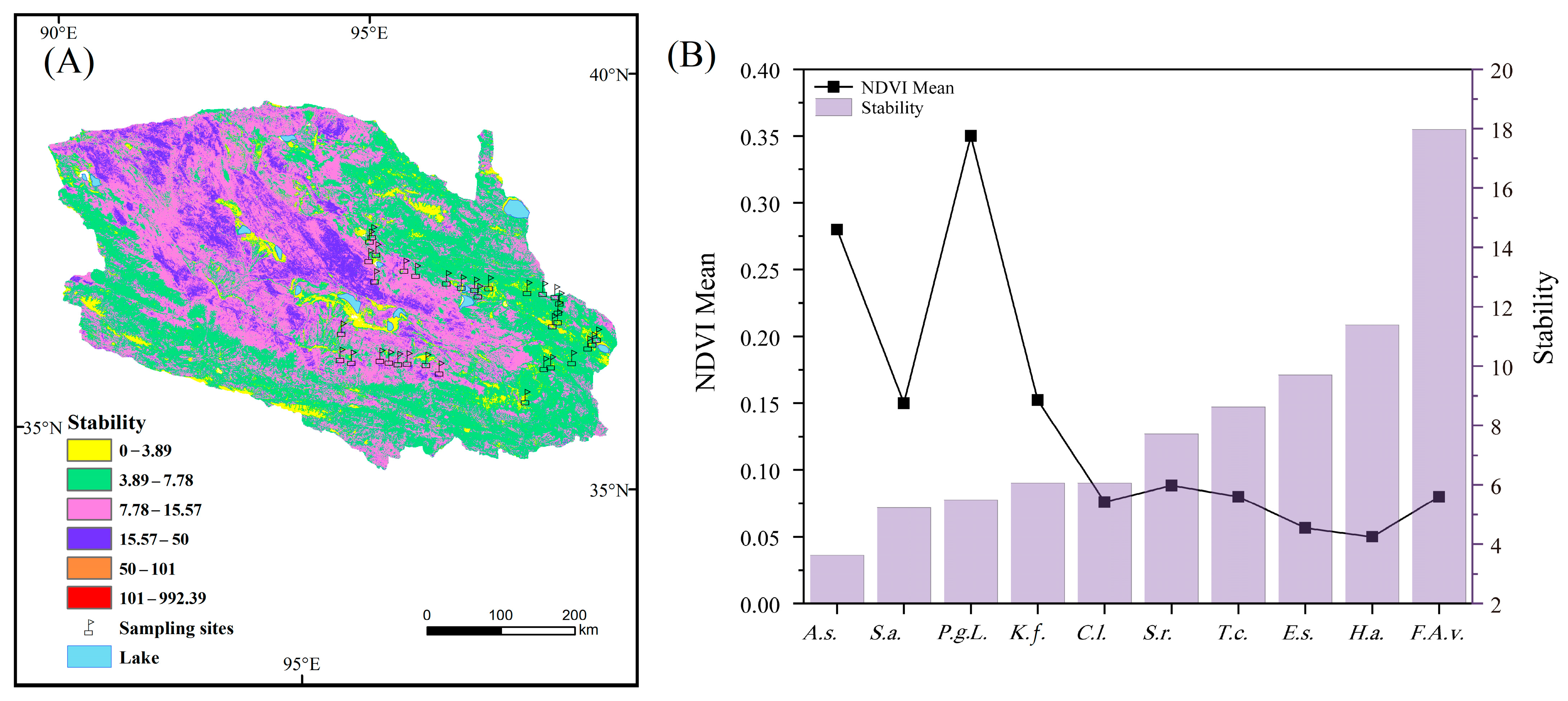
3.3. Productivity Change Trend and Ecosystem Stability in the Qaidam Basin
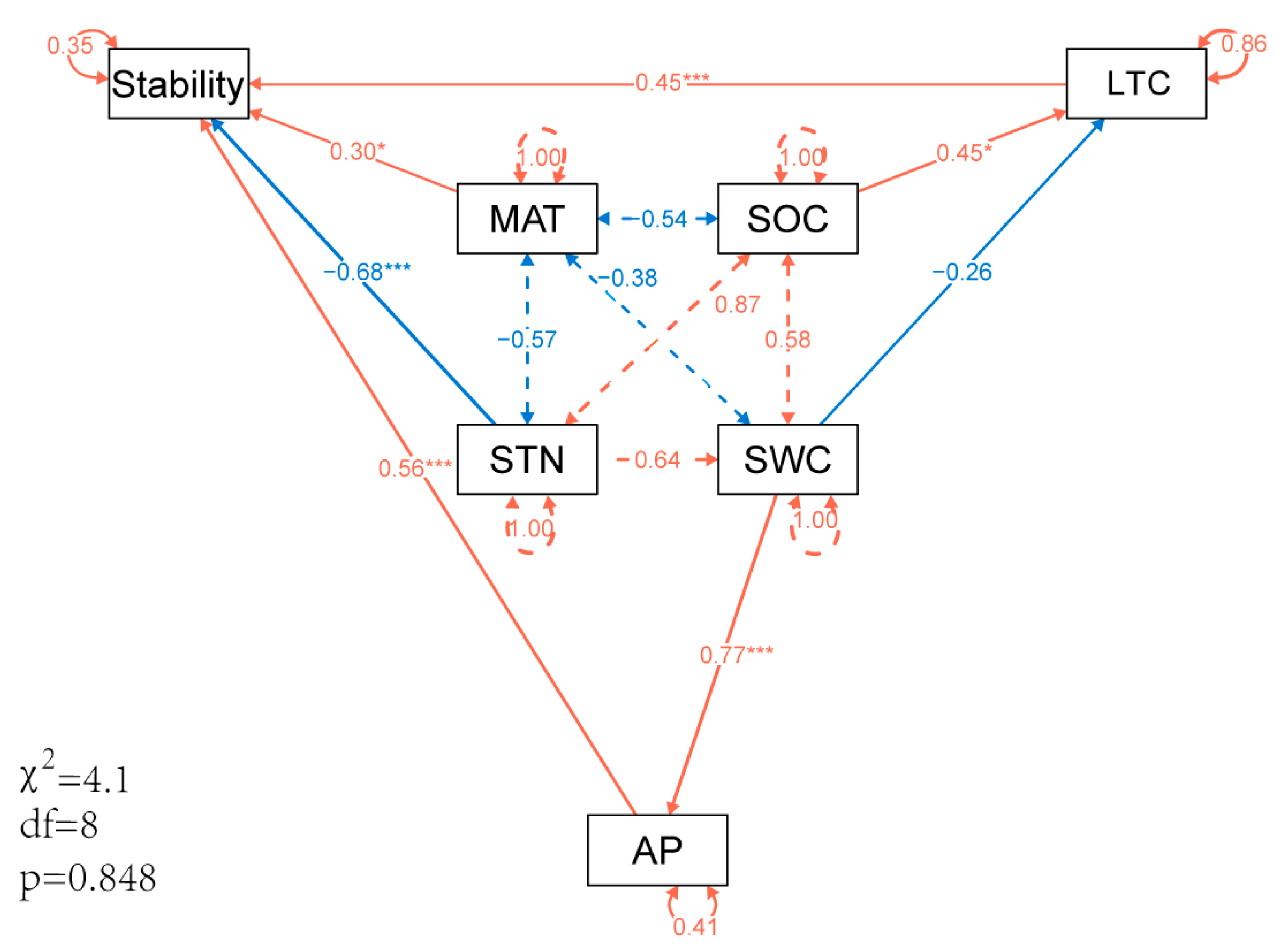
3.4. Effects of Soil Nutrients, Plant Nutrients, and Environmental Factors on Ecosystem Stability
4. Discussion
4.1. Effects of Arid Environment on Soil and Plant Nutrients
4.2. Soil–Plant Nutrient Element Relationship
4.3. Influence Pathway of Soil, Plant Nutrients and Climate Factors on Ecosystem Stability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feng, S.; Gu, X.; Luo, S.; Liu, R.; Gulakhmadov, A.; Slater, L.J.; Li, J.; Zhang, X.; Kong, D. Greenhouse gas emissions drive global dryland expansion but not spatial patterns of change in aridification. J. Clim. 2022, 35, 6501–6517. [Google Scholar] [CrossRef]
- Zeng, H.; Wu, B.; Zhang, M.; Zhang, N.; Elnashar, A.; Zhu, L.; Zhu, W.; Wu, F.; Yan, N.; Liu, W. Dryland ecosystem dynamic change and its drivers in mediterranean region. Curr. Opin. Environ. Sustain. 2021, 48, 59–67. [Google Scholar] [CrossRef]
- Berdugo, M.; Delgado-Baquerizo, M.; Soliveres, S.; Hernández-Clemente, R.; Zhao, Y.; Gaitán, J.J.; Gross, N.; Saiz, H.; Maire, V.; Lehmann, A.; et al. Global ecosystem thresholds driven by aridity. Science 2020, 367, 787–790. [Google Scholar] [CrossRef] [PubMed]
- Maestre, F.T.; Eldridge, D.J.; Soliveres, S.; Kéfi, S.; Delgado-Baquerizo, M.; Bowker, M.A.; García-Palacios, P.; Gaitán, J.; Gallardo, A.; Lázaro, R.; et al. Structure and functioning of dryland ecosystems in a changing world. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 215–237. [Google Scholar] [CrossRef] [PubMed]
- Báez, S.; Collins, S.L.; Pockman, W.T.; Johnson, J.E.; Small, E.E. Effects of experimental rainfall manipulations on chihuahuan desert grassland and shrubland plant communities. Oecologia 2013, 172, 1117–1127. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Zhou, H.; Cui, Y.; Zhao, W.; Hou, Y.; Tan, C.; Yang, G.; Ding, F. Stoichiometric characteristics of leaf, litter and soil during vegetation succession in maolan national nature reserve, guizhou, china. Sustainability 2022, 14, 16517. [Google Scholar] [CrossRef]
- Pugnaire, F.I.; Morillo, J.A.; Penuelas, J.; Reich, P.B.; Bardgett, R.D.; Gaxiola, A.; Wardle, D.A.; van der Putten, W.H. Climate change effects on plant-soil feedbacks and consequences for biodiversity and functioning of terrestrial ecosystems. Sci. Adv. 2019, 5, eaaz1834. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Chen, G.; Zhang, C.; Melillo, J.M.; Hall, C.A.S. Pattern and variation of c:N:P ratios in china’s soils: A synthesis of observational data. Biogeochemistry 2009, 98, 139–151. [Google Scholar] [CrossRef]
- Fan, B.; Westerband, A.C.; Wright, I.J.; Gao, P.; Ding, N.; Ai, D.; Tian, T.; Zhao, X.; Sun, K. Shifts in plant resource use strategies across climate and soil gradients in dryland steppe communities. Plant Soil 2023, 497, 277–296. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf n and p in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Han, W.; Tang, L.; Tang, Z.; Fang, J. Leaf nitrogen and phosphorus concentrations of woody plants differ in responses to climate, soil and plant growth form. Ecography 2013, 36, 178–184. [Google Scholar] [CrossRef]
- He, M.; Dijkstra, F.A.; Zhang, K.; Li, X.; Tan, H.; Gao, Y.; Li, G. Leaf nitrogen and phosphorus of temperate desert plants in response to climate and soil nutrient availability. Sci. Rep. 2014, 4, 6932. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Tang, G.; Luo, H.; Yin, H.; Zhang, Z.; Xue, J.; Huang, C.; Lu, Y.; Shareef, M.; Gao, X.; et al. Topsoil nutrients drive leaf carbon and nitrogen concentrations of a desert phreatophyte in habitats with different shallow groundwater depths. Water 2021, 13, 3093. [Google Scholar] [CrossRef]
- Xu, H.P.; Zhang, J.; Pang, X.P.; Wang, Q.; Zhang, W.N.; Wang, J.; Guo, Z.G. Responses of plant productivity and soil nutrient concentrations to different alpine grassland degradation levels. Environ. Monit. Assess. 2019, 191, 678. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, R.; Gao, J. Precipitation and soil nutrients determine the spatial variability of grassland productivity at large scales in china. Front. Plant Sci. 2022, 13, 996313. [Google Scholar] [CrossRef] [PubMed]
- Booth, M.S.; Stark, J.M.; Rastetter, E. Controls on nitrogen cycling in terrestrial ecosystems: A synthetic analysis of literature data. Ecol. Monogr. 2005, 75, 139–157. [Google Scholar] [CrossRef]
- Lambers, H.; Raven, J.A.; Shaver, G.R.; Smith, S.E. Plant nutrient-acquisition strategies change with soil age. Trends Ecol. Evol. 2008, 23, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Fujita, Y.; van Bodegom, P.M.; Witte, J.-P.M. Relationships between nutrient-related plant traits and combinations of soil n and p fertility measures. PLoS ONE 2014, 8, e83735. [Google Scholar] [CrossRef]
- Li, R.; Wu, P.-P.; Peng, C.; Shi, F.-X.; Mao, R. Shifted plant composition predominantly controls nitrogen addition effect on community-level leaf nutrient resorption in a boreal peatland. Plant Soil 2024, 494, 321–331. [Google Scholar] [CrossRef]
- Huang, M.J.; de Vries, J.; Zhou, S.; Hautier, Y. Aridity and soil fertility, not species richness, interact to affect temporal stability of primary productivity along a natural gradient in northern china. Oikos 2023, 2023, e10086. [Google Scholar] [CrossRef]
- Li, T.; Kamran, M.; Chang, S.; Peng, Z.; Wang, Z.; Ran, L.; Jiang, W.Q.; Jin, Y.; Zhang, X.; You, Y.; et al. Climate-soil interactions improve the stability of grassland ecosystem by driving alpine plant diversity. Ecol. Indic. 2022, 141, 109002. [Google Scholar] [CrossRef]
- Liu, L.; Zhu, K.; Wurzburger, N.; Zhang, J. Relationships between plant diversity and soil microbial diversity vary across taxonomic groups and spatial scales. Ecosphere 2020, 11, e02999. [Google Scholar] [CrossRef]
- Tognetti, P.M.; Prober, S.M.; Báez, S.; Chaneton, E.J.; Firn, J.; Risch, A.C.; Schuetz, M.; Simonsen, A.K.; Yahdjian, L.; Borer, E.T.; et al. Negative effects of nitrogen override positive effects of phosphorus on grassland legumes worldwide. Proc. Natl. Acad. Sci. USA 2021, 118, e2023718118. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jiang, L.; Wang, Z.; Song, M.; Wang, S. Phosphorus enrichment increased community stability by increasing asynchrony and dominant species stability in alpine meadow of qinghai-tibet plateau. J. Geophys. Res.-Biogeosci. 2022, 127, e2022JG006819. [Google Scholar] [CrossRef]
- Xu, Q.; Yang, X.; Yan, Y.; Wang, S.; Loreau, M.; Jiang, L. Consistently positive effect of species diversity on ecosystem, but not population, temporal stability. Ecol. Lett. 2021, 24, 2256–2266. [Google Scholar] [CrossRef] [PubMed]
- Orwin, K.H.; Dickie, I.A.; Wood, J.R.; Bonner, K.I.; Holdaway, R.J. Soil microbial community structure explains the resistance of respiration to a dry–rewet cycle, but not soil functioning under static conditions. Funct. Ecol. 2016, 30, 1430–1439. [Google Scholar] [CrossRef]
- Wang, X.; Yang, M.; Liang, X.; Pang, G.; Wan, G.; Chen, X.; Luo, X. The dramatic climate warming in the qaidam basin, northeastern tibetan plateau, during 1961–2010. Int. J. Climatol. 2014, 34, 1524–1537. [Google Scholar] [CrossRef]
- Zeng, F.; Zhang, X.; Zhan, T.; Zhang, Z.; Chen, L.; Chen, L.; Ji, M. Rapid warming and increasing moisture levels in the qaidam basin. Theor. Appl. Climatol. 2024. [Google Scholar] [CrossRef]
- Naeem, S.; Li, S. Biodiversity enhances ecosystem reliability. Nature 1997, 390, 507–509. [Google Scholar] [CrossRef]
- Niu, W.; Chen, H.; Wu, J. Soil moisture and soluble salt content dominate changes in foliar δ13c and δ15n of desert communities in the qaidam basin, qinghai-tibetan plateau. Front. Plant Sci. 2021, 12, 675817. [Google Scholar] [CrossRef]
- Kang, Y.; Guo, E.; Wang, Y.; Bao, Y.; Bao, Y.; Mandula, N. Monitoring vegetation change and its potential drivers in inner mongolia from 2000 to 2019. Remote Sens. 2021, 13, 3357. [Google Scholar] [CrossRef]
- Zhang, Y.; He, N.; Loreau, M.; Pan, Q.; Han, X. Scale dependence of the diversity-stability relationship in a temperate grassland. J. Ecol. 2018, 106, 1277–1285. [Google Scholar] [CrossRef] [PubMed]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef]
- Tan, Q.; Jia, Y.; Wang, G. Decoupling of soil nitrogen and phosphorus dynamics along a temperature gradient on the qinghai-tibetan plateau. Geoderma 2021, 396, 115084. [Google Scholar] [CrossRef]
- Woods, H.A.; Makino, W.; Cotner, J.B.; Hobbie, S.E.; Harrison, J.F.; Acharya, K.; Elser, J.J. Temperature and the chemical composition of poikilothermic organisms. Funct. Ecol. 2003, 17, 237–245. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, W.; Ge, X.; Zheng, X.; Zhou, X.; Ding, H.; Zhang, A. Response of plant and soil n, p, and n:P stoichiometry to n addition in china: A meta-analysis. Plants 2023, 12, 2104. [Google Scholar] [CrossRef]
- Joly, F.-X.; Kurupas, K.L.; Throop, H.L. Pulse frequency and soil-litter mixing alter the control of cumulative precipitation over litter decomposition. Ecology 2017, 98, 2255–2260. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Baquerizo, M.; Maestre, F.T.; Gallardo, A.; Bowker, M.A.; Wallenstein, M.D.; Quero, J.L.; Ochoa, V.; Gozalo, B.; García-Gómez, M.; Soliveres, S.; et al. Decoupling of soil nutrient cycles as a function of aridity in global drylands. Nature 2013, 502, 672–676. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Peng, Q.; Li, K.; Gong, Y.; Liu, Y.; Han, W. Patterns of nitrogen and phosphorus stoichiometry among leaf, stem and root of desert plants and responses to climate and soil factors in xinjiang, china. Catena 2021, 199, 105100. [Google Scholar] [CrossRef]
- Hedin, L.O. Global organization of terrestrial plant–nutrient interactions. Proc. Natl. Acad. Sci. USA 2004, 101, 10849–10850. [Google Scholar] [CrossRef] [PubMed]
- Sardans, J.; Peñuelas, J. Tree growth changes with climate and forest type are associated with relative allocation of nutrients, especially phosphorus, to leaves and wood. Glob. Ecol. Biogeogr. 2013, 22, 494–507. [Google Scholar] [CrossRef]
- Heineman, K.D.; Turner, B.L.; Dalling, J.W. Variation in wood nutrients along a tropical soil fertility gradient. New Phytol. 2016, 211, 440–454. [Google Scholar] [CrossRef] [PubMed]
- Andersen, K.M.; Corre, M.D.; Turner, B.L.; Dalling, J.W. Plant–soil associations in a lower montane tropical forest: Physiological acclimation and herbivore-mediated responses to nitrogen addition. Funct. Ecol. 2010, 24, 1171–1180. [Google Scholar] [CrossRef]
- Han, W.X.; Fang, J.Y.; Guo, D.L.; Zhang, Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in china. New Phytol. 2005, 168, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Hayes, P.; Turner, B.L.; Lambers, H.; Laliberte, E. Foliar nutrient concentrations and resorption efficiency in plants of contrasting nutrient-acquisition strategies along a 2-million-year dune chronosequence. J. Ecol. 2014, 102, 396–410. [Google Scholar] [CrossRef]
- Luo, W.; Li, M.-H.; Sardans, J.; Lü, X.-T.; Wang, C.; Peñuelas, J.; Wang, Z.; Han, X.-G.; Jiang, Y. Carbon and nitrogen allocation shifts in plants and soils along aridity and fertility gradients in grasslands of china. Ecol. Evol. 2017, 7, 6927–6934. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Gao, X.; Li, L.; Lu, Y.; Shareef, M.; Huang, C.; Liu, G.; Gui, D.; Zeng, F. Groundwater depth affects phosphorus but not carbon and nitrogen concentrations of a desert phreatophyte in northwest china. Front. Plant Sci. 2018, 9, 338. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Zheng, H.; Zhang, B.; Tariq, A.; Lv, G.; Zeng, F.; Graciano, C. Stoichiometry of c:N:P in the roots of alhagi sparsifolia is more sensitive to soil nutrients than aboveground organs. Front. Plant Sci. 2021, 12, 698961. [Google Scholar] [CrossRef] [PubMed]
- Schimel, D. Nutrient cycling and limitation: Hawai’i as a model system. Nature 2004, 431, 630–631. [Google Scholar] [CrossRef]
- He, M.; Zhang, K.; Tan, H.; Hu, R.; Su, J.; Wang, J.; Huang, L.; Zhang, Y.; Li, X. Nutrient levels within leaves, stems, and roots of the xeric species reaumuria soongorica in relation to geographical, climatic, and soil conditions. Ecol. Evol. 2015, 5, 1494–1503. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z. Relationship of Main Nutrient Elements in Soil and Vegetation in Qaidam Basin. Master’s Thesis, Hebei Normal University, Shijiazhuang, China, 2021; p. 84. [Google Scholar] [CrossRef]
- Wang, L.; Zhao, G.; Li, M.; Zhang, M.; Zhang, L.; Zhang, X.; An, L.; Xu, S. C:N:P stoichiometry and leaf traits of halophytes in an arid saline environment, northwest china. PLoS ONE 2015, 10, e0119935. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.S.; Shi, P.L.; Wu, J.S.; Xiong, D.P.; Zong, N.; Zhang, X.Z. Foliar nutrient resorption patterns of four functional plants along a precipitation gradient on the tibetan changtang plateau. Ecol. Evol. 2017, 7, 7201–7212. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Yang, X.; Sun, X.; Chen, W.Q.; Yang, G.W.; Liu, N.; Chen, J.S.; Zhang, Y.J. Increased precipitation modulates the influence of nitrogen and litter inputs on the nutrient resorption proficiency rather than efficiency of leymus chinensis. Plant Ecol. 2018, 219, 217–230. [Google Scholar] [CrossRef]
- Glenn, E.P.; Brown, J.J.; Blumwald, E. Salt tolerance and crop potential of halophytes. Crit. Rev. Plant Sci. 1999, 18, 227–255. [Google Scholar] [CrossRef]
- Pessarakli, M.; Harivandi, M.A.; Kopec, D.M.; Ray, D.T. Growth responses and nitrogen uptake by saltgrass (Distichlis spicata L.), a halophytic plant species, under salt stress, using the 15n technique. Int. J. Agron. 2012, 2012, 896971. [Google Scholar] [CrossRef]
- Rahnama, A.; James, R.A.; Poustini, K.; Munns, R. Stomatal conductance as a screen for osmotic stress tolerance in durum wheat growing in saline soil. Funct. Plant Biol. 2010, 37, 255–263. [Google Scholar] [CrossRef]
- Huang, K.; Xia, J. High ecosystem stability of evergreen broadleaf forests under severe droughts. Glob. Chang. Biol. 2019, 25, 3494–3503. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Chi, Y.; Chen, J.; Zhou, L. Global climate regulates dimensions of terrestrial ecosystem stability. Ecosphere 2023, 14, e4577. [Google Scholar] [CrossRef]
- Yu, H.; Ma, Q.; Liu, X.; Li, Y.; Li, L.; Qi, M.; Wu, W.; Wang, Y.; Xu, Z.; Zhou, G.; et al. Resistance, recovery, and resilience of desert steppe to precipitation alterations with nitrogen deposition. J. Clean. Prod. 2021, 317, 128434. [Google Scholar] [CrossRef]
- Chen, B.; Chen, H.; Li, M.; Fiedler, S.; Mărgărint, M.C.; Nowak, A.; Wesche, K.; Tietjen, B.; Wu, J. Climate sensitivity of the arid scrublands on the tibetan plateau mediated by plant nutrient traits and soil nutrient availability. Remote Sens. 2022, 14, 4601. [Google Scholar] [CrossRef]
- Balazs, K.R.; Munson, S.M.; Butterfield, B.J. Functional composition of plant communities mediates biomass effects on ecosystem service recovery across an experimental dryland restoration network. Funct. Ecol. 2022, 36, 2317–2330. [Google Scholar] [CrossRef]
- Song, M.-H.; Zong, N.; Jiang, J.; Shi, P.-L.; Zhang, X.-Z.; Gao, J.-Q.; Zhou, H.-K.; Li, Y.-K.; Loreau, M. Nutrient-induced shifts of dominant species reduce ecosystem stability via increases in species synchrony and population variability. Sci. Total Environ. 2019, 692, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, X.; Ma, Q.; Zhang, X.; Chen, Y.; Isbell, F.; Wang, D. Nitrogen addition reduced ecosystem stability regardless of its impacts on plant diversity. J. Ecol. 2019, 107, 2427–2435. [Google Scholar] [CrossRef]
- Zhang, T.; Ji, X.; Xu, M.; Zhao, G.; Zheng, Z.; Tang, Y.; Chen, N.; Zhu, J.; He, Y.; Zhang, Y. Influences of drought on the stability of an alpine meadow ecosystem. Ecosyst. Health Sustain. 2022, 8, 2110523. [Google Scholar] [CrossRef]
- Liu, X.; Ma, Q.; Yu, H.; Li, Y.; Li, L.; Qi, M.; Wu, W.; Zhang, F.; Wang, Y.; Zhou, G.; et al. Climate warming-induced drought constrains vegetation productivity by weakening the temporal stability of the plant community in an arid grassland ecosystem. Agric. For. Meteorol. 2021, 307, 108526. [Google Scholar] [CrossRef]
- Gao, Q.; Ganjurjav, H.; Hu, G.; Xu, H.; Schwartz, M.W.; Gornish, E.S.; Zhu, W. Warming diminishes the stability of primary productivity in global grass- and forb-dominated ecosystems. Environ. Res. Commun. 2022, 4, 121006. [Google Scholar] [CrossRef]
- Shi, Z.; Xu, X.; Souza, L.; Wilcox, K.; Jiang, L.; Liang, J.; Xia, J.; García-Palacios, P.; Luo, Y. Dual mechanisms regulate ecosystem stability under decade-long warming and hay harvest. Nat. Commun. 2016, 7, 11973. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Xiao, P.; Yu, H.; Zhao, T.; Liu, S.; Yang, L.; He, Y.; Luo, Y.; Wang, X.; Dong, W.; et al. Effects of climate changes on the pasture productivity from 1961 to 2016 in sichuan yellow river source, qinghai-tibet plateau, china. Front. Ecol. Evol. 2022, 10, 908924. [Google Scholar] [CrossRef]
- Gao, Y.; Zhou, X.; Wang, Q.; Wang, C.; Zhan, Z.; Chen, L.; Yan, J.; Qu, R. Vegetation net primary productivity and its response to climate change during 2001–2008 in the tibetan plateau. Sci. Total Environ. 2013, 444, 356–362. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Lat. (°N) | Long. (°E) | Alt. (m) | Soil Type | Community Type | MAP (mm) | MAT (°C) | AI | Drought Gradient |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 36.35 | 95.09 | 2889 | Brown calcic soil | Sympegma regelii | 58.49 | 5.15 | 27.80 | severe drought |
| 2 | 36.33 | 95.27 | 2968 | Grey brown desert soil | Ephedra sinica | 62.87 | 5.03 | 25.14 | severe drought |
| 3 | 36.38 | 95.70 | 2847 | Grey brown desert soil | Ephedra sinica | 60.8 | 4.8 | 26.49 | severe drought |
| 4 | 36.37 | 95.85 | 2890 | Grey brown desert soil | Sympegma regelii | 61.33 | 4.75 | 25.83 | severe drought |
| 5 | 36.36 | 95.98 | 2824 | Grey brown desert soil | Tamarix chinensis Lour | 61.01 | 4.74 | 26.28 | severe drought |
| 6 | 36.38 | 96.13 | 2765 | Grey brown desert soil | Tamarix chinensis Lour | 57.78 | 4.77 | 27.80 | severe drought |
| 7 | 36.38 | 96.41 | 2853 | Grey brown desert soil | Ceratoides latens | 53.31 | 4.91 | 29.82 | severe drought |
| 8 | 36.29 | 96.63 | 2847 | Grey brown desert soil | Haloxylon ammodendron | 85.12 | 4.57 | 18.48 | moderate drought |
| 9 | 36.03 | 97.97 | 3160 | Brown calcic soil | Achnatherum splendens | 203.83 | 3.11 | 7.09 | mild drought |
| 10 | 36.44 | 98.23 | 3255 | Brown calcic soil | Achnatherum splendens | 206.2 | 3.17 | 6.40 | mild drought |
| 11 | 36.47 | 98.33 | 3320 | Brown calcic soil | Kalidium foliatum | 216.75 | 2.96 | 5.78 | mild drought |
| 12 | 36.54 | 98.65 | 3530 | Chestnut soil | Kalidium foliatum | 244.51 | 2.38 | 4.47 | mild drought |
| 13 | 36.72 | 98.88 | 3173 | Chestnut soil | Kalidium foliatum | 267.19 | 1.9 | 4.96 | mild drought |
| 14 | 36.77 | 98.95 | 3069 | Chestnut soil | Kalidium foliatum | 273.07 | 1.79 | 4.78 | mild drought |
| 15 | 36.84 | 99.01 | 3125 | Chestnut soil | Achnatherum splendens | 280.46 | 1.69 | 4.72 | mild drought |
| 16 | 36.96 | 98.31 | 2945 | Chestnut soil | Sympegma regelii | 231.44 | 2.78 | 6.20 | mild drought |
| 17 | 37.02 | 98.39 | 3132 | Chestnut soil | Kalidium foliatum | 240.11 | 2.61 | 5.69 | mild drought |
| 18 | 37.13 | 98.39 | 3276 | Chestnut soil | Kalidium foliatum | 248.91 | 2.43 | 4.70 | mild drought |
| 19 | 37.24 | 98.40 | 3379 | Chestnut soil | Achnatherum splendens | 255.04 | 2.27 | 4.43 | mild drought |
| 20 | 37.32 | 98.32 | 3503 | Chestnut soil | Achnatherum splendens | 245.76 | 2.37 | 4.52 | mild drought |
| 21 | 37.35 | 98.13 | 3466 | Chestnut soil | Potentilla glabra Lodd | 237.33 | 2.51 | 4.61 | mild drought |
| 22 | 37.34 | 97.89 | 3204 | Chestnut soil | Salsola abrotanoides | 212.62 | 3.05 | 6.12 | mild drought |
| 23 | 37.37 | 97.28 | 2971 | Grey brown desert soil | Kalidium foliatum | 165.65 | 3.98 | 9.33 | mild drought |
| 24 | 37.26 | 97.13 | 2862 | Grey brown desert soil | Ceratoides latens | 145.96 | 4.14 | 10.65 | moderate drought |
| 25 | 37.33 | 97.07 | 2905 | Grey brown desert soil | Kalidium foliatum | 144.17 | 4.01 | 10.80 | moderate drought |
| 26 | 37.35 | 96.86 | 2829 | Inland salt soil | Kalidium foliatum | 124.76 | 3.94 | 12.71 | moderate drought |
| 27 | 37.38 | 96.62 | 2982 | Grey brown desert soil | Sympegma regelii | 114.6 | 3.79 | 13.57 | moderate drought |
| 28 | 37.44 | 96.13 | 3621 | Grey brown desert soil | Salsola abrotanoides | 87.46 | 3.38 | 14.73 | moderate drought |
| 29 | 37.49 | 95.95 | 3336 | Grey brown desert soil | Sympegma regelii | 82.94 | 3.14 | 17.28 | moderate drought |
| 30 | 37.65 | 95.49 | 3248 | Grey brown desert soil | Ceratoides latens | 76.76 | 2.56 | 18.87 | moderate drought |
| 31 | 37.86 | 95.39 | 3308 | Chestnut soil | Sympegma regelii | 85.11 | 2.08 | 15.80 | moderate drought |
| 32 | 37.80 | 95.36 | 3188 | Grey brown desert soil | Ceratoides latens | 80.93 | 2.18 | 18.20 | moderate drought |
| 33 | 37.56 | 95.38 | 3184 | Inland salt soil | Ceratoides latens | 67.39 | 2.81 | 21.38 | severe drought |
| 34 | 37.33 | 95.51 | 3035 | Salt crust | Ephedra sinica | 58.4 | 3.45 | 26.44 | severe drought |
| 35 | 36.66 | 95.07 | 2701 | Inland salt soil | Folium Apocyni Veneti | 36.02 | 5.14 | 45.72 | severe drought |
| d.f. | SS | F | p | η2 (%) | |
|---|---|---|---|---|---|
| Stability, R2 = 0.47 | |||||
| LTN | 1 | 2.13 | 4.03 | 0.06 | 6.46 |
| LTP | 1 | 0.52 | 0.99 | 0.33 | 1.58 |
| LTK | 1 | 1.91 | 3.62 | 0.07 | 5.79 |
| LTC | 1 | 4.95 | 9.36 | <0.01 | 15.00 |
| STP | 1 | 0.00 | 0.01 | 0.94 | 0.01 |
| STK | 1 | 0.01 | 0.02 | 0.89 | 0.03 |
| STN | 1 | 6.25 | 11.82 | <0.01 | 18.92 |
| MAT | 1 | 2.47 | 4.67 | <0.05 | 7.48 |
| AP | 1 | 2.49 | 4.71 | <0.05 | 7.54 |
| AHN | 1 | 1.86 | 3.52 | 0.08 | 5.64 |
| Soil C:N | 1 | 0.01 | 0.01 | 0.92 | 0.02 |
| SWC | 1 | 0.49 | 0.93 | 0.35 | 1.49 |
| SS | 1 | 0.02 | 0.04 | 0.84 | 0.07 |
| MAP | 1 | 0.36 | 0.67 | 0.42 | 1.08 |
| pH | 1 | 0.03 | 0.05 | 0.83 | 0.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Chen, H.; Sun, H.; Yang, F. In the Qaidam Basin, Soil Nutrients Directly or Indirectly Affect Desert Ecosystem Stability under Drought Stress through Plant Nutrients. Plants 2024, 13, 1849. https://doi.org/10.3390/plants13131849
Zhao Y, Chen H, Sun H, Yang F. In the Qaidam Basin, Soil Nutrients Directly or Indirectly Affect Desert Ecosystem Stability under Drought Stress through Plant Nutrients. Plants. 2024; 13(13):1849. https://doi.org/10.3390/plants13131849
Chicago/Turabian StyleZhao, Yunhao, Hui Chen, Hongyan Sun, and Fan Yang. 2024. "In the Qaidam Basin, Soil Nutrients Directly or Indirectly Affect Desert Ecosystem Stability under Drought Stress through Plant Nutrients" Plants 13, no. 13: 1849. https://doi.org/10.3390/plants13131849
APA StyleZhao, Y., Chen, H., Sun, H., & Yang, F. (2024). In the Qaidam Basin, Soil Nutrients Directly or Indirectly Affect Desert Ecosystem Stability under Drought Stress through Plant Nutrients. Plants, 13(13), 1849. https://doi.org/10.3390/plants13131849