
Profiling the Bioactive Compounds in Broccoli Heads with Varying Organ Sizes and Growing Seasons

Abstract
1. Introduction
2. Results and Discussion
2.1. Soluble Soilds and Dry Matter Contents in Broccoli Heads Are Influenced by Growing Seasons
2.2. Vitamin C, Total Phenols, Carotenoids, and Individual Phenolic Compound Contents in Broccoli Heads Are Influenced by Growing Seasons and Organ Size
2.3. Glucoraphanin Content in Broccoli Heads Is Influenced by Growing Seasons and Organ Size
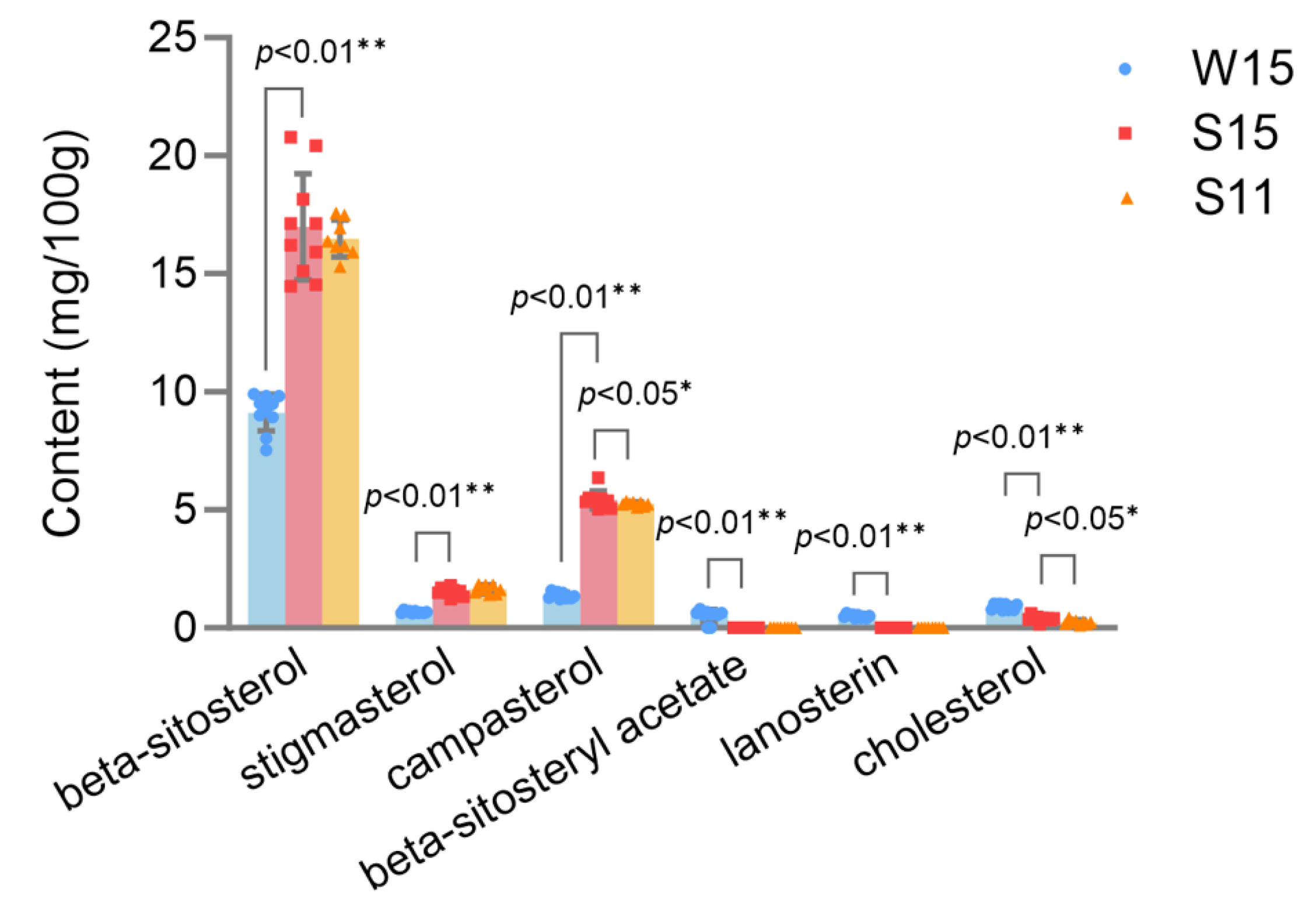
2.4. Phytosterol Profiles in Broccoli Heads Are Influenced by Growing Seasons and Organ Size
2.5. Mineral Contents in Broccoli Heads Are Influenced by Growing Seasons and Organ Size
3. Materials and Methods
3.1. Plant Materials
3.2. Sample Preparation and Determinations
3.2.1. Determination of Total Soluble Solids and Dry Matter Contents
3.2.2. Analysis of Vitamin C, Total Polyphenol, and Carotenoids
3.2.3. Analysis of Individual Phenolic and Glucosinolate Compounds by HPLC with Diode Array Detection (DAD)
3.2.4. Analysis of Individual Phytosterol Compound by HPLC-MS
3.2.5. Measurement of Mineral Content
3.2.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Martin, C.; Li, J. Medicine Is Not Health Care, Food Is Health Care: Plant Metabolic Engineering, Diet and Human Health. New Phytol. 2017, 216, 699–719. [Google Scholar] [CrossRef]
- Grosso, G.; Godos, J.; Currenti, W.; Micek, A.; Falzone, L.; Libra, M.; Giampieri, F.; Forbes-Hernández, T.Y.; Quiles, J.L.; Battino, M.; et al. The Effect of Dietary Polyphenols on Vascular Health and Hypertension: Current Evidence and Mechanisms of Action. Nutrients 2022, 14, 545. [Google Scholar] [CrossRef]
- Jeffery, E.H.; Brown, A.F.; Kurilich, A.C.; Keck, A.S.; Matusheski, N.; Klein, B.P.; Juvik, J.A. Variation in Content of Bioactive Components in Broccoli. J. Food Compos. Anal. 2003, 16, 323–330. [Google Scholar] [CrossRef]
- Shiva, R.B.; Jung-Ho, K. Seasonal Variation in Phytochemicals and Antioxidant Activities in Different Tissues of Various Broccoli Cultivars. Afr. J. Biotechnol. 2014, 13, 604–615. [Google Scholar] [CrossRef]
- Martínez-Castro, J.; De Haro-Bailón, A.; Obregón-Cano, S.; García Magdaleno, I.M.; Moreno Ortega, A.; Cámara-Martos, F. Bioaccessibility of Glucosinolates, Isothiocyanates and Inorganic Micronutrients in Cruciferous Vegetables through INFOGEST Static in Vitro Digestion Model. Food Res. Int. 2023, 166, 112598. [Google Scholar] [CrossRef]
- Almushayti, A.Y.; Brandt, K.; Carroll, M.A.; Scotter, M.J. Current Analytical Methods for Determination of Glucosinolates in Vegetables and Human Tissues. J. Chromatogr. A 2021, 1643, 462060. [Google Scholar] [CrossRef]
- Palliyaguru, D.L.; Yuan, J.; Kensler, T.W.; Fahey, J.W. Isothiocyanates: Translating the Power of Plants to People. Mol. Nutr. Food Res. 2018, 62, 1700965. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Rossi, M.; Liang, X.; Zhang, H.; Zou, L.; Ong, C.N. An Integrated Metabolomics Study of Glucosinolate Metabolism in Different Brassicaceae Genera. Metabolites 2020, 10, 313. [Google Scholar] [CrossRef] [PubMed]
- Oftadeh Harsin, A.; Firozian, F.; Soleimani, M.; Mehri, F.; Ranjbar, A. Evaluation of Ascorbic Acid Niosomes as Potential Detoxifiers in Oxidative Stress-Induced HEK-293 Cells by Arsenic Trioxide. Iran. J. Pharm. Res. 2022, 21, e127038. [Google Scholar] [CrossRef]
- Šalić, A.; Šamec, D. Changes in the Content of Glucosinolates, Polyphenols and Carotenoids during Lactic-Acid Fermentation of Cruciferous Vegetables: A Mini Review. Food Chem. X 2022, 16, 100457. [Google Scholar] [CrossRef]
- Bouafia, M. The Chemical Profile and Seasonal Variation of the Composition of the Phenolic Acids in Different Plant Parts of Centaurea sp. J. Pharm. Biomed. Anal. 2023, 236, 115686. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Goel, N. Phenolic Acids: Natural Versatile Molecules with Promising Therapeutic Applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef] [PubMed]
- Park, T.-K.; Kang, I.-A.; Park, C.-H.; Roh, J.; Lee, S.-H.; Kim, M.; Jin, E.; Kim, S.-K.; Kim, T.-W. Inhibition of 4-HYDROXYPHENYLPYRUVATE DIOXYGENASE Expression by Brassinosteroid Reduces Carotenoid Accumulation in Arabidopsis. J. Exp. Bot. 2022, 73, 1415–1428. [Google Scholar] [CrossRef] [PubMed]
- AbuMweis, S.S.; Marinangeli, C.P.F.; Frohlich, J.; Jones, P.J.H. Implementing Phytosterols into Medical Practice as a Cholesterol-Lowering Strategy: Overview of Efficacy, Effectiveness, and Safety. Can. J. Cardiol. 2014, 30, 1225–1232. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Yu, J.-W.; Song, M.-H.; Ahn, H.-Y.; Lee, J.-H.; Keum, Y.-S.; Lee, J.-H. Profiling of Redox-Active Lipophilic Constituents in Leaf Mustard (Brassica juncea (L.) Czern.) Cultivars Using LC-MS and GC-MS. Antioxidants 2022, 11, 2464. [Google Scholar] [CrossRef]
- Su, X.; Zheng, D.; Wang, M.; Zuo, Y.; Wen, J.; Zhai, Q.; Zhang, Y.; He, Y. Low Density Lipoprotein Cholesterol Is Associated with Increased Risk of Cardiovascular Disease in Participants over 70 Years Old: A Prospective Cohort Study. Nutr. Metab. Cardiovasc. Dis. 2022, 32, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Igl, W.; Kamal-Eldin, A.; Johansson, Å.; Liebisch, G.; Gnewuch, C.; Schmitz, G.; Gyllensten, U. Animal Source Food Intake and Association with Blood Cholesterol, Glycerophospholipids and Sphingolipids in a Northern Swedish Population. Int. J. Circumpolar Health 2013, 72, 21162. [Google Scholar] [CrossRef] [PubMed]
- Behrman, E.J.; Gopalan, V. Cholesterol and Plants. J. Chem. Educ. 2005, 82, 1791. [Google Scholar] [CrossRef]
- Šola, I.; Davosir, D.; Kokić, E.; Zekirovski, J. Effect of Hot- and Cold-Water Treatment on Broccoli Bioactive Compounds, Oxidative Stress Parameters and Biological Effects of Their Extracts. Plants 2023, 12, 1135. [Google Scholar] [CrossRef]
- Wermter, N.S.; Rohn, S.; Hanschen, F.S. Seasonal Variation of Glucosinolate Hydrolysis Products in Commercial White and Red Cabbages (Brassica oleracea Var. Capitata). Foods 2020, 9, 1682. [Google Scholar] [CrossRef]
- Wang, J.; Mao, S.; Wu, Q.; Yuan, Y.; Liang, M.; Wang, S.; Huang, K.; Wu, Q. Effects of LED Illumination Spectra on Glucosinolate and Sulforaphane Accumulation in Broccoli Seedlings. Food Chem. 2021, 356, 129550. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, G.; He, H.; Liu, Y.; Han, F.; Liu, W. Effects of Nanocarbon Solution Treatment on the Nutrients and Glucosinolate Metabolism in Broccoli. Food Chem. X 2022, 15, 100429. [Google Scholar] [CrossRef] [PubMed]
- Palmer, J.W.; Harker, F.R.; Tustin, D.S.; Johnston, J. Fruit Dry Matter Concentration: A New Quality Metric for Apples: Apple Fruit Dry Matter Concentration. J. Sci. Food Agric. 2010, 90, 2586–2594. [Google Scholar] [CrossRef]
- Valverde-Miranda, D.; Díaz-Pérez, M.; Gómez-Galán, M.; Callejón-Ferre, Á.-J. Total Soluble Solids and Dry Matter of Cucumber as Indicators of Shelf Life. Postharvest Biol. Technol. 2021, 180, 111603. [Google Scholar] [CrossRef]
- Gajc-Wolska, J.; Kowalczyk, K.; Bujalski, D. The Effect of Cultivation Term, Substrate and Cultivar on Chemical Composition of Cucumber Fruit (Cucumis sativus L.) in Greenhouse Production. Acta Hortic. 2010, 877, 239–244. [Google Scholar] [CrossRef]
- Ikeda, T.; Yamazaki, K.; Kumakura, H.; Hamamoto, H. Effect of High Temperature on Fruit Quality of Pot-Grown Strawberry Plants. Acta Hortic. 2009, 842, 679–682. [Google Scholar] [CrossRef]
- Menzel, C.M. Effect of Temperature on Soluble Solids Sontent in Strawberry in Queensland, Australia. Horticulturae 2022, 8, 367. [Google Scholar] [CrossRef]
- Rao, A.; Rao, L. Carotenoids and Human Health. Pharmacol. Res. 2007, 55, 207–216. [Google Scholar] [CrossRef]
- Fiedor, J.; Burda, K. Potential Role of Carotenoids as Antioxidants in Human Health and Disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef]
- Carr, A.; Maggini, S. Vitamin C and Immune Function. Nutrients 2017, 9, 1211. [Google Scholar] [CrossRef]
- Pullar, J.; Carr, A.; Vissers, M. The Roles of Vitamin C in Skin Health. Nutrients 2017, 9, 866. [Google Scholar] [CrossRef]
- Liu, X.; Wu, R.; Bulley, S.M.; Zhong, C.; Li, D. Kiwifruit MYBS1-like and GBF3 Transcription Factors Influence l -ascorbic Acid Biosynthesis by Activating Transcription of GDP-L-galactose Phosphorylase 3. New Phytol. 2022, 234, 1782–1800. [Google Scholar] [CrossRef]
- Xiang, N.; Hu, J.; Wen, T.; Brennan, M.A.; Brennan, C.S.; Guo, X. Effects of Temperature Stress on the Accumulation of Ascorbic Acid and Folates in Sweet Corn ( Zea mays L.) Seedlings. J. Sci. Food Agric. 2020, 100, 1694–1701. [Google Scholar] [CrossRef]
- Quian-Ulloa, R.; Stange, C. Carotenoid Biosynthesis and Plastid Development in Plants: The Role of Light. Int. J. Mol. Sci. 2021, 22, 1184. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Singh, S.K.; Pattanaik, S.; Wang, H.; Yuan, L. Light Regulation of the Biosynthesis of Phenolics, Terpenoids, and Alkaloids in Plants. Commun. Biol. 2023, 6, 1055. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kong, D.; Fu, Y.; Sussman, M.R.; Wu, H. The Effect of Developmental and Environmental Factors on Secondary Metabolites in Medicinal Plants. Plant Physiol. Biochem. 2020, 148, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Cai, W.; Wu, R.; Huang, Y.; Lu, Q.; Wang, H.; Huang, X.; Zhang, Y.; Wu, Q.; Cheng, X.; et al. Differential CaKAN3-CaHSF8 Associations Underlie Distinct Immune and Heat Responses under High Temperature and High Humidity Conditions. Nat. Commun. 2023, 14, 4477. [Google Scholar] [CrossRef]
- Bento-Silva, A.; Patto, M.C.V.; do Rosário Bronze, M. Relevance, Structure and Analysis of Ferulic Acid in Maize Cell Walls. Food Chem. 2018, 246, 360–378. [Google Scholar] [CrossRef]
- Singh, P.; Arif, Y.; Bajguz, A.; Hayat, S. The Role of Quercetin in Plants. Plant Physiol. Biochem. 2021, 166, 10–19. [Google Scholar] [CrossRef]
- Mnich, E.; Bjarnholt, N.; Eudes, A.; Harholt, J.; Holland, C.; Jørgensen, B.; Larsen, F.H.; Liu, M.; Manat, R.; Meyer, A.S.; et al. Phenolic Cross-Links: Building and de-Constructing the Plant Cell Wall. Nat. Prod. Rep. 2020, 37, 919–961. [Google Scholar] [CrossRef]
- Nuñez-Gómez, V.; Baenas, N.; Navarro-González, I.; García-Alonso, J.; Moreno, D.A.; González-Barrio, R.; Periago-Castón, M.J. Seasonal Variation of Health-Promoting Bioactives in Broccoli and Methyl-Jasmonate Pre-Harvest Treatments to Enhance Their Contents. Foods 2020, 9, 1371. [Google Scholar] [CrossRef]
- Chae, S.-H.; Lee, O.N.; Park, H.Y.; Ku, K.-M. Seasonal Effects of Glucosinolate and Sugar Content Determine the Pungency of Small-Type (Altari) Radishes (Raphanus sativus L.). Plants 2022, 11, 312. [Google Scholar] [CrossRef]
- Groenbaek, M.; Kidmose, U.; Tybirk, E.; Kristensen, H.L. Glucosinolate Content and Sensory Evaluation of Baby Leaf Rapeseed from Annual and Biennial White- and Yellow-flowering Cultivars with Repeated Harvesting in Two Seasons. J. Food Sci. 2019, 84, 1888–1899. [Google Scholar] [CrossRef] [PubMed]
- Jasper, J.; Wagstaff, C.; Bell, L. Growth Temperature Influences Postharvest Glucosinolate Concentrations and Hydrolysis Product Formation in First and Second Cuts of Rocket Salad. Postharvest Biol. Technol. 2020, 163, 111157. [Google Scholar] [CrossRef] [PubMed]
- Oloyede, O.O.; Wagstaff, C.; Methven, L. Influence of Cabbage (Brassica oleracea) Accession and Growing Conditions on Myrosinase Activity, Glucosinolates and Their Hydrolysis Products. Foods 2021, 10, 2903. [Google Scholar] [CrossRef] [PubMed]
- Rosa, E.A.S.; Rodrigues, P.M.F. The Effect of Light and Temperature on Glucosinolate Concentration in the Leaves and Roots of Cabbage Seedlings. J. Sci. Food Agric. 1998, 78, 208–212. [Google Scholar] [CrossRef]
- Del Carmen Martínez-Ballesta, M.; Moreno, D.; Carvajal, M. The Physiological Importance of Glucosinolates on Plant Response to Abiotic Stress in Brassica. Int. J. Mol. Sci. 2013, 14, 11607–11625. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yang, J.; Zhang, J.; Liu, Y.; Tian, C.; Qu, B.; Gao, C.; Xin, P.; Cheng, S.; Zhang, W.; et al. An Arabidopsis Secondary Metabolite Directly Targets Expression of the Bacterial Type III Secretion System to Inhibit Bacterial Virulence. Cell Host Microbe 2020, 27, 601–613. [Google Scholar] [CrossRef]
- Zangenberg, M.; Hansen, H.B.; Jørgensen, J.R.; Hellgren, L.I. Cultivar and Year-to-Year Variation of Phytosterol Content in Rye (Secale cereale L.). J. Agric. Food Chem. 2004, 52, 2593–2597. [Google Scholar] [CrossRef]
- Moreau, R.A.; Nyström, L.; Whitaker, B.D.; Winkler-Moser, J.K.; Baer, D.J.; Gebauer, S.K.; Hicks, K.B. Phytosterols and Their Derivatives: Structural Diversity, Distribution, Metabolism, Analysis, and Health-Promoting Uses. Prog. Lipid Res. 2018, 70, 35–61. [Google Scholar] [CrossRef]
- Weyh, C.; Krüger, K.; Peeling, P.; Castell, L. The Role of Minerals in the Optimal Functioning of the Immune System. Nutrients 2022, 14, 644. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Salcedo, E.M.; Mena, P.; García-Viguera, C.; Martínez, J.J.; Hernández, F. Phytochemical Evaluation of White (Morus alba L.) and Black (Morus nigra L.) Mulberry Fruits, a Starting Point for the Assessment of Their Beneficial Properties. J. Funct. Foods 2015, 12, 399–408. [Google Scholar] [CrossRef]
- Melse-Boonstra, A. Bioavailability of Micronutrients from Nutrient-Dense Whole Foods: Zooming in on Dairy, Vegetables, and Fruits. Front. Nutr. 2020, 7, 101. [Google Scholar] [CrossRef] [PubMed]
- Liu, D. Root Developmental Responses to Phosphorus Nutrition. J. Integr. Plant Biol. 2021, 63, 1065–1090. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.S. (Ed.) Food Analysis Laboratory Manual; Food Science Texts Series; Springer US: Boston, MA, USA, 2010; ISBN 978-1-4419-1462-0. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. [14] Analysis of Total Phenols and Other Oxidation Substrates and Antioxidants by Means of Folin-Ciocalteu Reagent. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1999; Volume 299, pp. 152–178. ISBN 978-0-12-182200-2. [Google Scholar]
- Zhou, F.; Jiang, A.; Feng, K.; Gu, S.; Xu, D.; Hu, W. Effect of Methyl Jasmonate on Wound Healing and Resistance in Fresh-Cut Potato Cubes. Postharvest Biol. Technol. 2019, 157, 110958. [Google Scholar] [CrossRef]
- Carvalho, L.; Gomes, P.; de Oliveira Godoy, R.; Pacheco, S.; Monte, P.; Carvalho, J.; Nutti, M.; Neves, A.; Vieira, A.; Ramos, S. Total Carotenoid Content, α-Carotene and β-Carotene, of Landrace Pumpkins (Cucurbita moschata Duch): A Preliminary Study. Food Res. Int. 2012, 47, 337–340. [Google Scholar] [CrossRef]
- Becerra-Moreno, A.; Benavides, J.; Cisneros-Zevallos, L.; Jacobo-Velázquez, D.A. Plants as Biofactories: Glyphosate-Induced Production of Shikimic Acid and Phenolic Antioxidants in Wounded Carrot Tissue. J. Agric. Food Chem. 2012, 60, 11378–11386. [Google Scholar] [CrossRef] [PubMed]
- Baenas, N.; García-Viguera, C.; Moreno, D.A. Biotic Elicitors Effectively Increase the Glucosinolates Content in Brassicaceae Sprouts. J. Agric. Food Chem. 2014, 62, 1881–1889. [Google Scholar] [CrossRef]
- Guan, Y.; Hu, W.; Jiang, A.; Xu, Y.; Sa, R.; Feng, K.; Zhao, M.; Yu, J.; Ji, Y.; Hou, M.; et al. Effect of Methyl Jasmonate on Phenolic Accumulation in Wounded Broccoli. Molecules 2019, 24, 3537. [Google Scholar] [CrossRef]
- Nor, N.D.M.; Lignou, S.; Bell, L.; Houston-Price, C.; Harvey, K.; Methven, L. The Relationship between Glucosinolates and the Sensory Characteristics of Steamed-Pureed Turnip (Brassica rapa Subsp. Rapa L.). Foods 2020, 9, 1719. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Hwang, K.T. Changes in Physicochemical Properties of Mulberry Fruits (Morus alba L.) during Ripening. Sci. Hortic. 2017, 217, 189–196. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Samples | TSS (%) | Dry Matter Contents (%) |
|---|---|---|
| W15 | 7.50 ± 0.47 | 11.88 ± 0.75 |
| S15 | 6.13 ± 0.33 | 11.13 ± 0.65 |
| S11 | 6.44± 0.26 | 11.56 ± 0.68 |
| W15 vs. S15 | p < 0.01 | p < 0.05 |
| S15 vs. S11 | p < 0.05 | n.s. |
| Samples | Individual Phenolic Content (μg/g FW) | |||||
|---|---|---|---|---|---|---|
| Ferulic Acid | Catechin | Quercetin | Caffeic Acid | Chlorogenic Acid | Rutin | |
| W15 | 19.53 ± 1.28 | 50.64 ± 10.93 | 21.56 ± 6.83 | 6.70 ± 1.25 | 150.29 ± 7.21 | 117.27 ± 3.82 |
| S15 | 23.20 ± 0.77 | 54.68 ± 2.43 | 24.89 ± 4.72 | 9.45 ± 2.02 | 152.03 ± 9.04 | 83.85 ± 11.71 |
| S11 | 34.01 ± 0.36 | 71.76 ± 15.41 | 44.60 ± 15.37 | 8.02 ± 1.29 | 130.66 ± 2.69 | 107.89 ± 2.04 |
| W15 vs. S15 | p < 0.05 | n.s. | n.s. | n.s. | n.s. | p < 0.01 |
| S15 vs. S11 | p < 0.01 | n.s. | n.s. | n.s. | p < 0.05 | p < 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, L.; Li, Y.; Lin, M.; Liang, Y.; Zhang, Z. Profiling the Bioactive Compounds in Broccoli Heads with Varying Organ Sizes and Growing Seasons. Plants 2024, 13, 1329. https://doi.org/10.3390/plants13101329
Shi L, Li Y, Lin M, Liang Y, Zhang Z. Profiling the Bioactive Compounds in Broccoli Heads with Varying Organ Sizes and Growing Seasons. Plants. 2024; 13(10):1329. https://doi.org/10.3390/plants13101329
Chicago/Turabian StyleShi, Lu, Yahui Li, Menghua Lin, Ying Liang, and Zhiyong Zhang. 2024. "Profiling the Bioactive Compounds in Broccoli Heads with Varying Organ Sizes and Growing Seasons" Plants 13, no. 10: 1329. https://doi.org/10.3390/plants13101329
APA StyleShi, L., Li, Y., Lin, M., Liang, Y., & Zhang, Z. (2024). Profiling the Bioactive Compounds in Broccoli Heads with Varying Organ Sizes and Growing Seasons. Plants, 13(10), 1329. https://doi.org/10.3390/plants13101329