
New Approach to the Systematics of the Section Psammiris (Iris, Iridaceae): What Does Chloroplast DNA Sequence Tell Us?



Abstract
1. Introduction
2. Materials and Methods
2.1. Taxa Used
2.2. Plant Samples, DNA Extraction, and Sequencing
2.3. Sequence Alignment and Phylogenetic Analysis
2.4. Morphological Data
2.5. Taxonomy and Distribution
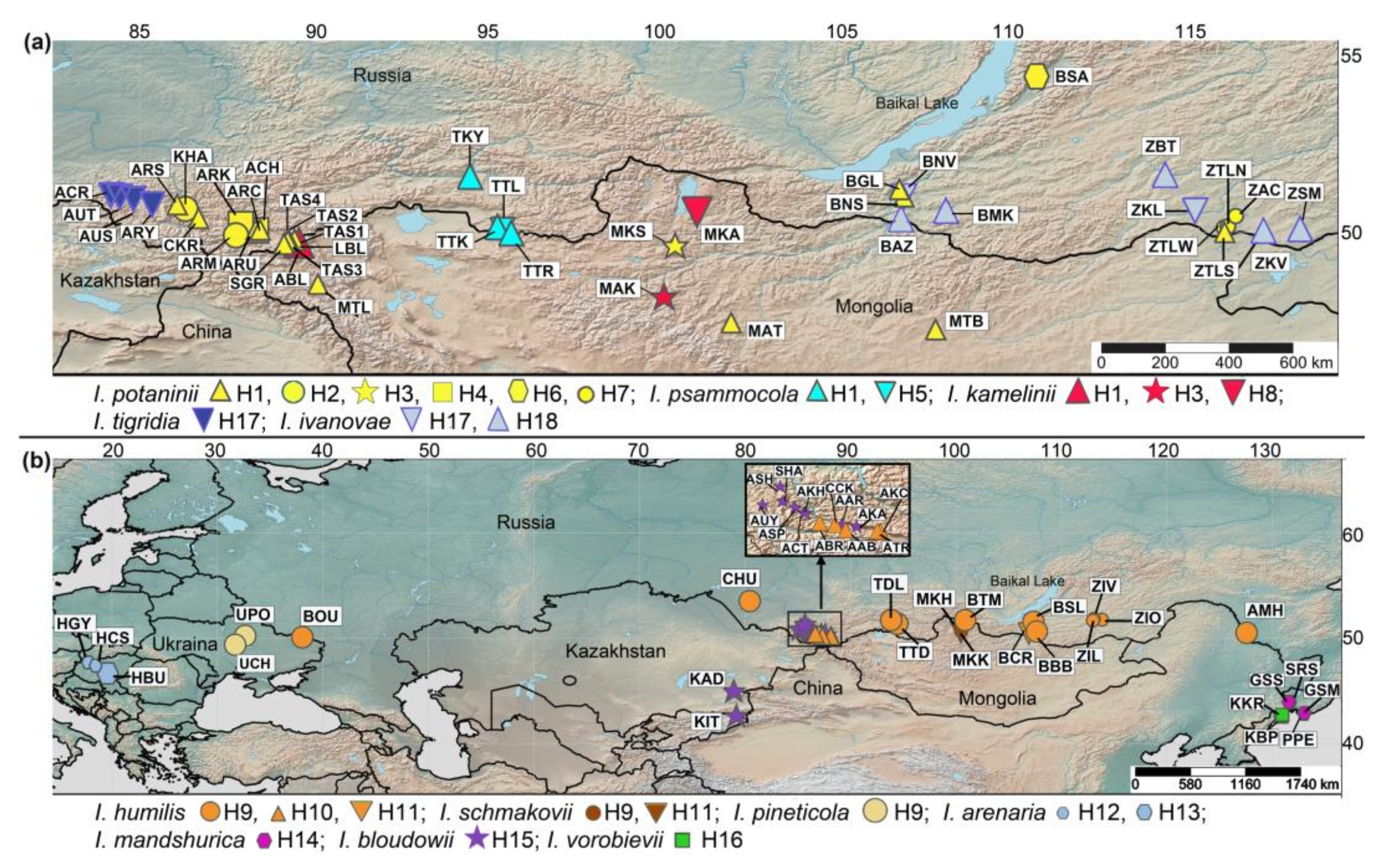
3. Results
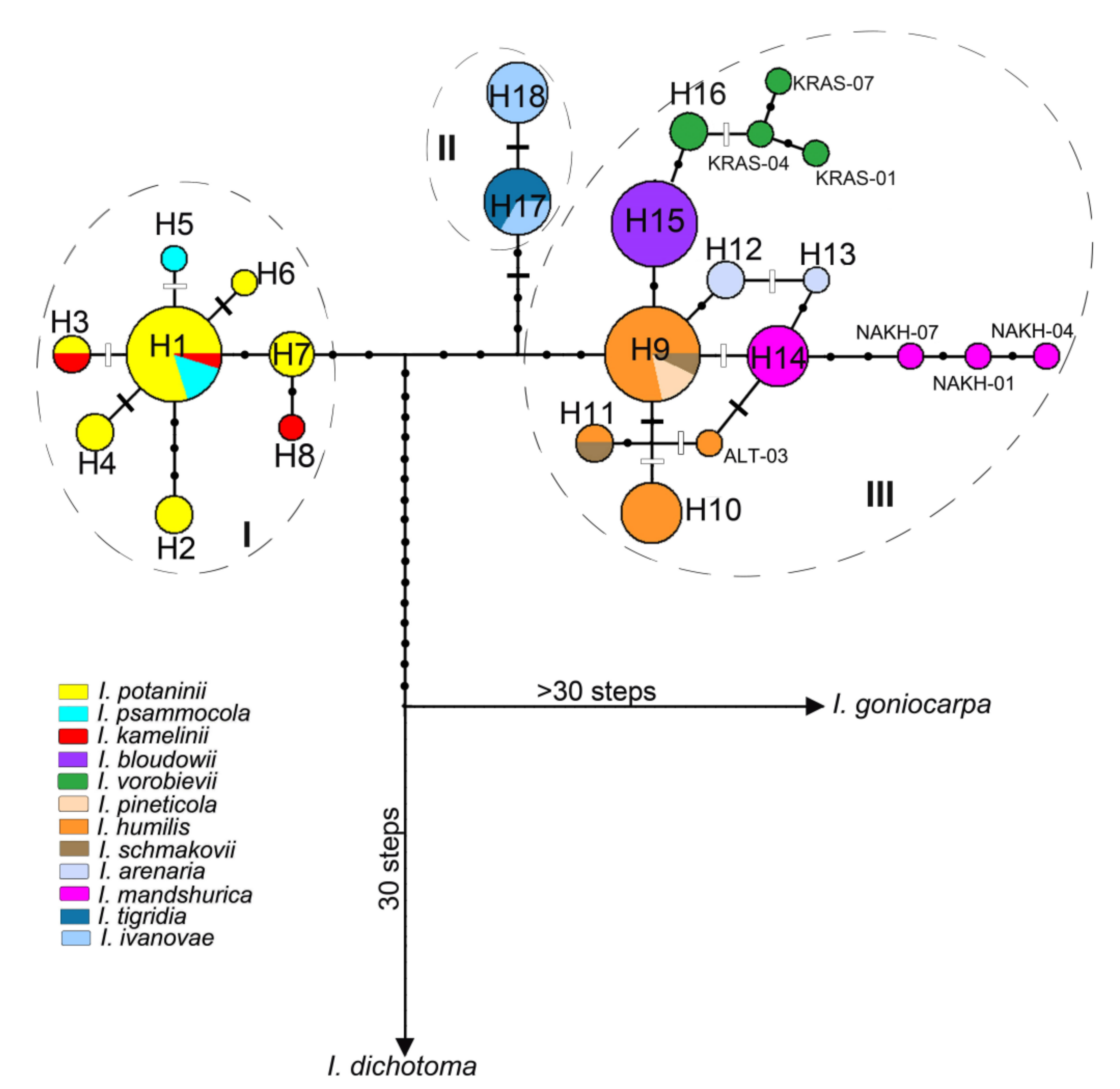
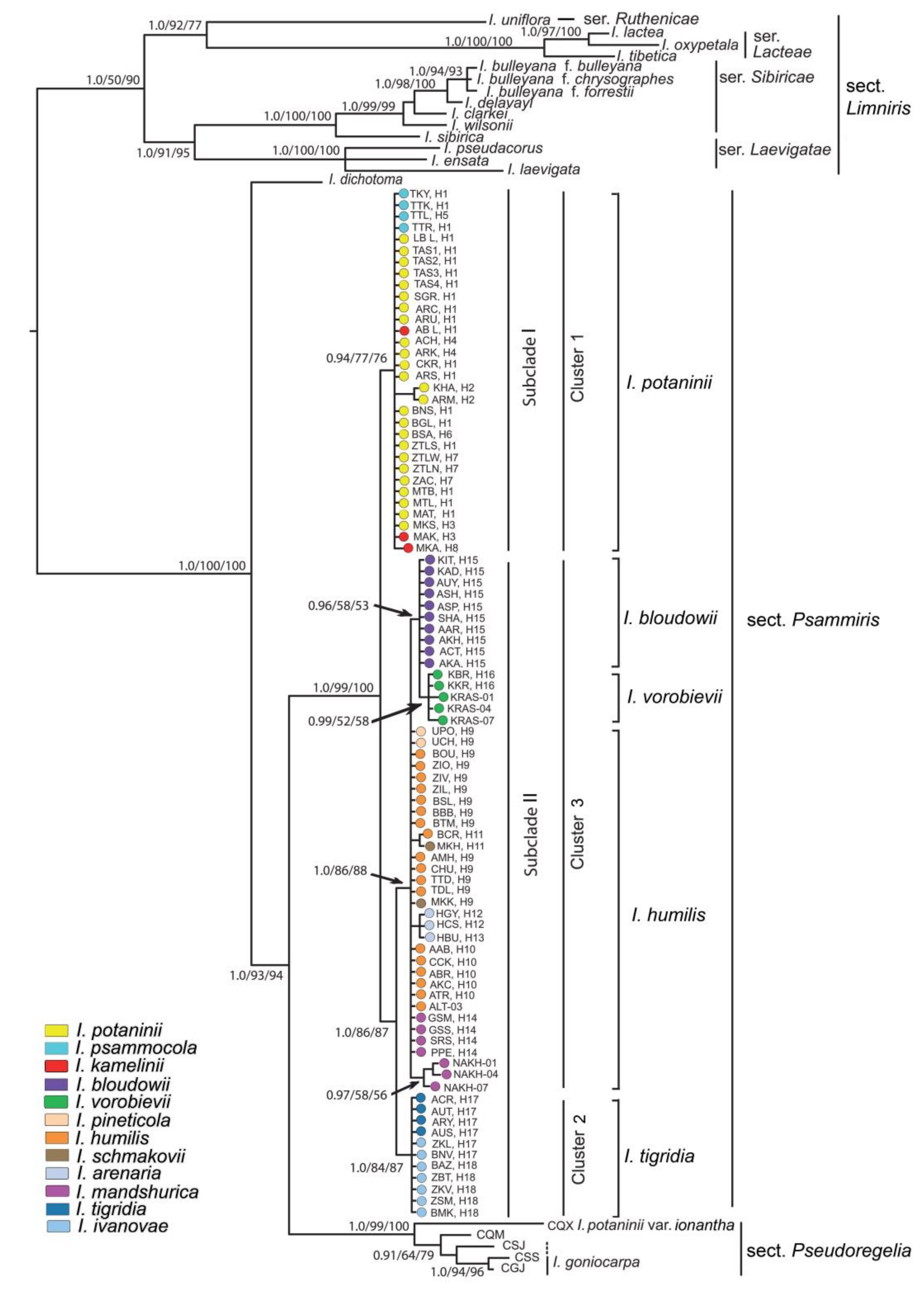
3.1. Genetic Divergence and Phylogenetic Relationships within Iris Sect. Psammiris
3.2. Morphological Comparison of the Iris Sect. Psammiris Species
4. Discussion
4.1. Taxonomic Treatment
4.1.1. List of Taxa
4.1.2. The Key
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Rodionenko, G.I. Comprehending the Secrets of Nature (My Fate—Irises); St. Petersburg Publishing and Printing College: St. Petersburg, Russia, 2013. [Google Scholar]
- Afanasieva, E.A.; Danilova, N.S. The seasonal rhythm characteristics of the Iris species in the Central Yakutia conditions. In Iris–2016, Proceedings of the III Moscow International Symposium, Moscow, Russia, 15–18 June 2016; Novikov, V.S., Ed.; MAKS Press: Moscow, Russia, 2016; pp. 75–80. [Google Scholar]
- Spach, E. Revisio generis Iris. Ann. Sci. Nat. Bot. Ser. 3 1846, 5, 89–111. [Google Scholar]
- Baker, J.G. A synopsis of the known species of Iris; XII. Gard. Chron. New Ser. 1876, 6, 647–648. [Google Scholar]
- Lynch, R.I. The Book of the Iris; John Lane: London, UK; New York, NY, USA, 1904. [Google Scholar] [CrossRef]
- Dykes, W.R. The Genus Iris; Cambridge University Press: Cambridge, UK, 1913. [Google Scholar] [CrossRef]
- Dykes, W.R. A Handbook of Garden Irises; M. Hopkinson & Co.: London, UK, 1924. [Google Scholar] [CrossRef]
- Lawrence, G.H.M. A reclassification of the genus Iris. Gentes Herbarum 1953, 8, 346–371. [Google Scholar]
- Taylor, J.J. A reclassification of Iris species bearing arillate seeds. Proc. Biol. Soc. Wash. 1976, 89, 411–420. [Google Scholar]
- Mathew, B. The Iris, 2nd ed.; Timber Press: Portland, OR, USA, 1989. [Google Scholar]
- Doronkin, V.M. The synopsis of Siberian species of the genus Iris (Iridaceae). Bot. Zhurn. 1990, 75, 409–416. [Google Scholar]
- Rodionenko, G.I. A new system of the genus Iris (Iridaceae). Bot. Zhurn. 2009, 94, 423–435. [Google Scholar]
- Service, N. Section Psammiris (Spach) J.Taylor. In A Guide to Species Irises: Their Identification and Cultivation; The Species Group of the British Iris Society, Ed.; Cambridge University Press: Cambridge, UK, 2012; pp. 58–62. [Google Scholar]
- Wilson, C.A. Sectional relationships in the Eurasian bearded Iris (subgen. Iris) based on phylogenetic analyses of sequence data. Syst. Bot. 2017, 42, 392–401. [Google Scholar] [CrossRef]
- Alexeeva, N.B. A taxonomic revision of Iris section Psammiris (Iridaceae) in Russia. Phytotaxa 2018, 340, 201–216. [Google Scholar] [CrossRef]
- Crespo, M.B.; Alexeeva, N.B.; Xiao, Y.E. Iris zhaoana, a new name for Iris potaninii var. ionantha (I. sect. Pseudoregelia ser. Tigridiae, Iridaceae) from China: Evidence from morphological and plastid DNA data. Phytotaxa 2020, 470, 282–289. [Google Scholar] [CrossRef]
- Alexeeva, N.B. New species of Iris L. (Iridaceae) from Mongolia. Turczaninowia 2018, 4, 145–149. [Google Scholar] [CrossRef]
- Georgi, J.G. Bemerkungen einer Reise im Russischen Reich in den Jahren 1773 und 1774; Kayserliche Academie der Wissenschaften: St. Petersburg, Russia, 1775; Volume 2. [Google Scholar]
- Georgi, J.G. Geographisch-Physikalische und Naturhistorische Beschreibung des Russischen Reichs zur Übersicht Bisheriger Kenntnisse von Demselben; Friedrich Nicolovius: Konigsberg, Russia, 1800; Volume 3. [Google Scholar]
- Maximowicz, C.J. Diagnoses plantarum novarum asiaticarum, III. Bull. Acad. Imp. Sci. Saint-Pétersbourg 1880, 26, 420–542. [Google Scholar]
- Baker, J.G. Handbook of the Irideae; George Bell & Sons: London, UK, 1892. [Google Scholar] [CrossRef]
- Ougrinsky, C. Notices critiques sur quelques plantes de la flore de Kharcoff, II. Trudy Obsc. Isp. Prir. Imp. Har’kovsk. Univ. 1911, 44, 287–318. [Google Scholar]
- Ugrinsky, K.A. Die Gesamtart Iris flavissima Pall. Repert. Spec. Nov. Regni Veg. Beih. 1922, 14, 1–40. [Google Scholar]
- Sand, W.W.A. A study of Pogoniris varieties. Mem. N. Y. Agric. Exp. Stn. 1926, 100, 1–159. [Google Scholar]
- Pavlov, N.V. Materials on the flora of Northern and Central Mongolia brought by the botanical expeditions in 1924 and 1926. Byull. Moskovsk. Obshch. Isp. Prir. Otd. Biol. 1929, 38, 1–153. [Google Scholar]
- Hasselbring, H. The Standard Cyclopedia of Horticulture; Bailey, L.H., Ed.; The Macmillan Company: New York, NY, USA, 1947; Volume 2, pp. 1663–1682. [Google Scholar]
- Grubov, V.I. Iridaceae. In Plantae Asiae Centralis; Grubov, V.I., Egorova, T.V., Eds.; Nauka: Leningrad, Russia, 1977; Volume 7, pp. 88–102. [Google Scholar]
- Pallas, P.S. Reise Durch Verschiedene Provinzen des Russischen Reichs; Kaiserlichen Akademie der Wissenschaften: St. Petersburg, Russia, 1776; Volume 3. [Google Scholar]
- Bobrov, E.G. Species Georgianae neglectae e “Flora Baicalensi”. Bot. Mater. Gerb. Bot. Inst. Komarova Akad. Nauk SSSR 1960, 20, 3–22. [Google Scholar]
- Lenz, L.W. Review of the taxa of the genus Iris. In The World of Irises; Warburton, B., Ed.; Publishers Press: Salt Lake City, UT, USA, 1978; pp. 8–42. [Google Scholar]
- Mathew, B. What is Iris vorobievii? In Iris Year Book; The British Iris Society: London, UK, 1979; pp. 72–73. [Google Scholar]
- Pavlova, N.S. Iridaceae Juss. In Sosudistye Rasteniya Sovetskogo Dal’nego Vostoka [Vascular Plants of the Soviet Far East]; Kharkevich, S.S., Ed.; Nauka: Leningrad, Russia, 1987; Volume 1, pp. 414–426. [Google Scholar]
- Pavlova, N.S. Iridaceae Juss. In Flora of the Russian Far East; Kozhevnikov, A.E., Probatova, N.S., Eds.; Dalnauka: Vladivostok, Russia, 2006; pp. 277–279. [Google Scholar]
- Kozyrenko, M.M.; Artyukova, E.V.; Zhuravlev, Y.N. Independent species status of Iris vorobievii N.S.Pavlova, Iris mandshurica Maxim., and Iris humilis Georgi (Iridaceae): Evidence from the nuclear and chloroplast genomes. Russ. J. Genet. 2009, 45, 1394–1402. [Google Scholar] [CrossRef]
- Webb, D.A.; Chater, A.O. Iris, L. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1980; Volume 5, pp. 87–92. [Google Scholar]
- Hegi, G. Illustriere Flora von Mittel-Europa; J.F. Lehmanns: Munich, Germany, 1909; Volume 2. [Google Scholar]
- Krahulcová, A. Chromosome numbers in selected monocotyledons (Czech Republic, Hungary, and Slovakia). Preslia 2003, 75, 97–113. [Google Scholar]
- Mayorov, S.R. Iridaceae Juss. In Flora of the Middle Zone of the European Part of Russia, 11th ed.; Maevskii, P.F., Ed.; KMK Scientific Press: Moscow, Russia, 2014; pp. 466–468. [Google Scholar]
- Fomin, A.V.; Bordzilowski, E.I. Iridaceae Lindl. In Flora URSR.; Kotov, M.I., Barbaricz, A.I., Eds.; Izdatel’stvo Akademii nauk Ukrainskoy SSR: Kyiv, Ukraine, 1950; Volume 3, pp. 276–312. [Google Scholar]
- Mądalski, J. Atlas Flory Polskiej i Ziem Ościennych; PAN: Kraków, Poland, 1990; Volume 2. [Google Scholar]
- Zhygalova, S.L.; Olshanskyi, I.G. Typification of the name Iris pineticola (Iridaceae). Phytotaxa 2016, 265, 93–94. [Google Scholar] [CrossRef]
- The Global Biodiversity Information Facility. Available online: https://www.gbif.org/ (accessed on 20 December 2022).
- Ledebour, C.F. Icones Plantarum Novarum vel Imperfecte Cognitarum Floram Rossicam, Imprimis Altaicam, Illustrantes; I. Deubner: Riga, Latvia, 1830; Volume 2. [Google Scholar] [CrossRef]
- Ledebour, C.F. Reise durch das Altai-Gibirge und die Soongorische Kirgisen-Steppe; G. Reimer: Berlin, Germany, 1829; Volume 1. [Google Scholar] [CrossRef]
- Peschkova, G.A. Iridaceae. In Flora Sibiriae Centralis; Malyschev, L.I., Peschkova, G.A., Eds.; Nauka: Novosibirsk, Russia, 1979; Volume 1, pp. 230–234. [Google Scholar]
- Zhao, Y.-T.; Noltie, H.J.; Mathew, B. Iridaceae. In Flora of China; Wu, Z.-Y., Raven, P.H., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2000; Volume 24, pp. 297–313. [Google Scholar]
- Fu, L.; Hong, T. Higher Plants of China; Qingdao Publishing House: Qingdao, China, 2002; Volume 13. [Google Scholar]
- Mathew, B. Iris Bloudowii. Bot. Mag. Ser. 6 2007, 24, 30–33. [Google Scholar] [CrossRef]
- Zhao, Y.-T. Some notes on the genus Iris of China. Acta Phytotax. Sin. 1980, 18, 53–62. Available online: https://www.jse.ac.cn/EN/Y1980/V18/I1/53 (accessed on 20 December 2022).
- Wilson, C.A. Subgeneric classification in Iris re-examined using chloroplast sequence data. Taxon 2011, 60, 27–35. [Google Scholar] [CrossRef]
- Mavrodiev, E.V.; Martínez-Azorín, M.; Dranishnikov, P.; Crespo, M.B. At least 23 genera instead of one: The case of Iris L. s.l. (Iridaceae). PLoS ONE 2014, 9, e106459. [Google Scholar] [CrossRef] [PubMed]
- Pirogov, Y. Iris potaninii Maxim. and Iris thoroldii Baker ex Hemsl. a history of confusion. In Iris–2011, Proceedings of the II Moscow International Symposium, Moscow, Russia, 14–17 June 2011; Novikov, V.S., Ed.; MAKS Press: Moscow, Russia, 2011; pp. 111–114. [Google Scholar]
- Zhao, Y.-T. A new species of Iris from China. Acta Phytotax. Sin. 1992, 30, 181–182. Available online: https://www.jse.ac.cn/EN/Y1992/V30/I2/181 (accessed on 20 December 2022).
- Subject Database of China Plant. Available online: http://www.plant.csdb.cn/ (accessed on 20 December 2022).
- Doronkin, V.M.; Shaulo, D.N. Iris psammocola (Iridaceae), a new species to the flora of Russia. Bot. Zhurn. 2007, 92, 435–439. [Google Scholar]
- Doronkin, V.; Vlasova, N.; Ochgerel, N.; Enkhtuya, L.; Munkh-Erdene, T. Rare species of psammophyte flora in transboundary areas of Southern Siberia and Mongolia. In Results and Prospects of Geobotanical Research in Siberia, BIO Web of Conferences, Novosibirsk, Russia, 13–17 May 2019; Lashinsky, N.N., Makunina, N.I., Eds.; EDP Sciences: Les Ulis, France, 2019; Volume 16, p. 6. [Google Scholar] [CrossRef]
- Doronkin, V.M.; Krasnikov, A.A. Cytotaxonomic studies in some Siberian species of the genus Iris (Iridaceae). Bot. Zhurn. 1984, 69, 683–685. [Google Scholar]
- Mitrenina, E.Y.; Boltenkov, E.V.; Erst, A.S.; Wang, W. IAPT chromosome data 38/6. Taxon 2022, 71, 1353–1360. [Google Scholar] [CrossRef]
- Zakhariyeva, O.I.; Makushenko, L.M. Chromosome numbers of monocotyledons belonging to the families Liliaceae, Iridaceae, Amaryllidaceae and Araceae. Bot. Zhurn. 1969, 54, 1213–1227. [Google Scholar]
- Krasnikov, A.A.; Doronkin, V.M. Chromosome numbers of some Iris species (Iridaceae) of Asian Russia. Bot. Zhurn. 2009, 94, 444–445. [Google Scholar]
- Alexeeva, N. Generis Iris L. (Iridaceae) species nova e republica Altai. Novosti Sist. Vyssh. Rast. 2006, 38, 116–119. [Google Scholar]
- Alexeeva, N. Successful expedition to Altai. SIGNA 2007, 78, 3908–3909. [Google Scholar]
- Alexeeva, N.B. In the Altai together with Rudolf Vladimirovich Kamelin. Turczaninowia 2016, 4, 47–52. [Google Scholar] [CrossRef][Green Version]
- Alexeeva, N.B. Seed morphology in the genus Iris (Iridaceae) from Russia. Vavilovia 2020, 3, 5–28. [Google Scholar] [CrossRef]
- Olonova, M.V.; Zhang, D.; Ulkhan, B. On the identification of important plant areas on Altai Mountain Country. Tomsk State Univ. J. Biol. 2013, 21, 59–73. [Google Scholar] [CrossRef]
- Wilson, C.A. Patterns of evolution in characters that define Iris subgenera and sections. Aliso 2006, 22, 425–433. [Google Scholar] [CrossRef]
- Tillie, N.; Chase, M.W.; Hall, T. Molecular studies in the genus Iris L.: A preliminary study. Ann. Bot. (Roma) New Ser. 2000, 58, 105–112. [Google Scholar] [CrossRef]
- Rix, M. Section Pseudoregelia Dykes. In A Guide to Species Irises: Their Identification and Cultivation; The Species Group of the British Iris Society, Ed.; Cambridge University Press: Cambridge, UK, 2012; pp. 98–108. [Google Scholar]
- Makarevich, I.; Golovnina, K.; Scherbik, S.; Blinov, A. Phylogenetic relationships of the Siberian Iris species inferred from noncoding chloroplast DNA sequences. Int. J. Plant Sci. 2003, 164, 229–237. [Google Scholar] [CrossRef]
- Alexeeva, N.B. New species of Iris L. (Iridaceae) for the flora of Mongolia. Turczaninowia 2011, 1, 59–60. [Google Scholar]
- Alexeeva, N.B. A new section of the genus Iris (Iridaceae) and nomenclatural combinations of the sectional rank. Bot. Zhurn. 2006, 91, 1095–1096. [Google Scholar]
- Zhao, W. A Phylogenetic Study of Partial Species in Genus Iris L. from China; D. Northeast Normal University: Changchun, China, 2020; pp. 23–38. [Google Scholar] [CrossRef]
- Doronkin, V.M. Iridaceae. In Flora of Siberia; Malyshev, L.I., Peschkova, G.A., Eds.; Nauka: Novosibirsk, Russia, 1987; Volume 4, pp. 113–125. [Google Scholar]
- Grubov, V.I. Opredelitel’ Sosudistykh Rasteniy Mongolii; Key to the Vascular Plants of Mongolia; Nauka: Leningrad, Russia, 1982. [Google Scholar]
- Byazrov, L.G.; Ganbold, E.; Gubanov, I.A.; Ulziykhutag, N. Flora of Khangai; Nauka: Leningrad, Russia, 1989. [Google Scholar]
- Gubanov, I.A. Conspectus of Flora of Outer Mongolia (Vascular Plants); Valang: Moscow, Russia, 1996. [Google Scholar]
- Grubov, V.I. Key to the Vascular Plants of Mongolia; Science Publishers: Enfield, NH, USA, 2001; Volume 1. [Google Scholar]
- Doronkin, V.; Shaulo, D.; Han, I.; Vlasova, N.; Ivleva, V.; Enkhtuya, L.; Munkh-Erdene, T.; Ochgerel, N.; Munkhjargal, B. New records for the flora of Selenge Province (Mongolia). Skvortsovia 2015, 2, 8–27. [Google Scholar]
- Kress, W.J.; Erickson, D.L. A two-locus global DNA barcode for land plants: The coding rbcL gene complements the non-coding trnH-psbA spacer region. PLoS ONE 2007, 2, e508. [Google Scholar] [CrossRef] [PubMed]
- Boltenkov, E.V.; Artyukova, E.V.; Kozyrenko, M.M.; Trias-Blasi, A. Iris tibetica, a new combination in I. ser. Lacteae (Iridaceae) from China: Evidence from morphological and chloroplast DNA analyses. Phytotaxa 2018, 338, 223–240. [Google Scholar] [CrossRef]
- Boltenkov, E.; Artyukova, E.; Kozyrenko, M.; Erst, A.; Trias-Blasi, A. Iris sanguinea is conspecific with I. sibirica (Iridaceae) according to morphology and plastid DNA sequence data. PeerJ 2020, 8, e10088. [Google Scholar] [CrossRef] [PubMed]
- Boltenkov, E.V.; Artyukova, E.V.; Trias-Blasi, A. Taxonomic composition of Iris subser. Chrysographes (Iridaceae) inferred from chloroplast DNA and morphological analyses. Plants 2021, 10, 2232. [Google Scholar] [CrossRef]
- Index Herbariorum. Available online: https://sweetgum.nybg.org/ih/ (accessed on 20 December 2022).
- Bronnikova, M.A.; Agatova, A.R.; Lebedeva, M.P.; Nepop, R.K.; Konoplianikova, Y.V.; Turova, I.V. Record of Holocene changes in high-mountain landscapes of southeastern Altai in the soil–sedimentary sequence of the Boguty river valley. Eurasian Soil Sci. 2018, 51, 1381–1396. [Google Scholar] [CrossRef]
- Kozyrenko, M.M.; Artyukova, E.V.; Boltenkov, E.V.; Lauve, L.S. Somaclonal variability of Iris pseudacorus L. according to RAPD and cytogenetic analyses. Biotechnol. Russ. 2004, 2, 11–22. [Google Scholar]
- Bonfield, J.K.; Smith, K.F.; Staden, R. A new DNA sequence assembly program. Nucleic Acids Res. 1995, 23, 4992–4999. [Google Scholar] [CrossRef]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef]
- Bandelt, H.-J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods), Version 4.0 b10; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE 2010), New Orleans, LA, USA, 14 November 2010; Volume 1, pp. 1–8. [Google Scholar] [CrossRef]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Krylov, P.N. Iridaceae. In Flora Zapadnoi Sibiri [Flora of Western Siberia]; Krylov, P.N., Ed.; Tomskoye Otdeleniye Russkogo Botanicheskogo Obshchestva: Tomsk, Russia, 1929; Volume 3, pp. 660–672. [Google Scholar]
- Fedtschenko, B.A. Iris. In Flora of the USSR.; Komarov, V.L., Ed.; Izdatel’stvo Akademii nauk SSSR: Leningrad, Russia, 1935; Volume 4, pp. 511–557. [Google Scholar]
- Sergievskaya, L.P. Flora Zabaykal’ya [Transbaikal Flora]; Izdatel’stvo Tomskogo Universiteta: Tomsk, Russia, 1972; Volume 4. [Google Scholar]
- The Plant Photo Bank of China. Available online: https://ppbc.iplant.cn/ (accessed on 20 December 2022).
- Chinese Virtual Herbarium. Available online: https://www.cvh.ac.cn/index.php (accessed on 20 December 2022).
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.-H.; Li, D.-Z.; Marhold, K.; et al. (Eds.) International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code) Adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017 [Regnum Vegetabile Volume 159]; Koeltz Botanical Books: Glashütten, Germany, 2018. [Google Scholar] [CrossRef]
- iNaturalist. Available online: https://www.inaturalist.org (accessed on 20 December 2022).
- Plantarium. Available online: https://www.plantarium.ru/lang/en.html (accessed on 20 December 2022).
- Virtual Guide to the Flora of Mongolia. Available online: https://floragreif.uni-greifswald.de/taxon/?flora_search=Record&fam=Iridaceae&gen=Iris&spec=potaninii (accessed on 20 December 2022).
- Murín, A.; Haberová, I.; Žamsran, C. Karyological studies of some species of the Mongolian flora. Folia Geobot. Phytotax. 1980, 15, 395–405. [Google Scholar] [CrossRef]
- Malakhova, L.A.; Markova, G.A. Chromosome numbers in the flowering plants of Tomsk Region, Monocotyledones. Bot. Zhurn. 1994, 79, 134–135. [Google Scholar]
- Krivenko, D.A.; Kotseruba, V.V.; Kazanovsky, S.G.; Verkhozina, A.V.; Stepanov, A.V. IAPT/IOPB chromosome data 11: Iris humilis. Taxon 2011, 60, 1222. [Google Scholar] [CrossRef]
- Probatova, N.S.; Gnutikov, A.A.; Rudyka, E.G.; Chepinoga, V.V. Chromosome numbers of some plant species from Baikal Siberia. Bot. Zhurn. 2008, 93, 162–181. [Google Scholar]
- Probatova, N.S. Chromosome Numbers in Vascular Plants of the Primorskii Krai; Dal’nauka: Vladivostok, Russia, 2014. [Google Scholar]
- Zhao, Y.; Lu, J. Karyotype studies of 3 species of genus Iris in China. J. Northeast Norm. Univ. (Nat. Sci.) 1986, 2, 71–78. [Google Scholar]
- Sokolovskaya, A.P.; Probatova, N.S. Chromosome numbers in some representatives of the Asteraceae, Iridaceae, Poaceae, Primulaceae, Violaceae families from the Far East of the USSR. Bot. Zhurn. 1986, 71, 1423–1425. [Google Scholar]
- Starodubtsev, V.N.; Mirinova, L.N. Chromosome numbers in the species of the genus Iris (Iridaceae) from the flora of the Primorye Territory. Bot. Zhurn. 1990, 75, 123. [Google Scholar]
- Shatokhina, A.V. Chromosome numbers of some vascular plant species rare to the Amur Region. Bot. Zhurn. 2007, 92, 1082–1086. [Google Scholar]
- Sennikov, A.; Khassanov, F.; Ortikov, E.; Kurbonaliyeva, M.; Tojibaev, K.S. The genus Iris L. s. l. (Iridaceae) in the Mountains of Central Asia biodiversity hotspot. Plant Divers. Cent. Asia 2023, 2, 1–104. [Google Scholar] [CrossRef]
- Spach, E. Histoire Naturelle des Végétaux. Phanerogames; Librairie Encyclopédique de Roret: Paris, France, 1846; Volume 13, pp. 69–70. [Google Scholar] [CrossRef]
- Alexeeva, N.B.; Mironova, L.N. Notes critical about some species of the genus Iris (Iridaceae) in Sibiria and Far East of Russia. Bot. Zhurn. 2007, 92, 916–925. [Google Scholar]
- Alexeeva, N.B. Iridaceae Juss. In Catalogue of the Type Specimens of the Vascular Plants from Siberia and the Russian Far East kept in the Herbarium of the Komarov Botanical Institute (LE); Sokolova, I.V., Ed.; KMK Scientific Press: Moscow—St. Petersburg, Russia, 2012; pp. 415–419. [Google Scholar]
- Waldstein, F.; Kitaibel, P. Descriptiones et Icones Plantarum Rariorum Hungariae; Typis Matthiae Andreae Schmidt: Viennae, Austria, 1802; Volume 1, p. 57. [Google Scholar]
- Neuffer, B.; Friesen, N.; Oyuntsetseg, B.; Jamsran, T.; Hurka, H. Osnabrück botanical expeditions to Mongolia. Erforsch. Biol. Ress. Mongolei 2012, 12, 307–333. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code (Haplotype) | Locality (Voucher *) ** | Coordinates: ° N, ° E | GenBank Accession Nos. trnH-psbA/rps4-trnS/trnS-trnG/trnL-trnF |
|---|---|---|---|
| Iris subgen. Iris | |||
| I. psammocola Y.T.Zhao | |||
| TKY (H1) | Russia, Tuva, Kyzyl, A.Yu. Astashenkov s.n. (VBGI) | 51.58211, 94.35711 | ON569443/ON569531/ON569619/ON569707 |
| TTK (H1) | Russia, Tuva, Lake Tore-Khol, A.Yu. Astashenkov s.n. (VBGI) | 50.15408, 95.13172 | ON569444/ON569532/ON569620/ON569708 |
| TTR (H1) | Russia, Tuva, near Tes River, A.Yu. Astashenkov s.n. (VBGI) | 49.98005, 95.52527 | ON569446/ON569534/ON569622/ON569710 |
| TTL (H5) | Russia, Tuva, Tsugeer-Els, D.N. Shaulo & V.M. Doronkin 17 (LE01072832) | 50.333333, 95.484722 | ON569445/ON569533/ON569621/ON569709 |
| I. potaninii Maxim. | |||
| LBL (H1) | Russia, Altai Republic, Kosh-Agach District, Lake Nizhniye Boguty, Boltenkov et al. 38 (VBGI) | 49.79277, 89.38888 | ON569447/ON569535/ON569623/ON569711 |
| TAS1 (H1) | Russia, Altai Republic, Kosh-Agach District, Tashanta, Boltenkov et al. 33 (VBGI) | 49.790833, 89.388611 | ON569448/ON569536/ON569624/ON569712 |
| TAS2 (H1) | Russia, Altai Republic, Kosh-Agach District, Tashanta, Boltenkov et al. 39 (VBGI) | 49.76333, 89.2375 | ON569449/ON569537/ON569625/ON569713 |
| TAS3 (H1) | Russia, Altai Republic, Kosh-Agach District, Tashanta, N.V. Shchegoleva s.n. (VBGI) | 49.73307, 89.1562 | ON569450/ON569538/ON569626/ON569714 |
| TAS4 (H1) | Russia, Altai Republic, Kosh-Agach District, 6 km north of Tashanta, P.A. Kosachev et al. s.n. (VBGI) | 49.75941, 89.19536 | ON569451/ON569539/ON569627/ON569715 |
| SGR (H1) | Russia, Altai Republic, Kosh-Agach District, near the Bol’shoy Sar-Gobo River estuary, R.V. Kamelin et al. 2613 (ALTB) | 49.66666, 89.09166 | ON569452/ON569540/ON569628/ON569716 |
| ARC (H1) | Russia, Altai Republic, Kosh-Agach District, Chagan-Uzun, Boltenkov et al. 24 (VBGI) | 50.06055, 88.29222 | ON569453/ON569541/ON569629/ON569717 |
| ARU (H1) | Russia, Altai Republic, Kosh-Agach District, Chagan-Uzun, Boltenkov et al. 26 (VBGI) | 50.0725, 88.41416 | ON569454/ON569542/ON569630/ON569718 |
| CKR (H1) | Russia, Altai Republic, Ongudaysky District, confluence of Chuya and Katun rivers, Boltenkov et al. 22 (VBGI) | 50.39722, 86.67444 | ON569458/ON569546/ON569634/ON569722 |
| ARS (H1) | Russia, Altai Republic, Ongudaysky District, Shashikman, Boltenkov et al. 19 (VBGI) | 50.78583, 86.06361 | ON569459/ON569547/ON569635/ON569723 |
| KHA (H2) | Russia, Altai Republic, Ongudaysky District, Khabarovka, I.M. Krasnoborov 188 (MHA) | 50.66666, 86.3 | ON569460/ON569548/ON569636/ON569724 |
| ARM (H2) | Russia, Altai Republic, Mohro-Oyuk Pass, A.S. Revushkin et al. s.n. (LE) | 49.91769, 87.7311 | ON569461/ON569549/ON569637/ON569725 |
| ACH (H4) | Russia, Altai Republic, Kosh-Agach District, Kyzyl-Chin, P.A. Kosachev et al. s.n. (VBGI) | 50.06021, 88.29927 | ON569456/ON569544/ON569632/ON569720 |
| ARK (H4) | Russia, Altai Republic, Kosh-Agach District, 7 km west of Kuray, P.A. Kosachev et al. s.n. (VBGI) | 50.23663, 87.87082 | ON569457/ON569545/ON569633/ON569721 |
| BNS (H1) | Russia, Buryatia, Novoselenginsk, Boltenkov 57 (VBGI) | 51.01166, 106.64027 | ON569462/ON569550/ON569638/ON569726 |
| BGL (H1) | Russia, Buryatia, northeast of Lake Gusinoye, Boltenkov 62 (VBGI) | 51.21861, 106.51472 | ON569463/ON569551/ON569639/ON569727 |
| BSA (H6) | Russia, Buryatia, Sakhuli, D.G. Chimitov & O.V. Imetkhenova s.n. (UUH) | 54.41666, 110.4 | ON569464/ON569552/ON569640/ON569728 |
| ZTLS (H1) | Russia, Zabaykalsky Krai, southern bank of Lake Zun-Torey, Boltenkov 83 (VBGI) | 50.00222, 115.72055 | ON569465/ON569553/ON569641/ON569729 |
| ZTLW (H7) | Russia, Zabaykalsky Krai, northwestern bank of Lake Zun-Torey, Boltenkov 77 (VBGI) | 50.12972, 115.70361 | ON569466/ON569554/ON569642/ON569730 |
| ZTLN (H7) | Russia, Zabaykalsky Krai, north of Lake Zun-Torey, Boltenkov 80 (VBGI) | 50.1675, 115.81583 | ON569467/ON569555/ON569643/ON569731 |
| ZAC (H7) | Russia, Zabaykalsky Krai, Adon-Chelon, Boltenkov 91 (VBGI) | 50.46388, 116.0375 | ON569468/ON569556/ON569644/ON569732 |
| MTB (H1) | Mongolia, Tow Aimag, Bayan, Ch. Sanchir s.n. (LE) | 47.25111, 107.53833 | ON569469/ON569557/ON569645/ON569733 |
| MTL (H1) | Mongolia, Bayan-Olgii Aimag, Lake Tolbo, A.I. Shmakov & M.G. Kutsev s.n. (ALTB) | 48.536658, 90.050327 | ON569470/ON569558/ON569646/ON569734 |
| MAT (H1) | Mongolia, Arkhangai Aimag, 20 km south of Tsenkher Sum, I.A. Gubanov 255 (MW) | 47.44527, 101.75027 | ON569471/ON569559/ON569647/ON569735 |
| MKS (H3) | Mongolia, Khuvsgul Aimag, 25 km north of Sumber, A.L. Budantsev et al. 208 (MW) | 49.63333, 100.16694 | ON569472/ON569560/ON569648/ON569736 |
| I. kamelinii Alexeeva | |||
| ABL (H1) | Russia, Altai Republic, Kosh-Agach District, Lake Verkniye Boguty, Boltenkov et al. 34 (VBGI) ** | 49.70583, 89.51333 | ON569455/ON569543/ON569631/ON569719 |
| MAK (H3) | Mongolia, Arkhangai Province, Khorgo Mountain, N.B. Alexeeva et al. 6 (LE01071966!) | 48.18888, 99.84833 | ON569473/ON569561/ON569649/ON569737 |
| MKA (H8) | Mongolia, Khuvsgul Aimag, between Khukhuu and Eg-Uur, N.B. Alexeeva et al. 36 (LE01071967!) | 50.57861, 100.78388 | ON569474/ON569562/ON569650/ON569738 |
| I. bloudowii Ledeb. | |||
| KIT (H15) | Kyrgyzstan, Issyk-Kul Region, northern slope of Terskey Ala-Too, A. Naumenko s.n. (VBGI) | 42.676717, 79.167452 | ON569475/ON569563/ON569651/ON569739 |
| KAD (H15) | Kazakhstan, Almaty Region, Dzhungraian Alatau, 10 km west of Qapal, A.V. Grebenjuk 161 (LE) | 45.02486, 78.94919 | ON569476/ON569564/ON569652/ON569740 |
| AUY (H15) | Russia, Altai Republic, Ust-Kansky District, Yaboganskiy Pass, Boltenkov et al. 15 (VBGI) ** | 50.85194, 85.24194 | ON569477/ON569565/ON569653/ON569741 |
| ASH (H15) | Russia, Altai Republic, Shebalinsky Districtn, Shebalino, Boltenkov et al. 7 (VBGI) | 51.31611, 85.67972 | ON569478/ON569566/ON569654/ON569742 |
| ASP (H15) | Russia, Altai Republic, Ongudaysky District, ascent to the Seminsky Pass, Boltenkov et al. 40 (VBGI) | 50.94472, 85.74111 | ON569479/ON569567/ON569655/ON569743 |
| SHA (H15) | Russia, Altai Republic, Ongudaysky District, Shashikman, L. Lamanova s.n. (LE) | 50.7916, 86.05772 | ON569480/ON569568/ON569656/ON569744 |
| AAR (H15) | Russia, Altai Republic, Ongudaysky District, near the Aygulak River estuary, P.A. Kosachev et al. s.n. (VBGI) | 50.35986, 87.24423 | ON569481/ON569569/ON569657/ON569745 |
| AKH (H15) | Russia, Altai Republic, Ongudaysky Districtn, Khabarovka, Boltenkov et al. 20 (VBGI) | 50.66388, 86.29305 | ON569482/ON569570/ON569658/ON569746 |
| ACT (H15) | Russia, Altai Republic, Ongudaysky District, Chike-Taman Pass, Boltenkov et al. 21 (VBGI) | 50.64388, 86.31083 | ON569483/ON569571/ON569659/ON569747 |
| AKA (H15) | Russia, Altai Republic, Kosh-Agach District, Aktash, A. Dedov s.n. (VBGI) | 50.31111, 87.59916 | ON569484/ON569572/ON569660/ON569748 |
| I. pineticola Klokov | |||
| UPO (H9) | Ukraine, Poltava Oblast, Deimanivka, “Kukvyn”, pine forest, S.L. Zygalova et al. s.n. (KW) ** | 50.21666, 32.63388 | ON569485/ON569573/ON569661/ON569749 |
| UCH (H9) | Ukraine, Cherkasy Oblast, Irdyn, pine forest, S.L. Zygalova et al. s.n. (KW) ** | 49.36916, 31.67916 | ON569486/ON569574/ON569662/ON569750 |
| I. humilis Georgi | |||
| BOU (H9) | Russia, Belgorod Oblast, west of Urazovo, s. coll. s.n. (MHA) | 50.07861, 38.04805 | ON569487/ON569575/ON569663/ON569751 |
| CHU (H9) | Russia, Altai Krai, Bayevsky District, Chumanka, A. Dedov s.n. (VBGI) | 53.5, 80.45 | ON569496/ON569584/ON569672/ON569760 |
| AAB (H10) | Russia, Altai Republic, Ongudaysky District, Ak-Boom, N.V. Shchegoleva s.n. (VBGI) | 50.21429, 87.32491 | ON569504/ON569592/ON569680/ON569768 |
| CCK (H10) | Russia, Altai Republic, Ongudaysky District, confluence of Chuya and Katun rivers, Boltenkov et al. 22 (VBGI) | 50.39722, 86.67444 | ON569505/ON569593/ON569681/ON569769 |
| ABR (H10) | Russia, Altai Republic, Ongudaysky District, Ak-Boom Rock, Boltenkov et al. 23 (VBGI) | 50.35361, 87.05694 | ON569506/ON569594/ON569682/ON569770 |
| AKC (H10) | Russia, Altai Republic, Kosh-Agach Districtn, between Kurai and Chagan-Uzun, Boltenkov et al. 28 (VBGI) | 50.16944, 88.20861 | ON569507/ON569595/ON569683/ON569771 |
| ATR (H10) | Russia, Altai Republic, Kosh-Agach District, 1 km west of the Tydtugem River estuary, P.A. Kosachev et al. s.n. (VBGI) | 50.18732, 88.12405 | ON569508/ON569596/ON569684/ON569772 |
| TTD (H9) | Russia, Tuva, Tandinsky District, Lake Dus-Khol’, Yu.S. Otmakhov 8 (VBGI) | 51.35563, 94.45398 | ON569497/ON569585/ON569673/ON569761 |
| TDL (H9) | Russia, Tuva, Lake Dus-Khol’, Yu.S. Otmakhov 40 (VBGI) | 51.35604, 94.44693 | ON569498/ON569586/ON569674/ON569762 |
| BSL (H9) | Russia, Buryatia, Tarbagataysky District, Mount Spyashchiy Lev, Boltenkov 63 (VBGI) | 51.53833, 107.34611 | ON569491/ON569579/ON569667/ON569755 |
| BBB (H9) | Russia, Buryatia, Bichursky District, Bichura, Boltenkov 112 (VBGI) | 50.62888, 107.66472 | ON569492/ON569580/ON569668/ON569756 |
| BTM (H9) | Russia, Buryatia, Tunkinsky District, Mondy, D.V. Sandanov s.n. (VBGI) | 51.69760, 100.86766 | ON569493/ON569581/ON569669/ON569757 |
| BCR (H11) | Russia, Buryatia, Kyakhtinsky District, Chikoy River, Khilgantuy, Boltenkov 114 (VBGI) ** | 50.44944, 106.91 | ON569494/ON569582/ON569670/ON569758 |
| ZIO (H9) | Russia, Zabaykalsky Krai, Ingoda River, Orlenok, E.V. Boltenkov 109 (VBGI) ** | 51.74722, 113.84638 | ON569488/ON569576/ON569664/ON569752 |
| ZIV (H9) | Russia, Zabaykalsky Krai, Ingoda River, Ingoda Village, Boltenkov 110 (VBGI) ** | 51.83055, 113.08638 | ON569489/ON569577/ON569665/ON569753 |
| ZIL (H9) | Russia, Zabaykalsky Krai, Ingoda River, Lesnoi Gorodok, Boltenkov 111 (VBGI) ** | 51.66722, 112.98166 | ON569490/ON569578/ON569666/ON569754 |
| AMH (H9) | Russia, Amur Oblast, Novotroitskoe Village, G.F. Darman s.n. (VBGI) | 50.428889, 127.549806 | ON569495/ON569583/ON569671/ON569759 |
| ALT-03 | Russia, Altai Republic, Ongudaysky District, Chuya River estuary, L.M. Pshennikova s.n. (VBGI, cult.) | – | FM253737/FM253420/FM864187/FM863912 |
| I. arenaria Waldst. et Kit. | |||
| HGY (H12) | Hungary, Győrszentiván, 24.04.2020, A. Mesterházy s.n. (VBGI) ** | 47.69777, 17.73638 | ON569501/ON569589/ON569677/ON569765 |
| HCS (H12) | Hungary, Csákvár, A. Mesterházy s.n. (VBGI) ** | 47.39332, 18.46049 | ON569502/ON569590/ON569678/ON569766 |
| HBU (H13) | Hungary, Bugac, 28.07.2020, B. Zoltán s.n. (VBGI) ** | 46.65944, 19.59880 | ON569503/ON569591/ON569679/ON569767 |
| I. schmakovii Alexeeva | |||
| MKK (H9) | Mongolia, Khuvsgul Aimag, Khatgal Sum, Sh. Baasanmunkh s.n. (VBGI) ** | 50.61924, 100.51207 | ON569499/ON569587/ON569675/ON569763 |
| MKH (H11) | Mongolia, Khuvsgul Aimag, Lake Khuvsgul, R.V. Kamelin et al. 23 (LE) ** | 50.56666, 100.46666 | ON569500/ON569588/ON569676/ON569764 |
| I. mandshurica Maxim. | |||
| GSS (H14) | Russia, Primorsky Krai, Oktyabrsky District, Mount Sen’kina Shapka, Boltenkov s.n. (VBGI) ** | 43.91833, 131.65943 | ON569510/ON569598/ON569686/ON569774 |
| SRS (H14) | Russia, Primorsky Krai, Oktyabrsky Districtn, Sinel’nikovo-1, Boltenkov s.n. (VBGI) ** | 43.96, 131.53361 | ON569511/ON569599/ON569687/ON569775 |
| GSM (H14) | Russia, Primorsky Krai, Nakhodka, Mount Sestra, Boltenkov s.n. (VBGI) | 42.82777, 132.99499 | ON569509/ON569597/ON569685/ON569773 |
| PPE (H14) | Russia, Primorsky Krai, Partizansky District, Ekaterinovka, Boltenkov 123 (VBGI) | 42.91527, 133.04944 | ON569512/ON569600/ON569688/ON569776 |
| NAKH-01 NAKH-04 | Russia, Primorsky Krai, vicinities of Nakhodka, R.V. Dudkin s.n. (VBGI, cult.) | – | FM253719/FM253402/FM864169/FM863894 FM253722/FM253405/ FM864173/FM863897 |
| NAKH-07 | FM253725/FM253408/FM864175/FM863900 | ||
| I. vorobievii N.S.Pavlova | |||
| KKR (H16) | Russia, Primorsky Krai, Khasansky District, Kraskino, Boltenkov s.n. (VBGI) ** | 42.725, 130.93361 | ON569514/ON569602/ON569690/ON569778 |
| KBR (H16) | Russia, Primorsky Krai, Khasansky District, Bay Pemzovaya, E.A. Chubar s.n. (VBGI) | 42.54667, 130.83971 | ON569513/ON569601/ON569689/ON569777 |
| KRAS-01 | Russia, Primorsky Krai, Khasansky District, Kraskino, R.V. Dudkin s.n. (VBGI, cult.) ** | – | FM253702 /FM253385/FM864152/FM863877 |
| KRAS-04 | FM253705/FM253388/FM864155/FM863880 | ||
| KRAS-07 | FM253708/FM253391/FM864158/FM863883 | ||
| I. tigridia Bunge | |||
| ACR (H17) | Russia, Altai Republic, Ust-Kansky District, right bank of Charysh River, Vladimirovka, Boltenkov et al. 11 (VBGI) ** | 51.05388, 84.19 | ON569515/ON569603/ON569691/ON569779 |
| AUT (H17) | Russia, Altai Republic, Ust-Kansky District, 3 km west of Tiudrala, Boltenkov et al. 10 (VBGI) | 51.01, 84.44138 | ON569516/ON569604/ON569692/ON569780 |
| ARY (H17) | Russia, Altai Republic, Ongudaysky District, 12 km west of Yelo, Boltenkov et al. 17 (VBGI) | 50.79055, 85.35777 | ON569517/ON569605/ON569693/ON569781 |
| AUS (H17) | Russia, Altai Republic, Ust-Kansky District, east of Ust-Kan, Boltenkov et al. 13 (VBGI) | 50.94722, 84.82944 | ON569518/ON569606/ON569694/ON569782 |
| I. ivanovae Doronkin | |||
| BNV (H17) | Russia, Buryatia, Novoselenginsk Village, Boltenkov 58 (VBGI) | 51.01166, 106.64027 | ON569523/ON569611/ON569699/ON569787 |
| BAZ (H18) | Russia, Buryatia, Kyakhtinsky District, shtab-lekarskaya zaimka, Boltenkov 59 (VBGI) | 50.38027, 106.55861 | ON569524/ON569612/ON569700/ON569788 |
| BMK (H18) | Russia, Buryatia, Maly Kunaley Village, Boltenkov 113 (VBGI) | 50.61361, 107.83111 | ON569525/ON569613/ON569701/ON569789 |
| ZKL (H17) | Russia, Zabaykalsky Krai, Aginsky District, Lake Khaptsagaytuy, Boltenkov 72 (VBGI) | 50.6167, 114.88777 | ON569520/ON569608/ON569696/ON569784 |
| ZKV (H18) | Russia, Zabaykalsky Krai, Kharanor Village, Boltenkov 93 (VBGI) ** | 50.04666, 116.8225 | ON569521/ON569609/ON569697/ON569785 |
| ZBT (H18) | Russia, Zabaykalsky Krai, Bol’shaya Tura Village, Boltenkov 66 (VBGI) | 51.63111, 114.0383 | ON569519/ON569607/ON569695/ON569783 |
| ZSM (H18) | Russia, Zabaykalsky Krai, Soktui-Milozan Village, Boltenkov 102 (VBGI) | 50.09916, 117.85861 | ON569522/ON569610/ON569698/ON569786 |
| I. goniocarpa Baker | |||
| CSS | China, Sichuan, Songpang County, Shuijing Village, T.G. Elumeeva s.n. (MW0735242) | 32.98123, 103.68576 | ON569526/ON569614/ON569702/ON569790 |
| CGJ | China, Gansu, Jonê County, Wanmaoxiang, SQAE 85 (E) | 34.8013, 103.20255 | ON569527/ON569615/ON569703/ON569791 |
| I. potaninii var. ionantha Y.T.Zhao | |||
| CQX | China, Qinghai, Xinghai County, northern slope of Jiangluling, D.G. Long et al. 148 (E00141064!) | 35.56576, 99.98481 | ON569530/ON569618/ON569706/ON569794 |
| Iris sp. | |||
| CQM | China, Qinghai, Madoi County, Heihe Town, M. Sun s.n. (NENU) | 34.797384, 98.133337 | ON569528/ON569616/ON569704/ON569792 |
| CSJ | China, Sichuan, Jiulong County, Tributary valley SW of Jiulong – Wuxuhai road, s.coll. 342 (E00424870!) | 28.96863, 101.40036 | ON569529/ON569617/ON569705/ON569793 |
| Outgroup specimens | |||
| I. subgen. Pardanthopsis (Hance) Baker | |||
| I. dichotoma Pall. | Russia, Amur Oblast, M. Baranova s.n. (LE, cult.) | – | LT978555/LT981297/LT984447/LT984483 |
| I. subgen. Limniris (Tausch) Spach | |||
| I. ser. Lacteae Doronkin | |||
| I. lactea Pall. | Russia, Zabaykalsky Krai, Kharanor, Chernova s.n. (IRK) | – | LT627854/LN871708/LN871662/LN871625 |
| I. oxypetala Bunge | China, Shaanxi, Suyde, Kabanov s.n. (LE) | – | LT627844/LT627950/LT627975/LT627911 |
| I. tibetica (Dykes) Bolt. | China, Qinghai, Xining to Ta Er, Long et al. 3 (E) | – | LT627893/LT627939/LT627998/LT627933 |
| I. ser. Laevigatae (Diels) G.H.M.Lawr. | |||
| I. ensata Thunb. | Russia, Primorsky Krai, Zarubino, Boltenkov s.n. (VBGI) | – | LT628002/LT628022/LT628012/LT627896 |
| I. laevigata Fisch. | Russia, Primorsky Krai, Roshchino, Pshennikova s.n. (VBGI) | – | LT628003/LT628024/LT628013/LT627897 |
| I. pseudacorus L. | Russia, Vladivostok, Boltenkov s.n. (VBGI) | – | LT628004/LT628025/LT628014/LT627898 |
| I. ser. Ruthenicae (Diels) G.H.M.Lawr. | |||
| I. uniflora | Russian Federation, Primorsky Krai, Zarubino, Boltenkov s.n. (VBGI) | – | LT628002/LT628022/LT628012/LT627896 |
| I. ser. Sibiricae (Diels) G.H.M.Lawr. | |||
| I. sibirica L. | Mongolia, Dornod, Bayan-Uul, Gubanov 550 (MW) | – | LT978556/LT981298/LT984448/LT984480 |
| I. bulleyana Dykes | China, Yunnan, Zhongdian, M.G. Pimenov et al. 432 (MW) | – | LT627895/LT628011/LT628021/LT628001 |
| I. bulleyana f. chrysographes (Dykes) Bolt. | China, Sichuan, Jiulong, Sichuan Expedition 331 (E) | – | LR597328/LR597344/LR597360/LR597376 |
| I. bulleyana f. forrestii (Dykes) Bolt. | China, Yunnan, Lijiang, Yulong Xueshan, P. Cox et al. 2633 (E, cult.) | – | LT978553/LT981295/LT984445/LT984478 |
| I. delavayi Micheli | China, Yunnan, Dali Xian, Yinglofen, Sino-Amer. Bot. Expedition 959 (MHA) | – | LT978552/LT981294/LT984444/LT984477 |
| I. clarkei Baker ex Hook.f. | Nepal, Trogsindho Pass, E.F. Needham 674 (E, cult.) | – | LR597338/LR597354/LR597370/LR597386 |
| I. wilsonii C.H.Wright | China, Yunnan, Little Zhongdian, E.J. Cowley 566 (Kew no. 1990-3457, cult.) | – | LR597339/LR597355/LR597371/LR597387 |
| No. | Character | I. humilis | I. bloudowii | I. vorobievii | I. potaninii | I. tigridia |
|---|---|---|---|---|---|---|
| 1 | Rhizome shape | Creeping | Creeping | Shortened | Compact | Creeping |
| 2 | Rhizome diameter | 0.35–0.8 | 0.4–1 | 0.5–1 | 0.45–1 | 0.25–0.65 |
| 3 | Root shape | Equal | Equal | Obconical | Contractile | Contractile |
| 4 | Root diameter | 0.05–0.2 | 0.06–0.24 | 0.12–0.27 | 0.07–0.34 | 0.12–0.44 |
| 5 | Leaf shape | Ensiform or subfalcate | Subfalcate | Subfalcate | Ensiform | Ensiform or subfalcate |
| 6 | Leaf apex | Acute, straight or incurved | Acute, incurved | Acute, incurved or straight | Narrowly acute, straight or incurved | Narrowly acute or acute, straight |
| 7 | Leaf texture | Thin, smooth | Thin, smooth | Thin, ribbed | Tough, smooth | Tough, smooth |
| 8 | Leaf length | 10.5–30 | 21–50 | 19–60 | 5–29 | 9.5–30 |
| 9 | Leaf width | 0.2–1.7 | 0.4–1.8 | 0.5–1.7 | 0.1–0.5 | 0.1–0.6 |
| 10 | Stem height | 2–19.5 | 6–28.5 | 6–25 | 0.5–2.5 | 2.5–20 |
| 11 | Stem branching | 0 | 0 | 0–2 | 0 | 0 |
| 12 | Number of flowers | 2–3 | 1–2 | 2–4 | 1 | 1 |
| 13 | Cauline leaf length | 4.3–14 | 11–19 | 5.2–13.5 | 2.5–6 | 3–10 |
| 14 | Number of bracteoles | (0) 1–3 | 1 | 1 | 0 | 0 |
| 15 | Bract length | 2–5.6 | 2.5–5.6 | 2.5–6.5 | 2.5–6 | 3–4.5 |
| 16 | Bract texture | Tough | Tough | Tough | Thin | Thin |
| 17 | Pedicel length | 0.2–3 | 0.8–6 | 0.5–1.5 | 0–0.2 | 0–0.7 |
| 18 | Tube length | 0.5–1.5 | 0.9–1.8 | 0.5–1.6 | 3.5–5.7 | 1.5–2.5 |
| 19 | Flower color | Yellow (white) | Yellow | Light yellow | Yellow | Blue to violet (white) |
| 20 | Fruit length | 2.7–6.5 | 3.3–6.5 | 5.2–6.5 | 2–4 | 2–4 |
| 21 | Fruit width | 1–2 | 1.3–2.5 | 1.2–2.4 | 0.8–2 | 0.8–1.8 |
| 22 | Fruit shape | Elliptical, tapering at apex | Elliptical | Oblong-elliptical, tapering at apex | Elliptical, apex obtuse | Elliptical, tapering at apex |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boltenkov, E.V.; Artyukova, E.V. New Approach to the Systematics of the Section Psammiris (Iris, Iridaceae): What Does Chloroplast DNA Sequence Tell Us? Plants 2023, 12, 1254. https://doi.org/10.3390/plants12061254
Boltenkov EV, Artyukova EV. New Approach to the Systematics of the Section Psammiris (Iris, Iridaceae): What Does Chloroplast DNA Sequence Tell Us? Plants. 2023; 12(6):1254. https://doi.org/10.3390/plants12061254
Chicago/Turabian StyleBoltenkov, Eugeny V., and Elena V. Artyukova. 2023. "New Approach to the Systematics of the Section Psammiris (Iris, Iridaceae): What Does Chloroplast DNA Sequence Tell Us?" Plants 12, no. 6: 1254. https://doi.org/10.3390/plants12061254
APA StyleBoltenkov, E. V., & Artyukova, E. V. (2023). New Approach to the Systematics of the Section Psammiris (Iris, Iridaceae): What Does Chloroplast DNA Sequence Tell Us? Plants, 12(6), 1254. https://doi.org/10.3390/plants12061254