
Autochthonous Apple Cultivars from the Campania Region (Southern Italy): Bio-Agronomic and Qualitative Traits

 ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Results and Discussion
2.1. Morphological, Pomological and Physicochemical Traits
2.2. Correlations among the Morphological and Qualitative Traits
2.3. Principal Component Analysis
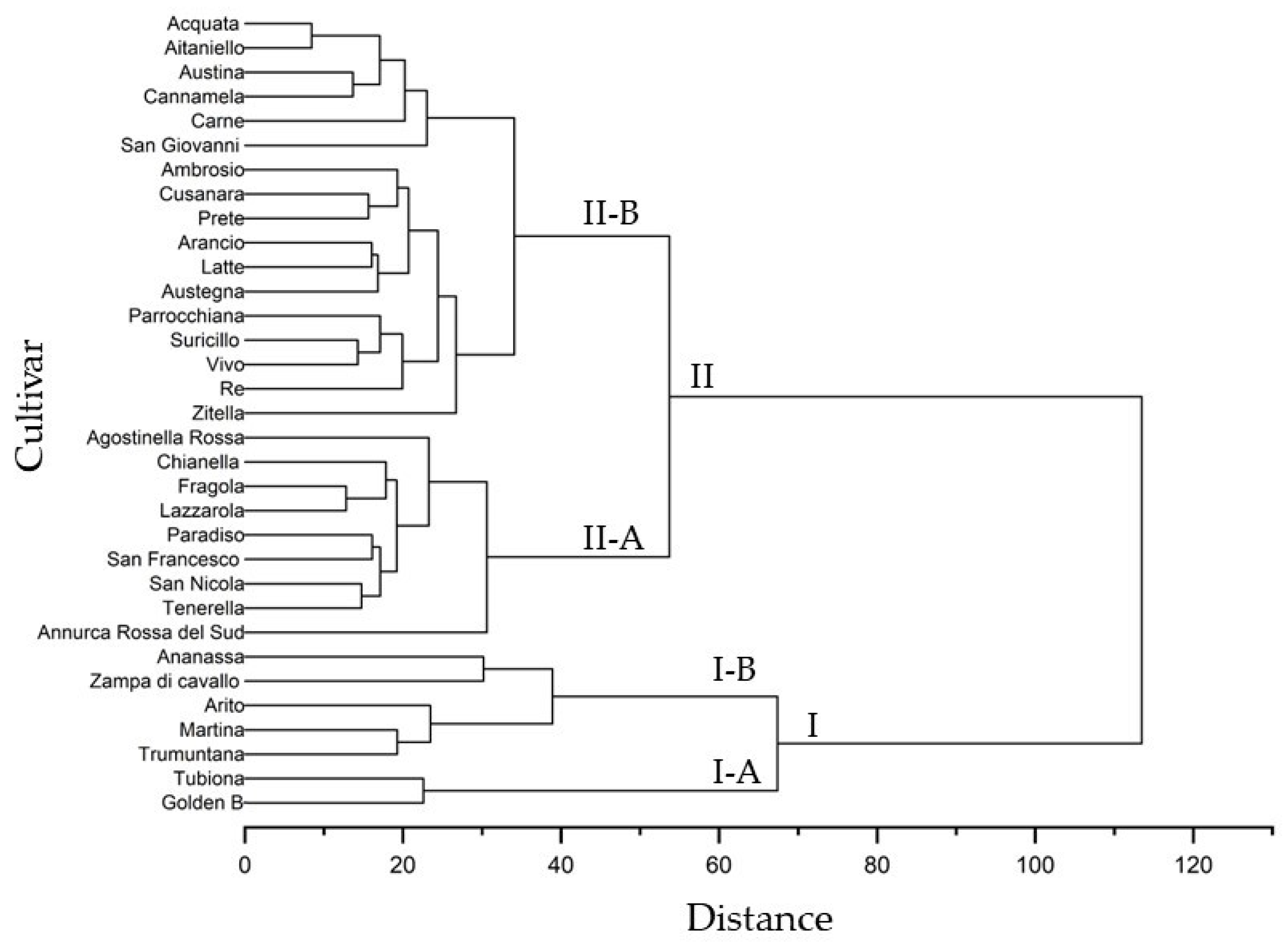
2.4. Cluster Analyses
3. Materials and Methods
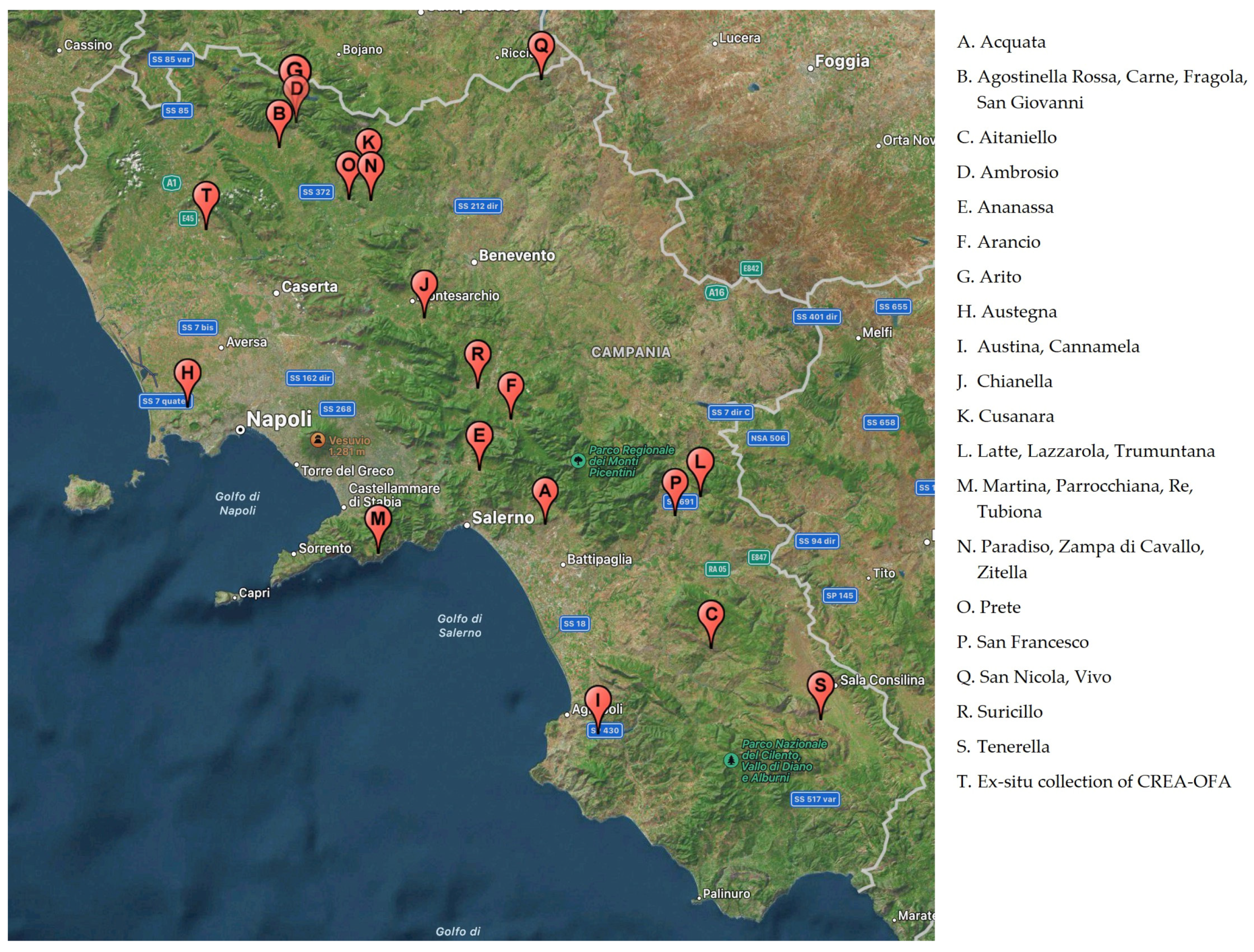
3.1. Plant Materials
3.2. Morphological Traits
3.3. Pomological and Physicochemical Analyses
3.4. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Harris, S.A.; Robinson, J.P.; Juniper, B.E. Genetic clues to the origin of the Apple. Trends Genet. 2002, 18, 426–430. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT 2021. Available online: https://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 6 February 2023).
- FAO2010; The Second Report on the State of the World’s. Plant Genetic Resource for Food and Agriculture: Rome, Italy, 2010.
- Hammer, K.; Gladis, T.; Diederichsen, A. In situ and on farm management of plant genetic resources. Eur. J. Agron. 2003, 19, 509–517. [Google Scholar] [CrossRef]
- Bhatia, S. Application of plant biotechnology. In Modern Applications of Plant Biotechnology in Pharmaceutical Sciences, 1st ed.; Academic Press: Cambridge, UK, 2015. [Google Scholar]
- Migicovsky, Z.; Myles, S. Exploiting wild relatives for genomics assisted breeding of perennial crops. Front. Plant Sci. 2017, 8, 460. [Google Scholar] [CrossRef] [PubMed]
- Petriccione, M.; Akkak, A.; Branca, L.; Rega, P. Caratterizzazione agronomica e molecolare delle mele dell’Irpina. Frutticoltura 2012, 11, 58–64. [Google Scholar]
- Illiano, A.; Pinto, G.; Carrera, M.A.; Palmese, A.; Di Novella, R.; Casoria, P.; Amoresano, A. LC–ms/MS-based quantification method of polyphenols for valorization of ancient apple cultivars from Cilento. ACS Food Sci. Technol. 2022, 2, 647–654. [Google Scholar] [CrossRef]
- Donno, D.; Beccaro, G.L.; Mellano, M.G.; Torello Marinoni, D.; Cerutti, A.K.; Canterino, S.; Bounous, G. Application of sensory, nutraceutical and genetic techniques to create a quality profile of ancient apple cultivars. J. Food Qual. 2012, 35, 169–181. [Google Scholar] [CrossRef]
- Cerrutti, A.K.; Bruun, S.; Donno, D.; Beccaro, G.L.; Bounous, G. Environmental sustainability of traditional foods: The case of ancient apple cultivars in Northern Italy assessed by multifunctional LCA. J. Clean. Prod. 2013, 52, 245–252. [Google Scholar] [CrossRef]
- Lo Piccolo, E.; Landi, M.; Massai, R.; Remorini, D.; Conte, G.; Guidi, L. Ancient apple cultivars from Garfagnana (Tuscany, Italy): A potential source for “nutrafruit” production. Food Chem. 2019, 294, 518–525. [Google Scholar] [CrossRef]
- Testolin, R.; Foria, S.; Baccichet, I.; Messina, R.; Danuso, F.; Losa, A.; Scarbolo, E.; Stocco, M.; Cipriani, G. Genotyping Apple (Malus x domestica borkh.) heirloom germplasm collected and maintained by the Regional Administration of friuli Venezia Giulia (Italy). Sci. Hortic. 2019, 252, 229–237. [Google Scholar] [CrossRef]
- Volk, G.M.; Bramel, P. Apple Genetic Resources: Diversity and Conservation. In The Apple Genome. Compendium of Plant Genomes, 1st ed.; Korban, S.S., Ed.; Springer Nature: Cham, Switzerland, 2021; pp. 33–45. [Google Scholar]
- Panzella, L.; Petriccione, M.; Rega, P.; Scortichini, M.; Napolitano, A. A reappraisal of traditional apple cultivars from southern Italy as a rich source of phenols with superior antioxidant activity. Food Chem. 2013, 140, 672–679. [Google Scholar] [CrossRef]
- D’Abrosca, B.; Scognamiglio, M.; Corrado, L.; Chiocchio, I.; Zampella, L.; Mastrobuoni, F.; Rega, P.; Scortichini, M.; Fiorentino, A.; Petriccione, M. Evaluation of different training systems on Annurca apple fruits revealed by agronomical, qualitative and NMR-based metabolomic approaches. Food Chem. 2017, 222, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Božović, D.; Lazović, B.; Ercisli, S.; Adakalić, M.; Jaćimović, V.; Sezer, I.; Koc, A. Morphological characterization of autochthonous Apple Genetic Resources in Montenegro. Erwerbs-Obstbau 2015, 58, 93–102. [Google Scholar] [CrossRef]
- Höfer, M.; Eldin Ali, M.A.; Sellmann, J.; Peil, A. Phenotypic evaluation and characterization of a collection of Malus species. Genet. Resour. Crop Evol. 2014, 61, 943–964. [Google Scholar] [CrossRef]
- Wagner, I.; Maurer, W.D.; Lemmer, P.; Schmitt, H.P.; Wagner, M. Hybridization and Genetic Diversity in Wild Apple (Malus sylvestris (L.) Mill.) from Various Regions in Germany and from Luxembourg. Silvae Genet. 2014, 63, 81–94. [Google Scholar] [CrossRef]
- Rotondi, A.; Magli, M.; Ricciolini, C.; Baldoni, L. Morphological and molecular analyses for the characterization of a group of Italian olive cultivars. Euphytica 2003, 132, 129–137. [Google Scholar] [CrossRef]
- Watkins, R.; Smith, R.A. Descriptors List for Apple (Malus); International Board for Plant Genetic Resources (IBPGR); Commission of the European Communities (CEC): Wageningen, The Netherlands, 1982. [Google Scholar]
- UPOV. Apple (Malus domestica Borkh.). In Guidelines for the Conduct of Tests for Distinctness, Uniformity and Stability; International Union for the Protection of New Varieties of Plants (UPOV): Geneva, Switzerland, 2005; p. 25. [Google Scholar]
- Gaši, F.; Šimon, S.; Pojskić, N.; Kurtović, M.; Pejić, I. Analysis of morphological variability in Bosnia and Herzegovina’s autochthonous apple germplasm. J. Food Agric. Environ. 2011, 9, 444–448. [Google Scholar]
- Watts, S.; Migicovsky, Z.; McClure, K.A.; Yu, C.H.; Amyotte, B.; Baker, T.; Bowlby, D.; Burgher-MacLellan, K.; Butler, L.; Donald, R.; et al. Quantifying apple diversity: A phenomic characterization of Canada’s Apple Biodiversity Collection. Plants People Planet 2021, 3, 747–760. [Google Scholar] [CrossRef]
- Király, I.; Ladányi, M.; Nagyistván, O.; Tóth, M. Assessment of diversity in a Hungarian apple gene bank using morphological markers. Org. Agric. 2015, 5, 143–151. [Google Scholar] [CrossRef]
- Dolker, T.; Kumar, D.; Chandel, J.S.; Angmo, S.; Chaurasia, O.P.; Stobdan, T. Phenological and Pomological Characteristics of native Apple (Malus domestica Borkh) cultivars of Trans Himalayan ladakh India. Def. Life Sci. J. 2021, 6, 64–69. [Google Scholar] [CrossRef]
- Farrokhi, J.; Darvishzadeh, R.; Hatami Maleki, H.; Naseri, L. Evaluation of Iranian nativeapple (Malus domestica Borkh.) germplasm using biochemical and morphological characteristics. Agric. Conspec. Sci. 2013, 78, 307–313. [Google Scholar]
- Martinelli, F.; Busconi, M.; Camangi, F.; Fogher, C.; Stefani, A. Ancient pomoideae (Malus domestica Borkh. and Pyrus communis L.) cultivars in “Appenino toscano. Caryologia 2008, 61, 320–331. [Google Scholar]
- Dar, J.A.; Wani, A.A.; Dhar, M.K. Morphological, biochemical and male-meiotic characterization of apple (Malus x domestica Borkh.) germplasm of Kashmir Valley. Chromosome Bot. 2015, 10, 39–49. [Google Scholar] [CrossRef]
- Kiprijanovski, M.; Arsov, T.; Saraginovski, N. Pomological, quality and organoleptic traits of some autochthonous apple cultivars in Prespa region, North Macedonia. Acta Hortic. 2020, 1289, 35–42. [Google Scholar] [CrossRef]
- Mratinić, E.; Akšić, M.F. Phenotypic diversity of Apple (Malus sp.) germplasm in South Serbia. Braz. Arch. Biol. Technol. 2012, 55, 349–358. [Google Scholar] [CrossRef]
- Karatas, N. Morphological, sensory and biochemical characteristics of summer apple genotypes. Braz. J. Biol. 2022, 82, 1–10. [Google Scholar] [CrossRef]
- Vavilov, N.I. Centres of origin of cultivated plants. Bull. Appl. Bot. Genet. Plant Breed. 1926, 16, 248. [Google Scholar]
- Maly, K.; Hammer, C.O. Lehmann Sammlung pflanzlicher genetischer Ressourcen in Süditalien ein Reisebericht aus dem Jahre 1950 mit Bemerkungen zur Erhaltung der Landsorten “in situ” und in der Genbank. Kulturpflanze 1987, 35, 109–134. [Google Scholar] [CrossRef]
- Hammer, K.; Knüpffer, H.; Xhuveli, L.; Perrino, P. Estimating genetic erosion in landraces—Two case studies. Genet. Resour. Crop Evol. 1996, 43, 329–336. [Google Scholar] [CrossRef]
- Petriccione, M.; Zampella, L.; Rega, P. Recuperato il patrimonio genetico di antiche mele di origine campana. Frutticoltura 2018, 8, 66–71. [Google Scholar]
- Muccillo, L.; Colantuoni, V.; Sciarrillo, R.; Baiamonte, G.; Salerno, G.; Marziano, M.; Sabatino, L.; Guarino, C. Molecular and environmental analysis of campania (Italy) sweet cherry (Prunus avium L.) cultivars for biocultural refugia identification and conservation. Sci. Rep. 2019, 9, 6796. [Google Scholar] [CrossRef]
- Nesme, T.; Plenet, D.; Hucbourg, B.; Fandos, G.; Lauri, P.E. A set of vegetative morphological variables to objectively estimate apple (Malus× domestica) tree orchard vigour. Sci. Hortic. 2005, 106, 76–90. [Google Scholar] [CrossRef]
- Asma, B.M.; Ozturk, K. Analysis of morphological, pomological and yield characteristics of some apricot germplasm in Turkey. Genet. Resour. Crop Evol. 2005, 52, 305–313. [Google Scholar] [CrossRef]
- Arnal, A.; Lazaro, A.; Tardío, J. Morphological characterization of 23 Malus domestica Borkh cultivars from central Spain. Plant Genet. Resour. 2022, 3, 22–37. [Google Scholar] [CrossRef]
- Zovko, M.; Vego, D.; Zovko, M. Pomological properties of autochthonous cultivars in the area Žepče. Glas. Zast. Bilja 2010, 33, 54–74. [Google Scholar]
- Takos, A.M.; Jaffe, F.W.; Jacob, S.R.; Bogs, J.; Robinson, S.P.; Walker, A.R. Light-induced expression of a MYB gene regulates anthocyanin biosynthesis in red apples. Plant Physiol. 2006, 142, 1216–1232. [Google Scholar] [CrossRef] [PubMed]
- Lin-Wang, K.; Micheletti, D.; Palmer, J.; Volz, R.; Lozano, L.; Espley, R.; Hellens, R.P.; Chagne, D.; Rowan, D.D.; Troggio, M.; et al. High temperature reduces apple fruit colour via modulation of the anthocyanin regulatory complex. Plant Cell Environ. 2011, 34, 1176–1190. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Keller, B.; Roth, M.; Aranzana, M.J.; Auwerkerken, A.; Guerra, W.; Al-Rifaï, M.; Lewandowski, M.; Sanin, N.; Rymenants, M.; et al. Genetic architecture and genomic predictive ability of apple quantitative traits across environments. Hort. Res. 2022, 9, uhac273. [Google Scholar] [CrossRef]
- Kaya, T.; Balta, F.; Şensoy, S. Fruit quality parameters and molecular analysis of Apple Germplasm Resources from Van Lake Basin, Turkey. Turk. J. Agric. For. 2015, 39, 864–875. [Google Scholar] [CrossRef]
- Hassan, S.; Bhat, K.M.; Dar, Z.A.; Mir, M.A.; Pandith, A.H.; Wani, W.M.; Jan, A. Morphological characterization of apple accessions in Kashmir region. Plant Arch. 2017, 17, 1071–1077. [Google Scholar]
- Pasquariello, M.S.; Rega, P.; Migliozzi, T.; Capuano, L.R.; Scortichini, M.; Petriccione, M. Effect of cold storage and shelf life on physiological and quality traits of early ripening pear cultivars. Sci. Hortic. 2013, 162, 341–350. [Google Scholar] [CrossRef]
- Valero, D.; Serrano, M. Growth and ripening stage at harvest modulates postharvest quality and bioactive compounds with antioxidant activity. Stewart Postharvest Rev. 2013, 3, 5. [Google Scholar]
- Zhang, B.; Han, Y. Genomics of Fruit Acidity and Sugar Content in Apple. In The Apple Genome. Compendium of Plant Genomes, 1st ed.; Korban, S.S., Ed.; Springer Nature: Cham, Switzerland, 2021; pp. 297–309. [Google Scholar]
- Koike, H.; Tamai, H.; Ono, T.; Shigehara, I. Influence of time of thinning on yield, fruit quality and return flowering of ‘Fuji’ apple. J. Am. Pom. Soc. 2003, 57, 169–173. [Google Scholar]
- Erdoğan, Ü.G.; Bolat, İ. Investigation on fenological and pomological properties of apple varieties grown in Çoruh valley. Bahçe 2002, 31, 25–32. [Google Scholar]
- Bostan, S.Z. Pomological traits of local apple and pear cultivars and types grown in Trabzon province (eastern Black Sea Region of Turkey). Acta Hort. 2009, 825, 293–298. [Google Scholar] [CrossRef]
- Fathizadeh, Z.; Aboonajmi, M.; Hassan-Beygi, S.R. Nondestructive methods for determining the firmness of apple fruit flesh. Inf. Process. Agric. 2021, 8, 515–527. [Google Scholar] [CrossRef]
- Kubìk, L.; Kažimírovà, V. Determination of Mechanical Properties of Apple cultivar Golden Delicious. J. Process. Energy Agric. 2015, 19, 17–20. [Google Scholar]
- Grotte, M.; Duprat, F.; Piétri, E.; Loonis, D. Young’s modulus, Poisson’s ratio, and lame’s coefficients of Golden Delicious Apple. Int. J. Food Prop. 2002, 5, 333–349. [Google Scholar] [CrossRef]
- Kunihisa, M.; Hayashi, T.; Hatsuyama, Y.; Fukasawa-Akada, T.; Uenishi, H.; Matsumoto, T.; Kon, T.; Kasai, S.; Kudo, T.; Oshino, H.; et al. Genome-wide association study for Apple Flesh browning: Detection, validation, and physiological roles of qtls. Tree Genet. Genomes 2021, 17, 11. [Google Scholar] [CrossRef]
- Daccord, N.; Celton, J.M.; Linsmith, G.; Becker, C.; Choisne, N.; Schijlen, E.; Van de Geest, H.; Bianco, L.; Micheletti, D.; Velasco, R.; et al. High-quality de novo assembly of the apple genome and methylome dynamics of early fruit development. Nat. Genet. 2017, 49, 1099–1106. [Google Scholar] [CrossRef]
- Pereira-Lorenzo, S.; Ramos-Cabrer, A.M.; Ascasiíbar-Errasti, J.; Pinñeiro-Andiόn, J. Analysis of apple germplasm in northwestern Spain. J. Am. Soc. Hortic. 2003, 128, 67–84. [Google Scholar] [CrossRef]
- Khadivi-Khub, A.; Barazandeh, M. A morphometric study of autochthonous plum genotypes based on multivariate analysis. Erwerbs-Obstbau 2015, 57, 185–194. [Google Scholar] [CrossRef]
- Khadivi, A.; Mirheidari, F.; Moradi, Y.; Paryan, S. Phenotypic and fruit characterizations of Prunus divaricata ledeb. germplasm from the north of Iran. Sci. Hort. 2020, 261, 109033. [Google Scholar] [CrossRef]
- Khadivi, A.; Safdari, L.; Hajian, M.-H.; Safari, F. Selection of the promising almond (Prunus amygdalus L.) genotypes among seedling origin trees. Sci. Hort. 2019, 256, 108587. [Google Scholar] [CrossRef]
- Pasquariello, M.S.; Mastrobuoni, F.; Di Patre, D.; Zampella, L.; Capuano, L.R.; Scortichini, M.; Petriccione, M. Agronomic, nutraceutical and molecular variability of Feijoa (Acca sellowiana (O. Berg) burret) germplasm. Sci. Hort. 2015, 191, 1–9. [Google Scholar] [CrossRef]
- Muranty, H.; Troggio, M.; Sadok, I.B.; Rifaï, M.A.; Auwerkerken, A.; Banchi, E.; Velasco, R.; Stevanato, P.; van de Weg, W.E.; Di Guardo, M.; et al. Accuracy and responses of genomic selection on Key traits in Apple breeding. Hortic. Res. 2015, 2, 15060. [Google Scholar] [CrossRef]
- Ganopoulos, I.; Tourvas, N.; Xanthopoulou, A.; Aravanopoulos, F.A.; Avramidou, E.; Zambounis, A.; Tsaftaris, A.; Madesis, P.; Sotiropoulos, T.; Koutinas, N. Phenotypic and molecular characterization of apple (Malus × domestica Borkh) genetic resources in Greece. Sci. Agric. 2018, 75, 509–518. [Google Scholar] [CrossRef]
- Magri, A.; Malorni, L.; Cozzolino, R.; Adiletta, G.; Siano, F.; Picariello, G.; Cice, D.; Capriolo, G.; Nunziata, A.; Di Matteo, M.; et al. Agronomic, Physicochemical, Aromatic and Sensory Characterization of Four Sweet Cherry Accessions of the Campania Region. Plants 2023, 12, 610. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Organ | Character | Abbreviation | Unit | Min. | Max. | Mean | SD | CV% |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Tree | Vigor | V | Code | 1 | 7 | 5.48 | 1.23 | 22.39 |
| 2 | Type | T | Code | 2 | 2 | 2 | 0 | 0 | |
| 3 | Only varieties with ramified tree type: habit | TH | Code | 1 | 3 | 1.51 | 0.62 | 40.82 | |
| 4 | Type of bearing | TB | Code | 1 | 3 | 2.24 | 0.75 | 33.50 | |
| 5 | One-year-old shoot | Thickness | THI | Code | 3 | 9 | 4.33 | 1.47 | 33.97 |
| 6 | Length of internode | L | Code | 1 | 7 | 4.57 | 1.20 | 26.22 | |
| 7 | Color on the sunny side | C | Code | 2 | 4 | 2.39 | 0.70 | 29.43 | |
| 8 | Pubescence (on the distal half of the shoot) | P | Code | 1 | 7 | 4.51 | 1.66 | 36.78 | |
| 9 | Number of lenticels | N | Code | 3 | 7 | 3.90 | 1.23 | 31.57 | |
| 10 | Leaf | Attitude in relation to shoot | A | Code | 1 | 3 | 1.12 | 0.41 | 37.03 |
| 11 | Length | LE | Code | 1 | 7 | 4.06 | 1.97 | 48.45 | |
| 12 | Width | W | Code | 3 | 7 | 4.51 | 1.50 | 33.28 | |
| 13 | Ratio length/width | R | Code | 3 | 7 | 5.60 | 1.46 | 25.98 | |
| 14 | Intensity of the green color | I | Code | 3 | 7 | 4.93 | 0.61 | 12.33 | |
| 15 | Incisions of margin (upper half) | IM | Code | 1 | 5 | 3.12 | 0.99 | 31.80 | |
| 16 | Pubescence on the lower side | PU | Code | 1 | 3 | 1.57 | 0.56 | 35.58 | |
| 17 | Petiole: length | PL | Code | 3 | 7 | 5.09 | 1.70 | 33.44 | |
| 18 | Petiole: the extent of anthocyanin coloration from base | PA | Code | 3 | 7 | 3.79 | 1.32 | 34.77 | |
| 19 | Flower | Predominant color at the balloon stage | PC | Code | 1 | 5 | 3.27 | 0.98 | 29.85 |
| 20 | Diameter with petals pressed into a horizontal position | D | Code | 5 | 7 | 5.06 | 0.35 | 6.88 | |
| 21 | Arrangement of petals | AP | Code | 1 | 3 | 1.67 | 0.69 | 41.53 | |
| 22 | Position of stigmas relative to anthers | PS | Code | 1 | 3 | 2.39 | 0.70 | 29.43 | |
| 23 | Fruit | Young fruit: the extent of anthocyanin overcolor | Y | Code | 1 | 9 | 1.42 | 1.48 | 103.89 |
| 24 | Size | S | Code | 1 | 9 | 3.58 | 2.95 | 82.44 | |
| 25 | Height | H | Code | 3 | 7 | 4.21 | 1.32 | 31.27 | |
| 26 | Diameter | DI | Code | 3 | 7 | 4.70 | 1.42 | 30.33 | |
| 27 | Ratio height/diameter | RH | Code | 1 | 9 | 4.15 | 2.87 | 69.22 | |
| 28 | General shape | G | Code | 2 | 7 | 5.06 | 2.19 | 43.33 | |
| 29 | Ribbing | RI | Code | 1 | 2 | 1.06 | 0.24 | 22.85 | |
| 30 | Crowning at the calyx end | CC | Code | 1 | 2 | 1.24 | 0.44 | 35.03 | |
| 31 | Size of eye | SE | Code | 3 | 7 | 4.88 | 1.58 | 32.31 | |
| 32 | Length of sepal | LS | Code | 3 | 7 | 3.36 | 0.93 | 27.63 | |
| 33 | Bloom of skin | BS | Code | 1 | 3 | 1.36 | 0.55 | 40.24 | |
| 34 | Greasiness of skin | GS | Code | 1 | 2 | 1.30 | 0.47 | 35.82 | |
| 35 | Ground color | GC | Code | 2 | 6 | 4.76 | 0.66 | 13.93 | |
| 36 | Relative area of overcolor | RA | Code | 1 | 9 | 3.06 | 2.57 | 84.07 | |
| 37 | Hue of overcolor—with bloom removed | HC | Code | 2 | 3 | 2.60 | 0.50 | 19.33 | |
| 38 | Intensity of overcolor | IO | Code | 3 | 5 | 4.27 | 0.98 | 23.05 | |
| 39 | Pattern of overcolor | PO | Code | 1 | 7 | 2.36 | 1.84 | 77.84 | |
| 40 | Width of stripes | WS | Code | 3 | 7 | 4.82 | 1.40 | 29.08 | |
| 41 | Area of russet around stalk attachment | AS | Code | 1 | 3 | 1.45 | 0.71 | 48.89 | |
| 42 | Area of russet on cheeks | AC | Code | 1 | 3 | 1.24 | 0.61 | 49.41 | |
| 43 | Area of russet around eye basin | AE | Code | 1 | 3 | 1.30 | 0.68 | 52.49 | |
| 44 | Number of lenticels | NL | Code | 3 | 7 | 4.58 | 1.56 | 34.13 | |
| 45 | Size of lenticels | SL | Code | 3 | 7 | 4.45 | 1.75 | 39.32 | |
| 46 | Length of stalk | LES | Code | 1 | 7 | 3.67 | 1.63 | 44.54 | |
| 47 | Thickness of stalk | TS | Code | 3 | 7 | 5.00 | 1.11 | 22.36 | |
| 48 | Depth of stalk cavity | DS | Code | 3 | 7 | 4.81 | 1.53 | 31.75 | |
| 49 | Width of stalk cavity | WC | Code | 3 | 7 | 4.21 | 1.49 | 35.49 | |
| 50 | Depth of eye basin | DE | Code | 3 | 7 | 4.39 | 1.17 | 26.65 | |
| 51 | Width of eye basin | WE | Code | 3 | 7 | 4.33 | 1.38 | 31.95 | |
| 52 | Firmness of flesh | F | Code | 3 | 9 | 6.09 | 1.67 | 27.34 | |
| 53 | Color of flesh | CF | Code | 1 | 2 | 1.15 | 0.36 | 31.62 | |
| 54 | Aperture of locules (in transverse section) | AL | Code | 1 | 3 | 1.54 | 0.71 | 46.01 | |
| 55 | Flower | Time of beginning of flowering | TF | Code | 3 | 9 | 5.67 | 1.47 | 25.98 |
| 56 | Fruit | Time for harvest | THA | Code | 1 | 9 | 5.84 | 2.24 | 38.26 |
| 57 | Time of eating maturity | TE | Code | 1 | 9 | 6.12 | 2.30 | 37.60 |
| Cultivar | Growing Area (Italy) | Harvest Time | Fruit Weight (g) | Ground Colour | Overcolour | Firmness (Kg/cm2) | SSC (°Brix) | TA (g Malic Acid L−1) | pH |
|---|---|---|---|---|---|---|---|---|---|
| Acquata | Montecorvino Pugliano (SA) | Late June | 42.40 ± 5.69 (abc) | Green-Yellow | - | 7.64 ± 0.53 (ghilm) | 9.27 ± 0.70 (cd) | 2.36 ± 0.40 (a) | 3.61 ± 0.05 (ilm) |
| Agostinella Rossa | Alife (CE) | Late August | 107.80 ± 13.06 (i) | Whitish-Green | Red | 7.32 ± 1.96 (fghil) | 12.60 ± 0.50 (lmn) | 4.30 ± 0.60 (de) | 2.90 ± 0.05 (abc) |
| Aitaniello | Ottati (SA) | Late June | 37.90 ± 11.12 (ab) | Green-Yellow | - | 7.25 ± 0.74 (fghil) | 9.00 ± 0.40 (bc) | 3.01 ± 0.57 (bc) | 3.58 ± 0.09 (ilm) |
| Ambrosio | Castello Matese (CE) | Late October | 76.60 ± 8.96 (g) | Whitish-Green | Pink-red | 9.28 ± 1.43 (op) | 12.00 ± 0.40 (hil) | 3.00 ± 0.81 (bc) | 3.52 ± 0.07 (hilm) |
| Ananassa | Fisciano (SA) | Late September | 178.70 ± 43.85 (n) | Green-Yellow | - | 8.76 ± 1.14 (no) | 8.20 ± 0.50 (a) | 4.03 ± 0.32 (d) | 3.73 ± 0.05 (mn) |
| Arancio | Serino (AV) | Early October | 110.00 ± 14.48 (fg) | Green-Yellow | - | 7.96 ± 1.92 (hilmn) | 13.80 ± 0.70 (o) | 4.69 ± 0.24 (ef) | 3.69 ± 0.18 (lmn) |
| Arito | San Gregorio Matese (CE) | Early October | 149.17 ± 16.13 (l) | Whitish-Green | Red | 8.59 ± 1.09 (no) | 13.30 ± 0.50 (no) | 3.35 ± 0.46 (c) | 3.39 ± 0.19 (fghi) |
| Austegna | Quarto (NA) | Late August | 61.00 ± 8.86 (ef) | Green-Yellow | Red | 7.29 ± 1.73 (fghil) | 11.56 ± 0.40 (gh) | 4.69 ± 0.16 (ef) | 3.10 ± 0.09 (cde) |
| Austina | Torchiara (SA) | Mid August | 46.93 ± 10.40 (bcd) | Green-Yellow | Pink-red | 6.55 ± 1.38 (def) | 8.40 ± 0.30 (ab) | 3.01 ± 0.53 (bc) | 3.67 ± 0.10 (lmn) |
| Cannamela | Torchiara (SA) | Mid September | 46.60 ± 7.77 (bcd) | Whitish-Green | Pink-red | 5.39 ± 0.83 (abc) | 10.50 ± 0.70 (ef) | 4.13 ± 0.38 (de) | 4.21 ± 0.25 (pq) |
| Carne | Alife (CE) | Mid August | 31.33 ± 5.71 (a) | Green-Yellow | Red | 5.61 ± 2.05 (bc) | 11.90 ± 0.42 (hil) | 2.34 ± 0.19 (a) | 4.05 ± 0.03 (opq) |
| Chianella | San Martino Valle Caudina (BN) | Mid September | 105.80 ± 10.61 (hi) | Green-Yellow | Red | 7.05 ± 1.09 (fgh) | 14.64 ± 0.30 (p) | 7.37 ± 0.29 (i) | 3.04 ± 0.08 (fghil) |
| Cusanara | San Lorenzello (BN) | Early September | 77.50 ± 17.04 (g) | Whitish-Green | - | 4.62 ± 0.68 (a) | 10.60 ± 0.5 (ef) | 2.68 ± 0.11 (ab) | 3.69 ± 0.09 (hilmn) |
| Fragola | Alife (CE) | Mid September | 94.28 ± 19.06 (g) | Green-Yellow | Pink-red | 6.83 ± 1.57 (efg) | 11.60 ± 0.30 (ghi) | 5.02 ± 0.22 (fg) | 3.23 ± 0.08 (defg) |
| Latte | Colliano (SA) | Early October | 63.50 ± 7.24 (ef) | Whitish-Green | - | 7.67 ± 1.29 (ghilm) | 12.00 ± 0.50 (hil) | 8.04 ± 0.32 (l) | 3.45 ± 0.21 (ghilm) |
| Lazzarola | Colliano (SA) | Late September | 97.80 ± 12.38 (h) | Green | Pink-red | 7.32 ± 0.88 (fghil) | 11.40 ± 0.20 (gh) | 5.36 ± 0.22 (g) | 3.28 ± 0.19 (defgh) |
| Martina | Agerola (NA) | Late August | 156.00 ± 18.77 (m) | Green-Yellow | Pink-red | 9.62 ± 1.21 (pq) | 13.00 ± 0.40 (mno) | 5.10 ± 0.30 (fg) | 3.73 ± 0.05 (mn) |
| Paradiso | Castelvenere (BN) | Late September | 103.10 ± 16.16 (hi) | Green-Yellow | Red | 8.17 ± 1.10 (lmn) | 8.60 ± 0.40 (abc) | 2.34 ± 0.49 (a) | 5.28 ± 0.15 (r) |
| Parrocchiana | Agerola (NA) | Late September | 52.40 ± 5.69 (cde) | Green-Yellow | - | 8.35 ± 1.28 (mn) | 11.90 ± 0.20 (hil) | 2.34 ± 0.12 (a) | 3.04 ± 0.15 (bcd) |
| Prete | San Salvatore Telesino (BN) | Late September | 70.27 ± 11.30 (fg) | Green-Yellow | Red | 4.96 ± 0.85 (ab) | 12.50 ± 0.25 (lm) | 2.36 ± 0.34 (a) | 4.27 ± 0.04 (q) |
| Re | Agerola (NA) | Late October | 56.70 ± 8.99 (def) | Whitish-Green | Red | 6.94 ± 0.59 (fg) | 11.50 ± 0.70 (gh) | 7.60 ± 0.20 (il) | 2.90 ± 0.06 (abc) |
| San Francesco | Oliveto Citra (SA) | Early October | 109.40 ± 10.09 (i) | Green-Yellow | - | 9.95 ± 1.03 (pq) | 10.20 ± 0.35 (e) | 5.02 ± 0.23 (fg) | 2.67 ± 0.15 (a) |
| San Giovanni | Alife (CE) | Late June | 32.90 ± 4.14 (a) | Green-Yellow | - | 8.07 ± 0.71 (lmn) | 8.00 ± 0.30 (a) | 3.35 ± 0.14 (c) | 3.57 ± 0.31 (ilm) |
| San Nicola | Castelvetere Valfortore (BN) | Early October | 107.60 ± 9.03 (i) | Green-Yellow | Red | 5.90 ± 0.53 (cd) | 13.50 ± 0.50 (o) | 4.67 ± 0.21 (ef) | 4.01 ± 0.20 (op) |
| Suricillo | Avellino | Mid September | 62.20 ± 8.87 (ef) | Green-Yellow | - | 10.24 ± 1.39 (q) | 11.00 ± 0.40 (fg) | 2.68 ± 0.82 (ab) | 4.30 ± 0.30 (q) |
| Tenerella | Sassano (SA) | Late August | 109.40 ± 19.94 (i) | Green-Yellow | Red | 7.13 ± 1.12 (fghi) | 9.90 ± 0.25 (de) | 5.00 ± 0.34 (efg) | 3.33 ± 0.31 (efghi) |
| Trumuntana | Colliano (SA) | Early August | 156.30 ± 35.20 (m) | Green-Yellow | - | 9.28 ± 0.84 (nopq) | 12.50 ± 0.20 (lm) | 6.70 ± 0.30 (h) | 2.85 ± 0.09 (abc) |
| Tubiona | Agerola (NA) | Mid August | 236.00 ± 31.63 (q) | Green-Yellow | - | 6.07 ± 1.21 (cde) | 12.40 ± 0.10 (ilm) | 4.36 ± 0.73 (de) | 3.18 ± 0.10 (def) |
| Vivo | Castelvetere Valfortore (BN) | Late August | 59.60 ± 5.53 (def) | Green-Yellow | Pink-red | 7.31 ± 1.28 (fghil) | 10.60 ± 0.30 (ef) | 10.38 ± 0.63 (n) | 2.86 ± 0.03 (abc) |
| Zampa di cavallo | Castelvenere (BN) | Mid September | 194.70 ± 43.61 (o) | Green-Yellow | - | 8.40 ± 1.21 (mn) | 13.00 ± 0.30 (mno) | 5.10 ± 0.30 (fg) | 2.80 ± 0.10 (ab) |
| Zitella | Castelvenere (BN) | Late September | 79.20 ± 3.83 (g) | Green-Yellow | Red | 9.39 ± 1.12 (opq) | 10.20 ± 0.30 (e) | 3.02 ± 0.25 (bc) | 4.20 ± 0.10 (pq) |
| Annurca Rossa del Sud | - | Late September | 122.62 ± 11.97 (p) | Green-Yellow | Red | 8.03 ± 1.48 (ilmn) | 13.50 ± 0.36 (hil) | 9.00 ± 0.65 (m) | 3.47 ± 0.02 (ghilm) |
| Golden B | - | Mid September | 223.75 ± 40.94 (l) | Green-Yellow | - | 6.86 ± 0.19 (efg) | 12.00 ± 0.70 (o) | 4.40 ± 0.40 (de) | 3.89 ± 0.04 (no) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cice, D.; Ferrara, E.; Magri, A.; Adiletta, G.; Capriolo, G.; Rega, P.; Di Matteo, M.; Petriccione, M. Autochthonous Apple Cultivars from the Campania Region (Southern Italy): Bio-Agronomic and Qualitative Traits. Plants 2023, 12, 1160. https://doi.org/10.3390/plants12051160
Cice D, Ferrara E, Magri A, Adiletta G, Capriolo G, Rega P, Di Matteo M, Petriccione M. Autochthonous Apple Cultivars from the Campania Region (Southern Italy): Bio-Agronomic and Qualitative Traits. Plants. 2023; 12(5):1160. https://doi.org/10.3390/plants12051160
Chicago/Turabian StyleCice, Danilo, Elvira Ferrara, Anna Magri, Giuseppina Adiletta, Giuseppe Capriolo, Pietro Rega, Marisa Di Matteo, and Milena Petriccione. 2023. "Autochthonous Apple Cultivars from the Campania Region (Southern Italy): Bio-Agronomic and Qualitative Traits" Plants 12, no. 5: 1160. https://doi.org/10.3390/plants12051160
APA StyleCice, D., Ferrara, E., Magri, A., Adiletta, G., Capriolo, G., Rega, P., Di Matteo, M., & Petriccione, M. (2023). Autochthonous Apple Cultivars from the Campania Region (Southern Italy): Bio-Agronomic and Qualitative Traits. Plants, 12(5), 1160. https://doi.org/10.3390/plants12051160