
Effects of Cesium on Physiological Traits of the Catherine’s Moss Atrichum undulatum Hedw.


 ,
,  ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Results
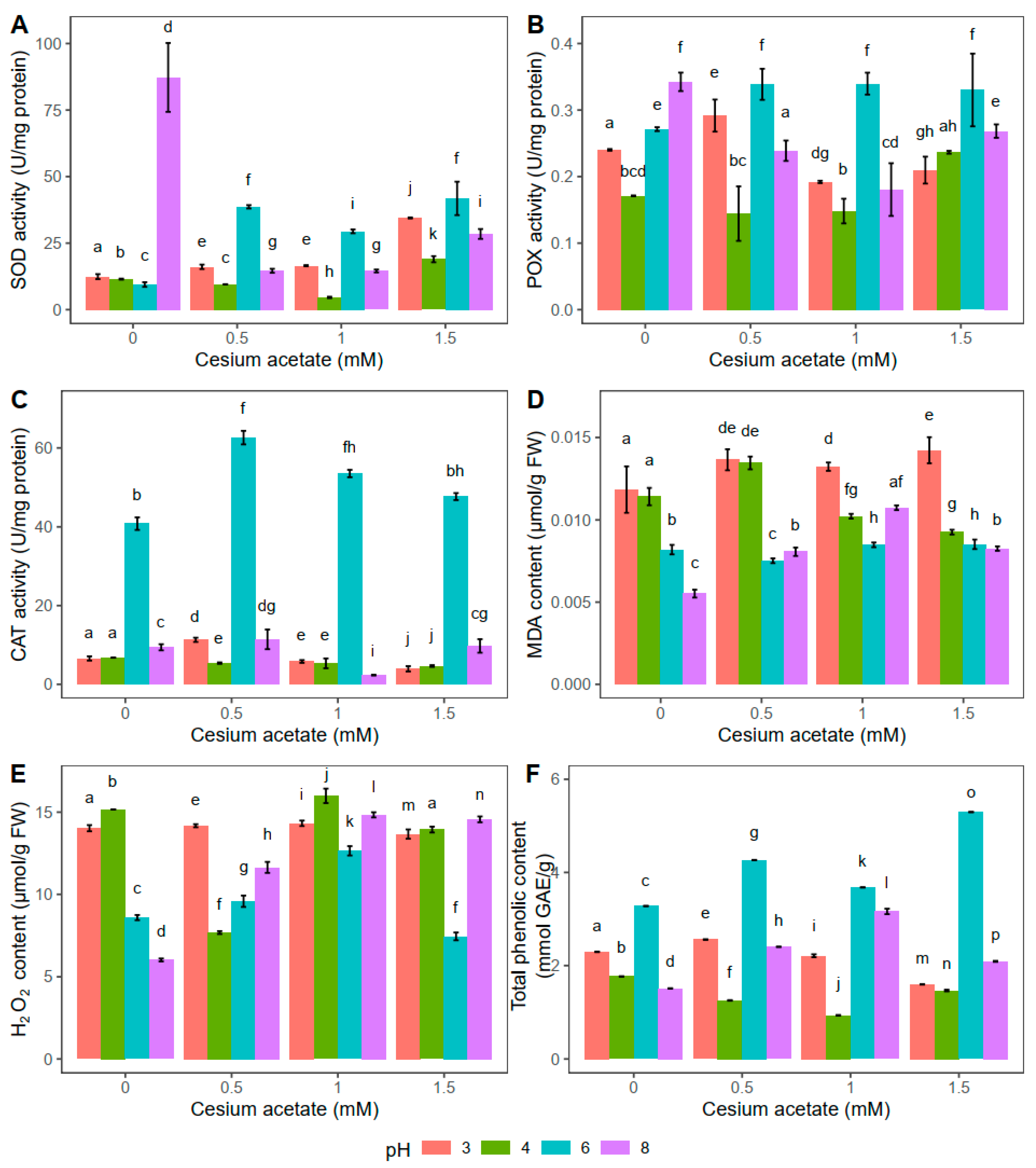
2.1. Effects of Long-Term Exposure to Cesium Acetate in Experiment I (Uptake from the Solid Substrate)
2.2. Effect of Short-Term Exposure for 2 h to Cesium Acetate in Experiment II (Light Rain Simulation)
2.3. Effect of Short-Term Exposure for 24 h to Cesium Acetate in Experiment II (Heavy Rain Simulation)
3. Discussion
4. Materials and Methods
4.1. Plant Material and Culture Conditions
4.2. Experimental Design
4.3. Determination of Antioxidant Enzyme Activities
4.3.1. Tissue Extract Preparation and Protein Content
4.3.2. Analytical Assays of Antioxidant Enzyme Activities
4.3.3. Determination of Malondialdehyde and Hydrogen Peroxide Concentrations
4.3.4. Spectrophotometric Analysis of Total Phenolic Content (Folin–Ciocalteu Test)
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Rühling, A.; Tyler, G. Ecological approach to the lead problem. Bot. Not. 1968, 121, 321–342. [Google Scholar]
- Shaw, A.J.; Szovenyi, P.; Shaw, B. Bryophyte diversity and evolution: Windows into the early evolution of land plants. Am. J. Bot. 2011, 98, 352–369. [Google Scholar] [CrossRef] [PubMed]
- Tyler, G. Bryophytes and heavy metals: A literature review. Bot. J. Linn. Soc. 1990, 104, 231–253. [Google Scholar] [CrossRef]
- Wolterbeek, B. Biomonitoring of trace element air pollution: Principles, possibilities and perspectives. Environ. Pollut. 2002, 120, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Szczepaniak, K.; Biziuk, M. Aspects of the biomonitoring studies using mosses and lichens as indicators of metal pollution. Environ. Res. 2003, 93, 221–230. [Google Scholar] [CrossRef] [PubMed]
- Petschinger, K.; Adlassnig, W.; Sabovljevic, M.; Lang, I. Lamina cell shape and cell wall thickness are useful indicators for metal tolerance—An example in bryophytes. Plants 2021, 10, 274. [Google Scholar] [CrossRef] [PubMed]
- Wattanavatee, K.; Krmar, M.; Bhongsuwan, T. A survey of natural terrestrial and airborne radionuclides in moss samples from the peninsular Thailand. J. Environ. Radioact. 2017, 177, 113–127. [Google Scholar] [CrossRef]
- Vitt, D.; Crandall-Stotler, B.; Wood, A. Bryophytes: Survival in a dry world through tolerance and avoidance. In Plant Ecology and Evolution in Harsh Environments; Rajakaruna, N., Boyd, R., Harris, T., Eds.; Nova Publishers: Hauppauge, NY, USA, 2014; pp. 267–295. ISBN 978-1-63321-955-7. [Google Scholar]
- Tessier, L.; Boisvert, J.L. Performance of terrestrial bryophytes as biomonitors of atmospheric pollution. A review. Toxicol. Envuron. Chem. 1999, 68, 179–220. [Google Scholar] [CrossRef]
- González, A.G.; Pokrovsky, O.S. Metal adsorption on mosses: Toward a universal adsorption model. J. Colloid Interface Sci. 2014, 415, 169–178. [Google Scholar] [CrossRef]
- Gorelova, S. Trace element accumulating ability of different moss species used to study atmospheric deposition of heavy metals in Central Russia: Tula region case study. Int. J. Biol. Biomed. Eng. 2016, 10, 271–285. [Google Scholar]
- Sabovljević, A.; Vujičić, M.; Stanković, J.; Sabovljević, M. Effects of zinc and copper on development and survival of the moss Atrichum undulatum in controlled conditions. Bot. Serbica 2018, 42, 181–184. [Google Scholar] [CrossRef]
- Sabovljević, M.; Weidinger, M.; Sabovljević, A.; Stanković, J.; Adlassnig, W.; Lang, I. Metal accumulation in the acrocarp moss Atrichum undulatum under controlled conditions. Environ. Pollut. 2020, 256, 113397. [Google Scholar] [CrossRef] [PubMed]
- Stankovic, J.; Janković, S.; Lang, I.; Vujičić, M.; Sabovljević, M.; Sabovljević, A. The toxic metal stress in two mosses of different growth forms under axenic and controlled conditions. Bot. Serbica 2021, 45, 31–47. [Google Scholar] [CrossRef]
- ATSDR. Toxicolgical Profile for Cesium, US Department of Health and Human Services. 2004. Available online: http://www.atsdr.cdc.gov/toxprofiles/tp.asp?id=578&tid=107 (accessed on 21 September 2023).
- Hu, Q.H.; Weng, J.Q.; Wang, J.S. Sources of anthropogenic radionuclides in the environment: A review. J. Environ. Radioact. 2010, 101, 426–437. [Google Scholar] [CrossRef] [PubMed]
- Saniewski, M.; Wietrzyk-Pełka, P.; Zalewska, T.; Olech, M.; Węgrzyn, M.H. Bryophytes and lichens as fallout originated radionuclide indicators in the Svalbard archipelago (High Arctic). Polar Sci. 2020, 25, 100536. [Google Scholar] [CrossRef]
- Burger, A.; Weidinger, M.; Adlassnig, W.; Puschenreiter, M.; Lichtscheidl, I. Response of Arabidopsis halleri to cesium and strontium in hydroponics: Extraction potential and effects on morphology and physiology. Ecotoxicol. Environ. Saf. 2019, 184, 109625. [Google Scholar] [CrossRef]
- Zhu, Y.; Smolders, E. Plant uptake of radiocaesium: A review of mechanisms, regulation and application. J. Exp. Bot. 2000, 51, 635–1645. [Google Scholar] [CrossRef]
- Hampton, C.R.; Bowen, H.C.; Broadley, M.R.; Hammond, J.P.; Mead, A.; Payne, K.A.; Pritchard, J.; White, P.J. Cesium toxicity in Arabidopsis. Plant Physiol. 2004, 136, 3824–3837. [Google Scholar] [CrossRef]
- Kanter, U.; Hauser, A.; Michalke, B.; Dräxl, S.; Schäffner, A.R. Caesium and strontium accumulation in shoots of Arabidopsis thaliana: Genetic and physiological aspects. J. Exp. Bot. 2010, 61, 3995–4009. [Google Scholar] [CrossRef]
- Atapaththu, K.S.S.; Rashid, M.H.; Asaeda, T. Growth and oxidative stress of Brittlewort (Nitella pseudoflabellata) in response to cesium exposure. Bull. Environ. Contam. Toxicol. 2016, 96, 347–353. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, G.J. Effects of cesium accumulation on chlorophyll content and fluorescence of Brassica juncea L. J. Environ. Radioact. 2018, 195, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Bystrzejewska-Piotrowska, G.; Urban, P.Ł. Accumulation of cesium in leaves of Lepidium sativum and its influence on photosynthesis and transpiration. Acta Biol. Crac. 2003, 45, 131–137. [Google Scholar]
- Chen, M.; Tang, Y.L.; Bing, A.N.; Wang, D. Effects of cesium on photosynthetic characteristics of wheat seedlings. J. Anhui Univ. 2012, 36, 103–108. [Google Scholar] [CrossRef]
- Li, H.; Tang, Y.J.; Zeng, F. Effects of high concentration of strontium and cesium on chlorophyll fluorescence characteristics of plants. Jiangsu Agric. Sci. 2013, 41, 349–352. [Google Scholar]
- Jing, X.U.; Tang, Y.; Wang, J.; Wang, D. Study on the effects of cesium on photosynthesis of spinach. J. Nucl. Agric. Sci. 2015, 181, 123–130. [Google Scholar]
- Cheng, X.; Chen, C.; Hu, Y.; Wang, J. Response of Amaranthus tricolor to cesium stress in hydroponic system: Growth, photosynthesis and cesium accumulation. Chemosphere 2022, 307, 135754. [Google Scholar] [CrossRef] [PubMed]
- Ivshina, I.B.; Peshkur, T.A.; Korobov, V.P. Efficient uptake of cesium ions by Rhodococcus cells. Microbiology 2002, 71, 357–361. [Google Scholar] [CrossRef]
- Bhaduri, A.M.; Fulekar, M.H. Antioxidant enzyme responses of plants to heavy metal stress. Rev. Environ. Sci. Biotechnol. 2012, 11, 55–69. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Alegre, L. The function of tocopherols and tocotrienols in plants. Crit. Rev. Plant Sci. 2002, 21, 31–57. [Google Scholar] [CrossRef]
- Guo, T.; Zhang, G.; Zhou, M.; Wu, F.; Chen, J. Effects of aluminum and cadmium toxicity on growth and antioxidant enzyme activities of two barley genotypes with different Al resistance. Plant Soil 2004, 258, 241–248. [Google Scholar] [CrossRef]
- Sun, S.Q.; He, M.; Cao, T.; Zhang, Y.C.; Han, W. Response mechanisms of antioxidants in bryophyte (Hypnum plumaeforme) under the stress of single or combined Pb and/or Ni. Environ. Monit. Assess. 2009, 149, 291–302. [Google Scholar] [CrossRef]
- Nikalje, G.C.; Shrivastava, M.; Nikam, T.D.; Suprasanna, P. Physiological Responses and Tolerance of Halophyte Sesuvium portulacastrum L. to Cesium. Adv. Agric. 2022, 2022, 9863002. [Google Scholar] [CrossRef]
- Lai, J.L.; Luo, X.G. High-efficiency antioxidant system, chelating system and stress-responsive genes enhance tolerance to cesium iontoxicity in Indian mustard (Brassica juncea L.). Ecotoxicol. Environ. Saf. 2019, 181, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Morales, M.; Munné-Bosch, S. Malondialdehyde: Facts and artifacts. Plant Physiol. 2019, 180, 1246–1250. [Google Scholar] [CrossRef]
- Panda, S.K.; Choudhury, S. Changes in nitrate reductase activity and oxidative stress response in the moss Polytrichum commune subjected to chromium, copper and zinc phytotoxicity. Braz. J. Plant Physiol. 2005, 17, 191–197. [Google Scholar] [CrossRef]
- Hossain, M.A.; Bhattacharjee, S.; Armin, S.M.; Qian, P.; Xin, W.; Li, H.Y.; Burritt, D.J.; Fujita, M.; Tran, L.S.P. Hydrogen peroxide priming modulates abiotic oxidative stress tolerance: Insights from ROS detoxification and scavenging. Front. Plant Sci. 2015, 6, 420. [Google Scholar] [CrossRef] [PubMed]
- Stanković, J.; Sabovljević, A.; Sabovljević, M. Bryophytes and heavy metals: A review. Acta Bot. Croat. 2018, 77, 109–118. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef]
- Mashkani, S.G.; Ghazvini, P.T.M. Biotechnological potential of Azolla filiculoides for biosorption of Cs and Sr: Application of micro-PIXE for measurement of biosorption. Bioresour. Technol. 2009, 100, 1915–1921. [Google Scholar] [CrossRef]
- Wang, X.; Rabung, T.; Geckeis, H. Effect of pH and humic acid on the adsorption of cesium onto g-Al2O3. J. Radioanal. Nucl. Chem. 2003, 258, 83–87. [Google Scholar] [CrossRef]
- Giannakopoulou, F.; Haidouti, C.; Chronopoulou, A.; Gasparatos, D. Sorption behavior of cesium on various soils under different pH levels. J. Hazard. Mater. 2007, 149, 553–556. [Google Scholar] [CrossRef] [PubMed]
- Saleh, H.M.; Moussa, H.R.; Mahmoud, H.H.; El-Saied, F.A.; Dawoud, M.; Wahed, R.S.A. Potential of the submerged plant Myriophyllum spicatum for treatment of aquatic environments contaminated with stable or radioactive cobalt and cesium. Prog. Nucl. Energy 2020, 118, 103147. [Google Scholar] [CrossRef]
- Sabovljević, A.; Cvetić, T.; Sabovljević, M. The establishment and development of the Catherine’s moss Atrichum undulatum (Hedw.) P. Beauv. (Polytrichaceae) in in vitro conditions. Arch. Biol. Sci. 2006, 58, 87–93. [Google Scholar] [CrossRef]
- Sabovljević, M.; Vujičić, M.; Živković, S.; Nerić, V.; Šinžar-Sekulić, J.; Lang, I.; Sabovljević, A. Genetic diversity within selected European populations of the moss species Atrichum undulatum as inferred by the isozymes. Wulfenia 2019, 26, 208–216. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Beyer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photooxidation in isolation chloroplast I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–194. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- R Core Team, R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 25 December 2022).
- Elkin, L.; Kay, M.; Higgins, J.; Wobbrock, J. An Aligned Rank Transform Procedure for Multifactor Contrast Tests. In Proceedings of the 34th Annual ACM Symposium on User Interface Software and Technology, Association for Computing Machinery, New York, NY, USA, 10–14 October 2021. [Google Scholar] [CrossRef]
- Wobbrock, J.; Findlater, L.; Gergle, D.; Higgins, J. The Aligned Rank Transform for Nonparametric Factorial Analyses Using Only ANOVA Procedures. In Proceedings of the ACM Conference on Human Factors in Computing Systems (CHI 2011), Vancouver, BC, Canada, 7–12 May 2011; ACM Press: New York, NY, USA, 2011; pp. 143–146. [Google Scholar] [CrossRef]
- Kay, M.; Elkin, L.; Higgins, J.; Wobbrock, J. mjskay/ARTool: ARTool 0.11.0 (v0.11.0). Zenodo. 2021. Available online: https://zenodo.org/records/4721941 (accessed on 20 December 2022).



{kind=link}
{kind=link}
{kind=link}
| CAT | POX | SOD | TPC | MDA | H2O2 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variable | 2 h | 24 h | 2 h | 24 h | 2 h | 24 h | 2 h | 24 h | 2 h | 24 h | 2 h | 24 h |
| C | 39.56 * | 156.46 * | 14.92 * | 37.99 * | 80.76 * | 36.29 * | 168.76 * | 401.48 * | 40.11 * | 35.78 * | 155.62 * | 198.15 * |
| pH | 114.27 * | 323.23 * | 138.79 * | 34.61 * | 162.21 * | 159.03 * | 402.27 * | 401.44 * | 117.01 * | 209.62 * | 324.74 * | 334.81 * |
| C × pH | 88.44 * | 223.95 * | 21.49 * | 32.53 * | 117.60 * | 224.35 * | 1296.86 * | 1093.28 * | 51.62 * | 172.16 * | 159.21 * | 437.98 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stanojković, J.N.; Ćosić, M.V.; Božović, D.P.; Sabovljević, A.D.; Sabovljević, M.S.; Čučulović, A.A.; Vujičić, M.M. Effects of Cesium on Physiological Traits of the Catherine’s Moss Atrichum undulatum Hedw. Plants 2024, 13, 54. https://doi.org/10.3390/plants13010054
Stanojković JN, Ćosić MV, Božović DP, Sabovljević AD, Sabovljević MS, Čučulović AA, Vujičić MM. Effects of Cesium on Physiological Traits of the Catherine’s Moss Atrichum undulatum Hedw. Plants. 2024; 13(1):54. https://doi.org/10.3390/plants13010054
Chicago/Turabian StyleStanojković, Jelena N., Marija V. Ćosić, Djordje P. Božović, Aneta D. Sabovljević, Marko S. Sabovljević, Ana A. Čučulović, and Milorad M. Vujičić. 2024. "Effects of Cesium on Physiological Traits of the Catherine’s Moss Atrichum undulatum Hedw." Plants 13, no. 1: 54. https://doi.org/10.3390/plants13010054
APA StyleStanojković, J. N., Ćosić, M. V., Božović, D. P., Sabovljević, A. D., Sabovljević, M. S., Čučulović, A. A., & Vujičić, M. M. (2024). Effects of Cesium on Physiological Traits of the Catherine’s Moss Atrichum undulatum Hedw. Plants, 13(1), 54. https://doi.org/10.3390/plants13010054