
Morphological and Genetic Clonal Diversity within the ‘Greco Bianco’ Grapevine (Vitis vinifera L.) Variety

 ,
,  ,
,  ,
,  ,
,
Abstract
1. Introduction
2. Results
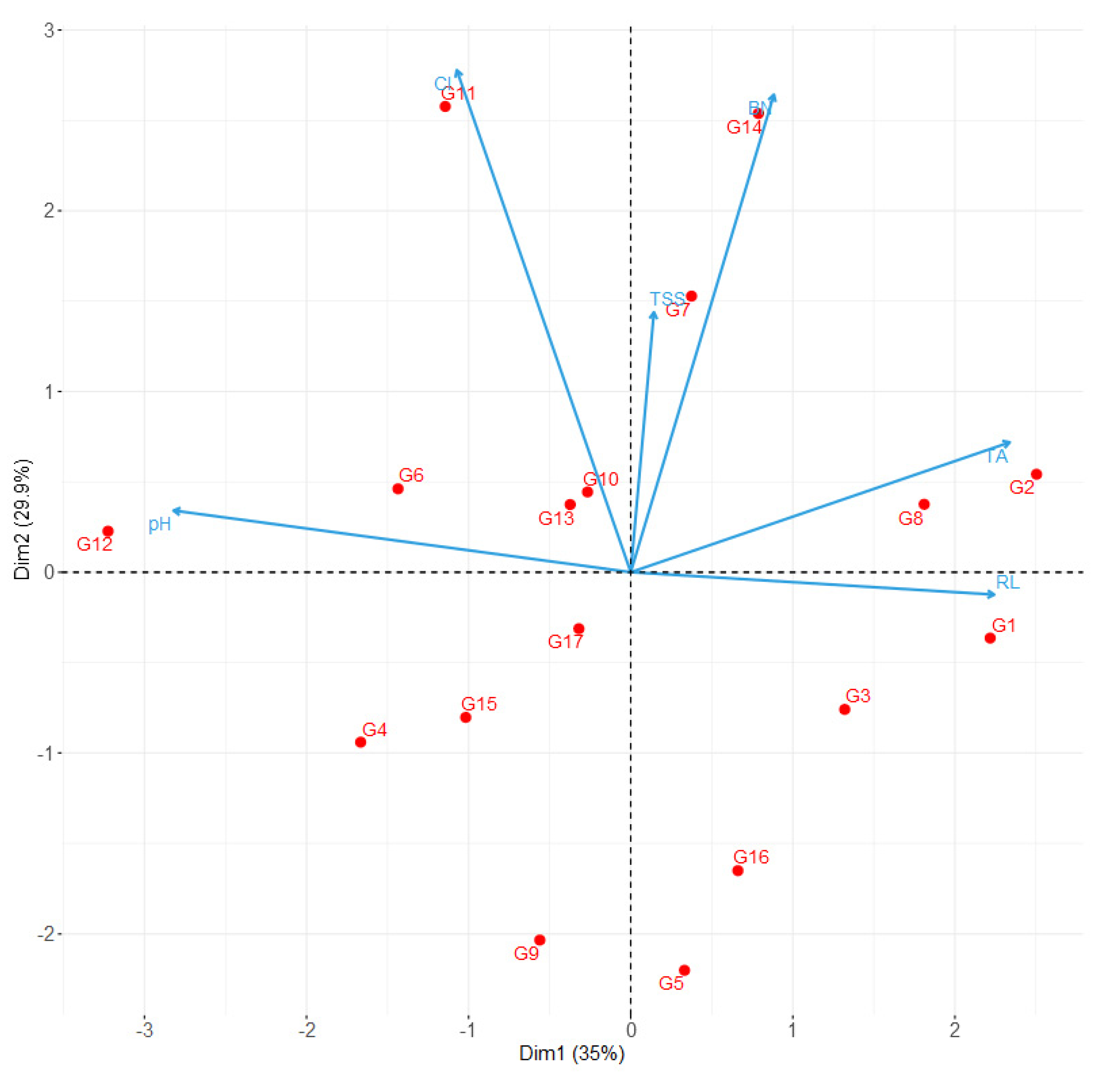
2.1. Diversity in Morphological and Berry Composition Traits
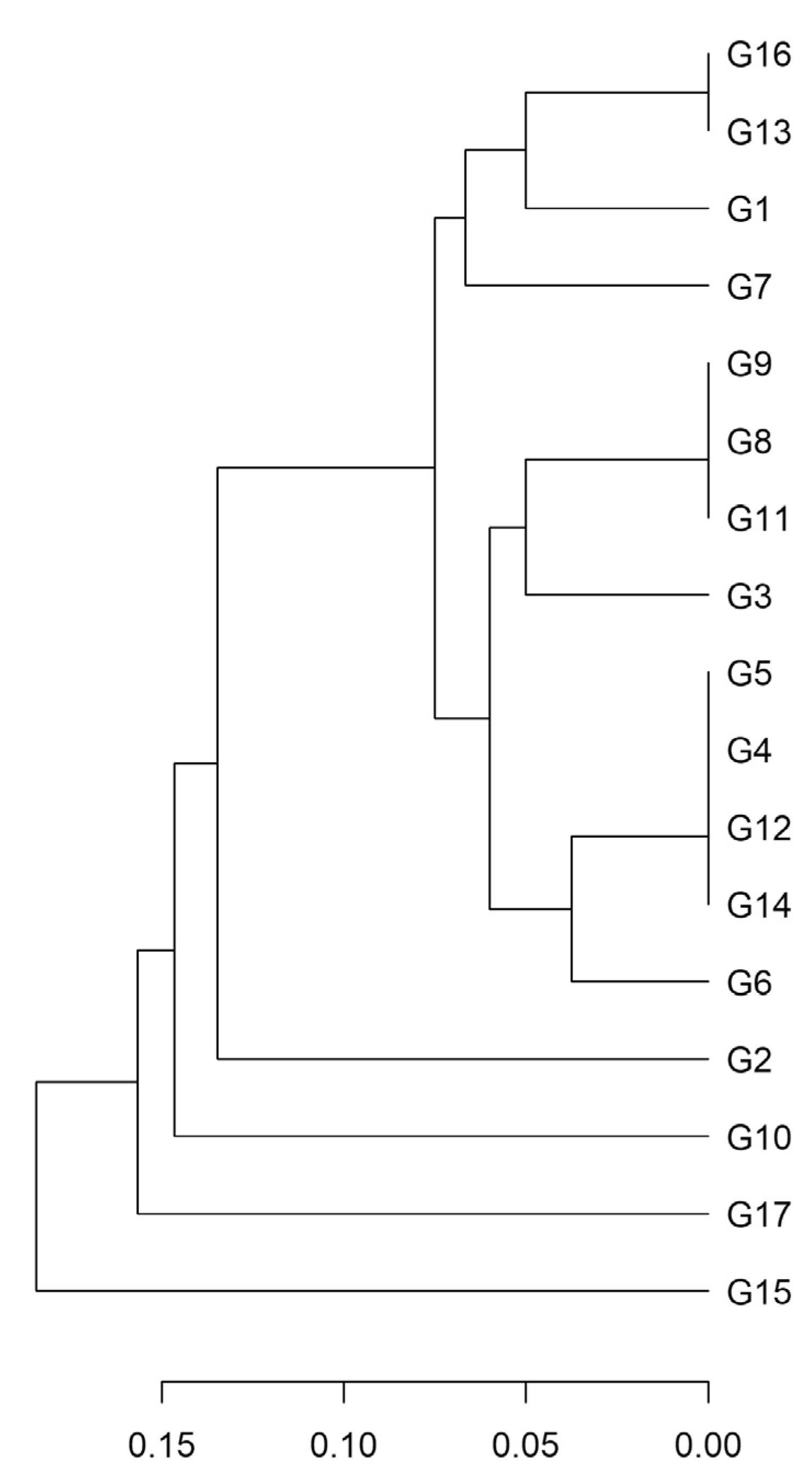
2.2. Genetic Diversity
2.2.1. Microsatellite Analysis
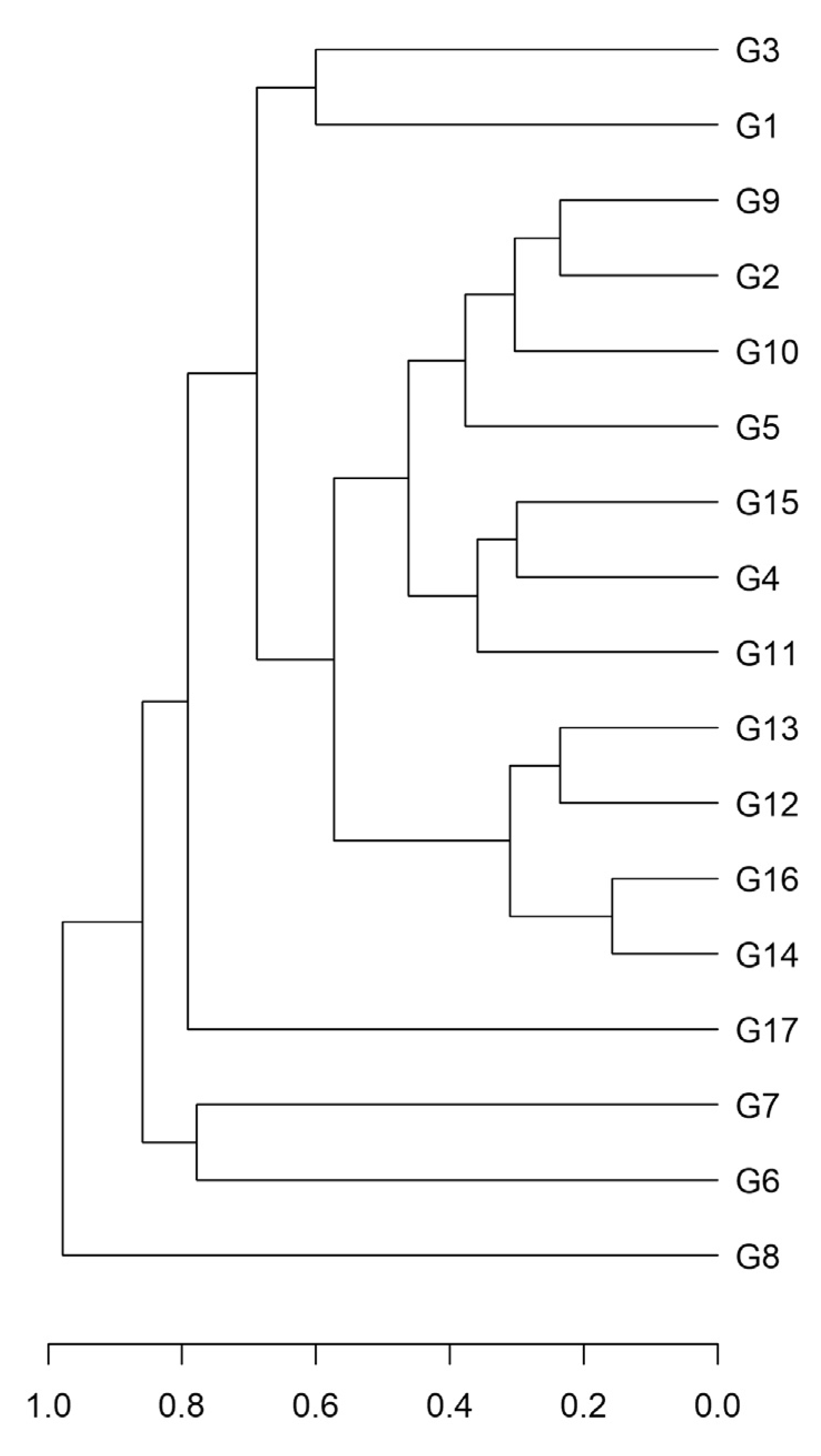
2.2.2. REMAP Analysis
2.3. Relationships between the Classification Obtained with the Different Marker Systems
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Site
4.2. Morphological and Berry Composition Analysis
4.3. Microsatellite Analysis
4.4. REMAP Analysis
4.5. Molecular Data Analysis
4.6. Concordance Test between Marker Systems
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reisch, B.I.; Owens, C.L.; Cousins, P.S. Grape. In Fruit Breeding; Springer: Berlin/Heidelberg, Germany, 2012; pp. 225–262. [Google Scholar]
- McKey, D.; Elias, M.; Pujol, B.; Duputié, A. The evolutionary ecology of clonally propagated domesticated plants. New Phytol. 2010, 186, 318–332. [Google Scholar] [CrossRef]
- Riaz, S.; Garrison, K.E.; Dangl, G.S.; Boursiquot, J.-M.; Meredith, C.P. Genetic divergence and chimerism within ancient asexually propagated winegrape cultivars. J. Am. Soc. Hortic. Sci. 2002, 127, 508–514. [Google Scholar] [CrossRef]
- Vondras, A.M.; Minio, A.; Blanco-Ulate, B.; Figueroa-Balderas, R.; Penn, M.A.; Zhou, Y.; Seymour, D.; Ye, Z.; Liang, D.; Espinoza, L.K. The genomic diversification of grapevine clones. BMC Genom. 2019, 20, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Pelsy, F. Molecular and cellular mechanisms of diversity within grapevine varieties. Heredity 2010, 104, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Chessa, I.; Nieddu, G. Analysis of diversity in the fruit tree genetic resources from a Mediterranean island. Genet. Resour. Crop Evol. 2005, 52, 267–276. [Google Scholar] [CrossRef]
- Leão, P.C.D.S.; Cruz, C.D.; Motoike, S.Y. Genetic diversity of table grape based on morphoagronomic traits. Sci. Agric. 2011, 68, 42–49. [Google Scholar] [CrossRef]
- Vafaee, Y.; Ghaderi, N.; Khadivi, A. Morphological variation and marker-fruit trait associations in a collection of grape (Vitis vinifera L.). Sci. Hortic. 2017, 225, 771–782. [Google Scholar] [CrossRef]
- Abiri, K.; Rezaei, M.; Tahanian, H.; Heidari, P.; Khadivi, A. Morphological and pomological variability of a grape (Vitis vinifera L.) germplasm collection. Sci. Hortic. 2020, 266, 109285. [Google Scholar] [CrossRef]
- Kupe, M.; Sayinci, B.; Demir, B.; Ercisli, S.; Aslan, K.A.; Gundesli, M.A.; Baron, M.; Sochor, J. Multivariate Analysis Approaches for Dimension and Shape Discrimination of Vitis vinifera Varieties. Plants 2021, 10, 1528. [Google Scholar] [CrossRef]
- Muganu, M.; Dangl, G.; Aradhya, M.; Frediani, M.; Scossa, A.; Stover, E. Ampelographic and DNA characterization of local grapevine accessions of the Tuscia area (Latium, Italy). Am. J. Enol. Vitic. 2009, 60, 110–115. [Google Scholar] [CrossRef]
- Villano, C.; Aiese Cigliano, R.; Esposito, S.; D’Amelia, V.; Iovene, M.; Carputo, D.; Aversano, R. DNA-Based Technologies for Grapevine Biodiversity Exploitation: State of the Art and Future Perspectives. Agronomy 2022, 12, 491. [Google Scholar] [CrossRef]
- Mondini, L.; Noorani, A.; Pagnotta, M.A. Assessing plant genetic diversity by molecular tools. Diversity 2009, 1, 19–35. [Google Scholar] [CrossRef]
- Grassi, F.; Labra, M.; Imazio, S.; Spada, A.; Sgorbati, S.; Scienza, A.; Sala, F. Evidence of a secondary grapevine domestication centre detected by SSR analysis. Theor. Appl. Genet. 2003, 107, 1315–1320. [Google Scholar] [CrossRef] [PubMed]
- Labra, M.; Failla, O.; Fossati, T.; Castiglione, S.; Scienza, A.; Sala, F. Phylogenetic analysis of grapevine cv. Ansonica growing on the island of Giglio, Italy, by AFLP and SSR markers. Vitis 1999, 38, 161–166. [Google Scholar]
- Pastore, C.; Fontana, M.; Raimondi, S.; Ruffa, P.; Filippetti, I.; Schneider, A. Genetic characterization of grapevine varieties from Emilia-Romagna (northern Italy) discloses unexplored genetic resources. Am. J. Enol. Vitic. 2020, 71, 334–343. [Google Scholar] [CrossRef]
- De Lorenzis, G.; Imazio, S.; Biagini, B.; Failla, O.; Scienza, A. Pedigree Reconstruction of the Italian Grapevine Aglianico (Vitis vinifera L.) from Campania. Mol. Biotechnol. 2012, 54, 634–642. [Google Scholar] [CrossRef]
- Crespan, M.; Calò, A.; Giannetto, S.; Sparacio, A.; Storchi, P.; Costacurta, A. Sangiovese’ and ‘Garganega’ are two key varieties of the Italian grapevine assortment evolution. Vitis 2008, 47, 97–104. [Google Scholar]
- Raimondi, S.; Tumino, G.; Ruffa, P.; Boccacci, P.; Gambino, G.; Schneider, A. DNA-based genealogy reconstruction of Nebbiolo, Barbera and other ancient grapevine cultivars from northwestern Italy. Sci. Rep. 2020, 10, 15782. [Google Scholar] [CrossRef]
- Imazio, S.; Labra, M.; Grassi, F.; Scienza, A.; Failla, O. Chloroplast microsatellites to investigate the origin of grapevine. Genet. Resour. Crop Evol. 2006, 53, 1003–1011. [Google Scholar] [CrossRef]
- Villano, C.; Carputo, D.; Frusciante, L.; Santoro, X.; Aversano, R. Use of SSR and retrotransposon-based markers to interpret the population structure of native grapevines from southern Italy. Mol. Biotechnol. 2014, 56, 1011–1020. [Google Scholar] [CrossRef]
- Muccillo, L.; Gambuti, A.; Frusciante, L.; Iorizzo, M.; Moio, L.; Raieta, K.; Rinaldi, A.; Colantuoni, V.; Aversano, R. Biochemical features of native red wines and genetic diversity of the corresponding grape varieties from Campania region. Food Chem. 2014, 143, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Kalendar, R.; Schulman, A.H. IRAP and REMAP for retrotransposon-based genotyping and fingerprinting. Nat. Protoc. 2006, 1, 2478–2484. [Google Scholar] [CrossRef] [PubMed]
- Kalendar, R.; Grob, T.; Regina, M.; Suoniemi, A.; Schulman, A. IRAP and REMAP: Two new retrotransposon-based DNA fingerprinting techniques. Theor. Appl. Genet. 1999, 98, 704–711. [Google Scholar] [CrossRef]
- D’Onofrio, C.; De Lorenzis, G.; Giordani, T.; Natali, L.; Cavallini, A.; Scalabrelli, G. Retrotransposon-based molecular markers for grapevine species and cultivars identification. Tree Genet. Genomes 2010, 6, 451–466. [Google Scholar] [CrossRef]
- Pereira, H.S.; Barão, A.; Delgado, M.; Morais-Cecílio, L.; Viegas, W. Genomic analysis of grapevine retrotransposon 1 (Gret1) in Vitis vinifera. Theor. Appl. Genet. 2005, 111, 871–878. [Google Scholar] [CrossRef]
- Kobayashi, S.; Goto-Yamamoto, N.; Hirochika, H. Retrotransposon-induced mutations in grape skin color. Science 2004, 304, 982. [Google Scholar] [CrossRef]
- Viala, P.; Vermorel, V. Traité Général de Viticulture: Ampélographie; Masson et Cie: Paris, France, 1901–1910; Volume 7. [Google Scholar]
- Rohlf, F.J. Consensus indices for comparing classifications. Math. Biosci. 1982, 59, 131–144. [Google Scholar] [CrossRef]
- Zombardo, A.; Meneghetti, S.; Morreale, G.; Calò, A.; Costacurta, A.; Storchi, P. Study of Inter-and Intra-varietal Genetic Variability in Grapevine Cultivars. Plants 2022, 11, 397. [Google Scholar] [CrossRef]
- Gonçalves, E.; Martins, A. Efficient Assessment and Large-Scale Conservation of Intra-Varietal Diversity of Ancient Grapevine Varieties: Case Study Portugal. Plants 2022, 11, 1917. [Google Scholar] [CrossRef]
- Vignani, R.; Bowers, J.; Meredith, C. Microsatellite DNA polymorphism analysis of clones of Vitis vinifera ‘Sangiovese’. Sci. Hortic. 1996, 65, 163–169. [Google Scholar] [CrossRef]
- Banilas, G.; Korkas, E.; Kaldis, P.; Hatzopoulos, P. Olive and grapevine biodiversity in Greece and Cyprus–a review. Clim. Chang. Intercropping Pest Control Benef. Microorg. 2009, 2, 401–428. [Google Scholar]
- Santiago, J.L.; Boso, S.; Gago, P.; Alonso-Villaverde, V.; Martínez, M.C. Molecular and ampelographic characterisation of Vitis vinifera L. “Albariño”, “Savagnin Blanc” and “Caíño Blanco” shows that they are different cultivars. Span. J. Agric. Res. 2007, 5, 333. [Google Scholar] [CrossRef]
- Sabir, A.; Tangolar, S.; Buyukalaca, S.; Kafkas, S. Ampelographic and molecular diversity among grapevine (Vitis spp.) cultivars. Czech J. Genet. Plant Breed. 2009, 45, 160–168. [Google Scholar] [CrossRef]
- Gago, P.; Santiago, J.-L.; Boso, S.; Alonso-Villaverde, V.; Grando, M.S.; Martínez, M.C. Biodiversity and Characterization of Twenty-two Vitis vinifera L. Cultivars in the Northwestern Iberian Peninsula. Am. J. Enol. Vitic. 2009, 60, 293–301. [Google Scholar] [CrossRef]
- Meneghetti, S.; Poljuha, D.; Frare, E.; Costacurta, A.; Morreale, G.; Bavaresco, L.; Calò, A. Inter-and intra-varietal genetic variability in Malvasia cultivars. Mol. Biotechnol. 2012, 50, 189–199. [Google Scholar] [CrossRef]
- Razi, M.; Amiri, M.E.; Darvishzadeh, R.; Doulati Baneh, H.; Alipour, H.; Martínez-Gómez, P. Assessment of genetic diversity of cultivated and wild Iranian grape germplasm using retrotransposon-microsatellite amplified polymorphism (REMAP) markers and pomological traits. Mol. Biol. Rep. 2020, 47, 7593–7606. [Google Scholar] [CrossRef]
- Tello, J.; Ibáñez, J. What do we know about grapevine bunch compactness? A state-of-the-art review. Aust. J. Grape Wine Res. 2018, 24, 6–23. [Google Scholar] [CrossRef]
- Šuklje, K.; Antalick, G.; Buica, A.; Langlois, J.; Coetzee, Z.A.; Gouot, J.; Schmidtke, L.M.; Deloire, A. Clonal differences and impact of defoliation on Sauvignon blanc (Vitis vinifera L.) wines: A chemical and sensory investigation. J. Sci. Food Agric. 2016, 96, 915–926. [Google Scholar] [CrossRef]
- Cipriani, G.; Marrazzo, M.T.; Di Gaspero, G.; Pfeiffer, A.; Morgante, M.; Testolin, R. A set of microsatellite markers with long core repeat optimized for grape (Vitis spp.) genotyping. BMC Plant Biol. 2008, 8, 1–13. [Google Scholar] [CrossRef]
- Laucou, V.; Lacombe, T.; Dechesne, F.; Siret, R.; Bruno, J.-P.; Dessup, M.; Dessup, T.; Ortigosa, P.; Parra, P.; Roux, C. High throughput analysis of grape genetic diversity as a tool for germplasm collection management. Theor. Appl. Genet. 2011, 122, 1233–1245. [Google Scholar] [CrossRef]
- Carrier, G.; Le Cunff, L.; Dereeper, A.; Legrand, D.; Sabot, F.; Bouchez, O.; Audeguin, L.; Boursiquot, J.-M.; This, P. Transposable Elements Are a Major Cause of Somatic Polymorphism in Vitis vinifera L. PLoS ONE 2012, 7, e32973. [Google Scholar] [CrossRef] [PubMed]
- The French–Italian Public Consortium for Grapevine Genome Characterization. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Hocquigny, S.; Pelsy, F.; Dumas, V.; Kindt, S.; Heloir, M.C.; Merdinoglu, D. Diversification within grapevine cultivars goes through chimeric states. Genome 2004, 47, 579–589. [Google Scholar] [CrossRef]
- Moncada, X.; Pelsy, F.; Merdinoglu, D.; Hinrichsen, P. Genetic diversity and geographical dispersal in grapevine clones revealed by microsatellite markers. Genome 2006, 49, 1459–1472. [Google Scholar] [CrossRef]
- Corrado, G.; La Mura, M.; Ambrosino, O.; Pugliano, G.; Varricchio, P.; Rao, R. Relationships of Campanian olive cultivars: Comparative analysis of molecular and phenotypic data. Genome 2009, 52, 692–700. [Google Scholar] [CrossRef] [PubMed]
- Basile, B.; Mataffo, A.; Forlani, M.; Corrado, G. Diversity in Morphometric, Pomological, and Fruit-Quality Traits of Apricot (Prunus armeniaca) Traditional Varieties: Implications for Landrace Differentiation at Regional Scale. Diversity 2022, 14, 608. [Google Scholar] [CrossRef]
- Manco, R.; Basile, B.; Capuozzo, C.; Scognamiglio, P.; Forlani, M.; Rao, R.; Corrado, G. Molecular and phenotypic diversity of traditional European plum (Prunus domestica L.) germplasm of Southern Italy. Sustainability 2019, 11, 4112. [Google Scholar] [CrossRef]
- Sefc, K.M.; Regner, F.; Turetschek, E.; Glössl, J.; Steinkellner, H. Identification of microsatellite sequences in Vitis riparia and their applicability for genotyping of different Vitis species. Genome 1999, 42, 367–373. [Google Scholar] [CrossRef]
- Thomas, M.; Scott, N. Microsatellite repeats in grapevine reveal DNA polymorphisms when analysed as sequence-tagged sites (STSs). Theor. Appl. Genet. 1993, 86, 985–990. [Google Scholar] [CrossRef]
- Merdinoglu, D.; Butterlin, G.; Bevilacqua, L.; Chiquet, V.; Adam-Blondon, A.-F.; Decroocq, S. Development and characterization of a large set of microsatellite markers in grapevine (Vitis vinifera L.) suitable for multiplex PCR. Mol. Breed. 2005, 15, 349–366. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef] [PubMed]
- Prevosti, A.; Ocana, J.; Alonso, G. Distances between populations of Drosophila subobscura, based on chromosome arrangement frequencies. Theor. Appl. Genet. 1975, 45, 231–241. [Google Scholar] [CrossRef]
- Romesburg, C. Cluster Analysis for Researchers; Lulu. com: Morrisville, NC, USA, 2004. [Google Scholar]
- Thioulouse, J.; Chessel, D.; Olivier, J.-M. ADE-4: A multivariate analysis and graphical display software. Stat. Comput. 1997, 7, 75–83. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Clone | Bud Potential Fertility Index | Components of Bunch Compactness | ||
|---|---|---|---|---|
| No Berries Per Bunch | Rachis Length (cm) | CI (No Berries/ cm Rachis) | ||
| G1 | 1.10 ± 0.14 a | 108 ± 23 ab | 21.1 ± 2.6 a | 4.99 ± 0.62 e |
| G2 | 1.00 ± 0.14 a | 114 ± 19 ab | 16.3 ± 0.8 abcd | 6.86 ± 0.85 bcde |
| G3 | 1.00 ± 0.15 a | 86 ± 13 b | 14.4 ± 2.0 abcd | 6.33 ± 1.42 bcde |
| G4 | 1.18 ± 0.18 a | 78 ± 13 b | 12.4 ± 2.0 cd | 6.53 ± 0.97 bcde |
| G5 | 1.27 ± 0.18 a | 77 ± 14 b | 16.0 ± 1.3 abcd | 4.77 ± 0.79 e |
| G6 | 1.19 ± 0.14 a | 110 ± 33 ab | 14.8 ± 2.7 abcd | 7.07 ± 1.14 bcde |
| G7 | 1.33 ± 0.19 a | 137 ± 27 ab | 17.8 ± 3.5 abc | 7.71 ± 0.03 abcd |
| G8 | 1.29 ± 0.167 a | 129 ± 15 ab | 19.9 ± 0.8 ab | 6.43 ± 0.55 bcde |
| G9 | 1.29 ± 0.17 a | 91 ± 20 b | 17.3 ± 2.4 abcd | 5.15 ± 0.47 de |
| G10 | 1.29 ± 0.19 a | 134 ± 1 ab | 19.3 ± 1.3 ab | 7.07 ± 0.45 bcde |
| G11 | 1.17 ± 0.14 a | 133 ± 21 ab | 14.0 ± 2.9 bcd | 9.90 ± 0.77 a |
| G12 | 1.20 ± 0.17 a | 87 ± 12 b | 10.6 ± 1.8 d | 8.28 ± 0.72 abc |
| G13 | 1.12 ± 0.17 a | 118 ± 15 ab | 15.9 ± 0.8 abcd | 7.37 ± 0.69 bcde |
| G14 | 1.13 ± 0.17 a | 157 ± 16 a | 18.0 ± 0.9 abc | 8.64 ± 0.50 ab |
| G15 | 1.29 ± 0.167 a | 91 ± 14 b | 15.9 ± 2.1 abcd | 5.91 ± 0.91 cde |
| G16 | 1.38 ± 0.18 a | 96 ± 5 b | 20.0 ± 1.9 ab | 4.98 ± 0.65 e |
| G17 | 1.19 ± 0.16 a | 91 ± 13 b | 15.9 ± 1.9 abcd | 5.76 ± 0.51 cde |
| C.V. (%) | 9.1 | 21.9 | 16.9 | 21.1 |
| Clone | TSS (°Brix) | pH | TA (g/L Tartaric Acid) | TSS/TA |
|---|---|---|---|---|
| G1 | 24.3 ± 0.3 a | 3.00 ± 0.06 cd | 10.4 ± 0.1 abcd | 2.35 ± 0.03 de |
| G2 | 22.6 ± 0.2 bcd | 2.96 ± 0.04 d | 14.1 ± 0.1 a | 1.60 ± 0.01 i |
| G3 | 21.6 ± 0.2 de | 3.01 ± 0.02 cd | 13.4 ± 2.2 ab | 1.62 ± 0.01 i |
| G4 | 22.3 ± 0.3 cd | 3.15 ± 0.01 abc | 8.9 ± 0.1 cd | 2.52 ± 0.04 bc |
| G5 | 21.1 ± 0.4 ef | 3.07 ± 0.01 abcd | 10.1 ± 0.4 abcd | 2.10 ± 0.04 f |
| G6 | 22.8 ± 0.5 bc | 3.21 ± 0.09 a | 9.3 ± 2.1 bcd | 2.45 ± 0.06 cd |
| G7 | 23.6 ± 0.2 ab | 3.07 ± 0.01 abcd | 9.0 ± 0.3 cd | 2.62 ± 0.02 b |
| G8 | 22.2 ± 0.3 cd | 3.03 ± 0.04 bcd | 11.0 ± 0.4 abcd | 2.03 ± 0.03 fg |
| G9 | 20.2 ± 0.4 f | 3.13 ± 0.04 abc | 8.0 ± 1.3 cd | 2.55 ± 0.05 bc |
| G10 | 21.2 ± 0.2 ef | 3.16 ± 0.01 ab | 8.6 ± 0.1 cd | 2.47 ± 0.026 c |
| G11 | 23.2 ± 0.2 bc | 3.15 ± 0.03 abc | 9.9 ± 0.9 abcd | 2.34 ± 0.02 de |
| G12 | 22.8 ± 0.3 bcd | 3.21 ± 0.04 a | 7.2 ± 0.1 d | 3.16 ± 0.046 a |
| G13 | 21.1 ± 0.6 ef | 3.16 ± 0.07 ab | 10.8 ± 2.7 abcd | 1.95 ± 0.06 gh |
| G14 | 22.5 ± 0.1 cd | 3.13 ± 0.07 abc | 12 ± 2.4 abc | 1.87 ± 0.01 h |
| G15 | 22.5 ± 0.7 bcd | 3.17 ± 0.02 ab | 8.9 ± 0.1 cd | 2.55 ± 0.08 bc |
| G16 | 21.1 ± 0.4 ef | 3.10 ± 0.01 abc | 9.3 ± 0.3 bcd | 2.27 ± 0.04 e |
| G17 | 24.4 ± 0.3 a | 3.13 ± 0.04 abc | 9.6 ± 0.9 bcd | 2.54 ± 0.03 bc |
| C.V. (%) | 5.2 | 2.3 | 18.1 | 17.2 |
| Principal Component | Eigenvalue | Variance Explained (%) | Correlation | ||||||
|---|---|---|---|---|---|---|---|---|---|
| BN | RL | CI | TSS | pH | TA | TSS/TA | |||
| Dim. 1 | 2.10 | 35.0 | 0.28 NS | 0.72 ** | −0.34 NS | 0.05 NS | −0.91 *** | 0.75 *** | 0.28 NS |
| Dim. 2 | 1.79 | 29.9 | 0.85 *** | −0.04 NS | 0.89*** | 0.46 NS | 0.11 NS | 0.23 NS | 0.85 NS |
| Dim. 3 | 1.06 | 17.7 | 0.44 NS | 0.64 ** | −0.10 NS | −0.40 NS | 0.30 NS | −0.44 NS | 0.44 NS |
| Dim. 4 | 0.88 | 14.7 | −0.06 NS | 0.25 NS | −0.24 NS | 0.79 ** | −0.01 NS | −0.37 NS | −0.06 NS |
| Dim. 5 | 0.15 | 2.5 | 0.01 NS | 0.04 NS | −0.12 NS | 0.06 NS | 0.28 NS | 0.23 NS | 0.01 NS |
| Dim. 6 | 0.01 | 0.2 | −0.07 NS | 0.06 NS | 0.07 NS | 0.01 NS | 0.01 NS | 0.01 NS | −0.07 NS |
| Locus | Na | ASR | Ho | He | I | F | E |
|---|---|---|---|---|---|---|---|
| VVS2 | 4 | 128–126 | 1.00 | 0.56 | 0.92 | −0.80 | 0.83 |
| VVMD7 | 4 | 244–250 | 0.65 | 0.58 | 1.05 | −0.12 | 0.75 |
| VRZAG21 | 2 | 188–200 | 1.00 | 0.50 | 0.69 | −1.00 | 1.00 |
| VVS4 | 2 | 166–172 | 1.00 | 0.50 | 0.69 | −1.00 | 1.00 |
| VRZAG29 | 2 | 112–118 | 1.00 | 0.50 | 0.69 | −1.00 | 1.00 |
| VVLN16 | 2 | 149–157 | 1.00 | 0.50 | 0.69 | −1.00 | 1.00 |
| VVLH54 | 3 | 163–167 | 1.00 | 0.53 | 0.82 | −0.88 | 0.90 |
| VVS5 | 4 | 147–167 | 0.06 | 0.28 | 0.60 | 0.78 | 0.47 |
| VVIC05 | 2 | 162–166 | 1.00 | 0.50 | 0.69 | −1.00 | 1.00 |
| VVLP60 | 4 | 312–320 | 1.00 | 0.56 | 0.94 | −0.78 | 0.83 |
| Quantitative | SSR | REMAP | |
|---|---|---|---|
| Quantitative | - | −0.034 (0.54) | −0.074 (0.64) |
| SSR | −0.120 (0.76) | - | −0.084 (0.64) |
| REMAP | −0.150 (0.82) | 0.028 (0.43) | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villano, C.; Corrado, G.; Basile, B.; Di Serio, E.; Mataffo, A.; Ferrara, E.; Aversano, R. Morphological and Genetic Clonal Diversity within the ‘Greco Bianco’ Grapevine (Vitis vinifera L.) Variety. Plants 2023, 12, 515. https://doi.org/10.3390/plants12030515
Villano C, Corrado G, Basile B, Di Serio E, Mataffo A, Ferrara E, Aversano R. Morphological and Genetic Clonal Diversity within the ‘Greco Bianco’ Grapevine (Vitis vinifera L.) Variety. Plants. 2023; 12(3):515. https://doi.org/10.3390/plants12030515
Chicago/Turabian StyleVillano, Clizia, Giandomenico Corrado, Boris Basile, Ermanno Di Serio, Alessandro Mataffo, Elvira Ferrara, and Riccardo Aversano. 2023. "Morphological and Genetic Clonal Diversity within the ‘Greco Bianco’ Grapevine (Vitis vinifera L.) Variety" Plants 12, no. 3: 515. https://doi.org/10.3390/plants12030515
APA StyleVillano, C., Corrado, G., Basile, B., Di Serio, E., Mataffo, A., Ferrara, E., & Aversano, R. (2023). Morphological and Genetic Clonal Diversity within the ‘Greco Bianco’ Grapevine (Vitis vinifera L.) Variety. Plants, 12(3), 515. https://doi.org/10.3390/plants12030515