
Drip Fertigation Enhances the Responses of Grain Yield and Quality to Nitrogen Topdressing Rate in Irrigated Winter Wheat in North China

 and
and
Abstract
1. Introduction
2. Results
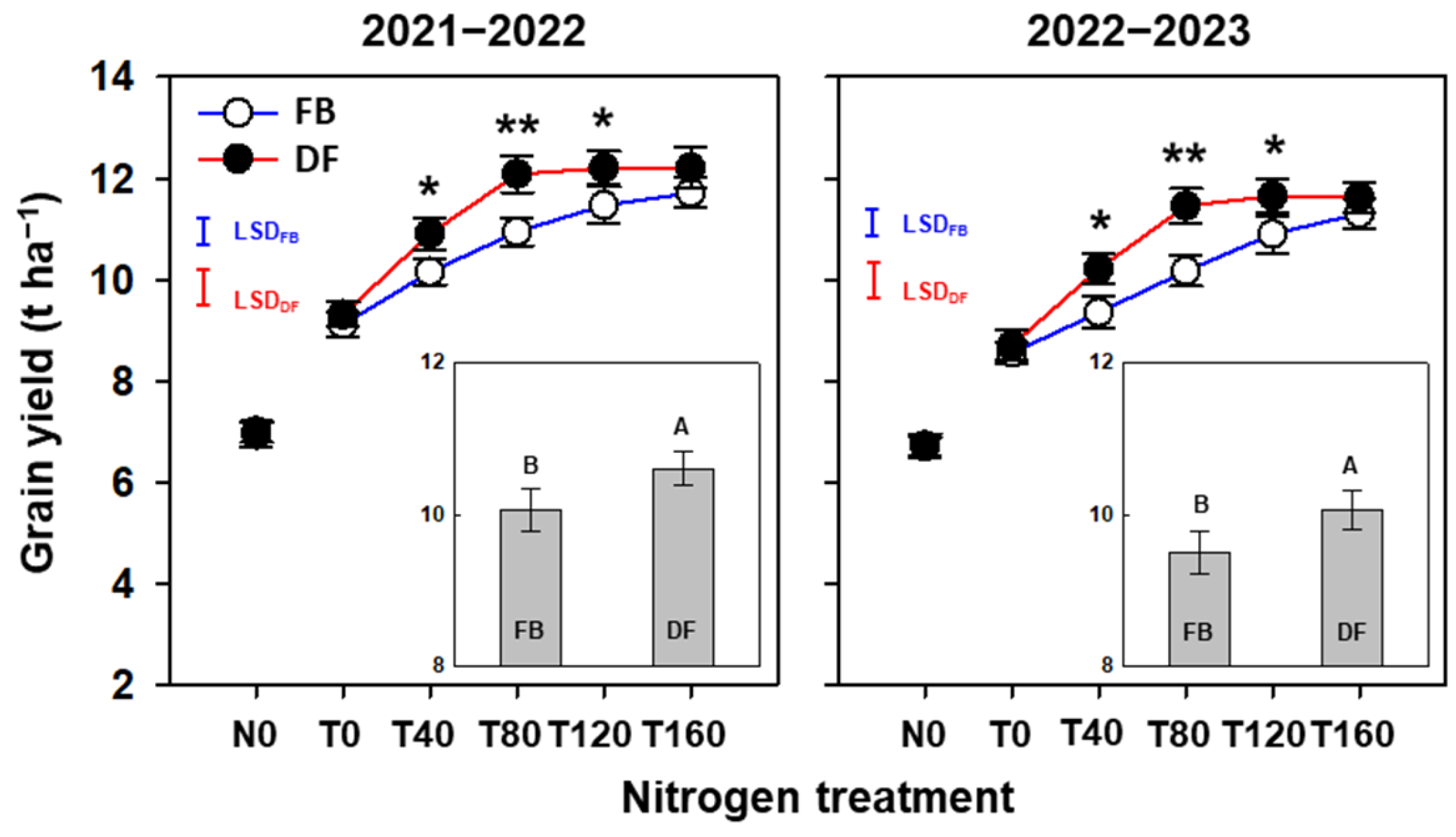
2.1. Yield and Yield-Related Attributes
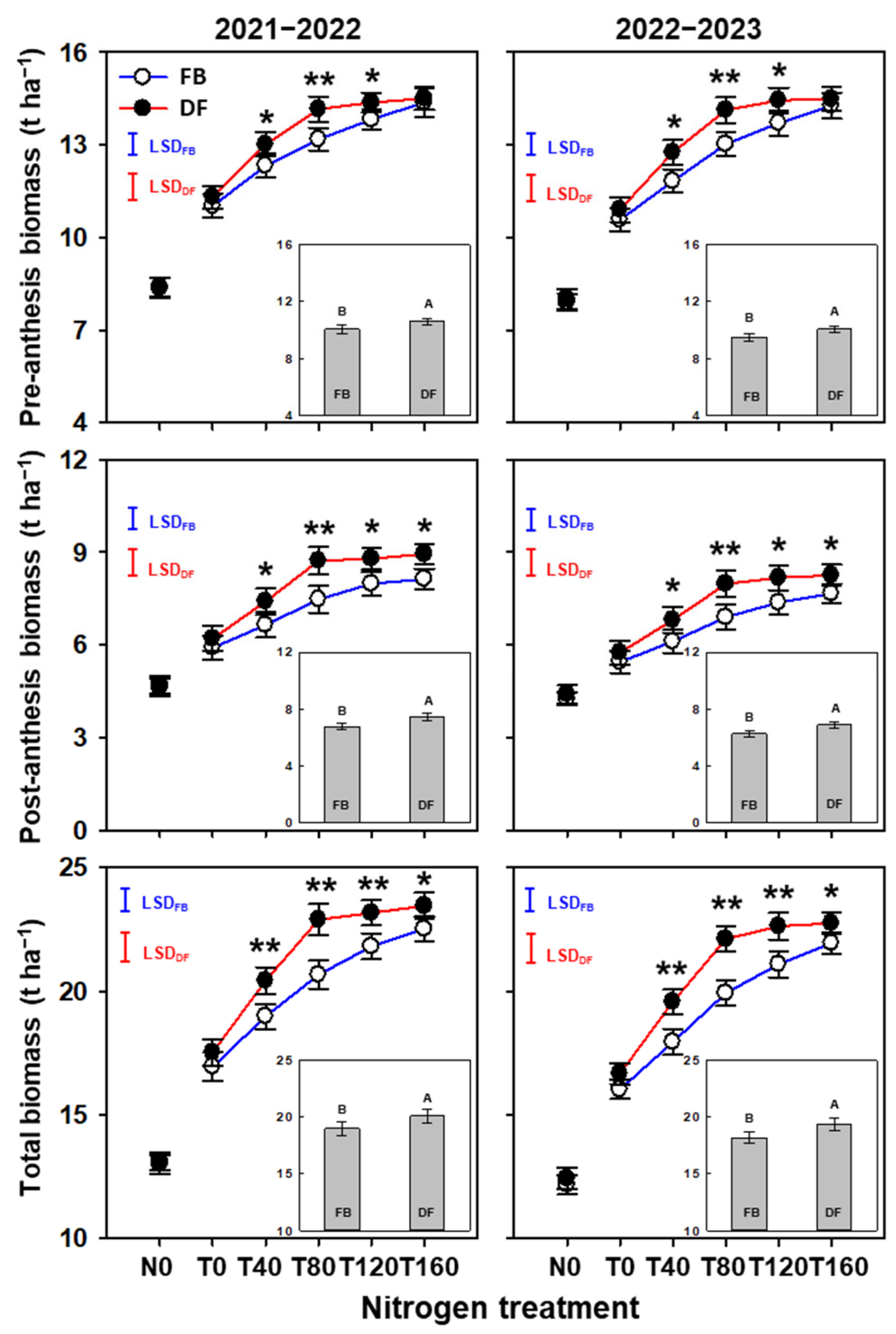
2.2. Biomass Accumulation
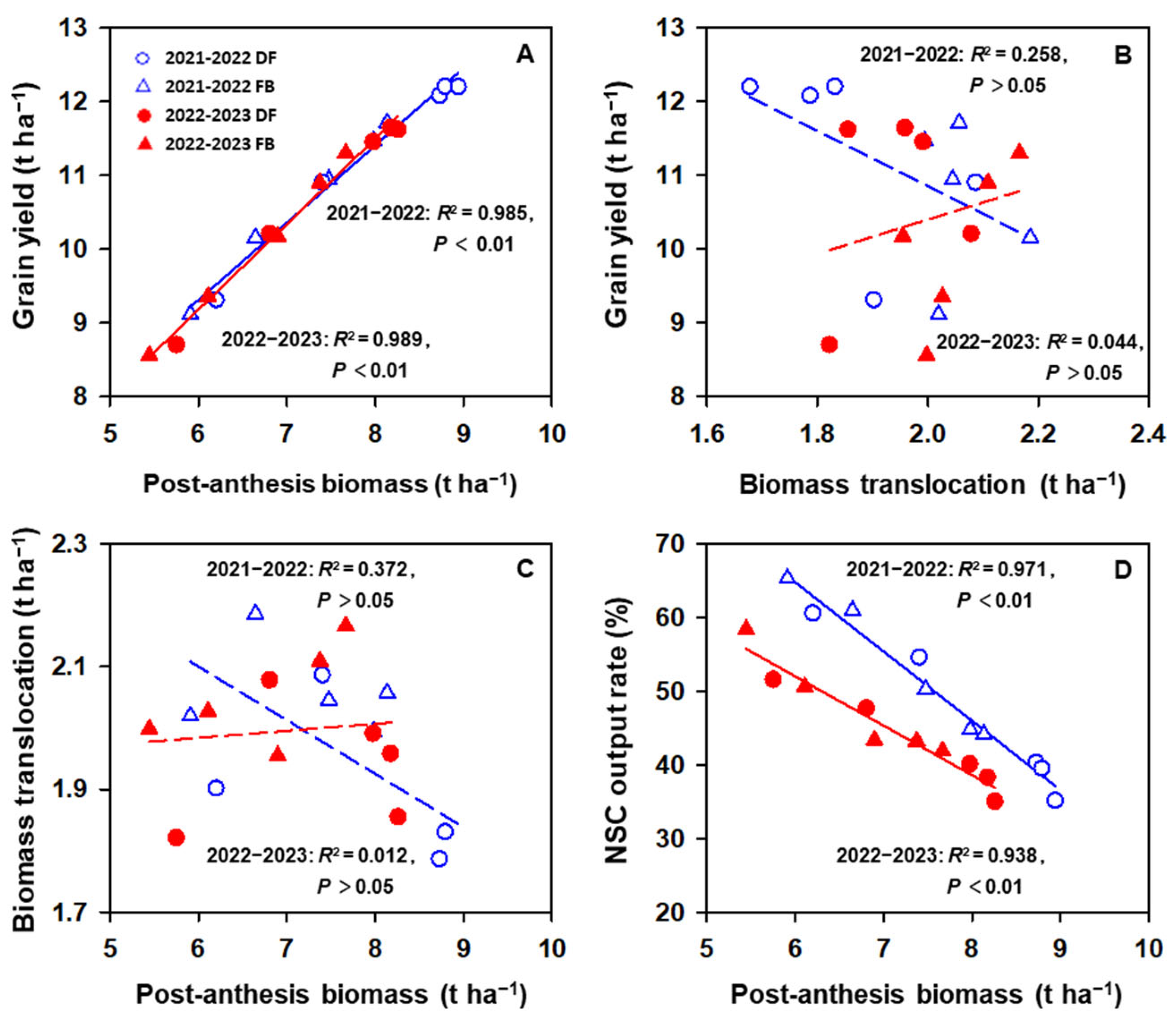
2.3. NSC Accumulation and Biomass Translocation and Partitioning
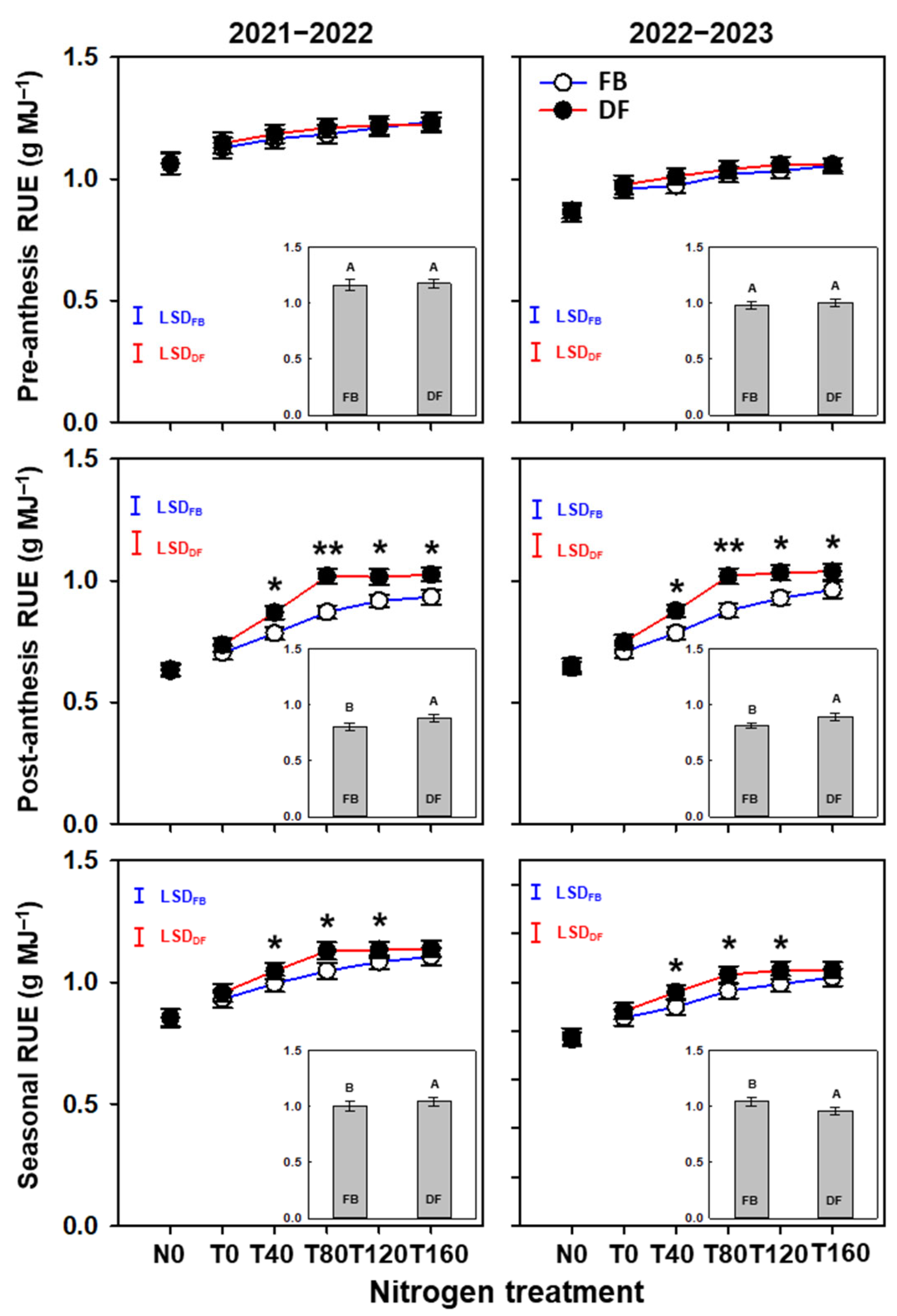
2.4. PAR Interception and Use Efficiency
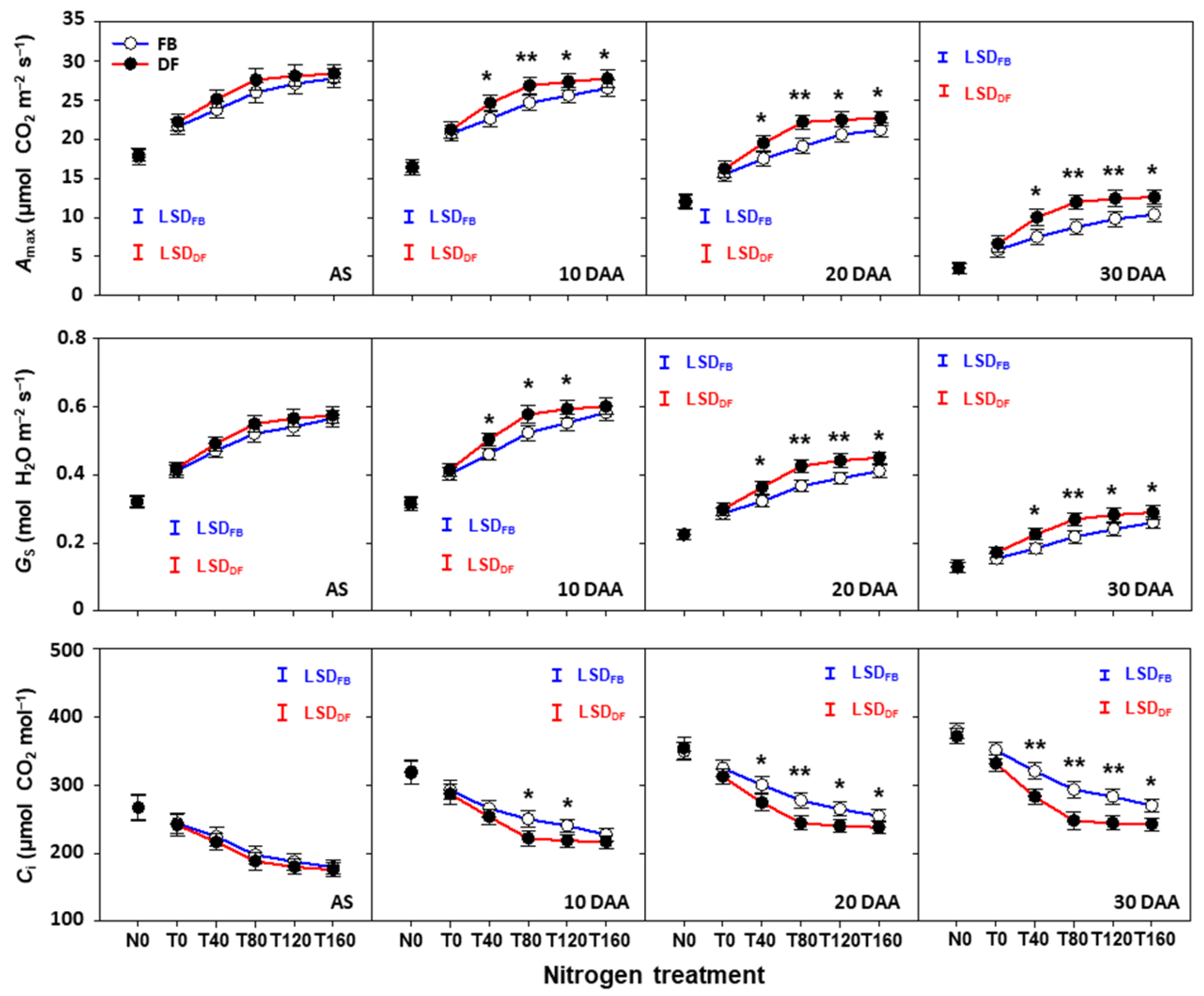
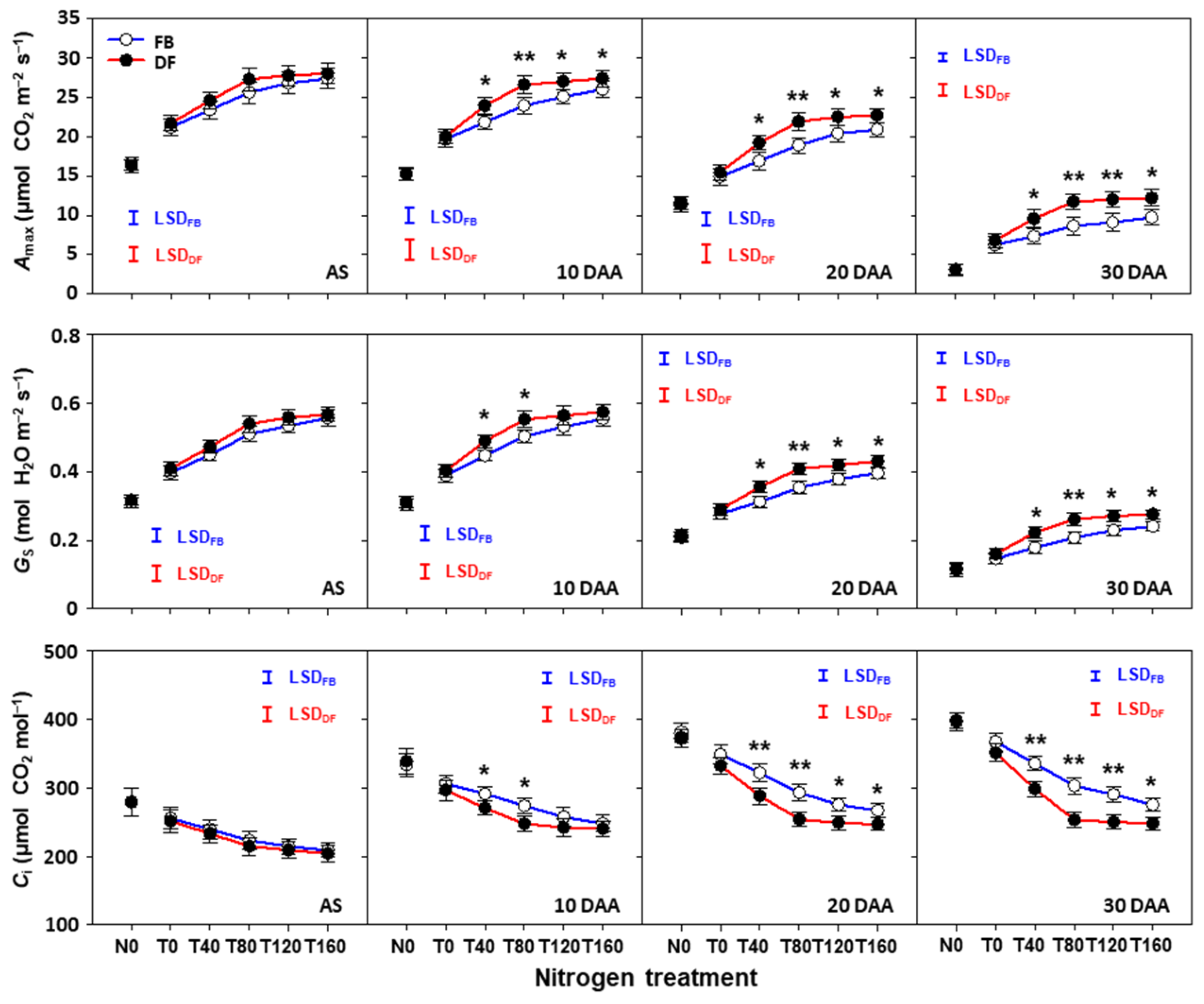
2.5. Photosynthetic Characteristics of Flag Leaves after Anthesis
2.6. SLN Contents of Flag Leaf after Anthesis
2.7. CAT and SOD Activities and MDA Contents of Flag Leaves after Anthesis
2.8. Grain Protein Content and Protein Components
2.9. Grain Processing Quality
3. Discussion
4. Materials and Methods
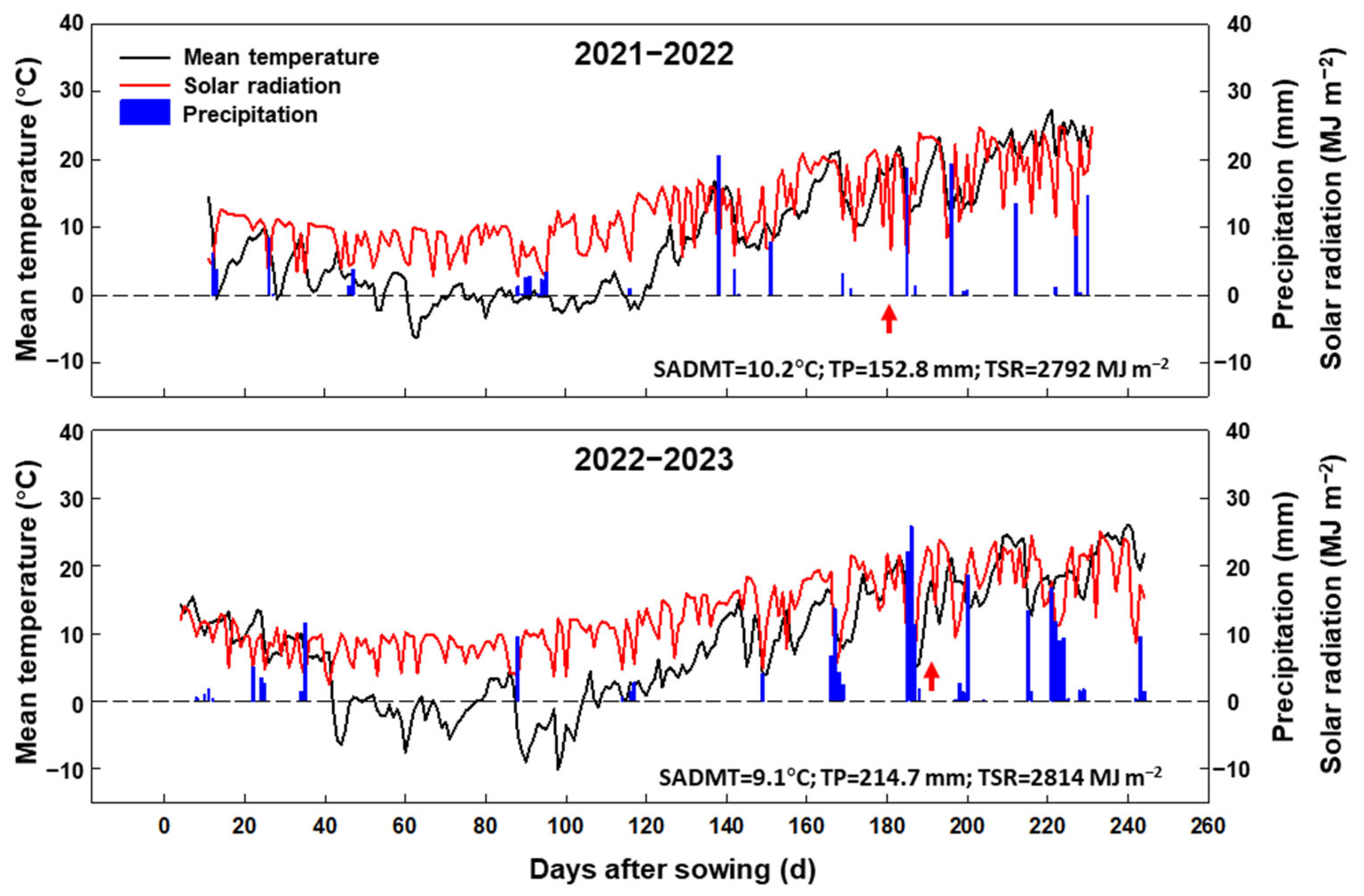
4.1. Site Description
4.2. Experimental Design and Field Management
4.3. Sampling and Measurements
4.3.1. Grain Yield and Yield Components
4.3.2. Biomass
4.3.3. Maximum Number of Stems and LAI
4.3.4. SLN
4.3.5. Light Interception and Radiation Use Efficiency
4.3.6. Photosynthetic Characteristics
4.3.7. Catalase (CAT) and Superoxide Dismutase (SOD) Activities, and Malondialdehyde (MDA) Contents
4.3.8. Protein Content of Grains and Protein Components
4.3.9. Grain Quality
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO (Food and Agriculture Organization). Online Statistical Database: Crops and Livestock Products. FAOSTAT. 2023. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 19 March 2024).
- National Bureau of Statistics of China. China Statistical Yearbook; China Statistics Press: Beijing, China, 2022; p. 453. [Google Scholar]
- Kang, S.Z.; Zhang, L.; Liang, Y.L.; Hu, X.T.; Cai, H.J.; Gu, B.T. Effects of limited irrigation on yield and water use efficiency of winter wheat in the Loess Plateau of China. Agric. Water Manag. 2002, 55, 203–216. [Google Scholar] [CrossRef]
- Dang, J.Y.; Pei, X.X.; Zhang, D.Y.; Zhang, J.; Cheng, M.F.; Wang, J.A.; Gao, L. Effects of integration of micro-sprinkler irrigation and nitrogen on growth and development of winter wheat and water and fertilizer use efficiency. Chin. J. Appl. Ecol. 2020, 31, 3700–3710. [Google Scholar]
- Peng, S.B.; Bouman, B.; Visperas, R.M.; Castañeda, A.; Nie, L.X.; Park, H.K. Comparison between aerobic and flooded rice in the tropics: Agronomic performance in an eight-season experiment. Field Crops Res. 2006, 96, 252–259. [Google Scholar] [CrossRef]
- Dalin, C.; Wada, Y.; Kastner, T.; Puma, M.J. Groundwater depletion embedded in international food trade. Nature 2017, 543, 700–704. [Google Scholar] [CrossRef] [PubMed]
- Shahdany, S.M.H.; Firoozfar, A.; Maestre, J.M.; Mallakpour, I.; Taghvaeian, S.; Karimi, P. Operational performance improvements in irrigation canals to overcome groundwater overexploitation. Agric. Water Manag. 2018, 204, 234–246. [Google Scholar] [CrossRef]
- Uhart, S.A.; Andrade, F.H. Nitrogen deficiency in maize I. Effects on crop growth, development, dry matter partitioning, and kernel set. Crop Sci. 1995, 35, 1376–1383. [Google Scholar] [CrossRef]
- Prystupa, P.; Slafer, G.A.; Savin, R. Leaf appearance, tillering and their coordination in response to NxP fertilization in barley. Plant Soil 2003, 255, 587–594. [Google Scholar] [CrossRef]
- Yang, D.Q.; Cai, T.; Luo, Y.L.; Wang, Z.L. Optimizing plant density and nitrogen application to manipulate tiller growth and increase grain yield and nitrogen-use efficiency in winter wheat. PeerJ 2019, 7, e6484. [Google Scholar] [CrossRef] [PubMed]
- Matson, P.A.; Naylor, R.; Ortiz-Monasterio, I. Integration of environmental, agronomic, and economic aspects of fertilizer management. Science 1998, 280, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Dar, E.A.; Brar, A.S.; Mishra, S.K.; Singh, K.B. Simulating response of wheat to timing and depth of irrigation water in drip irrigation system using CERES-wheat model. Field Crops Res. 2017, 214, 149–163. [Google Scholar] [CrossRef]
- Lv, H.F.; Zhao, Y.M.; Wang, Y.F.; Wan, L.; Wang, J.G.; Butterbach-Bahl, K.; Lin, S. Conventional flooding irrigation and over fertilization drives soil pH decrease not only in the top-but also in subsoil layers in solar greenhouse vegetable production systems. Geoderma 2020, 363, 114156. [Google Scholar] [CrossRef]
- Lu, J.S.; Xiang, Y.Z.; Fan, J.L.; Zhang, F.C.; Hu, T.T. Sustainable high grain yield, nitrogen use efficiency and water productivity can be achieved in wheat-maize rotation system by changing irrigation and fertilization strategy. Agric. Water Manag. 2021, 258, 107177. [Google Scholar] [CrossRef]
- Si, Z.Y.; Zain, M.; Mehmood, F.; Wang, G.S.; Gao, Y.; Duan, A.W. Effects of nitrogen application rate and irrigation regime on growth, yield, and water-nitrogen use efficiency of drip-irrigated winter wheat in the North China Plain. Agric. Water Manag. 2020, 231, 106002. [Google Scholar] [CrossRef]
- Yan, F.L.; Zhang, F.C.; Fan, X.K.; Fan, J.L.; Wang, Y.; Zou, H.Y.; Wang, H.D.; Li, G.D. Determining irrigation amount and fertilization rate to simultaneously optimize grain yield, grain nitrogen accumulation and economic benefit of drip-fertigated spring maize in northwest China. Agric. Water Manag. 2021, 243, 106440. [Google Scholar] [CrossRef]
- Hagin, J.; Lowengart, A. Fertigation for minimizing environmental pollution by fertilizers. Fertil. Res. 1995, 43, 5–7. [Google Scholar] [CrossRef]
- Fan, J.L.; Wu, L.F.; Zhang, F.C.; Yan, S.C.; Xiang, Y.Z. Evaluation of drip fertigation uniformity affected by injector type, pressure difference and lateral layout. Irrig. Drain. 2017, 66, 520–529. [Google Scholar] [CrossRef]
- Yan, S.C.; Wu, Y.; Fan, J.L.; Zhang, F.C.; Guo, J.J.; Zheng, J.; Wu, L.F. Quantifying grain yield, protein, nutrient uptake and utilization of winter wheat under various drip fertigation regimes. Agric. Water Manag. 2022, 261, 107380. [Google Scholar] [CrossRef]
- Wang, X.K.; Yun, J.; Shi, P.; Li, Z.B.; Li, P.; Xing, Y.Y. Root growth, fruit yield and water use efficiency of greenhouse grown tomato under different irrigation regimes and nitrogen levels. J. Plant Growth Regul. 2019, 38, 400–415. [Google Scholar] [CrossRef]
- Fanish, S.A.; Muthukrishnan, P.; Santhi, P. Effect of drip fertigation on field crops—A review. Agric. Rev. 2011, 32, 14–25. [Google Scholar]
- Jha, S.K.; Ramatshaba, T.S.; Wang, G.S.; Liang, Y.; Liu, H.; Gao, Y.P.; Liu, H.; Gao, Y.; Duan, A.W. Response of growth, yield and water use efficiency of winter wheat to different irrigation methods and scheduling in North China Plain. Agric. Water Manag. 2019, 217, 292–302. [Google Scholar]
- Soni, J.K.; Raja, N.A.; Kumar, V. Improving productivity of groundnut (Arachis hypogaea L.) under drip and micro sprinkler fertigation system. Legume Res.-Int. J. 2019, 42, 90–95. [Google Scholar] [CrossRef]
- Zheng, J.; Zhou, M.H.; Zhu, B.; Fan, J.L.; Lin, H.Y.; Ren, B.; Zhang, F.C. Drip fertigation sustains crop productivity while mitigating reactive nitrogen losses in Chinese agricultural systems: Evidence from a meta-analysis. Sci. Total Environ. 2023, 886, 163804. [Google Scholar] [CrossRef] [PubMed]
- Delbaz, R.; Ebrahimian, H.; Abbasi, F.; Ghameshlou, A.N.; Liaghat, A.; Ranazadeh, D. A global meta-analysis on surface and drip fertigation for annual crops under different fertilization levels. Agric. Water Manag. 2023, 289, 108504. [Google Scholar] [CrossRef]
- Li, H.R.; Mei, X.R.; Wang, J.D.; Huang, F.; Hao, W.P.; Li, B.G. Drip fertigation significantly increased crop yield, water productivity and nitrogen use efficiency with respect to traditional irrigation and fertilization practices: A meta-analysis in China. Agric. Water Manag. 2021, 244, 106534. [Google Scholar] [CrossRef]
- Cassman, K.G. Ecological intensification of cereal production systems: Yield potential, soil quality, and precision agriculture. Proc. Natl. Acad. Sci. USA 1999, 96, 5952–5959. [Google Scholar] [CrossRef] [PubMed]
- Parry, M.A.; Reynolds, M.; Salvucci, M.E.; Raines, C.; Andralojc, P.J.; Zhu, X.G.; Price, G.D.; Condon, A.G.; Furbank, R.T. Raising yield potential of wheat. II. Increasing photosynthetic capacity and efficiency. J. Exp. Bot. 2011, 62, 453–467. [Google Scholar] [CrossRef] [PubMed]
- Monteith, J.L. Solar radiation and productivity in tropical ecosystems. J. Appl. Ecol. 1972, 9, 747–766. [Google Scholar] [CrossRef]
- Monteith, J.L. Climate and the efficiency of crop production in Britain. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1977, 281, 277–294. [Google Scholar]
- Zhu, X.G.; Long, S.P.; Ort, D.R. Improving photosynthetic efficiency for greater yield. Annu. Rev. Plant Boil. 2010, 61, 235. [Google Scholar] [CrossRef]
- Allard, V.; Martre, P.; Le Gouis, J. Genetic variability in biomass allocation to roots in wheat is mainly related to crop tillering dynamics and nitrogen status. Eur. J. Agron. 2013, 46, 68–76. [Google Scholar] [CrossRef]
- Raymbek, A.; Saljnikov, E.; Kenenbayev, S.; Perovic, V.; Cakmak, D.; Ramazanova, S. Protein content changes in wheat grain as influenced by nitrogen fertilization. Agrochimica 2017, 61, 180–189. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Muchow, R.C. Radiation use efficiency. Adv. Agron. 1999, 65, 215–265. [Google Scholar]
- Fletcher, A.L.; Johnstone, P.R.; Chakwizira, E.; Brown, H.E. Radiation capture and radiation use efficiency in response to N supply for crop species with contrasting canopies. Field Crops Res. 2013, 150, 126–134. [Google Scholar] [CrossRef]
- Ullah, H.; Santiago-Arenas, R.; Ferdous, Z.; Attia, A.; Datta, A. Improving water use efficiency, nitrogen use efficiency, and radiation use efficiency in field crops under drought stress: A review. Adv. Agron. 2019, 156, 109–157. [Google Scholar]
- Wen, M.X.; Chen, D.A.; Li, D.S.; Qu, Z.X.; Cai, J.H. Regulating effect of nitrogen topdressing and planting density on population quality, yield and quality of Zhenmai 168. J. Triticeae Crops 2014, 34, 528–534. [Google Scholar]
- Zhao, G.C.; Chang, X.H.; Yang, Y.S.; Li, Z.H.; Feng, M.; Ma, S.K.; Yang, G.X. Grain yield and quality responding to the nitrogen fertilizer operation in different quality type wheat. J. Plant Nutr. Fert. 2010, 16, 859–865. [Google Scholar]
- Ma, R.Q.; Tao, Z.Q.; Wang, D.M.; Wang, Y.J.; Yang, Y.S.; Nu, Z.L.; Zhao, G.C.; Chang, X.H. Effects of topdressing nitrogen rate on photosynthetic characteristics and yield of flag leaves of wheat in different regions. J. Nucl. Agric. Sci. 2020, 34, 1281–1293. [Google Scholar]
- Ma, R.Q.; Tao, Z.Q.; Wang, D.M.; Wang, Y.J.; Yang, Y.S.; Zhu, Y.J.; Zhao, K.N.; Li, J.Z.; Wang, Y.J.; Chang, X.H.; et al. Effect of nitrogen topdressing rate on yield and quality of medium and strong gluten wheat cultivars. J. Plant Nutr. Fert. 2019, 25, 1799–1807. [Google Scholar]
- Guo, R.; Huang, X.G.; Wen, M.X.; Chen, C.; Liu, J.G.; Shan, Y.B.; Qu, Z.X.; Li, D.S. Effect of the nitrogen topdressing and plant density on grain yield and quality of spring strong gluten wheat cultivar zhenmai 12. J. Nucl. Agric. Sci. 2020, 34, 1834–1839. [Google Scholar]
- Zhang, L.; He, X.R.; Liang, Z.Y.; Zhang, W.; Zou, C.Q.; Chen, X.P. Tiller development affected by nitrogen fertilization in a high-yielding wheat production system. Crop Sci. 2020, 60, 1034–1047. [Google Scholar] [CrossRef]
- Yang, H.K.; Xiao, Y.; He, P.; Ai, D.L.; Zou, Q.S.; Hu, J.; Liu, L.; Huang, X.L.; Zheng, T.; Fan, G.Q. Straw mulch-based no-tillage improves tillering capability of dryland wheat by reducing asymmetric competition between main stem and tillers. Crop J. 2022, 10, 864–878. [Google Scholar] [CrossRef]
- Bai, S.S.; Kang, Y.H.; Wan, S.Q. Drip fertigation regimes for winter wheat in the North China Plain. Agric. Water Manag. 2020, 228, 105885. [Google Scholar] [CrossRef]
- Hamani, A.K.M.; Abubakar, S.A.; Si, Z.Y.; Kama, R.; Gao, Y.; Duan, A.W. Responses of grain yield and water-nitrogen dynamic of drip-irrigated winter wheat to different nitrogen fertigation and water regimes in the North China Plain. Agric. Water Manag. 2023, 288, 108494. [Google Scholar] [CrossRef]
- Dordas, C. Dry matter, nitrogen and phosphorus accumulation, partitioning and remobilization as affected by N and P fertilization and source-sink relations. Eur. J. Agron. 2009, 30, 129–139. [Google Scholar] [CrossRef]
- Manschadi, A.M.; Soltani, A. Variation in traits contributing to improved use of nitrogen in wheat: Implications for genotype by environment interaction. Field Crops Res. 2021, 270, 108211. [Google Scholar] [CrossRef]
- Ercoli, L.; Lulli, L.; Mariotti, M.; Masoni, A.; Arduini, I. Post-anthesis dry matter and nitrogen dynamics in durum wheat as affected by nitrogen supply and soil water availability. Eur. J. Agron. 2008, 28, 138–147. [Google Scholar] [CrossRef]
- Duan, J.Z.; Wu, Y.P.; Zhou, Y.; Ren, X.X.; Shao, Y.H.; Feng, W.; Zhu, Y.J.; Wang, Y.H.; Guo, T.C. Grain number responses to pre-anthesis dry matter and nitrogen in improving wheat yield in the Huang-Huai Plain. Sci. Rep. 2018, 8, 7126. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Wu, X.L.; Li, C.S.; Li, M.; Xiong, T.; Tang, Y.L. Dry matter and nitrogen accumulation, partitioning, and translocation in synthetic-derived wheat cultivars under nitrogen deficiency at the post-jointing stage. Field Crops Res. 2020, 248, 107720. [Google Scholar] [CrossRef]
- Li, H.T.; Shao, L.W.; Liu, X.W.; Sun, H.Y.; Chen, S.Y.; Zhang, X.Y. What matters more, biomass accumulation or allocation, in yield and water productivity improvement for winter wheat during the past two decades? Eur. J. Agron. 2023, 149, 126910. [Google Scholar] [CrossRef]
- Evans, J.R. Nitrogen and photosynthesis in the flag leaf of wheat (Triticum aestivum L.). Plant Physiol. 1983, 72, 297–302. [Google Scholar] [CrossRef]
- Carmo-Silva, E.; Andralojc, P.J.; Scales, J.C.; Driever, S.M.; Mead, A.; Lawson, T.; Raines, C.A.; Parry, M.A. Phenotyping of field-grown wheat in the UK highlights contribution of light response of photosynthesis and flag leaf longevity to grain yield. J. Exp. Bot. 2017, 68, 3473–3486. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.H.; Ma, L.L.; Yang, J.H.; Ding, W.J.; He, W.; Tang, Y.; Cui, G.J.; Zhang, W.J.; Ma, S.Y.; Ma, C.X.; et al. Night warming from tillering to jointing increases post-anthesis flag leaf photosynthetic capacity and wheat yield. Eur. J. Agron. 2023, 150, 126926. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, J.; Gong, S.; Xu, D.; Mo, Y.; Zhang, B. Straw mulching improves soil water content, increases flag leaf photosynthetic parameters and maintaines the yield of winter wheat with different irrigation amounts. Agric. Water Manag. 2021, 249, 106809. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, Y.; Guo, Q.; Zhang, P.; Cai, T.; Jia, Z. Adopting nitrogen deep placement based on different simulated precipitation year types enhances wheat yield and resource utilization by promoting photosynthesis capacity. Field Crops Res. 2023, 294, 108862. [Google Scholar] [CrossRef]
- Wang, H.Q.; Wang, R.R.; Jiang, G.Y.; Yin, H.J.; Yan, S.J.; Che, Z.Q. Effect of amount of nitrogen fertilizer applied on photosynthetic physiological characteristics of drip irrigated spring wheat leaves. Acta Agron. Sin. 2023, 49, 211–224. [Google Scholar]
- Sinclair, T.R.; Horie, T. Leaf nitrogen, photosynthesis, and crop radiation use efficiency: A review. Crop Sci. 1989, 29, 90–98. [Google Scholar] [CrossRef]
- Wang, C.Y.; Liu, W.X.; Li, Q.X.; Ma, D.Y.; Lu, H.F.; Feng, W.; Xie, Y.X.; Zhu, Y.J.; Guo, T.C. Effects of different irrigation and nitrogen regimes on root growth and its correlation with above-ground plant parts in high-yielding wheat under field conditions. Field Crops Res. 2014, 165, 138–149. [Google Scholar] [CrossRef]
- Zhang, P.P.; Ma, G.; Wang, C.Y.; Lu, H.F.; Li, S.S.; Xie, Y.X.; Ma, D.Y.; Zhu, Y.J.; Guo, T.C. Effect of irrigation and nitrogen application on grain amino acid composition and protein quality in winter wheat. PLoS ONE 2017, 12, e0178494. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, G.H.; Chen, Z.J.; Xiong, Y.W.; Huang, Q.Z.; Xu, X.; Huo, Z.L. Effects of irrigation and fertilization on grain yield, water and nitrogen dynamics and their use efficiency of spring wheat farmland in an arid agricultural watershed of Northwest China. Agric. Water Manag. 2022, 260, 107277. [Google Scholar] [CrossRef]
- Coventry, D.R.; Yadav, A.; Poswal, R.S.; Sharma, R.K.; Gupta, R.K.; Chhokar, R.S.; Gill, S.C.; Kumar, V.; Kumar, A.; Mehta, A.; et al. Irrigation and nitrogen scheduling as a requirement for optimising wheat yield and quality in Haryana, India. Field Crops Res. 2011, 123, 80–88. [Google Scholar] [CrossRef]
- Tari, A.F. The effects of different deficit irrigation strategies on yield, quality, and water-use efficiencies of wheat under semi-arid conditions. Agric. Water Manag. 2016, 167, 1–10. [Google Scholar] [CrossRef]
- Wieser, H.; Seilmeier, W. The influence of nitrogen fertilisation on quantities and proportions of different protein types in wheat flour. J. Sci. Food Agric. 1998, 76, 49–55. [Google Scholar] [CrossRef]
- Reynolds, M.; Bonnett, D.; Chapman, S.C.; Furbank, R.T.; Manès, Y.; Mather, D.E.; Parry, M.A. Raising yield potential of wheat. I. Overview of a consortium approach and breeding strategies. J. Exp. Bot. 2011, 62, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.X.; Luo, Y.L.; Li, C.H.; Chang, Y.L.; Jin, M.; Li, Y.; Wang, Z.L. Effects of nitrogen forms on nitrogen utilization, yield, and quality of two wheat varieties with different gluten characteristics. Eur. J. Agron. 2023, 149, 126919. [Google Scholar] [CrossRef]
- Millard, P. The accumulation and storage of nitrogen by herbaceous plants. Plant Cell Environ. 1988, 11, 1–8. [Google Scholar] [CrossRef]
- Pask, A.J.D.; Sylvester-Bradleyb, R.; Jamiesonc, P.D.; Foulkesa, M.J. Quantifying how winter wheat crops accumulate and use nitrogen reserves during growth. Field Crops Res. 2012, 126, 104–118. [Google Scholar] [CrossRef]
- Dier, M.; Hüther, L.; Schulze, W.X.; Erbs, M.; Köhler, P.; Weigel, H.J.; Manderscheid, R.; Zörb, C. Elevated atmospheric CO2 concentration has limited effect on wheat grain quality regardless of nitrogen supply. J. Agric. Food Chem. 2020, 68, 3711–3721. [Google Scholar] [CrossRef] [PubMed]
- Lyu, X.K.; Liu, Y.; Li, N.; Ku, L.B.; Hou, Y.T.; Wen, X.X. Foliar applications of various nitrogen (N) forms to winter wheat affect grain protein accumulation and quality via N metabolism and remobilization. Crop J. 2022, 10, 1165–1177. [Google Scholar] [CrossRef]
- Cock, J.H.; Yoshida, S.; Forno, D.A. Laboratory Manual for Physiological Studies of Rice; International Rice Research Institute: Los Baños, Philippines, 1976. [Google Scholar]
- Li, X.; Zhang, R.C.; Chen, G.; Xie, J.X.; Xiao, Z.W.; Cao, F.B.; Ali, I.; Anas, I.; Wahab, A.; Huang, M.; et al. Increasing grain weight and yield stability by increasing pre-heading non-structural carbohydrate reserves per spikelet in short-growth duration rice. Crop J. 2023, 11, 1912–1920. [Google Scholar] [CrossRef]
- Chen, Z.; Newman, I.; Zhou, M.; Mendham, N.; Zhang, G.; Shabala, S. Screening plants for salt tolerance by measuring K+ flux: A case study for barley. Plant Cell Environ. 2005, 28, 1230–1246. [Google Scholar] [CrossRef]
- Wang, X.K.; Huang, J.L. Principles and Techniques of Plant Physiological Biochemical Experiment; Higher Education Press: Beijing, China, 2000. [Google Scholar]
- Ahmad, I.; Kamran, M.; Yang, X.; Meng, X.; Ali, S.; Ahmad, S.; Zhang, X.; Bilegjargal, B.; Ahmad, B.; Liu, T.; et al. Effects of applying uniconazole alone or combined with manganese on the photosynthetic efficiency, antioxidant defense system, and yield in wheat in semiarid regions. Agric. Water Manag. 2019, 216, 400–414. [Google Scholar] [CrossRef]
- Luo, L.C.; Hui, X.L.; Wang, Z.H.; Zhang, X.; Xie, Y.H.; Gao, Z.Q.; Chai, S.X.; Lu, Q.L.; Li, T.L.; Sun, M.; et al. Multi-site evaluation of plastic film mulch and nitrogen fertilization for wheat grain yield, protein content and its components in semiarid areas of China. Field Crops Res. 2019, 240, 86–94. [Google Scholar] [CrossRef]
- National Grain and Oil Standardization Technical Committee. Method for Determination of Wet Gluten in Flour. GB/T 14608-1993. 1993. Available online: https://std.samr.gov.cn/gb/search/gbDetailed?id=71F772D78CA3D3A7E05397BE0A0AB82A (accessed on 19 May 2024).
- Ministry of Internal Trade of the PRC. Method for the Determination of Wet Gluten Quality in Wheat Flour-Gluten Index. LS/T 6102-1995. 1995. Available online: https://std.samr.gov.cn/hb/search/stdHBDetailed?id=8B1827F178E4BB19E05397BE0A0AB44A (accessed on 19 May 2024).
- State Administration for Market Regulation. Inspection of Grain and Oils-Doughs Rheological Properties Determination of Wheat Flour-Farinograph Test. GB/T 14614-2019. 2019. Available online: https://openstd.samr.gov.cn/bzgk/gb/newGbInfo?hcno=9C02083394E746C264A72D8717617CF4 (accessed on 19 May 2024).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growing Season | Management Pattern | N Treatment | Spikes Number | Grains per Spike | Grain Weight | Maximum Stem Number | Productive Stem Percentage | LAI at Anthesis |
|---|---|---|---|---|---|---|---|---|
| (m–2) | (mg) | (m–2) | (%) | (m2 m–2) | ||||
| 2021–2022 | FB | N0 | 435.0 e | 29.9 e | 53.9 a | 1356 b | 32.1 a | 4.21 d |
| T0 | 517.6 d | 34.4 d | 51.3 b | 1985 a | 26.1 d | 5.55 c | ||
| T40 | 555.8 c | 36.6 c | 49.9 c | 1973 a | 28.2 c | 6.06 b | ||
| T80 | 590.2 b | 37.6 b | 49.3 cd | 1986 a | 29.7 b | 6.47 a | ||
| T120 | 611.6 ab | 38.5 ab | 48.6 d | 1927 a | 31.7 a | 6.56 a | ||
| T160 | 620.0 a | 39.2 a | 48.2 d | 1989 a | 31.2 ab | 6.62 a | ||
| Mean | 555.0 B | 36.0 A | 50.2 B | 1869 A | 29.8 B | 5.91 A | ||
| DF | N0 | 439.0 d | 29.0 d | 54.6 a | 1319 b | 33.3 a | 4.15 d | |
| T0 | 531.7 c | 33.4 c | 52.4 ab | 1965 a | 27.1 c | 5.70 c | ||
| T40 | 589.3 b | 35.0 b | 52.8 b | 1912 a | 30.8 b | 6.26 b | ||
| T80 | 634.3 a | 36.4 a | 52.4 b | 1927 a | 32.9 a | 6.78 a | ||
| T120 | 649.3 a | 36.5 a | 51.5 c | 1909 a | 34.0 a | 6.84 a | ||
| T160 | 653.0 a | 37.2 a | 50.2 d | 1974 a | 33.1 a | 6.83 a | ||
| Mean | 582.8 A | 34.6 A | 52.3 A | 1834 A | 31.5 A | 6.09 A | ||
| ANOVA | ||||||||
| P | ** | ns | ** | ns | ** | ns | ||
| N | ** | ** | ** | ** | ** | ** | ||
| P × N | ** | * | ** | ns | * | ns | ||
| 2022–2023 | FB | N0 | 460.0 e | 28.7 e | 50.9 a | 1499 b | 30.7 a | 4.30 e |
| T0 | 552.5 d | 32.2 d | 48.1 b | 2190 a | 25.2 d | 5.73 d | ||
| T40 | 586.1 c | 34.0 c | 46.9 c | 2213 a | 26.5 c | 6.16 c | ||
| T80 | 611.8 b | 36.6 b | 45.5 d | 2197 a | 27.9 b | 6.62 a | ||
| T120 | 637.8 ab | 38.0 a | 45.0 d | 2196 a | 29.0 ab | 6.74 a | ||
| T160 | 657.4 a | 38.5 a | 44.7 d | 2181 a | 30.1 a | 6.90 a | ||
| Mean | 583.3 B | 34.7 A | 46.9 B | 2079 A | 28.2 B | 6.07 A | ||
| DF | N0 | 457.7 d | 28.8 d | 51.1 a | 1506 b | 30.4 b | 4.27 d | |
| T0 | 572.1 c | 30.8 c | 49.4 b | 2210 a | 25.9 d | 5.89 c | ||
| T40 | 628.6 b | 33.2 b | 48.9 b | 2162 a | 29.1 c | 6.41 b | ||
| T80 | 671.0 a | 35.2 a | 48.5 b | 2160 a | 31.1 ab | 6.85 a | ||
| T120 | 676.5 a | 36.0 a | 47.8 c | 2151 a | 31.4 ab | 6.95 a | ||
| T160 | 688.6 a | 36.1 a | 46.7 d | 2132 a | 32.3 a | 7.03 a | ||
| Mean | 615.7 A | 33.4 A | 48.7 A | 2053 A | 30.0 A | 6.23 A | ||
| ANOVA | ||||||||
| P | ** | ns | ** | ns | ** | ns | ||
| N | ** | ** | ** | ** | ** | ** | ||
| P × N | ** | * | ** | ns | * | ns |
| Growing Season | Management Pattern | N Treatment | NSC Accumulation at Anthesis | Biomass Translocation | NSC Output Rate | The Contribution Rate of Biomass Translocation to Grain | Harvest Index |
|---|---|---|---|---|---|---|---|
| (t ha–1) | (t ha–1) | (%) | (%) | (%) | |||
| 2021–2022 | FB | N0 | 2.13 e | 1.47 b | 69.2 a | 24.2 a | 46.9 a |
| T0 | 3.09 d | 2.02 a | 65.3 b | 25.5 a | 46.8 a | ||
| T40 | 3.59 c | 2.19 a | 60.9 c | 24.7 a | 46.6 a | ||
| T80 | 4.07 b | 2.05 a | 50.2 d | 21.5 b | 46.1 ab | ||
| T120 | 4.46 a | 1.99 a | 44.8 e | 20.0 b | 45.8 ab | ||
| T160 | 4.66 a | 2.06 a | 44.1 e | 20.2 b | 45.3 b | ||
| Mean | 3.67 A | 1.96 A | 55.8 A | 22.7 A | 46.2 A | ||
| DF | N0 | 1.97 e | 1.33 c | 67.5 a | 22.1 a | 46.1 a | |
| T0 | 3.14 d | 1.92 a | 60.6 b | 23.4 a | 46.5 a | ||
| T40 | 3.82 c | 2.09 a | 54.6 c | 22.0 a | 46.5 a | ||
| T80 | 4.43 b | 1.79 ab | 40.3 d | 17.0 b | 45.9 a | ||
| T120 | 4.63 ab | 1.83 ab | 39.5 d | 17.2 b | 45.9 a | ||
| T160 | 4.77 a | 1.68 b | 35.2 e | 15.8 b | 45.3 a | ||
| Mean | 3.79 A | 1.77 B | 49.6 B | 19.6 B | 46.0 A | ||
| ANOVA | |||||||
| P | ns | * | ** | ** | ns | ||
| N | ** | ** | ** | ** | * | ||
| P × N | ** | ** | ** | ** | ns | ||
| 2022–2023 | FB | N0 | 2.67 f | 1.60 b | 59.8 a | 27.3 a | 48.1 a |
| T0 | 3.42 e | 2.00 a | 58.4 a | 26.8 a | 46.5 b | ||
| T40 | 4.01 d | 2.03 a | 50.6 b | 24.9 b | 45.4 bc | ||
| T80 | 4.51 c | 1.96 a | 43.4 c | 22.1 c | 44.4 c | ||
| T120 | 4.88 b | 2.11 a | 43.2 c | 22.2 c | 45.0 bc | ||
| T160 | 5.17 a | 2.17 a | 41.9 c | 22.0 c | 44.8 bc | ||
| Mean | 4.11 A | 1.98 A | 49.5 A | 24.2 A | 45.7 A | ||
| DF | N0 | 2.69 e | 1.52 b | 54.2 a | 25.7 a | 47.7 a | |
| T0 | 3.53 d | 1.95 a | 51.6 b | 25.3 a | 46.3 b | ||
| T40 | 4.36 c | 2.08 a | 47.7 c | 23.4 b | 45.4 bc | ||
| T80 | 4.96 b | 1.99 a | 40.1 d | 20.0 c | 45.1 c | ||
| T120 | 5.11 ab | 1.96 a | 38.3 de | 19.3 c | 44.8 c | ||
| T160 | 5.30 a | 1.86 a | 35.0 e | 18.3 d | 44.5 c | ||
| Mean | 4.33 A | 1.89 A | 44.5 B | 22.0 B | 45.6 A | ||
| ANOVA | |||||||
| P | ns | ns | ** | ** | ns | ||
| N | ** | ** | ** | ** | ** | ||
| P × N | ** | ** | ** | ** | ns |
| Growing Season | Management Pattern | N Treatment | Protein Components (%) | Total Protein (%) | ||
|---|---|---|---|---|---|---|
| Albumin + Globulin | Gliadin | Glutenin | ||||
| 2021–2022 | FB | N0 | 3.18 a | 3.62 c | 4.86 c | 11.66 c |
| T0 | 3.22 a | 3.81 bc | 5.04 bc | 12.07 bc | ||
| T40 | 3.24 a | 4.03 ab | 5.26 ab | 12.53 ab | ||
| T80 | 3.26 a | 4.16 a | 5.39 a | 12.81 a | ||
| T120 | 3.28 a | 4.20 a | 5.43 a | 12.91 a | ||
| T160 | 3.27 a | 4.21 a | 5.46 a | 12.94 a | ||
| Mean | 3.25 A | 4.08 B | 5.32 B | 12.65 B | ||
| DF | N0 | 3.22 a | 3.70 e | 4.95 d | 11.87 e | |
| T0 | 3.26 a | 3.87 de | 5.05 d | 12.18 de | ||
| T40 | 3.25 a | 4.09 cd | 5.39 c | 12.73 cd | ||
| T80 | 3.27 a | 4.31 bc | 5.59 bc | 13.17 bc | ||
| T120 | 3.30 a | 4.51 ab | 5.80 ab | 13.61 ab | ||
| T160 | 3.28 a | 4.72 a | 6.03 a | 14.03 a | ||
| Mean | 3.27 A | 4.30 A | 5.57 A | 13.15 A | ||
| ANOVA | ||||||
| P | ns | ** | ** | ** | ||
| N | ns | ** | ** | ** | ||
| P × N | ns | ** | ** | ** | ||
| 2022–2023 | FB | N0 | 2.95 a | 3.52 c | 4.57 c | 11.04 c |
| T0 | 3.00 a | 3.71 bc | 4.78 bc | 11.49 bc | ||
| T40 | 3.03 a | 3.88 ab | 5.00 ab | 11.91 ab | ||
| T80 | 3.05 a | 4.03 a | 5.15 a | 12.23 a | ||
| T120 | 3.07 a | 4.07 a | 5.20 a | 12.34 a | ||
| T160 | 3.06 a | 4.11 a | 5.24 a | 12.41 a | ||
| Mean | 3.04 A | 3.96 B | 5.07 B | 12.08 B | ||
| DF | N0 | 2.98 a | 3.60 e | 4.66 e | 11.24 e | |
| T0 | 3.00 a | 3.80 de | 4.90 de | 11.71 de | ||
| T40 | 3.01 a | 4.01 cd | 5.08 cd | 12.09 cd | ||
| T80 | 3.05 a | 4.18 bc | 5.31 bc | 12.54 bc | ||
| T120 | 3.07 a | 4.39 b | 5.57 ab | 13.03 ab | ||
| T160 | 3.11 a | 4.63 a | 5.76 a | 13.50 a | ||
| Mean | 3.05 A | 4.20 A | 5.33 A | 12.58 A | ||
| ANOVA | ||||||
| P | ns | ** | ** | ** | ||
| N | ns | ** | ** | ** | ||
| P × N | ns | ** | ** | ** | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tong, J.; Xiong, Y.; Lu, Y.; Li, W.; Lin, W.; Xue, J.; Sun, M.; Wang, Y.; Gao, Z. Drip Fertigation Enhances the Responses of Grain Yield and Quality to Nitrogen Topdressing Rate in Irrigated Winter Wheat in North China. Plants 2024, 13, 1439. https://doi.org/10.3390/plants13111439
Tong J, Xiong Y, Lu Y, Li W, Lin W, Xue J, Sun M, Wang Y, Gao Z. Drip Fertigation Enhances the Responses of Grain Yield and Quality to Nitrogen Topdressing Rate in Irrigated Winter Wheat in North China. Plants. 2024; 13(11):1439. https://doi.org/10.3390/plants13111439
Chicago/Turabian StyleTong, Jin, Yulei Xiong, Yu Lu, Wen Li, Wen Lin, Jianfu Xue, Min Sun, Yuechao Wang, and Zhiqiang Gao. 2024. "Drip Fertigation Enhances the Responses of Grain Yield and Quality to Nitrogen Topdressing Rate in Irrigated Winter Wheat in North China" Plants 13, no. 11: 1439. https://doi.org/10.3390/plants13111439
APA StyleTong, J., Xiong, Y., Lu, Y., Li, W., Lin, W., Xue, J., Sun, M., Wang, Y., & Gao, Z. (2024). Drip Fertigation Enhances the Responses of Grain Yield and Quality to Nitrogen Topdressing Rate in Irrigated Winter Wheat in North China. Plants, 13(11), 1439. https://doi.org/10.3390/plants13111439