

Analysis of the Complete Mitochondrial Genome of the Bitter Gourd (Momordica charantia)
 and
and
Abstract
1. Introduction
2. Results
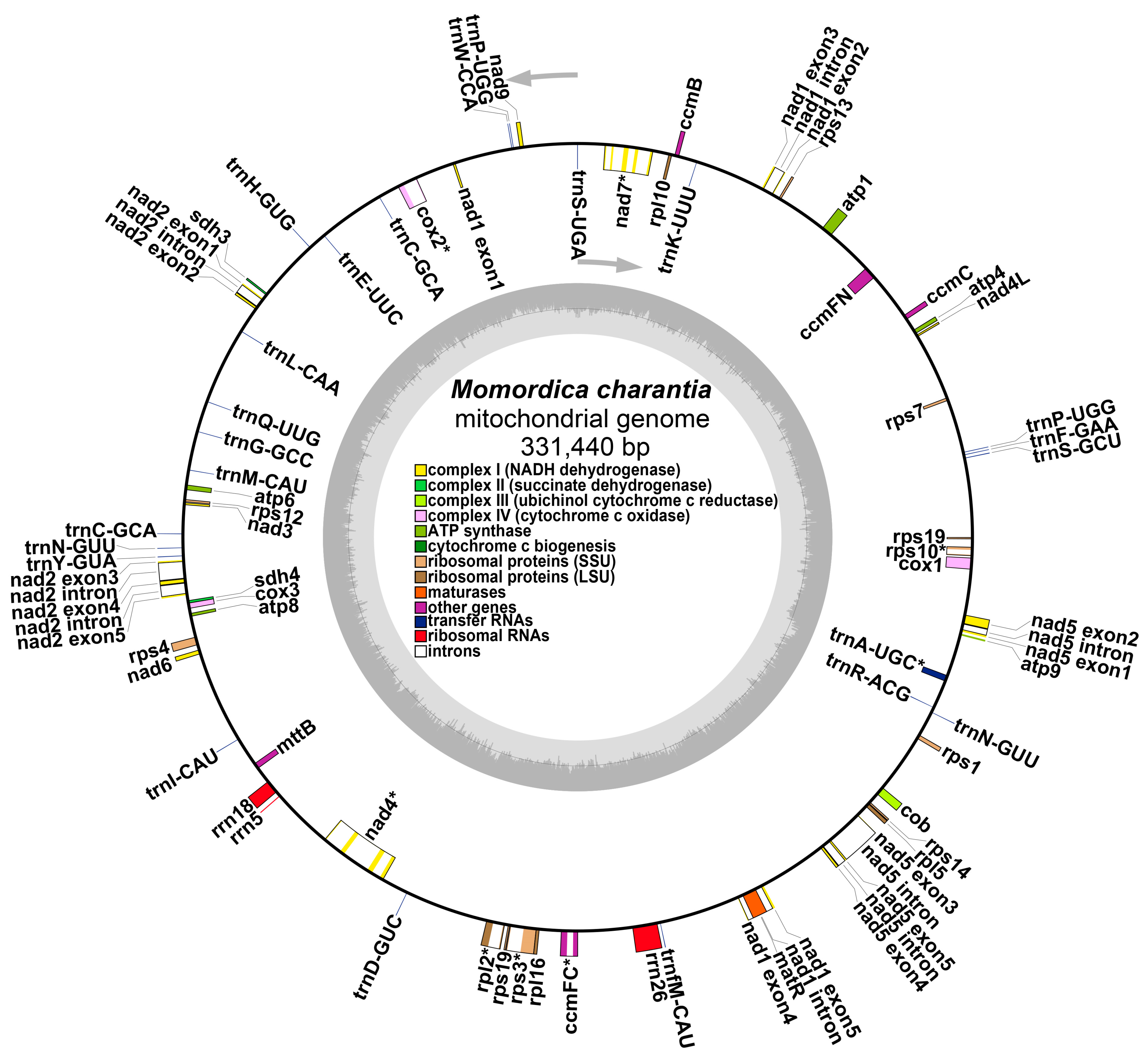
2.1. Mitochondrial Genome Assembly and Annotation of Bitter Gourd
2.2. The Structure and Codon Preference of the Mitochondrial Genome
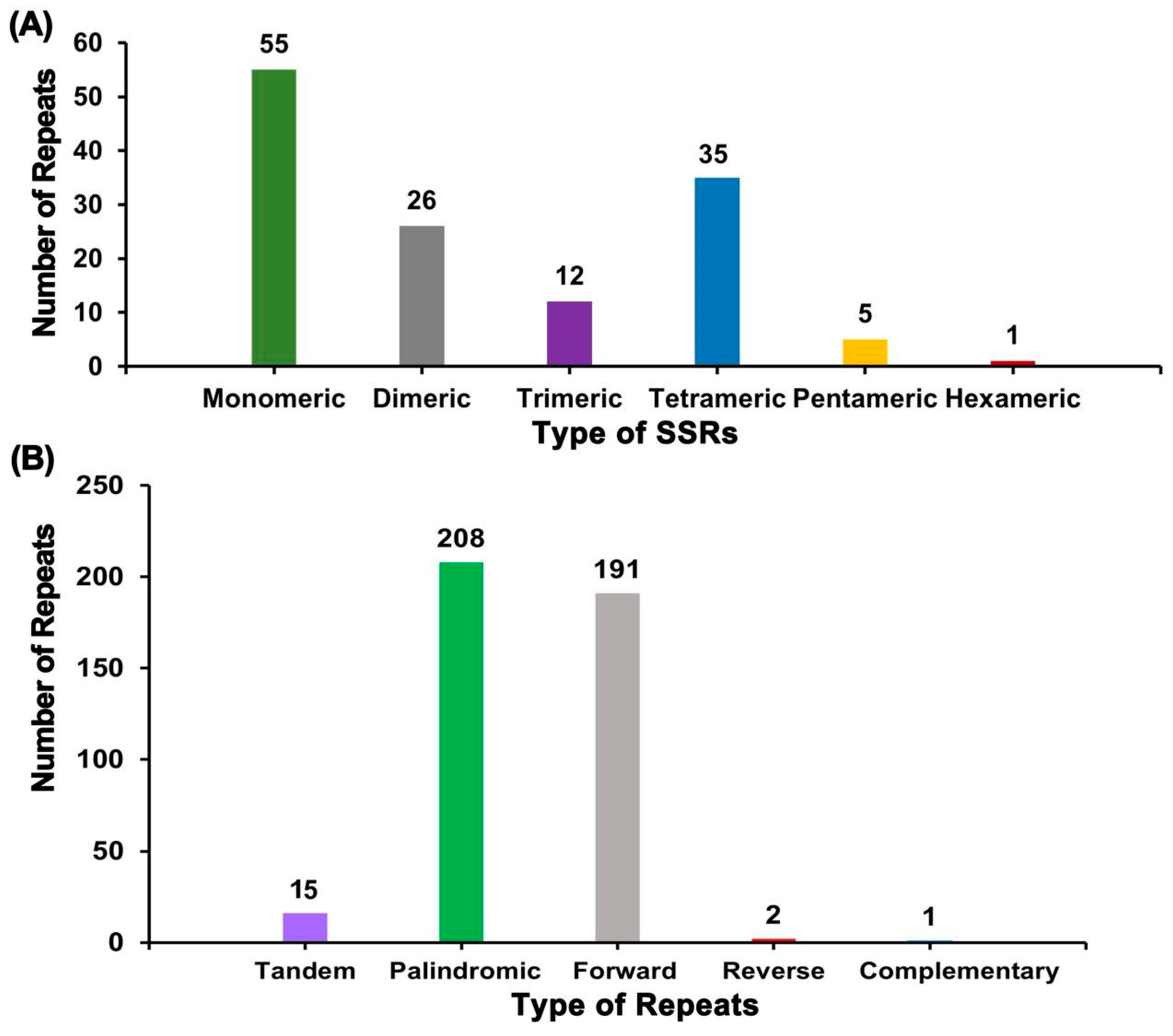
2.3. Repeat Elements and DNA Transfer Analysis
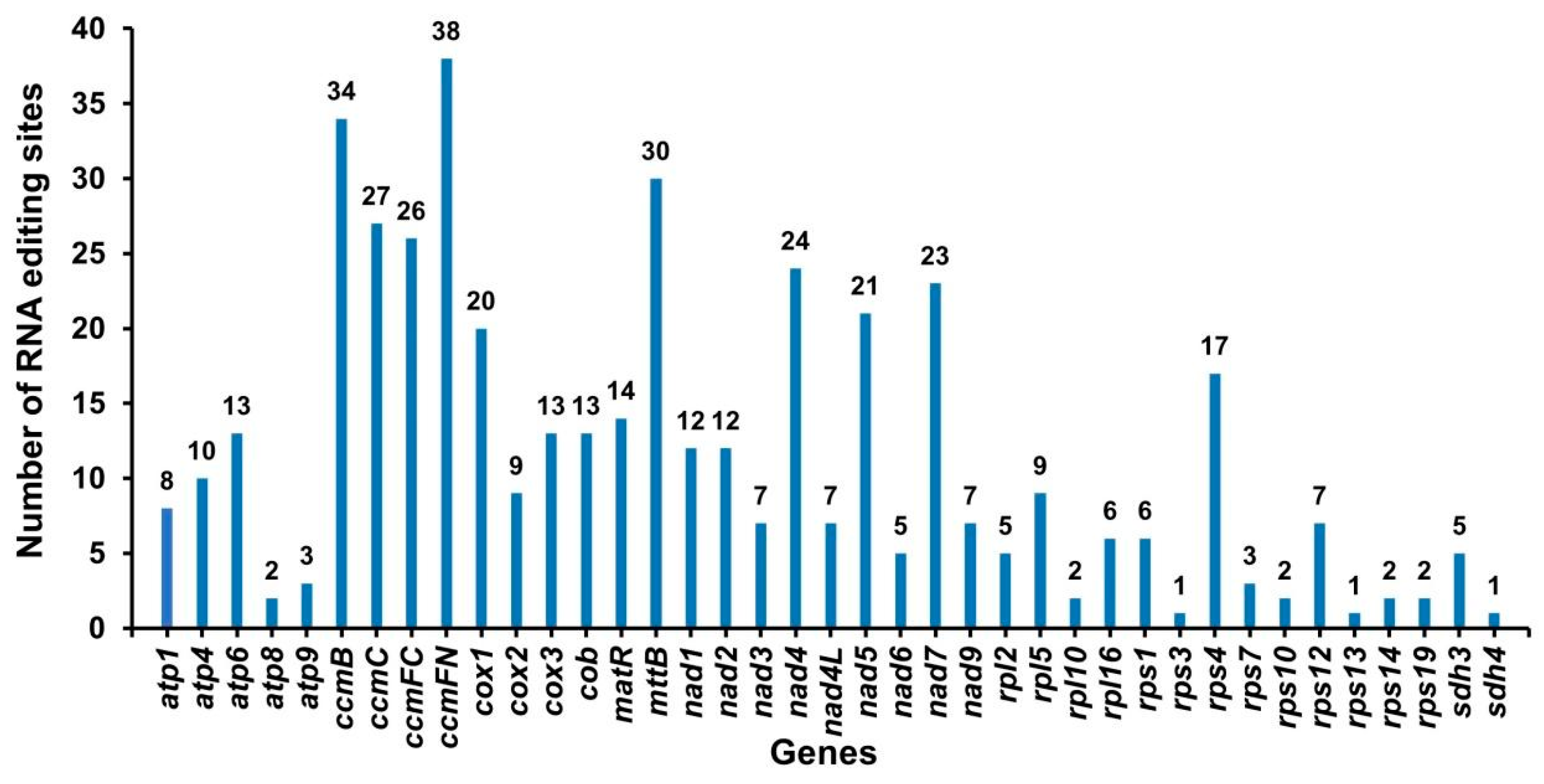
2.4. Phylogenetic Analysis and RNA Editing Events
2.5. Covariance Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Materials, DNA Extraction, and Sequencing
4.2. Genome Assembly and Annotation
4.3. Structural Analysis and Codon Preference Analysis
4.4. Repeat and Homologous DNA Analysis
4.5. Phylogenetic Analysis and RNA Editing Site Prediction
4.6. Covariance Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Raina, K.; Kumar, D.; Agarwal, R. Promise of Bitter Melon (Momordica charantia) Bioactives in Cancer Prevention and Therapy. Semin. Cancer Biol. 2016, 40-41, 116–129. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.-S. Comparison of Antioxidant Capacity and α -Glucosidase Inhibitory Activity between Bitter Melon (Momordica Charanti) Fruit and Leaf Extract. Asian Pac. J. Trop. Biomed. 2018, 8, 189. [Google Scholar] [CrossRef]
- Perez, J.L.; Jayaprakasha, G.K.; Crosby, K.; Patil, B.S. Evaluation of Bitter Melon (Momordica charantia) Cultivars Grown in Texas and Levels of Various Phytonutrients. J. Sci. Food Agric. 2019, 99, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Kenny, O.; Smyth, T.J.; Hewage, C.M.; Brunton, N.P. Antioxidant Properties and Quantitative UPLC-MS Analysis of Phenolic Compounds from Extracts of Fenugreek (Trigonella Foenum-Graecum) Seeds and Bitter Melon (Momordica charantia) Fruit. Food Chem. 2013, 141, 4295–4302. [Google Scholar] [CrossRef] [PubMed]
- Grover, J.K.; Yadav, S.P. Pharmacological Actions and Potential Uses of Momordica charantia: A Review. J. Ethnopharmacol. 2004, 93, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Fang, E.F.; Ng, T.B. Bitter Gourd (Momordica charantia) Is a Cornucopia of Health: A Review of Its Credited Antidiabetic, Anti-HIV, and Antitumor Properties. Curr. Mol. Med. 2011, 11, 417–436. [Google Scholar] [CrossRef]
- Leung, L.; Birtwhistle, R.; Kotecha, J.; Hannah, S.; Cuthbertson, S. Anti-Diabetic and Hypoglycaemic Effects of Momordica charantia (bitter Melon): A Mini Review. Br. J. Nutr. 2009, 102, 1703–1708. [Google Scholar] [CrossRef]
- Begum, S.; Ahmed, M.; Siddiqui, B.S.; Khan, A.; Saify, Z.S.; Arif, M. Triterpenes, A Sterol and A Monocyclic Alcohol from Momordica charantia. Phytochemistry 1997, 44, 1313–1320. [Google Scholar] [CrossRef]
- Douce, R. Mitochondria in Higher Plants: Structure, Function, and Biogenesis; Elsevier: Amsterdam, The Netherlands, 2012; ISBN 9780323155052. [Google Scholar]
- Birky, C.W., Jr. The Inheritance of Genes in Mitochondria and Chloroplasts: Laws, Mechanisms, and Models. Annu. Rev. Genet. 2001, 35, 125–148. [Google Scholar] [CrossRef]
- Bi, C.; Lu, N.; Xu, Y.; He, C.; Lu, Z. Characterization and Analysis of the Mitochondrial Genome of Common Bean (Phaseolus vulgaris) by Comparative Genomic Approaches. Int. J. Mol. Sci. 2020, 21, 3778. [Google Scholar] [CrossRef]
- Ye, N.; Wang, X.; Li, J.; Bi, C.; Xu, Y.; Wu, D.; Ye, Q. Assembly and Comparative Analysis of Complete Mitochondrial Genome Sequence of an Economic Plant. PeerJ 2017, 5, e3148. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; He, X.; Priyadarshani, S.V.G.N.; Wang, Y.; Ye, L.; Shi, C.; Ye, K.; Zhou, Q.; Luo, Z.; Deng, F.; et al. Assembly and Comparative Analysis of the Complete Mitochondrial Genome of Suaeda Glauca. BMC Genom. 2021, 22, 167. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.-M.; Staub, J.E.; Fazio, G. Inheritance of Chilling Injury: A Maternally Inherited Trait in Cucumber. J. Am. Soc. Hortic. Sci. 2003, 128, 526–530. [Google Scholar] [CrossRef]
- Ali, A.; Bang, S.W.; Yang, E.M.; Chung, S.-M.; Staub, J.E. Putative Paternal Factors Controlling Chilling Tolerance in Korean Market-Type Cucumber (Cucumis sativus L.). Sci. Hortic. 2014, 167, 145–148. [Google Scholar] [CrossRef]
- Levi, A.; Thomas, C.E.; Thies, J.A.; Simmons, A.M.; Ling, K.-S.; Harrison, H.F.; Hassell, R.; Keinath, A.P. Novel Watermelon Breeding Lines Containing Chloroplast and Mitochondrial Genomes Derived from the Desert Species Citrullus Colocynthis. HortScience 2006, 41, 463–464. [Google Scholar] [CrossRef]
- Alverson, A.J.; Rice, D.W.; Dickinson, S.; Barry, K.; Palmer, J.D. Origins and Recombination of the Bacterial-Sized Multichromosomal Mitochondrial Genome of Cucumber. Plant Cell 2011, 23, 2499–2513. [Google Scholar] [CrossRef]
- Alverson, A.J.; Wei, X.; Rice, D.W.; Stern, D.B.; Barry, K.; Palmer, J.D. Insights into the Evolution of Mitochondrial Genome Size from Complete Sequences of Citrullus Lanatus and Cucurbita Pepo (Cucurbitaceae). Mol. Biol. Evol. 2010, 27, 1436–1448. [Google Scholar] [CrossRef]
- Urasaki, N.; Takagi, H.; Natsume, S.; Uemura, A.; Taniai, N.; Miyagi, N.; Fukushima, M.; Suzuki, S.; Tarora, K.; Tamaki, M.; et al. Draft Genome Sequence of Bitter Gourd (Momordica charantia), a Vegetable and Medicinal Plant in Tropical and Subtropical Regions. DNA Res. 2017, 24, 51–58. [Google Scholar] [CrossRef]
- Cui, J.; Yang, Y.; Luo, S.; Wang, L.; Huang, R.; Wen, Q.; Han, X.; Miao, N.; Cheng, J.; Liu, Z.; et al. Whole-Genome Sequencing Provides Insights into the Genetic Diversity and Domestication of Bitter Gourd (Momordica Spp.). Hortic. Res. 2020, 7, 85. [Google Scholar] [CrossRef]
- Matsumura, H.; Hsiao, M.-C.; Lin, Y.-P.; Toyoda, A.; Taniai, N.; Tarora, K.; Urasaki, N.; Anand, S.S.; Dhillon, N.P.S.; Schafleitner, R.; et al. Long-Read Bitter Gourd Genome and the Genomic Architecture of Nonclassic Domestication. Proc. Natl. Acad. Sci. USA 2020, 117, 14543–14551. [Google Scholar] [CrossRef]
- Morgante, M.; Hanafey, M.; Powell, W. Microsatellites Are Preferentially Associated with Nonrepetitive DNA in Plant Genomes. Nat. Genet. 2002, 30, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Moreno, L.; González, V.M.; Benjak, A.; Martí, M.C.; Puigdomènech, P.; Aranda, M.A.; Garcia-Mas, J. Determination of the Melon Chloroplast and Mitochondrial Genome Sequences Reveals That the Largest Reported Mitochondrial Genome in Plants Contains a Significant Amount of DNA Having a Nuclear Origin. BMC Genom. 2011, 12, 424. [Google Scholar] [CrossRef] [PubMed]
- Giegé, P.; Brennicke, A. RNA Editing in Arabidopsis Mitochondria Effects 441 C to U Changes in ORFs. Proc. Natl. Acad. Sci. USA 1999, 96, 15324–15329. [Google Scholar] [CrossRef] [PubMed]
- Notsu, Y.; Masood, S.; Nishikawa, T.; Kubo, N.; Akiduki, G.; Nakazono, M.; Hirai, A.; Kadowaki, K. The Complete Sequence of the Rice (Oryza sativa L.) Mitochondrial Genome: Frequent DNA Sequence Acquisition and Loss during the Evolution of Flowering Plants. Mol. Genet. Genom. 2002, 268, 434–445. [Google Scholar] [CrossRef]
- Handa, H. The Complete Nucleotide Sequence and RNA Editing Content of the Mitochondrial Genome of Rapeseed (Brassica napus L.): Comparative Analysis of the Mitochondrial Genomes of Rapeseed and Arabidopsis Thaliana. Nucleic Acids Res. 2003, 31, 5907–5916. [Google Scholar] [CrossRef]
- Bartoszewski, G.; Malepszy, S.; Havey, M.J. Mosaic (MSC) Cucumbers Regenerated from Independent Cell Cultures Possess Different Mitochondrial Rearrangements. Curr. Genet. 2004, 45, 45–53. [Google Scholar] [CrossRef]
- Bartoszewski, G.; Havey, M.J.; Ziółkowska, A.; Długosz, M.; Malepszy, S. The Selection of Mosaic (MSC) Phenotype after Passage of Cucumber (Cucumis sativus L.) through Cell Culture—A Method to Obtain Plant Mitochondrial Mutants. J. Appl. Genet. 2007, 48, 1–9. [Google Scholar] [CrossRef]
- Pawełkowicz, M.; Zieliński, K.; Zielińska, D.; Pląder, W.; Yagi, K.; Wojcieszek, M.; Siedlecka, E.; Bartoszewski, G.; Skarzyńska, A.; Przybecki, Z. Next Generation Sequencing and Omics in Cucumber (Cucumis sativus L.) Breeding Directed Research. Plant Sci. 2016, 242, 77–88. [Google Scholar] [CrossRef]
- Dewey, R.E.; Timothy, D.H.; Levings, C.S. A Mitochondrial Protein Associated with Cytoplasmic Male Sterility in the T Cytoplasm of Maize. Proc. Natl. Acad. Sci. USA 1987, 84, 5374–5378. [Google Scholar] [CrossRef]
- Satoh, M.; Kubo, T.; Mikami, T. The Owen Mitochondrial Genome in Sugar Beet (Beta vulgaris L.): Possible Mechanisms of Extensive Rearrangements and the Origin of the Mitotype-Unique Regions. Theor. Appl. Genet. 2006, 113, 477–484. [Google Scholar] [CrossRef]
- Pruitt, K.D.; Hanson, M.R. Transcription of the Petunia Mitochondrial CMS-Associated Pcf Locus in Male Sterile and Fertility-Restored Lines. Mol. Gen. Genet. MGG 1991, 227, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.-J.; Yu, W.-B.; Yang, J.-B.; Song, Y.; dePamphilis, C.W.; Yi, T.-S.; Li, D.-Z. GetOrganelle: A Fast and Versatile Toolkit for Accurate de Novo Assembly of Organelle Genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Wick, R.R.; Schultz, M.B.; Zobel, J.; Holt, K.E. Bandage: Interactive Visualization of de Novo Genome Assemblies. Bioinformatics 2015, 31, 3350–3352. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and Accurate Short Read Alignment with Burrows-Wheeler Transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq—Versatile and Accurate Annotation of Organelle Genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef]
- Shi, L.; Chen, H.; Jiang, M.; Wang, L.; Wu, X.; Huang, L.; Liu, C. CPGAVAS2, an Integrated Plastome Sequence Annotator and Analyzer. Nucleic Acids Res. 2019, 47, W65–W73. [Google Scholar] [CrossRef]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A Program for Improved Detection of Transfer RNA Genes in Genomic Sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef]
- Chen, Y.; Ye, W.; Zhang, Y.; Xu, Y. High Speed BLASTN: An Accelerated MegaBLAST Search Tool. Nucleic Acids Res. 2015, 43, 7762–7768. [Google Scholar] [CrossRef]
- Thornton, J. Faculty Opinions Recommendation of Apollo: A Sequence Annotation Editor. Faculty Opinions—Post-Publication Peer Review of the Biomedical Literature; BioMed Central Ltd.: London, UK, 2003. [Google Scholar]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An Integrated and Scalable Desktop Platform for Streamlined Molecular Sequence Data Management and Evolutionary Phylogenetics Studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-Web: A Web Server for Microsatellite Prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef] [PubMed]
- Benson, G. Tandem Repeats Finder: A Program to Analyze DNA Sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef]
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The Manifold Applications of Repeat Analysis on a Genomic Scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Meltzer, P.; Davis, S. RCircos: An R Package for Circos 2D Track Plots. BMC Bioinform. 2013, 14, 244. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent Updates and New Developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef]
- Lenz, H.; Hein, A.; Knoop, V. Plant Organelle RNA Editing and Its Specificity Factors: Enhancements of Analyses and New Database Features in PREPACT 3.0. BMC Bioinform. 2018, 19, 1–18. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group of Genes | Name of Genes | |
|---|---|---|
| Core genes | ATP synthase | atp1, atp4, atp6, atp8, atp9 |
| NADH dehydrogenase | nad1, nad2, nad3, nad4, nad4L, nad5, nad6, nad7, nad9 | |
| Cytochrome c biogenesis | ccmB, ccmC, ccmFC, ccmFN | |
| Ubiquinol cytochrome c reductase | cob | |
| Cytochrome c oxidase | cox1, cox2, cox3 | |
| Maturases | matR | |
| Transport membrane protein | mttB | |
| Variable genes | Large subunit of ribosome | rpl2, rpl5, rpl10, rpl16 |
| Small subunit of ribosome | rps1, rps3, rps4, rps7, rps10, rps12, rps13, rps14, rps19 (×2) | |
| Succinate dehydrogenase | sdh3, sdh4 | |
| rRNA genes | Ribosome RNA | rrn5, rrn18, rrn26 |
| tRNA genes | Transfer RNA | trnA-UGC, trnC-GCA (×2), trnD-GUC, trnEUUC, trnF-GAA, trnfM-CAU, trnG-GCC, trnHGUG, trnI-CAU, trnK-UUU, trnL-CAA, trnMCAU, trnN-GUU (×2), trnP-UGG (×2), trnQUUG, trnR-ACG, trnS-GCU, trnS-UGA, trnWCCA, trnY-GU |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, Y.; Zhang, T.; Chen, M.; Chen, G.; Liu, Z.; Yu, R.; Han, X.; Chen, K.; Huang, A.; Chen, C.; et al. Analysis of the Complete Mitochondrial Genome of the Bitter Gourd (Momordica charantia). Plants 2023, 12, 1686. https://doi.org/10.3390/plants12081686
Niu Y, Zhang T, Chen M, Chen G, Liu Z, Yu R, Han X, Chen K, Huang A, Chen C, et al. Analysis of the Complete Mitochondrial Genome of the Bitter Gourd (Momordica charantia). Plants. 2023; 12(8):1686. https://doi.org/10.3390/plants12081686
Chicago/Turabian StyleNiu, Yu, Ting Zhang, Muxi Chen, Guoju Chen, Zhaohua Liu, Renbo Yu, Xu Han, Kunhao Chen, Aizheng Huang, Changming Chen, and et al. 2023. "Analysis of the Complete Mitochondrial Genome of the Bitter Gourd (Momordica charantia)" Plants 12, no. 8: 1686. https://doi.org/10.3390/plants12081686
APA StyleNiu, Y., Zhang, T., Chen, M., Chen, G., Liu, Z., Yu, R., Han, X., Chen, K., Huang, A., Chen, C., & Yang, Y. (2023). Analysis of the Complete Mitochondrial Genome of the Bitter Gourd (Momordica charantia). Plants, 12(8), 1686. https://doi.org/10.3390/plants12081686