
Identification and Validation of Quantitative Trait Loci Associated with Fruit Puffiness in a Processing Tomato Population


Abstract
1. Introduction
2. Materials and Methods
2.1. Mapping Populations
2.2. Field Trials
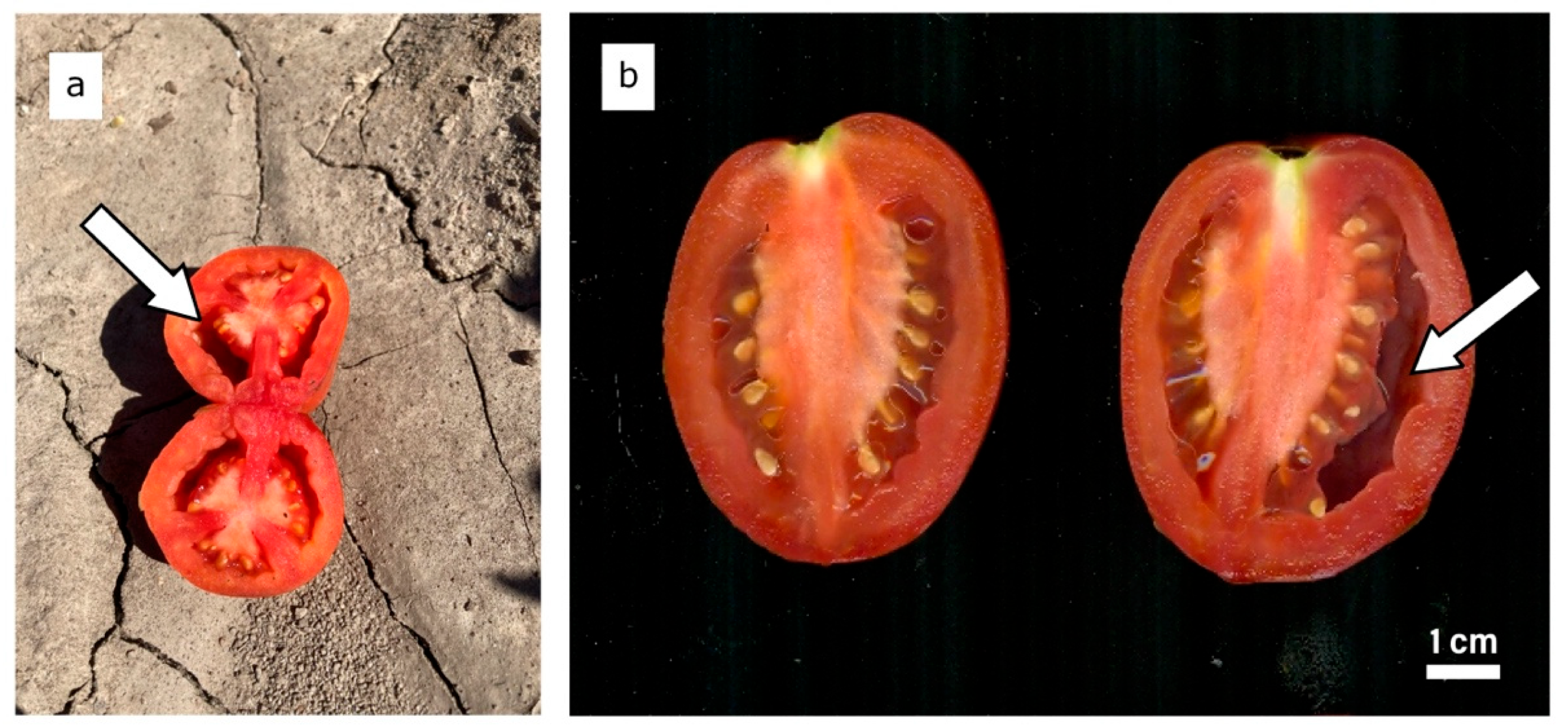
2.3. Fruit Puffiness Measurements
2.4. Phenotypic Data Analysis
2.5. DNA Isolation and Genotyping
2.6. Genetic Linkage Map Construction
2.7. QTL Mapping
2.8. Interaction between QTLs
2.9. QTL Validation and QTL Action
2.10. Comparison of Selection Strategies for Decreased Fruit Puffiness
3. Results
3.1. Fruit Puffiness Assessment
3.2. Linkage Map Quality
3.3. QTL Identification
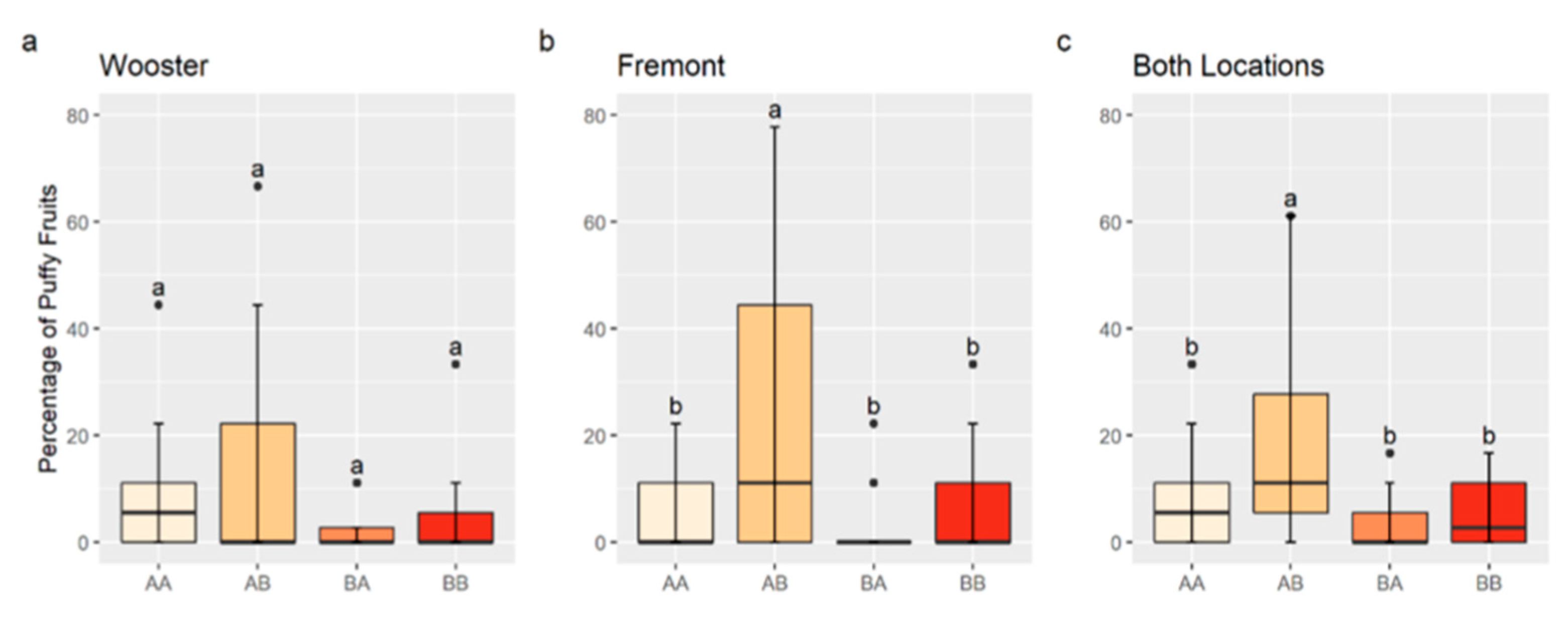
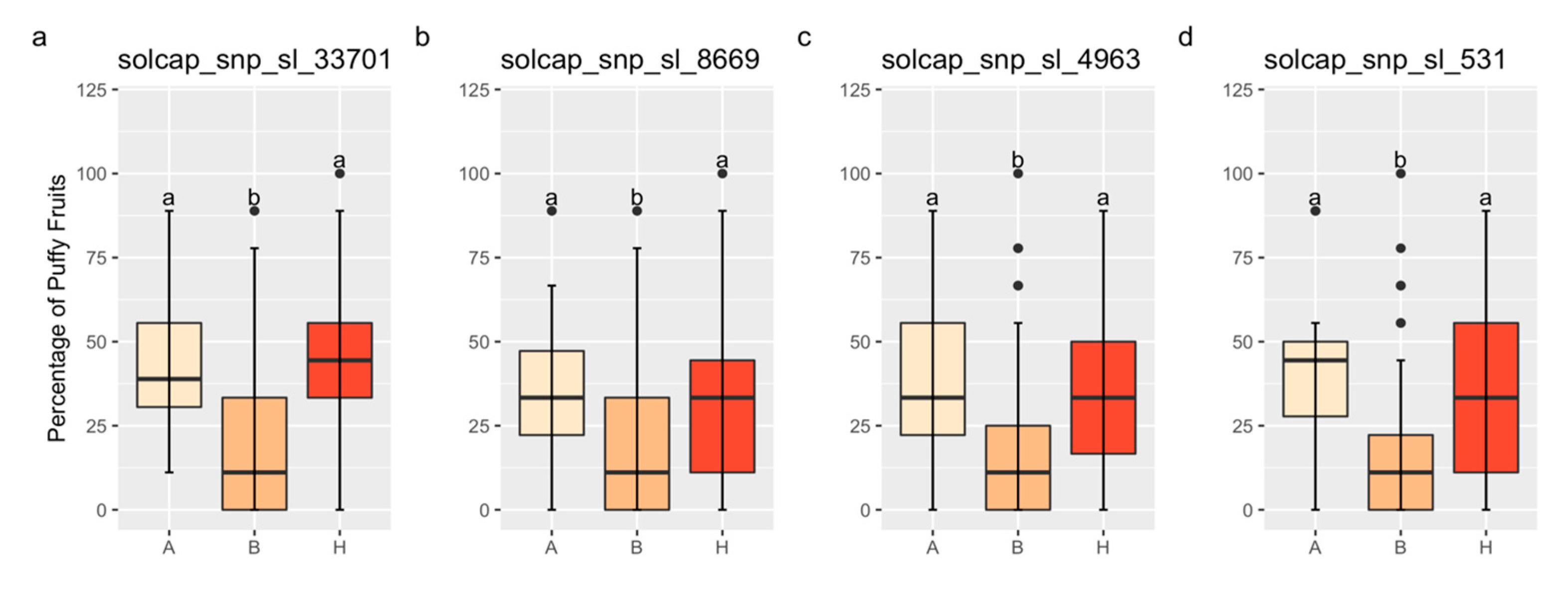
3.4. QTL Validation and Interaction
3.5. QTL Action
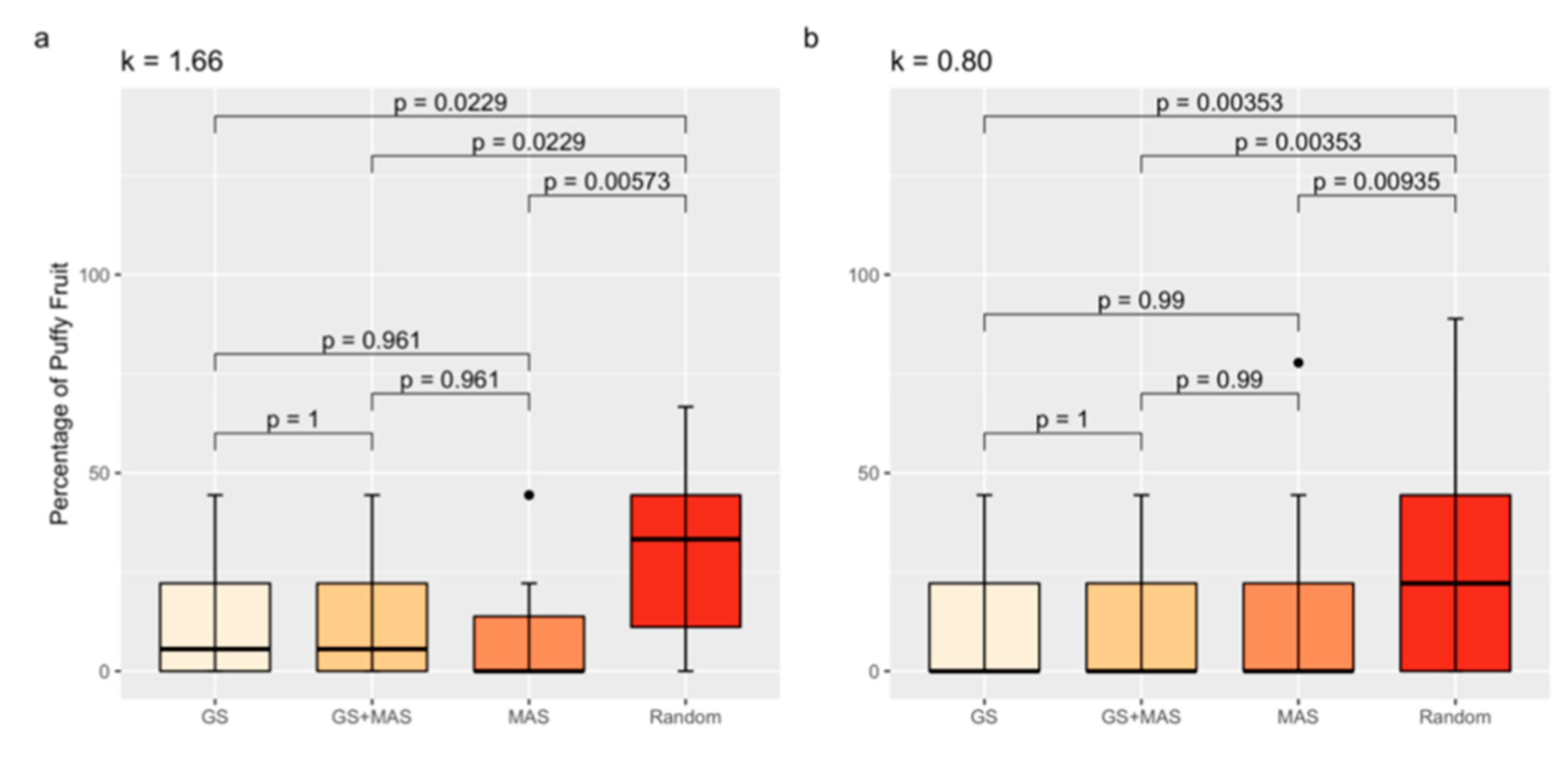
3.6. Marker-Assisted and Genomic Selection
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Masarirambi, M.; Mhazo, N.; Oseni, T.; Shongwe, V. Common Physiological Disorders of Tomato (Lycopersicon esculentum) Fruit Found in Swaziland. Agric. Soc. Sci. 2009, 5, 123–127. [Google Scholar]
- Peet, M.M. Physiological Disorders in Tomato Fruit Development. Acta Hortic. 2009, 821, 151–160. [Google Scholar] [CrossRef]
- Gatahi, D.M. Challenges and Opportunities in Tomato Production Chain and Sustainable Standards Introduction. Int. J. Hortic. Sci. Technol. 2020, 7, 235–262. [Google Scholar] [CrossRef]
- Gangadhara, K.; Selvakumar, R.; Singh, P.K.; Manjunathagowda, D.C.; Kumar, J. Tomato: Physiological Disorders and Their Management. Biot. Res. Today 2021, 3, 944–948. [Google Scholar]
- USDA. Tomatoes. Shipping Point and Market Inspection Instructions; United States Department of Agriculture: Washington, DC, USA, 2005. [Google Scholar]
- Doganlar, S.; Frary, A.; Ku, H.M.; Tanksley, S.D. Mapping Quantitative Trait Loci in Inbred Backcross Lines of Lycopersicon pimpinellifolium (LA1589). Genome 2002, 45, 1189–1202. [Google Scholar] [CrossRef] [PubMed]
- Nawata, E.; Inden, H.; Asahira, T. Effects of ccc on the Occurrence of Tomato Puffy Fruits and the Endogenous Cytokinin Activities. Sci. Hortic. 1985, 26, 119–127. [Google Scholar] [CrossRef]
- Olson, S.M. Physiological, Nutritional, and Other Disorders of Tomato Fruit; HS-954; University of Florida: Gainesville, FL, USA, 2004. [Google Scholar]
- Shabtai, S.; Salts, Y.; Kaluzky, G.; Barg, R. Improved Yielding and Reduced Puffiness under Extreme Temperatures Induced by Fruit-Specific Expression of RolB in Processing Tomatoes. Theor. Appl. Genet. 2007, 114, 1203–1209. [Google Scholar] [CrossRef] [PubMed]
- Capel, C.; Yuste-Lisbona, F.J.; López-Casado, G.; Angosto, T.; Cuartero, J.; Lozano, R.; Capel, J. Multi-Environment QTL Mapping Reveals Genetic Architecture of Fruit Cracking in a Tomato RIL Solanum lycopersicum × S. pimpinellifolium Population. Theor. Appl. Genet. 2017, 130, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Topcu, Y.; Sapkota, M.; Illa-Berenguer, E.; Nambeesan, S.U.; van der Knaap, E. Identification of Blossom-End Rot Loci Using Joint QTL-Seq and Linkage-Based QTL Mapping in Tomato. Theor. Appl. Genet. 2021, 134, 2931–2945. [Google Scholar] [CrossRef]
- Asíns, M.J. Present and Future of Quantitative Trait Locus Analysis in Plant Breeding. Plant Breed. 2002, 121, 281–291. [Google Scholar] [CrossRef]
- Pessoa, H.P.; Da Silva, L.J.; Chagas, R.R.; Ferreira Copati, M.G.; Dariva, F.D.; de Oliveira Dias, F.; Nick Gomes, C. Uncovering Tomato Candidate Genes for Seed Germination Performance and Seedling Vigour Using the Solanum pennellii Introgression Line Population. Sci. Hortic. 2021, 284, 110134. [Google Scholar] [CrossRef]
- Robbins, M.D.; Masud, M.A.; Panthee, D.R.; Gardner, R.G.; Francis, D.M.; Stevens, M.R. Marker-Assisted Selection for Coupling Phase Resistance to Tomato Spotted Wilt Virus and Phytophthora infestans (Late Blight) in Tomato. HortScience 2010, 45, 1424–1428. [Google Scholar] [CrossRef]
- Bernal, E.; Francis, D.M. Processing Tomato Germplasm with Improved Resistance to Bacterial Spot. HortScience 2021, 56, 519–520. [Google Scholar] [CrossRef]
- Rodríguez, G.R.; Moyseenko, J.B.; Robbins, M.D.; Morejón, N.H.; Francis, D.M.; van der Knaap, E. Tomato Analyzer: A Useful Software Application to Collect Accurate and Detailed Morphological and Colorimetric Data from Two-Dimensional Objects. J. Vis. Exp. 2010, 37, e1856. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, G.R.; Muños, S.; Anderson, C.; Sim, S.C.; Michel, A.; Causse, M.; McSpadden Gardener, B.B.; Francis, D.; van der Knaap, E. Distribution of SUN, OVATE, LC, and FAS in the Tomato Germplasm and the Relationship to Fruit Shape Diversity. Plant Physiol. 2011, 156, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Sim, S.C.; Robbins, M.D.; Wijeratne, S.; Wang, H.; Yang, W.; Francis, D.M. Association Analysis for Bacterial Spot Resistance in a Directionally Selected Complex Breeding Population of Tomato. Phytopathology 2015, 105, 1437–1445. [Google Scholar] [CrossRef] [PubMed]
- Gill, U.; Scott, J.W.; Shekasteband, R.; Ogundiwin, E.; Schuit, C.; Francis, D.M.; Sim, S.C.; Smith, H.; Hutton, S.F. Ty-6, a Major Begomovirus Resistance Gene on Chromosome 10, Is Effective against Tomato Yellow Leaf Curl Virus and Tomato Mottle Virus. Theor. Appl. Genet. 2019, 132, 1543–1554. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.C.; Durstewitz, G.; Plieske, J.; Wieseke, R.; Ganal, M.W.; van Deynze, A.; Hamilton, J.P.; Buell, C.R.; Causse, M.; Wijeratne, S.; et al. Development of a Large SNP Genotyping Array and Generation of High-Density Genetic Maps in Tomato. PLoS ONE 2012, 7, e40563. [Google Scholar] [CrossRef]
- Sim, S.C.; van Deynze, A.; Stoffel, K.; Douches, D.S.; Zarka, D.; Ganal, M.W.; Chetelat, R.T.; Hutton, S.F.; Scott, J.W.; Gardner, R.G.; et al. High-Density SNP Genotyping of Tomato (Solanum lycopersicum L.) Reveals Patterns of Genetic Variation Due to Breeding. PLoS ONE 2012, 7, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Broman, K.W.; Wu, H.; Sen, Ś.; Churchill, G.A. R/Qtl: QTL Mapping in Experimental Crosses. Bioinformatics 2003, 19, 889–890. [Google Scholar] [CrossRef] [PubMed]
- R Core Team R: A Language and Environment for Statistical Computing 2021. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R-project.org/ (accessed on 19 May 2024).
- Hosmani, P.S.; Flores-Gonzalez, M.; Van De Geest, H.; Maumus, F.; Bakker, L.V.; Schijlen, E.; Van Haarst, J.; Cordewener, J.; Sanchez-Perez, G.; Peters, S.; et al. An Improved de Novo Assembly and Annotation of the Tomato Reference Genome Using Single-Molecule Sequencing, Hi-C Proximity Ligation and Optical Maps. bioRxiv 2019, 767764. [Google Scholar] [CrossRef]
- Kosambi, D.D. The Estimation of Map Distances from Recombination Values. In Selected Works in Mathematics and Statistics; Ramaswamy, R., Ed.; Springer: New Delhi, India, 2016; pp. 125–130. ISBN 978-81-322-3676-4. [Google Scholar]
- Haley, C.S.; Knott, S.A. A Simple Regression Method for Mapping Quantitative Trait Loci in Line Crosses Using Flanking Markers. Heredity 1992, 69, 315–324. [Google Scholar] [CrossRef]
- Adhikari, L.; Missaoui, A.M. Quantitative Trait Loci Mapping of Leaf Rust Resistance in Tetraploid Alfalfa. Physiol. Mol. Plant Pathol. 2019, 106, 238–245. [Google Scholar] [CrossRef]
- Fenstemaker, S.; Sim, L.; Cooperstone, J.; Francis, D. Solanum galapagense-Derived Purple Tomato Fruit Color Is Conferred by Novel Alleles of the Anthocyanin Fruit and Atroviolacium Loci. Plant Direct 2022, 6, e394. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Wani, S.H.; Collins, P.J.; Wen, Z.; Gu, C.; Chilvers, M.I.; Wang, D. Mapping Quantitative Trait Loci for Tolerance to Pythium irregulare in Soybean (Glycine max L.). G3 Genes Genomes Genet. 2018, 8, 3155–3161. [Google Scholar] [CrossRef] [PubMed]
- Churchill, G.A.; Doerge, R.W. Empirical Threshold Values for Quantitative Trait Mapping. Genetics 1994, 138, 963–971. [Google Scholar] [CrossRef] [PubMed]
- de Mendiburu, F. Agricolae: Statistical Procedures for Agricultural. Research. R Package Version 1.3-5 2021. Available online: https://CRAN.R-project.org/package=agricolae (accessed on 19 May 2024).
- Endelman, J.B. Ridge Regression and Other Kernels for Genomic Selection with R Package RrBLUP. Plant Genome 2011, 4, 250–255. [Google Scholar] [CrossRef]
- Baral, K.; Coulman, B.; Biligetu, B.; Fu, Y.B. Advancing Crested Wheatgrass [Agropyron cristatum (L.) Gaertn.] Breeding through Genotyping-by-Sequencing and Genomic Selection. PLoS ONE 2020, 15, e0239609. [Google Scholar] [CrossRef] [PubMed]
- Voorrips, R.E. MapChart: Software for the Graphical Presentation of Linkage Maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef]
- Doerge, R.W. Mapping and Analysis of Quantitative Trait Loci in Experimental Populations. Nat. Rev. Genet. 2002, 3, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Zou, F. QTL Mapping in Intercross and Backcross Populations. In Cardiovascular Genomics; Methods in Molecular Biology Volume 573; DiPetrillo, K., Ed.; Humana Press: Totowa, NJ, USA, 2009; pp. 157–173. [Google Scholar]
- Rebaï, A. Comparison of Methods for Regression Interval Mapping in QTL Analysis with Non-Normal Traits. Genet. Res. 1997, 69, 69–74. [Google Scholar] [CrossRef]
- Bernacchi, D.; Beck-Bunn, T.; Eshed, Y.; Lopez, J.; Petiard, V.; Uhlig, J.; Zamir, D.; Tanksley, S. Advanced Backcross QTL Analysis in Tomato. I. Identification of QTLs for Traits of Agronomic Importance from Lycopersicon hirsutum. Theor. Appl. Genet. 1998, 97, 381–397. [Google Scholar] [CrossRef]
- Frary, A.; Fulton, T.M.; Zamir, D.; Tanksley, S.D. Advanced Backcross QTL Analysis of a Lycopersicon esculentum x L. pennellii Cross and Identification of Possible Orthologs in the Solanaceae. Theor. Appl. Genet. 2004, 108, 485–496. [Google Scholar] [CrossRef] [PubMed]
- Fulton, T.M.; Grandillo, S.; Beck-Bunn, T.; Fridman, E.; Frampton, A.; Lopez, J.; Petiard, V.; Uhlig, J.; Zamir, D.; Tanksley, S.D. Advanced backcross QTL analysis of a Lycopersicon esculentum × Lycopersicon parviflorum cross. Theor. Appl. Genet. 2000, 100, 1025–1042. [Google Scholar] [CrossRef]
- Tanksley, S.D.; Grandillo, S.; Fulton Zamir, T.D.; Eshed, Y.; Petiard -J Lopez Beck-Bunn, V.T.; Zamir, D.; Eshed, T.; Lopez Hispareco, J.S. Advanced Backcross QTL Analysis in a Cross between an Elite Processing Line of Tomato and Its Wild Relative L. pimpinellifolium. Theor. Appl. Genet. 1996, 92, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Fulton, T.M.; Beck-Bunn, T.; Emmatty, D.; Eshed, Y.; Lopez, J.; Petiard, V.; Uhlig, J.; Zamir, D.; Tanksley, S.D. QTL analysis of an advanced backcross of Lycopersicon peruvianum to the cultivated tomato and comparisons with QTLs found in other wild species. Theor. Appl. Genet. 1997, 95, 881–894. [Google Scholar] [CrossRef]
- Heffner, E.L.; Sorrells, M.E.; Jannink, J.L. Genomic Selection for Crop Improvement. Crop Sci. 2009, 49, 1–12. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genetic Map vs. Physical Map (Sl4.0) Correlation | |||||||
|---|---|---|---|---|---|---|---|
| Chr | Number of Markers | Chr Length (cM) | Average Distance between Markers (cM) | Largest Distance between Markers (cM) | a p-Value | b R2 | c ρ |
| 1 | 12 | 54.2 | 4.9 | 10.6 | 0.0001 | 0.7581 | 1.0000 |
| 2 | 14 | 81.2 | 6.2 | 34.3 | 0.0000 | 0.7522 | 1.0000 |
| 3 | 6 | 41 | 8.2 | 28.7 | 0.0006 | 0.9509 | 1.0000 |
| 4a | 27 | 37.9 | 1.5 | 13.6 | 0.0000 | 0.4790 | 1.0000 |
| 4b | 3 | 0.5 | 0.2 | 0.5 | 0.2305 | 0.7491 | 1.0000 |
| 5 | 16 | 35 | 2.3 | 9.9 | 0.0000 | 0.7728 | 1.0000 |
| 6 | 9 | 93.3 | 11.7 | 46.8 | 0.0001 | 0.8960 | 1.0000 |
| 7 | 11 | 47.7 | 4.8 | 17.2 | 0.0002 | 0.7813 | 1.0000 |
| 8 | 7 | 55.1 | 9.2 | 33 | 0.0061 | 0.7671 | 1.0000 |
| 9 | 13 | 62.6 | 5.2 | 51.4 | 0.0000 | 0.9945 | 1.0000 |
| 10 | 10 | 63.6 | 7.1 | 31 | 0.0012 | 0.7198 | 1.0000 |
| 11 | 3 | 11.1 | 5.6 | 9.1 | 0.5260 | −0.0803 | 1.0000 |
| 12 | 5 | 14.9 | 3.7 | 8.2 | 0.1730 | 0.3516 | 1.0000 |
| Overall | 136 | 598 | 4.9 | 51.4 | |||
| Location | Variable | Mapping Method | Chr | QTL Peak Position (cM) | LOD Peak | a LOD Cutoff | b Nearest Marker | c PVE | d Additive Effect | e Odds Ratio | f Beneficial Allele | g Physical Position of Nearest Marker (bp) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fremont | Percentage of puffy fruit | CIM | 1 | 17 | 2.69 | 2.46 | solcap_snp_sl_20440 | 6.40 | −4.40 | - | B | 3,637,274 |
| 2 | 32 | 3.06 | 2.46 | solcap_snp_sl_13625 | 8.30 | −5.00 | - | B | 40,480,064 | |||
| 4 | 0 | 2.83 | 2.46 | solcap_snp_sl_21372 | 7.10 | 4.70 | - | A | 2,951,634 | |||
| Percentage of puffy fruit | IM-NP | 1 | 20 | 2.86 | 2.40 | solcap_snp_sl_18619 | 5.00 | −3.90 | - | B | 68,611,189 | |
| 2 | 32 | 2.63 | 2.40 | solcap_snp_sl_13625 | 8.30 | −5.00 | - | B | 40,480,064 | |||
| Incidence of puffy fruit | IM-BI | 1 | 21 | 2.67 | 2.48 | solcap_snp_sl_18619 | - | - | 3.40 | B | 68,611,189 | |
| Both Locations | Percentage of puffy fruit | CIM | 2 | 32 | 3.59 | 2.40 | solcap_snp_sl_13625 | 9.50 | −4.10 | - | B | 40,480,064 |
| Percentage of puffy fruit | IM-NP | 1 | 29 | 3.50 | 2.46 | solcap_snp_sl_531 | 5.40 | −3.10 | - | B | 73,097,261 | |
| 2 | 32 | 3.18 | 2.46 | solcap_snp_sl_13625 | 9.50 | −4.10 | - | B | 40,480,064 | |||
| Incidence of puffy fruit | IM-BI | 1 | 28 | 3.25 | 2.49 | solcap_snp_sl_4963 | - | - | 3.60 | B | 70,083,502 | |
| 2 | 32 | 2.55 | 2.49 | solcap_snp_sl_13625 | - | - | 3.90 | B | 40,480,064 |
| p-Values of ANOVA F Tests from Marker–Trait Regressions | |||||
|---|---|---|---|---|---|
| Chr | Marker | Validation Population | 2K20-8312 Sub-Population | 2K20-8322 Sub-Population | 2K20-8357 Sub-Population |
| 1 | solcap_snp_sl_33701 | 5.59 × 10−7 | 0.83 | monomorphic B | monomorphic B |
| 1 | solcap_snp_sl_8669 | 1.04 × 10−4 | 0.71 | 0.33 | monomorphic B |
| 1 | solcap_snp_sl_100164 | 1.32 × 10−9 | monomorphic A | 0.31 | monomorphic B |
| 1 | solcap_snp_sl_20440 | 3.02 × 10−11 | monomorphic A | monomorphic A | monomorphic B |
| 1 | solcap_snp_sl_18619 | 3.02 × 10−11 | monomorphic A | monomorphic A | monomorphic B |
| 1 | solcap_snp_sl_4963 | 4.54 × 10−7 | 0.85 | 0.45 | monomorphic B |
| 1 | solcap_snp_sl_531 | 5.05 × 10−9 | 0.75 | 0.04 | monomorphic B |
| 2 | solcap_snp_sl_8505 | 0.80 | monomorphic A | 0.92 | monomorphic A |
| 2 | solcap_snp_sl_8510 | 0.80 | monomorphic A | 0.92 | monomorphic A |
| 2 | solcap_snp_sl_23850 | 0.68 | monomorphic A | 0.25 | monomorphic A |
| 2 | solcap_snp_sl_13625 | 0.76 | monomorphic A | 0.59 | monomorphic A |
| 4 | solcap_snp_sl_21372 | 0.94 | 0.44 | 0.94 | 0.36 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dalprá Dariva, F.; Subode, S.; Cho, J.; Nick, C.; Francis, D. Identification and Validation of Quantitative Trait Loci Associated with Fruit Puffiness in a Processing Tomato Population. Plants 2024, 13, 1454. https://doi.org/10.3390/plants13111454
Dalprá Dariva F, Subode S, Cho J, Nick C, Francis D. Identification and Validation of Quantitative Trait Loci Associated with Fruit Puffiness in a Processing Tomato Population. Plants. 2024; 13(11):1454. https://doi.org/10.3390/plants13111454
Chicago/Turabian StyleDalprá Dariva, Françoise, Su Subode, Jihuen Cho, Carlos Nick, and David Francis. 2024. "Identification and Validation of Quantitative Trait Loci Associated with Fruit Puffiness in a Processing Tomato Population" Plants 13, no. 11: 1454. https://doi.org/10.3390/plants13111454
APA StyleDalprá Dariva, F., Subode, S., Cho, J., Nick, C., & Francis, D. (2024). Identification and Validation of Quantitative Trait Loci Associated with Fruit Puffiness in a Processing Tomato Population. Plants, 13(11), 1454. https://doi.org/10.3390/plants13111454