1. Introduction
Plants are integral components of terrestrial ecosystems [
1]. Global climate change (GCC) exerts nonlinear, dynamic, and multilevel effects on plant distribution, which in turn alter the structure and function of terrestrial ecosystems. Xinjiang is situated in the hinterland of the Asian–European continent and is a arid or semi-arid region with low precipitation, strong evaporation, relatively sparse vegetation, and fragile ecosystems, which makes it a region of high global ecological sensitivity [
2].
Trollius is a genus of flowering plants belonging to the
Ranunculaceae family. It grows mostly in mountains, and four species are produced in Xinjiang:
T. altaicus,
T. asiaticus,
T. dschungaricus, and
T. lilacinus. The mountain flower landscape formed by them has become a characteristic tourism project in Xinjiang. Recently, due to the sharp rise in emissions caused by human activities, which lead to climate anomalies,
Trollius wildflowers have become highly susceptible to extinction because of their high climatic sensitivity [
3], yet there is a paucity of knowledge regarding the distributional changes and conservation strategies of Trollius wildflowers in Xinjiang in response to GCC.
ITAP (Ili River Valley, Tacheng, Altay Prefecture) is situated in northwestern Xinjiang, with a total area of 325,117.01 km
2 and the highest biodiversity in Xinjiang [
4], making it an ideal location for studying ecosystem heterogeneity in buffering or accelerating the impacts of GCC owing to its climatic sensitivity. ITAP is also the main producing area for
Trollius wildflowers, and is comprised of three parts. The northern part (Altay) is characterized by a cooler climate and high habitat heterogeneity, with complex flora on Altay Mountain [
1,
5], and serves as a refuge for numerous endangered and endemic species. The central part (Tacheng) is situated in the west of the Junggar Basin, where the climate is drier in the plains and more humid in the mountains, resulting in the mountainous areas becoming isolated islands for the survival of species [
6]. The southern part (Ili River Valley) belongs to western Tien Shan, which is marked by a high degree of species richness and distinctive features, with significant vegetation differentiation, and it is a global biodiversity hotspot [
7,
8]. Climate change is anticipated to result in increased warming or precipitation in ITAP in the future, which may present challenges to biodiversity conservation.
Previous studies on
Trollius have mostly concentrated on its medical properties. The efficacy of
Trollius in clearing heat and detoxifying toxins [
9], as well as its antibacterial and anti-inflammatory effects [
10], were documented in the
Compendium of Materia Medica. More recent molecular chemistry research has demonstrated that
Trollius is rich in flavonoids and alkaloids [
11]. Additionally, its medical efficacy in antiviral and antioxidant properties has been confirmed in the context of pharmacology [
10,
12]. Due to their medical value, the resource of
Trollius species has been over-harvested anthropogenically and their populations have dwindled dramatically. In the context of plant physiology, scientists have investigated the response of
Trollius to various environmental stresses, including drought [
13], flooding [
14], and salinity [
15]. Additionally, they have examined the impact of train exhaust emissions and soil heavy metal pollution on its photosynthetic pigment content [
16]. Previous studies have paid little attention to the macro-level ecological characteristics and landscape features of
Trollius wildflowers, but environmental changes have already adversely affected and threatened the habitats of
Trollius wildflowers. Therefore, it is important to accurately assess the impact of GCC on suitable habitats and ecological niches of
Trollius wildflowers. This assessment is necessary to protect, develop, and utilize these resources in a rational manner.
The MaxEnt model has been employed with considerable success in the delineation of protected areas for endangered and relict species [
17,
18], the mapping of invasive species distributions [
19,
20], crop-designated geographical indications [
21,
22], and plant disease control [
23]. The objective of this study was to determine the intra- and inter-species impacts of GCC on the four species of
Trollius in ITAP. To achieve this, we developed separate models for each species based on different distribution points and environmental variables. As herbaceous flowers,
Trollius have a much lower dispersal capacity than animals and lower immunity to GCC than woody plants. Thus, we attempted to explore the magnitude of suitable habitat changes between species under the premise of overlapping survival zones to determine the tolerance level of species to GCC. In addition, we explored the critical role of the transition zone between the more heterogeneous mountainous areas and the more homogeneous plains in the survival of species. Furthermore, different climate scenarios present distinct pathways for the future protection of species. In this study, we identified the effects of the new scenario SSP3-7.0 on the distribution of
Trollius wildflowers, which emphasizes land-use change for sustainable development and reduced precipitation [
24]. Finally, we attempted to determine the evolutionary potential and dispersal movements of species in the face of GCC impacts through changes in species’ ecological niche breadth and overlap indexes.
This study employed the R-optimized MaxEnt model to analyze the impacts of the last glacial period, current conditions, and future scenarios on suitable habitats and ecological niche dynamics for Trollius wildflowers. The study aims to achieve the following scientific objectives: (1) to obtain the potential distribution range of Trollius under a baseline climate, the most suitable distribution area, and the key factors affecting its distribution; (2) to clarify the impact of GCC on the suitable distribution range and spatial patterns of Trollius; and (3) to determine the degree of ecological differentiation of Trollius under climate change. The study aims to scientifically delineate the scope of development and protection of Trollius wildflower resources and realize the high-quality development of territorial resources.
2. Results
2.1. Model Accuracy and Dominant Environmental Factors
The results showed that the AUC values were 0.908 for T. altaicus, 0.939 for T. asiaticus, 0.926 for T. dschungaricus, and 0.954 for T. lilacinus, indicating that the model prediction accuracy is very high and can be used to simulate the periods of suitable Trollius wildflower habitat distribution.
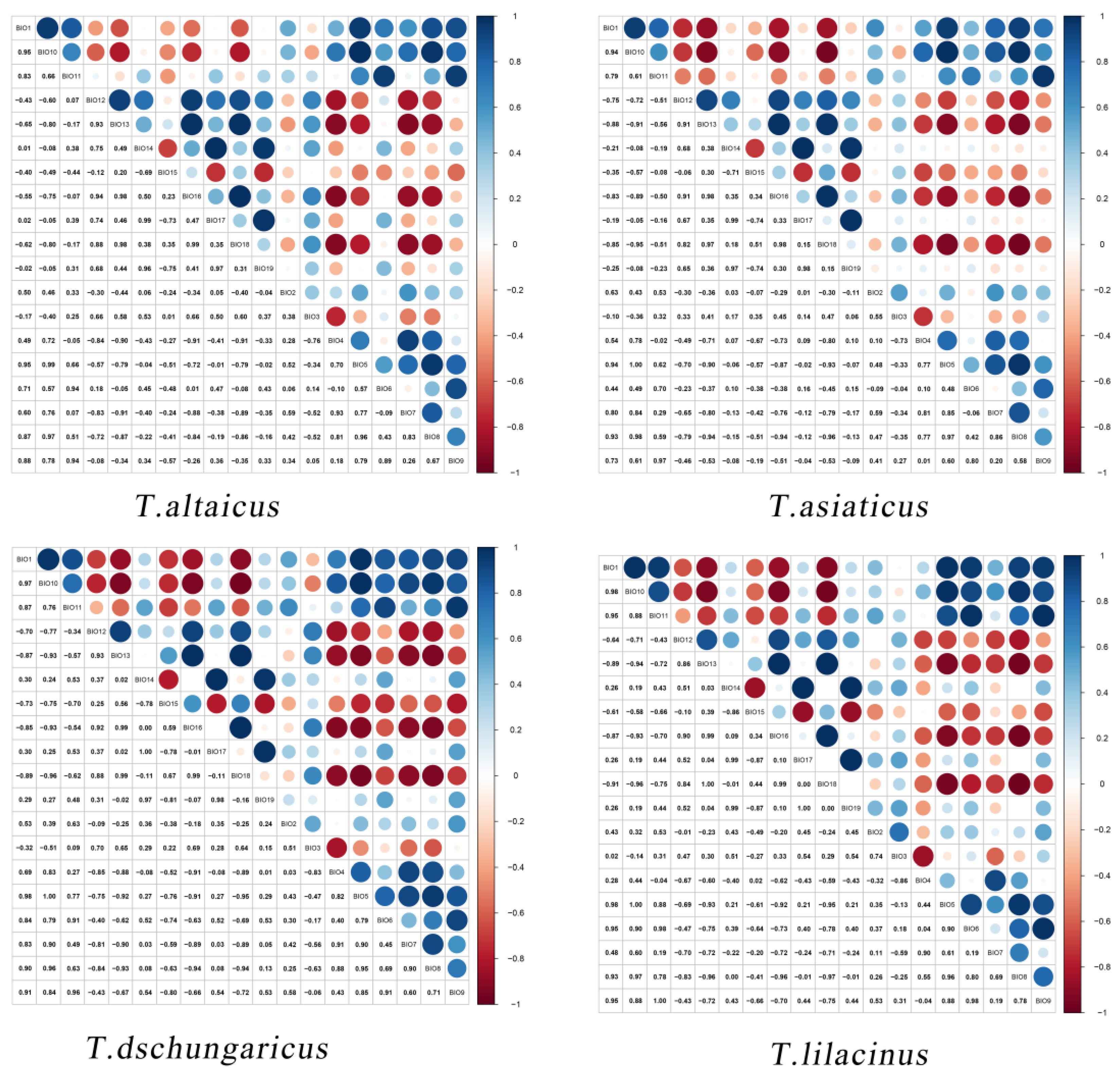
We accumulated the environmental variables with a sum of contributions greater than 80%, and the results indicated that the climatic factors affecting
Trollius were primarily Bio18, Bio3, Bio4, Bio1, Bio19, and Bio15 (
Table 1). First, the largest contributor was Bio18, which contributed 43.2% to
T. altaicus and 19.0% to
T. lilacinus. This was followed by Bio3 and Bio4, with Bio3 mainly affecting
T. aisaticus and Bio4 affecting the distribution of all four species, with a contribution rate ranging from 4.8% to 14.0%. These results indicated that the four species are more sensitive to seasonal changes in temperature. Furthermore, the NDVI influenced the distribution of all four species, with contribution rates ranging from 4.2% to 24.4%. This was particularly evident in
T. altaicus and
T. dschungaricus.
T. lilacinus and
T. dschungaricus exhibited a strong sensitivity to soil pH, with both species thriving in humus-rich, slightly acidic sandy loam. However, excessive acidity or alkalinity can lead to plant death.
Species distribution thresholds were selected based on distribution probabilities exceeding 50%, then the means were then calculated. T. altaicus exhibited a lower Bio18 requirement with a mean of 125.76 mm, while T. lilacinus demonstrated a higher requirement for the Bio18 factor with a higher upper threshold limit of 281.20 mm. The difference in the means of the four Trollius species for the Bio4 requirement was relatively minor, ranging from 937.17 to 1139.31, with the upper threshold of T. altaicus being higher. Furthermore, T. asiaticus was more sensitive to the precipitation of the driest month and quarter (Bio14 and Bio17).
2.2. Current Distribution of Suitable Habitat for Trollius Wildflowers
The results of the simulation indicated that the total area of suitable habitats for
Trollius wildflowers in ITAP exhibited considerable variation (
Table 2).
T. altaicus displayed the largest total area of suitable habitats, accounting for 40.68%, while
T. lilacinus showed the smallest total area of suitable habitats, accounting for 20.46%. In the highly suitable habitats,
T. asiaticus accounted for the largest percentage (8.25%), with a concentration in western Tien Shan in Ili River Valley, the Alatau Mountain in Bole, and the southwestern foothills of the Baluk Mountains in Tacheng (
Figure 1).
T. altaicus accounted for the second-largest percentage (7.61%), with a distribution concentrated in the southern and middle branches of Tien Shan, as well as in the Xialshili and Sayram Lake areas. The Talbahadai Mountains, the Urgarsar Mountains, the Saighur Mountains, the Baluk Mountains, and the northern Altay Mountains, all of which were highly suitable habitats for
T. altaicus, moderately suitable habitats, and lowly suitable habitats, were identified on the edges of the patches of highly suitable habitats.
The smallest proportion of highly suitable habitat was observed in T. dschungaricus (5.12%), which was mainly distributed in western Tien Shan in Ili. Additionally, sporadic distributions of highly suitable habitats were observed in northwest Altay Prefecture (Baihaba scenic area). However, no distribution of moderate or highly suitable habitats for T. dschungaricus was observed in Tacheng. In contrast, the highly suitable habitat of T. lilacinus was exclusively distributed in western Tien Shan in the Ili River Valley, while only lowly suitable habitats were distributed in Tacheng and Altay Prefecture.
2.3. Impacts of GCC on Area Alteration in Suitable Habitats for Trollius Wildflowers
The alterations in the suitable habitats of the
Trollius wildflowers exhibited a notable discrepancy from the past to future periods, largely due to the climatic sensitivity of mountainous regions and the habitat preferences of species. Specifically, the totally suitable habitat percentage of the four
Trollius species varied considerably, ranging from 0.14% to 70.97% (
Figure 2a). The smallest change in totally suitable habitat was observed in
T. altaicus, ranging from 25.27% to 40.68%. In contrast, the largest variation was observed in
T. lilacinus, ranging from 8.16% to 52.42%. Western Tien Shan and the Altay Mountains were identified as the most comprehensive refuges for
Trollius wildflowers, while the distribution in other regions varied according to climate conditions (
Figure 2b).
The Last Glacial Maximum (LGM) was a period of transition from a cold to a warm Earth, with average temperatures 5 to 10 °C lower than the current era. However, the average temperature in the Middle Holocene (MID) was 3–5 °C higher. From the LGM to the current era, the areas of highly suitable habitats for the four Trollius species showed an upward trend, while the areas of moderate and lowly suitable habitats were relatively fluctuating. It was evident that the totally suitable habitats of T. altaicus and T. lilacinus exhibited a decline and subsequent increase. Both species exhibited low values in the MID, indicating that they are more sensitive to warming. The totally suitable habitat of T. asiaticus increased and then decreased, while T. dschungaricus exhibited a nearly absent distribution of suitable habitat during the glacial era.
The greatest proportion of total and highly suitable habitats was exhibited by T. altaicus in the SSP1-2.6 scenario, while the lowest proportion was observed in the SSP3-7.0 scenario. In the 2050s, T. altaicus exhibits a declining trend in totally suitable habitat, as evidenced by a reduction in highly and moderately suitable habitats and fluctuations in lowly suitable habitats. The percentage of totally suitable habitats under the three scenarios for the 2070s remains relatively consistent, with the SSP1-2.6 scenario still exhibiting the highest percentage of totally and highly suitable habitats (37.85% and 1.93%). The percentage of highly suitable habitats under the three scenarios falls below 2% in the 2070s, representing a significant decline from the current era (7.61%). However, by the 2090s, the percentage of highly suitable habitats rebounds to more than 4%, with the percentage of highly suitable habitats under the SSP1-2.6 scenario rising to 6.33%.
For T. asiaticus, it is predicted that the area of both moderately and highly suitable habitats will increase. In the 2050s, the area of moderately suitable habitats increases to over 14% in three scenarios, exceeding the current level. In contrast, the proportion of lowly suitable habitats decreases to 13.99% under the SSP3-7.0 scenario, while the proportion of highly suitable habitats increased to 25.46%. In the 2070s, there is less difference between the classes of suitable habitats. However, the percentage of highly suitable habitats exceeds 35%. Totally suitable habitats exceed 70%, making this the most widely distributed period for T. asiaticus. In the 2090s, totally suitable habitats for T. asiaticus decrease to 48.27% under the SSP1-2.6 scenario, yet remain higher than the 37.83% observed at present.
For T. dschungaricus, the proportion of totally suitable habitats exhibits a decline in the 2050s and 2070s, followed by a recovery in the 2090s. In the 2050s, the habitat conditions of T. dschungaricus demonstrate a high degree of maladaptation to the SSP3-7.0 scenario, with only 3.45% of the totally suitable habitats under the scenario and a distribution confined to the Ili River Valley. In the 2070s, the proportion of highly suitable habitats under all three scenarios drops below 1%, with the species confined to western Tien Shan and the Altay Mountains. By the 2090s, however, the proportion of highly suitable habitats rebounds to more than 23%, with the SSP1-2.6 scenario displaying the greatest total area of suitable habitats, yet the smallest proportion of highly suitable habitats and lower than the proportions of highly suitable habitats under the SSP3-7.0 and SSP5-8.5 scenarios.
The suitable habitats for T. lilacinus will undergo a decline in the future. By the 2070s, the proportion of highly suitable habitats declines to approximately 3%. The highest proportion of highly suitable habitats is observed in SSP5-8.5, but the lowest proportion of totally suitable habitats is also evident. In the 2090s, the greatest proportion of totally suitable habitats and all classes of suitable habitat are exhibited under the SSP1-2.6 scenario, with a rebound from the 2070s SSP1-2.6 scenario. Nevertheless, the proportion of all classes of suitable habitat in the SSP3-7.0 and SSP5-8.5 scenarios continue to decline in comparison to the 2070s.
2.4. Impacts of GCC on Spatial Transfer of Suitable Habitats for Trollius Wildflowers
The result of the spatial transfer of suitable habitats indicates that the edges of habitat patches are susceptible to GCC. They are also the main areas where habitat is added or lost (
Figure 3b). Habitat additions and losses varied among species.
T. dschungaricus and
T. lilacinus had the lowest average habitat additions, while
T. asiaticus had the most habitat additions. Habitat changes in
T. altaicus were relatively fluctuating.
For
T. altaicus, the extent of its habitat has diminished by 12.50% during the LGM–MID period. This indicates that warmer temperatures have a significant impact on alpine flowers. During the MID to the current period, the extent of suitable habitats increased by 21.07%, with the majority of this recovery occurring in mountainous regions. By the 2050s, the spatial changes in suitable habitats are not apparent in the SSP1-2.6 or SSP5-8.5 scenarios. In contrast, in the SSP3-7.0 scenario, there is a dramatic shift in habitats. The habitats lost in the 2070s represent approximately 17% to 18% of the total, with the majority of these losses occurring in the higher-elevation areas of the mountains. The 2090s are characterized by a dominance of stable habitats, with a mere 1% of increased habitats and a loss of habitats amounting to 3% to 5% (
Figure 3a).
For T. asiaticus, habitat changes were dominated by loss. The increased habitats during the LGM–MID period were primarily situated on the edges of mountainous areas, where the warmth and light were more conducive to harvesting. In the MID–current period, a significant expansion in habitats occurred in the Ili River Valley, with a subsequent contraction in Altay. From the current period to the 2050s, the proportion of lost habitats ranges from 0.01% to 1.21%. Conversely, there was a notable increase in the number of added habitats, which occurred mainly in the Altay Mountains and northern Tien Shan. By the 2070s, the proportion of lost habitats ranges from 0.01% to 0.08%, while the increased habitats are distributed across a range of areas, with the largest patches located in Altay. By the 2090s, the proportion of lost habitats ranges from 0 to 0.32%, with the greatest increase occurring under the SSP5-8.5 scenario.
For T. dschungaricus, habitat changes are dominated by additions. The percentage of non-habitat during the LGM–MID period was 99.26%, while the percentage of increased habitats in the MID–current period was 27.32%, which was concentrated in mountainous areas. During the current–2050s period, the largest percentage of lost habitats (24.42%) and the lowest percentage of increased habitats (0.02%) are observed under SSP3-7.0. By the 2070s, the number of lost habitats continues to rise, with the majority of these habitats located in the low-elevation areas of the Ili River Valley. By the 2090s, the greatest increased habitats are observed in SSP5-8.5. However, this scenario still had the highest number of lost habitats, in contrast to the SSP1-2.6 scenario, which was more stable.
For T. lilacinus, suitable habitats declined by 16.63% during the LGM–MID period and increased by 85.47% during the MID–current period. From the current to the 2050s, the increased habitats under the SSP3-7.0 rise by 37.46%, with the majority concentrated in the Ili River Valley and the Junggar Basin. The remaining two scenarios exhibited a similarly low percentage of increased habitats, with less than 1%. By the 2070s, the increased habitats are all 0, while the lost habitats constitute approximately 8%. The lost habitats are primarily located on the edge of western Tien Shan. By the 2090s, the SSP1-2.6 scenario exhibits the greatest increase in habitats, at 0.58%. This is primarily observed in the Alatau Mountains, the Xialshili area, the Saighur Mountains, and the Altay Mountains. In contrast, the SSP5-8.5 scenario exhibits the fewest new additions and the most lost habitats.
2.5. Impacts of GCC on Gravity Center of Suitable Habitats for Trollius Wildflowers
For
T. altaicus, overall, the gravity center shifted to low latitudes (
Figure 4). The direction of the gravity center of suitable habitats exhibited a notable shift, with a predominant northeast–southwest trajectory. During the LGM–MID period, the suitable habitats in the southern Tacheng experienced a substantial reduction, prompting a northward shift in the gravity center of
T. altaicus by 39.47 km. However, in the MID–current period, the suitable habitats in southern Tacheng demonstrated a notable recuperation, resulting in a return of the gravity center to its original position. In the SSP1-2.6 and SSP5-8.5 scenarios, shorter migration distances and less spatial variability of the gravity center were observed. In the SSP3-7.0, the gravity center exhibited a significant shift of 96.64 km to high latitudes to avoid the intense heat. However, by the 2070s, the gravity center shifts southwest by 73.08 km, and by the 2090s, it shifts further, reaching 63.17 km to the southwest
(Table S3).
The gravity center of T. asiaticus shifted from the southern to the northern part of Karamay during the LGM–MID–current period. In the SSP1-2.6 scenario, the expansion of habitats was primarily observed in the Sagur Mountains and the Altay Mountains. Consequently, the gravity center shifted to higher latitudes by 124.25 km in the 2050s. In the 2070s and 2090s, the gravity center exhibits a meandering shift from southwest to northeast. In the SSP3-7.0 scenario, the gravity center undergoes a southwest–northeast–southwest shift with a shorter distance. In contrast, in the SSP5-8.5 scenario, the gravity center undergoes a sharp shift to the northwest by 125.51 km in the 2050s, followed by a return to the current gravity center after two migrations from the 2070s to the 2090s.
The migration of the gravity center of suitable habitats for T. dschungaricus exhibited fluctuations and a meandering pattern. During the LGM–MID–current period, the gravity center initially migrated from Tacheng to Ili River Valley and subsequently reverted to eastern Tacheng. In the SSP1-2.6 scenario, the gravity center initially shifts 220.14 km to the southwest, followed by a 124.60 km shift back to the northeast in the 2090s. In the SSP3-7.0 scenario, the increased suitable habitats occur exclusively in the Ili River Valley during the 2050s, and consequently, the gravity center shifts abruptly 310.67 km to the Ili River Valley. There is a restoration of suitable habitats in the mountainous regions in the eastern part of the Ili River Valley during the 2070s, resulting in a further shift in the gravity center by 310.67 km eastward. In the SSP5-8.5 scenario, the gravity center migrates 67.52 km southwest in the 2050s, and the extent of degradation of suitable habitats in Altay and Tacheng is considerable in the 2070s, with the gravity center shifting southward to lower latitudes. In the 2090s, suitable habitats in Altay and Tacheng experience a recovery, resulting in the gravity center returning to a position near its original location.
For T. lilacinus, during the LGM–MID–current period, the gravity center of suitable habitats for T. lilacinus shifted southwest from Karamay to Bole by a distance of 180.76 km, and then moved northwest to the central part of Tacheng. In the future SSP1-2.6 scenario, the gravity center moves 65.66 km southwest in the 2050s and continues to move southwest to Ili River Valley in the 2070s by a distance of 202.99 km. In the 2090s, it moves back to the area where the 2050s center is located. In the SSP3-7.0 scenario, the gravity center shifts 117.64 km to the northeast in the 2050s, migrates 373.55 km to the southwest of Ili River Valley in the 2070s, and travels 214.13 km back to the northwest in the 2090s. In the SSP5-8.5 scenario, the gravity center is initially displaced slightly to the southwest by 64.61 km in the 2050s, subsequently continuing to migrate southward to Ili River Valley in the 2070s, and finally moving to the south of Bole in the 2090s.
2.6. Impacts of GCC on Ecological Niche Breadth, Overlap, and Equivalency for Trollius Wildflowers
The maximum threshold of niche breadth for
T. altaicus and
T. asiaticus was consistently above 0.9 in each period, while the minimum threshold exhibited fluctuations (
Figure 5a). The minimum threshold of niche breadth for
T. altaicus exhibited a “falling–rising–falling” pattern, with a minimum value of 0.24 reached in the 2050s under the SSP3-7.0 scenario. Conversely, the minimum threshold of niche breadth for
T. asiaticus exhibited a continuous increase, reaching a peak in the 2070s, followed by a decrease in the 2090s, and finally reaching a minimum value of 0.48 in the 2090s under the SSP1-2.6 scenario. The maximum and minimum threshold changes in niche breadth of
T. dschungaricus both exhibited a similar wave-like pattern, with peaks of 0.92 and 0.26 observed in the present and troughs of 0.55 and 0.00 observed during the LGM, respectively. The niche breadth of
T. lilacinus exhibited greater stability, although it displayed a pronounced peak in the 2050s under the SSP3-7.0 scenario, due to the rapid expansion in suitable habitats for
T. lilacinus in this scenario.
The results of principal component analysis (PCA) indicated that PCA (PC1 and PC2) based on screened climatic variables explained 84.63% of the climatic variance at the distribution site of
T. altaicus, 81.50% of
T. asiaticus, 81.70% of
T. dschungaricus, and 86.00% of
T. lilacinus. In particular, temperature and precipitation were found to influence the geographic distribution and ecological niche variation of
Trollius wildflowers (
Figure S2).
The results of the climatic ecological niche overlap dynamics and equivalency test, based on species distribution data and climatic variables, revealed that the degree of climatic ecological niche differentiation of the four
Trollius species varied greatly (
Figure 5b). For
T. altaicus, comparing past with present niches, there was a differentiation characterized by deletion. In the future, except for the SSP3-7.0 scenario in the 2050s and all scenarios in the 2070s, the overlap indexes for rest periods and scenarios were above 0.9 and essentially undifferentiated [
25]. With regard to
T. asiaticus, the differentiation of ecological niches in the future was mainly dominated by expansion, which was attributed to the substantial growth in suitable habitats. In contrast,
T. dschungaricus was distributed scarcely in the past glacial period, and thus there was a large deletion in the ecological niche compared to the current. Under the SSP3-7.0 scenario in 2050s, the suitable habitat of
T. dschungaricus retreated to the Ili River Valley, resulting in a severe ecological niche deletion. However, the overlap index for the remainder of the period was approximately 0.7–0.8, indicating a slight degree of differentiation. The type of ecological niche differentiation observed in
T. lilacinus was characterized by deletions, which were related to its spatial distribution and its capacity to hold ecological resources at different times.
4. Discussion
Trollius wildflowers, with their beautiful color and appearance, are an important regional landscape resource in Xinjiang. The mountain flower communities formed by
Trollius and their companion species are also important engines for maintaining the health of grassland and understory ecosystems in Xinjiang [
35]. This study predicted the impacts of GCC on the evolution of suitable habitats and ecological niche changes for
Trollius wildflowers in ITAP from the past to the future. The results indicated that precipitation and temperature are the dominant environmental variables affecting the distribution of
Trollius. For
T. altaicus and
T. lilacinus, the precipitation factor was a deeper influence, potentially due to their adaptations to extreme temperatures in Xinjiang. These adaptations include entering dormancy or seeking shelter under extremely hot or cold conditions, which would allow them to survive despite these harsh conditions [
36]. For instance,
T. altaicus exhibits a capacity to dethatch its leaves and reduce its height, retaining only the number of leaves necessary for photosynthesis to survive the summer. In contrast, during the winter months, it enters a dormant state and withers, preparing for germination in the following spring.
T. lilacinus displays greater tolerance to cold weather due to its prevalence at high altitudes. Nevertheless, both
T. altaicus and
T. lilacinus are highly susceptible to precipitation during the warmest season. In 2020, a field study was conducted in Xinjiang, where the climate was found to be abnormal, with increased temperature and decreased precipitation. This resulted in
T. altaicus plants in Tacheng Bayimuzha and Urumqi South Tien Shan having low levels of growth, with smaller flower diameters and overall sub-healthy conditions compared to the previous wet year (
Figure 9). This suggests that during the peak growing season of the plant, water and heat conditions are indispensable [
37]. Previous large-scale species distribution prediction studies have not considered soil factors. However, soil physicochemical properties play a crucial role in species distribution. Simulation studies have shown that soil texture and pH are also among the environmental variables influencing the distribution of
Trollius, particularly for
T. dschungaricus and
T. lilacinus. According to the research,
T. dschungaricus is predominantly found in the understory of temperate mixed coniferous and broad forests, while
T. lilacinus is found in alpine meadows, both of which are characterized by slightly acidic humus soils. Additionally, elevation and vegetation elements are also significant environmental variables influencing the distribution of
Trollius wildflowers.
With greenhouse gas emissions, some species will migrate to higher latitudes or elevations, while others will maintain their current location through physiological or phenological changes [
38]. The future climate scenario indicated that the changes in suitable habitat for
Trollius wildflowers in the 2050s, 2070s, and 2090s exhibits uneven fluctuations. In general, the SSP1-2.6 scenario demonstrated a higher level of habitat maintenance. It was demonstrated that a certain degree of increased CO
2 emissions could promote the growth of terrestrial plants and enhance photosynthetic efficiency [
39]. This suggests that a moderate increase in CO
2 concentration is favorable for the population expansion of
Trollius, but a scenario of emissions that are too high may be detrimental to the survival of herbaceous plants because of their weaker carbon sequestration capacity than that of woody plants [
40]. The establishment of the SSP3-7.0 scenario was associated with a decrease in future precipitation and an increase in temperature [
24], yet the proportion of suitable habitat area of
T. asiaticus remained consistently larger, which indicates that
T. asiaticus is more susceptible to temperature changes. This result is consistent with a previous report by Li [
41], which reveals an evolutionary bias in the response to environmental changes between the different species in the same genus. Furthermore, the transition zones between mountains and plains, such as hilly areas, are often the origin of increased habitats, and habitats are also the first to disappear in the transition zone. The edge theory indicates that the edge of an ecosystem often exhibits high biodiversity, yet it is inherently unstable and susceptible to external disturbances, particularly in recent times [
42]. In the future, all four species of
Trollius will have some degree of increased habitats in the Altay Mountains, which may be related to the increases in precipitation in the Altay Mountains in the future. Concerning Tacheng, the more arid climate results in plant communities being more sensitive to external random disturbances, which will lead to the disappearance of more habitats in the mountainous areas in Tacheng. Ili River Valley is characterized by a relatively humid climate and fertile soil and water resources, which provide a refuge for
Trollius to cope with a range of extreme climatic conditions. It also serves as a concentrated distribution point for
Trollius, but in the future, it may also face the problem of habitat degradation at the edge of the mountainous area, which needs to be prevented.
The climatic ecological niche is the ecological magnitude occupied by species in a multifactorial climate space [
34]. The differentiation of climatic ecological niches not only represents the change in species distribution but also records the alteration in species’ dependence and occupancy on climatic elements. This is essentially the evolution of species due to genetic changes as a result of natural selection [
43]. The ecological niches of
T. asiaticus were predominantly expansive, occupying the potentially available space, indicating the spatial migration and resource-holding capacity of
T. asiaticus when facing climate change, which can be also illustrated by the widest geographic distribution in China. However, considering the conservation of resource allocation, the expansion of
T. asiaticus has the potential to affect other species in the community. The ecological niches of
T. lilacinus were predominantly degraded, which revealed the vulnerability of alpine meadow ecosystems near the snow line. An overlap index greater than 0.9 can be considered to represent no differentiation. The results of this study indicate that the overlap indexes of
T. altaicus are greater than 0.9 in some scenarios, suggesting a greater probability of stability. In contrast, the overlap indexes of the remaining three species range from 0.6 to 0.9, indicating a relatively large amount of potentially usable environmental space.
In addition to climate warming, the development of tourism, livestock, and illegal harvesting have also significantly reduced the habitat of Trollius. In recent years, the Trollius flower sea has become a popular tourist destination (the established species are mainly T. altaicus and T. asiaticus); however, the construction of scenic facilities and the influx of tourists have harmed the local populations of Trollius. Furthermore, the habitats of Trollius overlap with grazing land in Xinjiang due to a lack of targeted protection mechanisms, which results in the populations of Trollius being seriously trampled and gnawed by herds. The medicinal and food value of Trollius has made illegal harvesting a major factor in the sharp decline in Trollius populations. For example, T. lilacinus, growing near the snow line, has been subjected to severe poaching for its tea properties. As a result, T. lilacinus has been classified as a vulnerable (VU) species in China. Based on the simulation results of the existing and potential habitats of Trollius, it is strongly recommended that the wild resources of Trollius be protected holistically, taking into account the Xinjiang Territorial Spatial Planning Document (2021–2035) and the results of the Third Xinjiang Scientific Expedition. Firstly, it is essential to scientifically divide the source areas of Trollius habitat by combining the scope of existing nature reserves and highly suitable habitat distribution and establishing ecological corridors between the source areas for migration, for example, making lowly and moderately suitable habitats become stepping stones and connections for the migration process of Trollius. Secondly, building ecological barriers such as Tien Shan, Altay Mountains, etc., to construct a biodiversity conservation network, to strengthen the protection of natural forests and meadow in the native range of Trollius, to scientifically demarcate the ecological red line, and to limit the harm of tourism, livestock increases, and the expansion of construction on the Trollius population. Finally, it is necessary to implement protection regulations, strictly combat illegal poaching, and carry out artificial cultivation of Trollius.
In conclusion, this study systematically explored the impacts of environmental variables on Trollius wildflowers and changes in suitable habitats and ecological niches and made relevant recommendations for the future conservation of Trollius. However, additional studies still need to be enhanced. In the future, field research should be strengthened to obtain real-time distribution data and local climatic data. Additionally, concerning the selection of environmental factors, it would be beneficial to consider the inclusion of human activities and land-cover change factors in future studies. Furthermore, the core patches and potential corridors of suitable habitats for the species could be integrated with the nature reserves, thus forming a systematic habitat network that would be more conducive to the conservation of the rare species.
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}