
Seed Longevity—The Evolution of Knowledge and a Conceptual Framework

 , and
, and 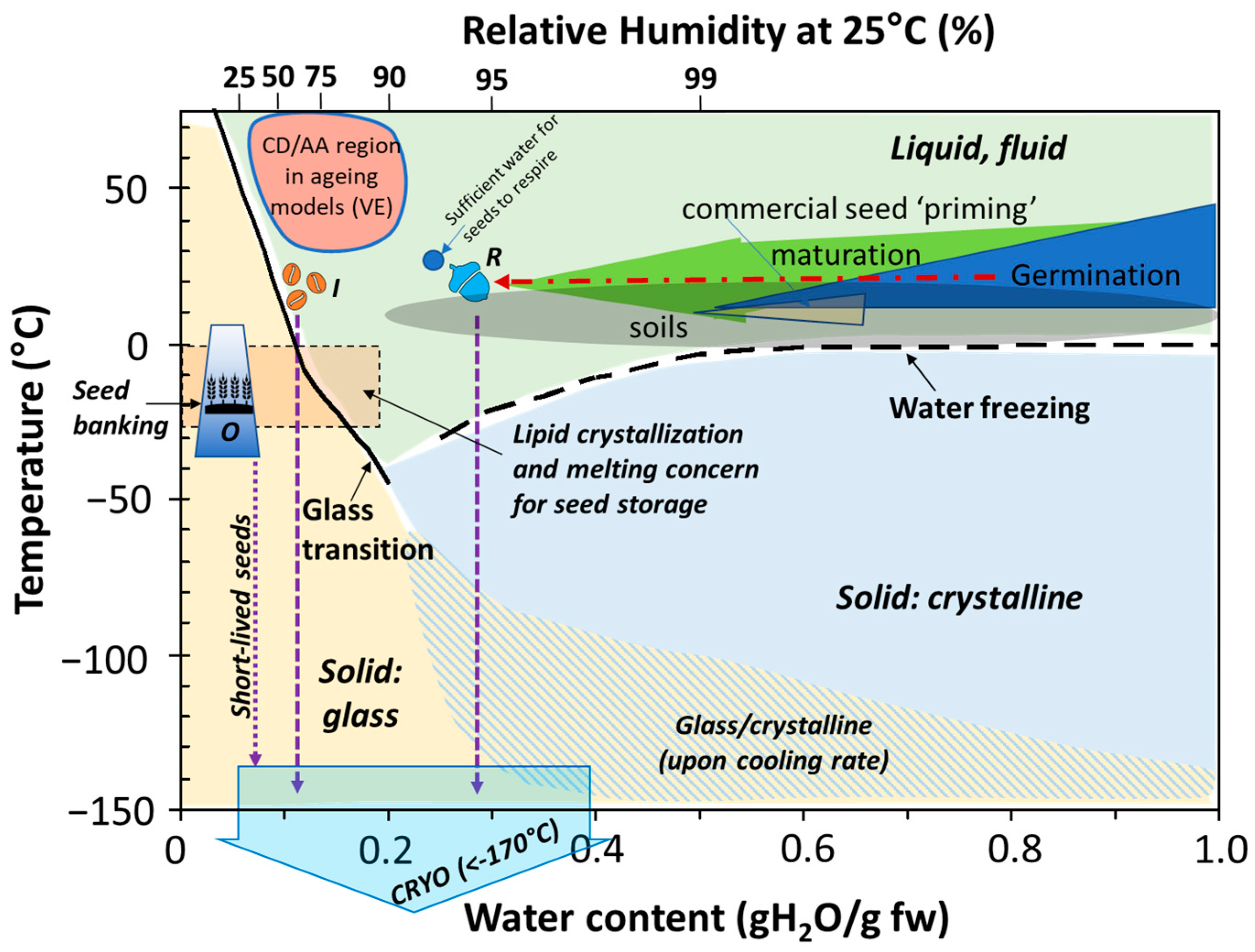{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
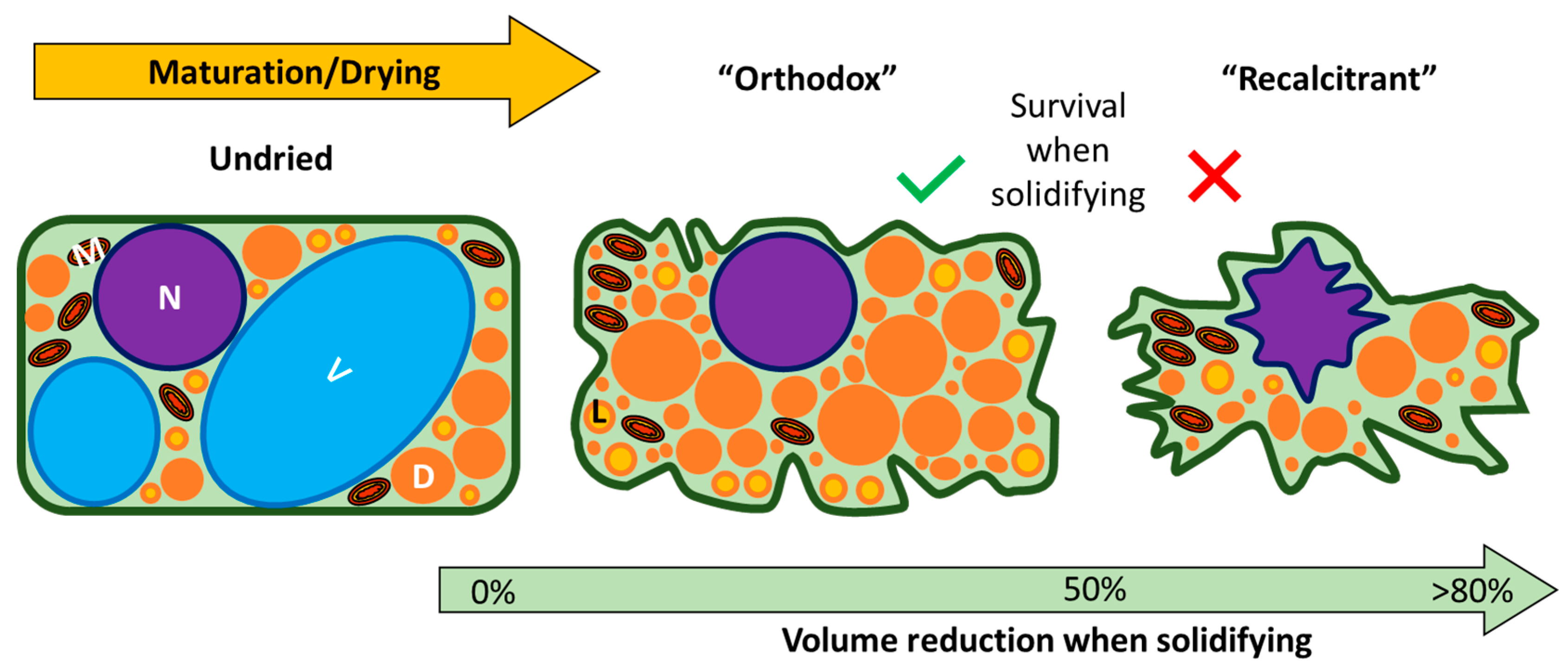
2. Seed Storage Classification
3. Prediction of Seed Storage Behaviour
4. Relationship among Seed Moisture, RH and Temperature
5. Long-Term Conservation and Molecular Motion
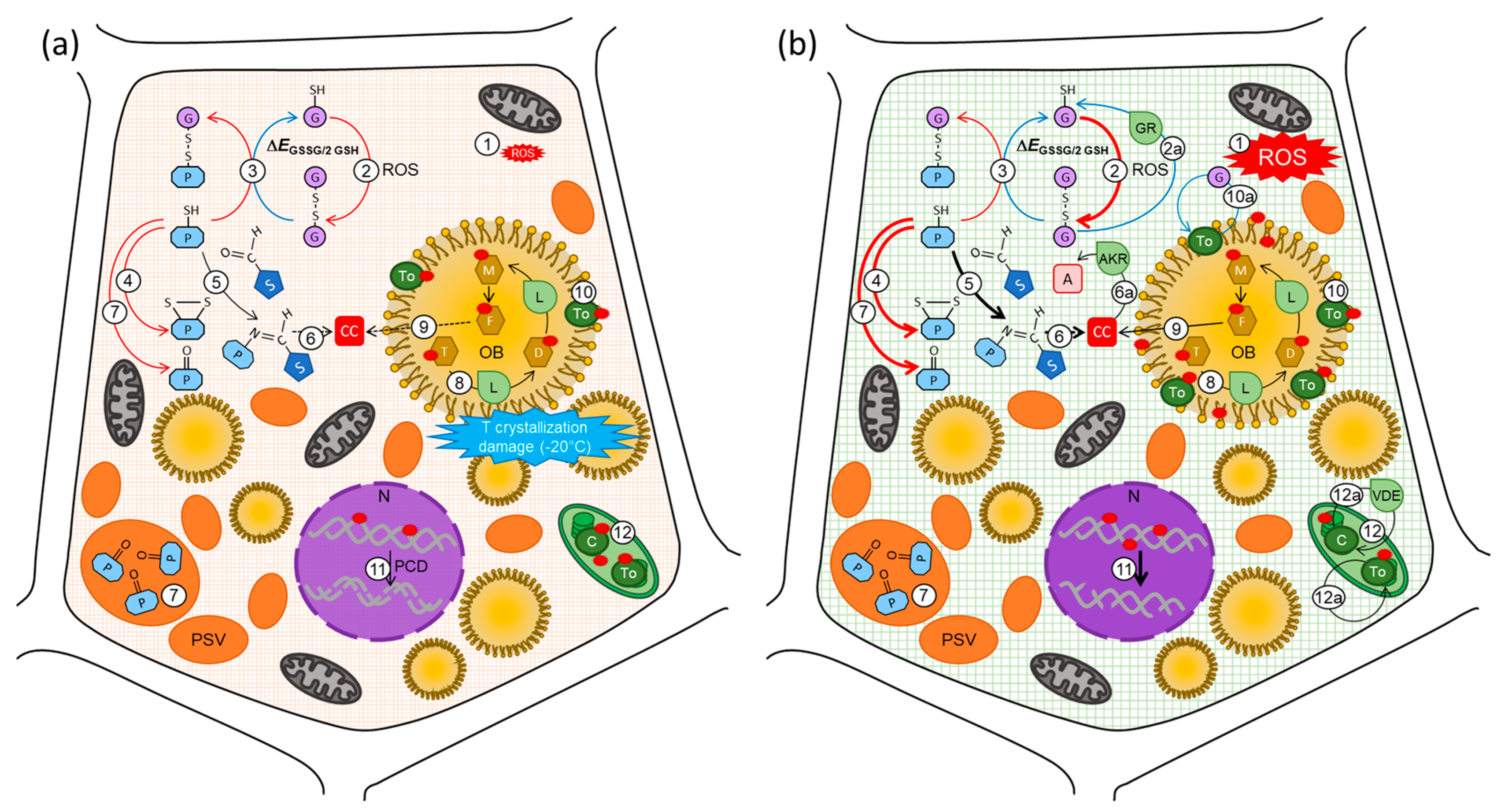
6. Ageing during Storage of Orthodox Seeds
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walters, C.; Pence, V.C. The unique role of seed banking and cryobiotechnologies in plant conservation. Plants People Planet 2021, 3, 83–91. [Google Scholar] [CrossRef]
- Ewart, A.J. On the longevity of seeds. Proc. R. Soc. Vic. 1908, 21, 1–120. [Google Scholar] [CrossRef]
- Harrington, J.F. Thumb rules of drying seed. Crops Soils 1960, 13, 16–17. [Google Scholar]
- Justice, O.L.; Bass, L.N. Principles and Practices of Seed Storage (No. 506); US Department of Agriculture Press: Washington, DC, USA, 1978.
- Roberts, E.H.; Ellis, R.H. Water and seed survival. Ann. Bot. 1989, 63, 39. [Google Scholar] [CrossRef]
- Walters, C. Understanding the mechanisms and kinetics of seed aging. Seed Sci. Res. 1998, 8, 223–244. [Google Scholar] [CrossRef]
- Walters, C.; Ballesteros, D.; Vertucci, V.A. Structural mechanics of seed deterioration: Standing the test of time. Plant Sci. 2010, 179, 565–573. [Google Scholar] [CrossRef]
- Ellis, R.H. Invited Review: Seed ageing, survival and the improved seed viability equation; forty years on. Seed Sci. Tech. 2022, 50 (Suppl. 1), 1–20. [Google Scholar] [CrossRef]
- Harrington, J.F. Seed storage and longevity. In Seed Biology III; Kozlowski, T.T., Ed.; Academic Press: New York, NY, USA, 1972; pp. 145–245. [Google Scholar]
- Liu, U.; Cossu, T.A.; Dickie, J.B. Royal Botanic Gardens, Kew’s Seed Information Database (SID): A compilation of taxon-based biological seed characteristics or traits. Biodivers. Inf. Sci. Stand. 2019, 3, e37030. [Google Scholar] [CrossRef]
- Vertucci, C.W.; Roos, E.E. Theoretical basis of protocols for seed storage. Plant Phys. 1990, 94, 1019–1023. [Google Scholar] [CrossRef]
- Roberts, E.H. Predicting the storage life of seeds. Seed Sci. Tech. 1973, 1, 499–514. [Google Scholar]
- Ellis, R.H.; Hong, T.D.; Roberts, E.H. An intermediate category of seed storage behaviour. II. Effects of provenance, immaturity, and imbibition on desiccation-tolerance in coffee. J. Exp. Bot. 1991, 42, 653–657. [Google Scholar] [CrossRef]
- FAO. Genebank Standards for Plant Genetic Resources for Food and Agriculture, Revised Edition; Food and Agricultural Organization of the United Nations: Rome, Italy, 2014. [Google Scholar]
- Center for Plant Conservation (CPC). CPC Best Plant Conservation Practices to Support Species Survival in the Wild; CPC: Escondido, CA, USA, 2019; Available online: https://saveplants.org/cpc-best-plant-conservation-practices-to-support-species-survival-in-the-wild/ (accessed on 7 December 2022).
- The Millennium Seed Bank Partnership (MSBP) Seed Conservation Standards for ‘MSBP Collections’. Available online: http://brahmsonline.kew.org/Content/Projects/msbp/resources/Training/MSBP-Seed-Conservation-Standards.pdf (accessed on 7 December 2022).
- Daws, M.I.; Garwood, N.C.; Pritchard, H.W. Prediction of desiccation sensitivity in seeds of woody species: A probabilistic model based on two seed traits and 104 species. Ann. Bot. 2006, 97, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Tweddle, J.C.; Dickie, J.B.; Baskin, C.C.; Baskin, J.M. Ecological aspects of seed desiccation sensitivity. J. Ecol. 2003, 91, 294–304. [Google Scholar] [CrossRef]
- Lan, Q.Y.; Xia, K.; Wang, X.F.; Liu, J.W.; Zhao, J.; Tan, Y.H. Seed storage behaviour of 101 woody species from the tropical rainforest of southern China: A test of the seed-coat ratio–seed mass (SCR–SM) model for determination of desiccation sensitivity. Aus. J. Bot. 2014, 62, 305–311. [Google Scholar] [CrossRef]
- Wyse, S.V.; Dickie, J.B. Predicting the global incidence of seed desiccation sensitivity. J. Ecol. 2017, 105, 1082–1093. [Google Scholar] [CrossRef]
- Pence, V.C.; Meyer, A.; Linsky, J.; Gratzfeld, J.; Pritchard, H.W.; Westwood, M.; Bruns, E.B. Defining exceptional species-A conceptual framework to expand and advance ex situ conservation of plant diversity beyond conventional seed banking. Biol. Conserv. 2022, 266, 109440. [Google Scholar] [CrossRef]
- Ballesteros, D.; Pritchard, H.W.; Walters, C. Dry architecture: Towards the understanding of the variation of longevity in desiccation-tolerant germplasm. Seed Sci. Res. 2020, 30, 142–155. [Google Scholar] [CrossRef]
- Bailly, C.; Benamar, A.; Corbineau, F.; Côme, D. Changes in malondialdehyde content and in superoxide dismutase, catalase and glutathione reductase activities in sunflower seeds as related to deterioration during accelerated aging. Phys. Plant. 1996, 97, 104–110. [Google Scholar] [CrossRef]
- Murthy, U.M.N.; Sun, W.Q. Protein modification by Amadori and Maillard reactions during seed storage: Roles of sugar hydrolysis and lipid peroxidation. J. Exp. Bot. 2000, 51, 1221–1228. [Google Scholar] [CrossRef]
- Kranner, I.; Chen, H.; Pritchard, H.W.; Pearce, S.R.; Birtić, S. Inter-nucleosomal DNA fragmentation and loss of RNA integrity during seed ageing. Plant Growth Regul. 2011, 63, 63–72. [Google Scholar] [CrossRef]
- Chen, H.; Osuna, D.; Colville, L.; Lorenzo, O.; Graeber, K.; Kuester, H.; Leubner-Metzger, G.; Kranner, I. Transcriptome-wide mapping of pea seed ageing reveals a pivotal role for genes related to oxidative stress and programmed cell death. PLoS ONE 2013, 8, e78471. [Google Scholar] [CrossRef] [PubMed]
- Fleming, M.B.; Hill, L.M.; Walters, C. The kinetics of ageing in dry-stored seeds: A comparison of viability loss and RNA degradation in unique legacy seed collections. Ann. Bot. 2019, 123, 1133–1146. [Google Scholar] [CrossRef] [PubMed]
- Kranner, I.; Kastberger, G.; Hartbauer, M.; Pritchard, H.W. Noninvasive diagnosis of seed viability using infrared thermography. Proc. Natl. Acad. Sci. USA 2010, 107, 3912–3917. [Google Scholar] [CrossRef] [PubMed]
- Colville, L.; Bradley, E.L.; Lloyd, A.S.; Pritchard, H.W.; Castle, L.; Kranner, I. Volatile fingerprints of seeds of four species indicate the involvement of alcoholic fermentation, lipid peroxidation, and Maillard reactions in seed deterioration during ageing and desiccation stress. J. Exp. Bot. 2012, 63, 6519–6530. [Google Scholar] [CrossRef] [PubMed]
- Mira, S.; Hill, L.M.; González-Benito, M.E.; Ibáñez, M.A.; Walters, C. Volatile emission in dry seeds as a way to probe chemical reactions during initial asymptomatic deterioration. J. Exp. Bot. 2016, 67, 1783–1793. [Google Scholar] [CrossRef]
- Michalak, M.; Plitta-Michalak, B.P.; Nadarajan, J.; Colville, L. Volatile signature indicates viability of dormant orthodox seeds. Phys. Plant. 2021, 173, 788–804. [Google Scholar] [CrossRef]
- Berjak, P.; Farrant, J.M.; Pammenter, N.W. The basis of recalcitrant seed behaviour. In Recent Advances in the Development and Germination of Seeds; Taylorson, R.B., Ed.; Springer: Boston, MA, USA, 1989; pp. 89–108. [Google Scholar]
- Vertucci, C.W.; Farrant, J.M. Acquisition and loss of desiccation tolerance. In Seed Development and Germination; Kigel, J., Galili, G., Eds.; Marcel Dekker: New York, NY, USA, 1995; pp. 237–271. [Google Scholar]
- Berjak, P.; Pammenter, N.W. From Avicennia to Zizania: Seed recalcitrance in perspective. Ann. Bot. 2008, 101, 213–228. [Google Scholar] [CrossRef]
- Leprince, O.; Pellizzaro, A.; Berriri, S.; Buitink, J. Late seed maturation: Drying without dying. J. Exp. Bot. 2017, 68, 827–841. [Google Scholar] [CrossRef]
- Li, D.Z.; Pritchard, H.W. The science and economics of ex situ plant conservation. Trends Plant Sci. 2009, 14, 614–621. [Google Scholar] [CrossRef]
- Colville, L.; Pritchard, H.W. Seed life span and food security. New Phytol. 2019, 224, 557–562. [Google Scholar] [CrossRef]
- Hay, F.R.; Sershen. New technologies to improve the ex situ conservation of plant genetic resources. In Plant Genetic Resources; Dulloo, M.E., Ed.; Burleigh Dodds Science Publishing: London, UK, 2021; pp. 185–216. [Google Scholar]
- Jouret, M.F. Ecologie de la dormance séminale et de la germination chez diverces espèces du genre Impatiens L. Bull. Soc. R. Bot. Belge 1976, 109, 213–225. [Google Scholar]
- Albrecht, M.A.; McCarthy, B.C. Effects of storage on seed dormancy and survivorship in black cohosh (Actaea racemosa L.) and goldenseal (Hydrastis canadensis L.). Seed Sci. Tech. 2007, 35, 414–422. [Google Scholar] [CrossRef]
- Ma, X.; Bucalo, K.; Determann, R.O.; Cruse-Sanders, J.M.; Pullman, G.S. Somatic embryogenesis, plant regeneration, and cryopreservation for Torreya taxifolia, a highly endangered coniferous species. In Vitro Cell. Dev. Biol. Plant 2012, 48, 324–334. [Google Scholar] [CrossRef]
- Pence, V.C. In Vitro methods and cryopreservation: Tools for endangered exceptional species preservation and restoration. Acta Hortic. 2014, 1039, 73–79. [Google Scholar] [CrossRef]
- Issac, M.; Kuriakose, P.; Leung, S.; Costa, A.B.; Johnson, S.; Bucalo, K.; Stober, J.M.; Determann, R.O.; Rogers, W.L.; Cruse-Sanders, J.M.; et al. Seed Cryopreservation, Germination, and Micropropagation of Eastern Turkeybeard, Xerophyllum asphodeloides (L.) Nutt.: A Threatened Species from the Southeastern United States. Plants 2021, 10, 1462. [Google Scholar] [CrossRef] [PubMed]
- Hong, T.D.; Ellis, R.H.; Linington, S. Compendium of Information on Seed Storage Behaviour; The Royal Botanic Gardens: London, UK, 1998; p. 901. [Google Scholar]
- Pritchard, H.W.; Wood, C.B.; Hodges, S.; Vautier, H.J. 100-seed test for desiccation tolerance and germination: A case study on eight tropical palm species. Seed Sci. Tech. 2004, 32, 393–403. [Google Scholar] [CrossRef]
- Oliver, M.J.; Farrant, J.M.; Hilhorst, H.W.; Mundree, S.; Williams, B.; Bewley, J.D. Desiccation tolerance: Avoiding cellular damage during drying and rehydration. Ann. Rev. Plant Biol. 2020, 71, 435–460. [Google Scholar] [CrossRef] [PubMed]
- Farrant, J.M.; Hilhorst, H.W. What is dry? Exploring metabolism and molecular mobility at extremely low water contents. J. Exp. Bot. 2021, 72, 1507–1510. [Google Scholar] [CrossRef]
- Berjak, P.; Vertucci, C.W.; Pammenter, N.W. Effects of developmental status and dehydration rate on characteristics of water and desiccation-sensitivity in recalcitrant seeds of Camellia sinensis. Seed Sci. Res. 1993, 3, 155–166. [Google Scholar] [CrossRef]
- Walters, C. Orthodoxy, recalcitrance and in-between: Describing variation in seed storage characteristics using threshold responses to water loss. Planta 2015, 242, 397–406. [Google Scholar] [CrossRef]
- Burke, M.J. The glass state and survival of anhydrous biological systems. In Membranes, Metabolism, and Dry Organisms; Leopold, A.C., Ed.; Cornell University Press: New York, NY, USA, 1986; pp. 358–363. [Google Scholar]
- Koster, K.L. Glass formation and desiccation tolerance in seeds. Plant Phys. 1991, 96, 302–304. [Google Scholar] [CrossRef] [PubMed]
- Buitink, J.; Leprince, O. Glass formation in plant anhydrobiotes: Survival in the dry state. Cryobiology 2004, 48, 215–228. [Google Scholar] [CrossRef]
- Sun, W.Q.; Irving, T.C.; Leopold, A.C. The role of sugar, vitrification and membrane phase transition in seed desiccation tolerance. Phys. Plant. 1994, 90, 621–628. [Google Scholar] [CrossRef]
- Buitink, J.; Leprince, O. Intracellular glasses and seed survival in the dry state. Comptes Rendus Biol. 2008, 331, 788–795. [Google Scholar] [CrossRef]
- Farrant, J.M.; Pammenter, N.W.; Berjak, P.; Walters, C. Subcellular organization and metabolic activity during the development of seeds that attain different levels of desiccation tolerance. Seed Sci. Res. 1997, 7, 135–144. [Google Scholar] [CrossRef]
- Walters, C.; Koster, K.L. Structural dynamics and desiccation damage in plant reproductive organs. In Plant Desiccation Tolerance; Jenks, M.A., Wood, A.J., Eds.; Blackwell Publishing: Ames, IA, USA, 2007; pp. 251–280. [Google Scholar]
- Ballesteros, D.; Hill, L.M.; Lynch, R.T.; Pritchard, H.W.; Walters, C. Longevity of preserved germplasm: The temperature dependency of aging reactions in glassy matrices of dried fern spores. Plant Cell Phys. 2019, 60, 376–392. [Google Scholar] [CrossRef]
- Meryman, H.T. Freezing injury and its prevention in living cells. Annu. Rev. Biophys. Bioeng. 1974, 3, 341–363. [Google Scholar] [CrossRef]
- Steponkus, P.L.; Uemura, M.; Webb, M.S. Freeze-induced destabilization of cellular membranes and lipid bilayers. In Permeability and Stability of Lipid Bilayers; Disalvo, E.A., Simon, S.A., Eds.; CRC Press: Boca Raton, FL, USA, 1995; pp. 77–104. [Google Scholar]
- Walters, C. Temperature dependency of molecular mobility in preserved seeds. Biophys. J. 2004, 86, 1253–1258. [Google Scholar] [CrossRef]
- Ellis, R.H.; Roberts, E.H. The quantification of ageing and survival in orthodox seeds. Seed Sci. Tech. 1981, 9, 373–409. [Google Scholar]
- Niedzielski, M.; Walters, C.; Luczak, W.; Hill, L.M.; Wheeler, L.J.; Puchlaski, J. Assessment of variation in seed longevity within rye, wheat and the intergeneric hybrid triticale. Seed Sci. Res. 2009, 19, 213–224. [Google Scholar] [CrossRef]
- Ballesteros, D.; Hill, L.M.; Walters, C. Variation of desiccation tolerance and longevity in fern spores. J. Plant Physiol. 2017, 211, 53–62. [Google Scholar] [CrossRef]
- Ballesteros, D.; Walters, C. Detailed characterization of mechanical properties and molecular mobility within dry seed glasses: Relevance to the physiology of dry biological systems. Plant J. 2011, 68, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Ballesteros, D.; Walters, C. Solid-state biology and seed longevity: A mechanical analysis of glasses in pea and soybean embryonic axes. Front. Plant Sci. 2019, 10, 920. [Google Scholar] [CrossRef] [PubMed]
- Zheng, G.H.; Jing, X.M.; Tao, K.L. Ultradry seed storage cuts cost of gene bank. Nature 1998, 393, 223–224. [Google Scholar] [CrossRef]
- Ellis, R.H.; Hong, T.D.; Roberts, E.H. An intermediate category of seed storage behaviour? I. Coffee. J. Exp. Bot. 1990, 41, 1167–1174. [Google Scholar] [CrossRef]
- Ellis, R.H.; Hong, T.D.; Roberts, E.H. Effect of storage temperature and moisture on the germination of papaya seeds. Seed Sci. Res. 1991, 1, 69–72. [Google Scholar] [CrossRef]
- Chau, M.M.; Chambers, T.; Weisenberger, L.; Keir, M.; Kroessig, T.I.; Wolkis, D.; Kam, R.; Yoshinaga, A.Y. Seed freeze sensitivity and ex situ longevity of 295 species in the native Hawaiian flora. Amer. J. Bot. 2019, 106, 1248–1270. [Google Scholar] [CrossRef]
- Crane, J.; Miller, A.L.; Van Roekel, J.W.; Walters, C. Triacylglycerols determine the unusual storage physiology of Cuphea seed. Planta 2003, 217, 699–708. [Google Scholar] [CrossRef]
- Volk, G.M.; Crane, J.; Caspersen, A.M.; Hill, L.M.; Gardner, C.; Walters, C. Massive cellular disruption occurs during early imbibition of Cuphea seeds containing crystallized triacylglycerols. Planta 2006, 224, 1415–1426. [Google Scholar] [CrossRef]
- Sacandé, M.; Buitink, J.; Hoekstra, F.A. A study of water relations in neem (Azadirachta indica) seed that is characterized by complex storage behaviour. J. Exp. Bot. 2000, 51, 635–643. [Google Scholar] [CrossRef]
- Buitink, J.; Walters, C.; Hoekstra, F.A.; Crane, J. Storage behavior of Typha latifolia pollen at low water contents: Interpretation on the basis of water activity and glass concepts. Phys. Plant. 1998, 103, 145–153. [Google Scholar] [CrossRef]
- Hong, T.D.; Ellis, R.H.; Buitink, J.; Walters, C.; Hoekstra, F.A.; Crane, J. A model of the effect of temperature and moisture on pollen longevity in air-dry storage environments. Ann. Bot. 1999, 83, 167–173. [Google Scholar] [CrossRef]
- Maroder, H.L.; Prego, I.A.; Facciuto, G.R.; Maldonado, S.B. Storage behaviour of Salix alba and Salix matsudana seeds. Ann. Bot. 2000, 86, 1017–1021. [Google Scholar] [CrossRef]
- Ballesteros, D.; Pence, V.C. Survival and death of seeds during liquid nitrogen storage: A case study on seeds with short lifespans. CryoLetters 2017, 38, 278–289. [Google Scholar] [PubMed]
- Davies, R.M.; Hudson, A.R.; Dickie, J.B.; Cook, C.; O’Hara, T.; Trivedi, C. Exploring seed longevity of UK native trees: Implications for ex situ conservation. Seed Sci. Res. 2020, 30, 101–111. [Google Scholar] [CrossRef]
- Walters, C.; Berjak, P.; Pammenter, N.; Kennedy, K.; Raven, P. Preservation of recalcitrant seeds. Science 2013, 339, 915–916. [Google Scholar] [CrossRef]
- Pence, V.C.; Ballesteros, D.; Walters, C.; Reed, B.M.; Philpott, M.; Dixon, K.W.; Pritchard, H.W.; Culley, T.M.; Vanhove, A.C. Cryobiotechnologies: Tools for expanding long-term ex situ conservation to all plant species. Biol. Conserv. 2020, 250, 108736. [Google Scholar] [CrossRef]
- Royal Botanic Gardens Kew (2022) Seed Information Database (SID), Version 7.1. Available online: http://data.kew.org/sid/ (accessed on 12 August 2022).
- Pritchard, H.W.; Daws, M.I.; Fletcher, B.; Gaméné, C.S.; Msanga, H.P.; Omondi, W. Ecological correlates of seed desiccation tolerance in tropical African dryland trees. Am. J. Bot. 2004, 91, 863–870. [Google Scholar] [CrossRef]
- Davies, R.M.; Newton, R.J.; Hay, F.R.; Probert, R.J. 150-seed comparative longevity protocol–a reduced seed number screening method for identifying short-lived seed conservation collections. Seed Sci. Tech. 2016, 44, 569–584. [Google Scholar] [CrossRef]
- Poincaré, H. The Foundations of Science: Science and Hypothesis. The Value of Science and Method. 332. Henri Poincaré Quotes on Fact from—97 Science Quotes—Dictionary of Science Quotations and Scientist Quotes. 1946. Available online: https://todayinsci.com/P/Poincare_Henri/PoincareHenri-Fact-Quotations.htm (accessed on 7 December 2022).
- Haldane, J.B.S. Possible Worlds. Science and Theology as Art-Forms. 227. J.B.S. Haldane Quotes—50 Science Quotes—Dictionary of Science Quotations and Scientist Quotes. 1927. Available online: https://todayinsci.com/H/Haldane_JBS/HaldaneJBS-Quotations.htm (accessed on 7 December 2022).
- Roberts, E.H. An investigation of inter-varietal differences in dormancy and viability of rice seed. Ann. Bot. 1963, 27, 365–369. [Google Scholar] [CrossRef]
- Ellis, R.H.; Roberts, E.H. A revised seed viability nomograph for onion. Seed Res. 1977, 5, 93–103. [Google Scholar]
- Ellis, R.H.; Roberts, E.H. Improved equations for the prediction of seed longevity. Ann. Bot. 1980, 45, 13–30. [Google Scholar] [CrossRef]
- Dickie, J.B.; Ellis, R.H.; Kraak, H.L.; Ryder, K.; Tompsett, P.B. Temperature and seed storage longevity. Ann. Bot. 1990, 65, 197–204. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis, 3rd ed.; Cambridge University Press: Cambridge, UK, 1971. [Google Scholar]
- Probert, R.J.; Daws, M.I.; Hay, F.R. Ecological correlates of ex situ seed longevity: A comparative study on 195 species. Ann. Bot. 2009, 104, 57–69. [Google Scholar] [CrossRef] [PubMed]
- Hay, F.R.; Valdez, R.; Lee, J.S.; Sta. Cruz, P.C. Seed longevity phenotyping: Recommendations on research methodology. J. Exp. Bot. 2019, 70, 425–434. [Google Scholar] [CrossRef]
- Bueso, E.; Muñoz-Bertomeu, J.; Campos, F.; Brunaud, V.; Martínez, L.; Sayas, E.; Ballester, P.; Yenush, L.; Serrano, R. ARABIDOPSIS THALIANA HOMEOBOX25 uncovers a role for gibberellins in seed longevity. Plant Phys. 2014, 164, 999–1010. [Google Scholar] [CrossRef]
- Nagel, M.; Kranner, I.; Neumann, K.; Rolletschek, H.; Seal, C.E.; Colville, L.; Fernández-Marín, B.; Börner, A. Genome-wide association mapping and biochemical markers reveal that seed ageing and longevity are intricately affected by genetic background and developmental and environmental conditions in barley. Plant Cell Env. 2015, 38, 1011–1022. [Google Scholar] [CrossRef]
- Lee, J.-S.; Kwak, J.; Hay, F.R. Genetic markers associated with seed longevity and vitamin E in diverse Aus rice varieties. Seed Sci. Res. 2020, 30, 133–141. [Google Scholar] [CrossRef]
- Hay, F.R.; Whitehouse, K.J.; Ellis, R.H.; Sackville Hamilton, N.R.; Lusty, C.; Ndjiondjop, M.N.; Tia, D.; Wenzl, P.; Santos, L.G.; Yazbek, M.; et al. CGIAR genebank viability data reveal inconsistencies in seed collection management. Glob. Food Sec. 2021, 30, 100557. [Google Scholar] [CrossRef]
- Pritchard, H.W.; Dickie, J.B. Predicting seed longevity: The use and abuse of seed viability equations. In Seed Conservation: Turning Science into Practice; Smith, R.D., Dickie, J.B., Probert, R.J., Pritchard, H.W., Eds.; Royal Botanic Gardens, Kew: London, UK, 2003; pp. 653–722. [Google Scholar]
- Buijs, G.; Willems, L.A.J.; Kodde, J.; Groot, S.P.C.; Bentsink, L. Evaluating the EPPO method for seed longevity analyses in Arabidopsis. Plant Sci. 2020, 301, 110644. [Google Scholar] [CrossRef]
- Roach, T.; Nagel, M.; Börner, A.; Eberle, C.; Kranner, I. Changes in tocochromanols and glutathione reveal differences in the mechanisms of seed ageing under seedbank conditions and controlled deterioration in barley. Environ. Exp. Bot. 2018, 156, 8–15. [Google Scholar] [CrossRef]
- Gerna, D.; Ballesteros, D.; Arc, E.; Stöggl, W.; Seal, C.E.; Marami-Zonouz, N.; Na, C.S.; Kranner, I.; Roach, T. Does oxygen affect ageing mechanisms of Pinus densiflora seeds? A matter of cytoplasmic physical state. J. Exp. Bot. 2022, 73, 2631–2649. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.H.; Hong, T.D.; Roberts, E.H. A comparison of the low-moisture-content limit to the logarithmic relation between seed moisture and longevity in twelve species. Ann. Bot. 1989, 63, 601–611. [Google Scholar] [CrossRef]
- Walters, C.; Wheeler, L.; Stanwood, P.C. Longevity of cryogenically stored seeds. Cryobiology 2004, 48, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Dussert, S.; Chabrillange, N.; Rocquelin, G.; Engelmann, F.; Lopez, M.; Hamon, S. Tolerance of coffee (Coffea spp.) seeds to ultra-low temperature exposure in relation to calorimetric properties of tissue water, lipid composition, and cooling procedure. Phys. Plant. 2001, 112, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Crane, J.; Kovach, D.; Gardner, C.; Walters, C. Triacylglycerol phase and ‘intermediate’ seed storage physiology: A study of Cuphea carthagenensis. Planta 2006, 223, 1081–1089. [Google Scholar] [CrossRef]
- Pritchard, H.W.; Seaton, P.T. Orchid seed storage: Historical perspective, current status, and future prospects for long-term conservation. Selbyana 1993, 14, 89–104. [Google Scholar]
- Mira, S.; Nadarajan, J.; Liu, U.; González-Benito, M.E.; Pritchard, H.W. Lipid thermal fingerprints of long-term stored seeds of Brassicaceae. Plants 2019, 8, 414. [Google Scholar] [CrossRef]
- Pritchard, H.W. Classification of seed storage ‘types’ for ex situ conservation in relation to temperature and moisture. In Ex Situ Plant Conservation. Supporting Species Survival in the Wild; Guerrant, E.O., Havens, K., Maunder, M., Eds.; Island Press: Washington, DC, USA, 2004; pp. 139–161. [Google Scholar]
- Tompsett, P.B. Desiccation studies in relation to the storage of Araucaria seed. Ann. Appl. Biol. 1984, 105, 581–586. [Google Scholar] [CrossRef]
- Tompsett, P.B. The effect of moisture content and temperature on the seed storage life of Araucaria columnaris. Seed Sci. Tech. 1984, 12, 801–816. [Google Scholar]
- Tompsett, P.B.; Kemp, R. Database of Tropical Tree Seed Research (DABATTS). Database Contents, Royal Botanic Gardens Kew: Kew, UK, 1996; 263p. [Google Scholar]
- Bonner, F.T. Storage of seeds: Potential and limitations for germplasm conservation. For. Ecol. Manag. 1990, 35, 35–43. [Google Scholar] [CrossRef]
- Pritchard, H.W.; Moat, J.; Ferraz, J.B.S.; Marks, T.R.; Camargo, J.L.C.; Nadarajan, J.; Ferraz, I.D.K. Innovative approaches to the preservation of tree species. For. Ecol. Manag. 2014, 33, 88–98. [Google Scholar] [CrossRef]
- Pritchard, H.W.; Sershen; Tsan, F.Y.; Wen, B.; Jaganathan, G.K.; Calvi, G.; Pence, V.C.; Mattana, E.; Ferraz, I.D.K.; Seal, C.E. Regeneration in recalcitrant-seeded species and risks from climate change. In Plant Regeneration from Seeds: A Global Warming Perspective; Baskin, C.C., Baskin, J.M., Eds.; Elsevier: New York, NY, USA, 2022; p. 15. [Google Scholar]
- Hong, T.D.; Ellis, R.H. Contrasting seed storage behaviour among different species of Meliaceae. Seed Sci. Tech. 1998, 26, 77–95. [Google Scholar]
- Hamilton, K.N.; Offord, C.; Cuneo, P.; Deseo, M.A. A comparative study of seed morphology in relation to desiccation tolerance and other physiological responses in 71 Eastern Australian rainforest species. Plant Species Biol. 2013, 28, 51–62. [Google Scholar] [CrossRef]
- De Lima Jr, M.; Hong, T.D.; Arruda, Y.M.B.C.; Mendes, A.M.S.; Ellis, R.H. Classification of seed storage behaviour of 67 Amazonian tree species. Seed Sci. Tech. 2014, 42, 363–392. [Google Scholar]
- Mattana, E.; Peguero, B.; Di Sacco, A.; Agramonte, W.; Encarnación Castillo, W.R.; Jiménez, F.; Clase, T.; Pritchard, H.W.; Gómez-Barreiro, P.; Castillo-Lorenzo, E.; et al. Assessing seed desiccation responses of native trees in the Caribbean. New For. 2020, 51, 705–721. [Google Scholar] [CrossRef]
- Wyse, S.V.; Dickie, J.B. Taxonomic affinity, habitat and seed mass strongly predict seed desiccation response: A boosted regression trees analysis based on 17 539 species. Ann. Bot. 2018, 121, 71–83. [Google Scholar] [CrossRef]
- Cromarty, A.S.; Ellis, R.H.; Roberts, E.H. The design of seed storage facilities for genetic conservation. In Handbooks for Genebanks: No 1; Ellis, R.H., Hong, T.D., Roberts, E.H., Eds.; International Board for Plant Genetic Resources: Rome, Italy, 1985; p. 100. [Google Scholar]
- Vertucci, C.W.; Leopold, A.C. Water binding in legume seeds. Plant Phys. 1987, 85, 224–231. [Google Scholar] [CrossRef]
- Ellis, R.H.; Hong, T.D. Temperature sensitivity of the low-moisture-content limit to negative seed longevity–moisture content relationships in hermetic storage. Ann. Bot. 2006, 97, 785–791. [Google Scholar] [CrossRef]
- Walters, C.; Wheeler, L.M.; Grotenhuis, J.M. Longevity of seeds stored in a genebank: Species characteristics. Seed Sci. Res. 2005, 15, 1–20. [Google Scholar] [CrossRef]
- Fernández-Marín, B.; Kranner, I.; Sebastián, M.S.; Artetxe, U.; Laza, J.M.; Vilas, J.L.; Pritchard, H.W.; Nadarajan, J.; Míguez, F.; Becerril, J.M.; et al. Evidence for the absence of enzymatic reactions in the glassy state. A case study of xanthophyll cycle pigments in the desiccation-tolerant moss Syntrichia ruralis. J. Exp. Bot. 2013, 64, 3033–3043. [Google Scholar] [CrossRef] [PubMed]
- Buitink, J.; Leprince, O.; Hemminga, M.A.; Hoekstra, F.A. Molecular mobility in the cytoplasm: An approach to describe and predict lifespan of dry germplasm. Proc. Natl. Acad. Sci. USA 2000, 97, 2385–2390. [Google Scholar] [CrossRef] [PubMed]
- Walters, C.; Hill, L.M.; Wheeler, L.J. Dying while dry: Kinetics and mechanisms of deterioration in desiccated organisms. Integr. Comp. Biol. 2005, 45, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Davies, R.M.; Dickie, J.B.; Ballesteros, D. Evaluation of short-lived seeds’ cryopreservation as alternative to conventional seed banking. Cryobiology 2018, 85, 140–141. [Google Scholar] [CrossRef]
- Yoshioka, S.; Miyazaki, T.; Aso, Y. Beta relaxation of insulin molecule in lyophilized formulations containing trehalose or dextran as a determinant of chemical reactivity. Pharm. Res. 2006, 23, 961–966. [Google Scholar] [CrossRef]
- Chang, L.L.; Pikal, M.J. Mechanisms of protein stabilization in the solid state. J. Pharm. Sci. 2009, 98, 2886–2908. [Google Scholar] [CrossRef]
- Hor, Y.L.; Kim, Y.J.; Ugap, A.; Chabrillange, N.; Sinniah, U.R.; Engelmann, F.; Dussert, S. Optimal hydration status for cryopreservation of intermediate oily seeds: Citrus as a case study. Ann. Bot. 2005, 95, 1153–1161. [Google Scholar] [CrossRef] [PubMed]
- Walters, C.; Landré, P.; Hill, L.; Corbineau, F.; Bailly, C. Organization of lipid reserves in cotyledons of primed and aged sunflower seeds. Planta 2005, 222, 397–407. [Google Scholar] [CrossRef]
- Hamilton, K.N.; Ashmore, S.E.; Pritchard, H.W. Thermal analysis and cryopreservation of seeds of Australian wild Citrus species (Rutaceae): Citrus australasica, C. inodora and C. garrawayi. CryoLetters 2009, 30, 268–279. [Google Scholar]
- Nadarajan, J.; Pritchard, H.W. Biophysical characteristics of successful oilseed embryo cryoprotection and cryopreservation using vacuum infiltration vitrification: An innovation in plant cell preservation. PLoS ONE 2014, 9, e96169. [Google Scholar] [CrossRef]
- Ballesteros, D.; Sershen, S.; Varghese, B.; Berjak, P.; Pammenter, N.W. Uneven drying of zygotic embryos and embryonic axes of recalcitrant seeds: Challenges and considerations for cryopreservation. Cryobiology 2014, 69, 100–109. [Google Scholar] [CrossRef]
- Pammenter, N.W.; Vertucci, C.W.; Berjak, P. Homeohydrous (recalcitrant) seeds: Dehydration, the state of water and viability characteristics in Landolphia kirkii. Plant Physiol. 1991, 96, 1093–1098. [Google Scholar] [CrossRef] [PubMed]
- Sershen; Pammenter, N.W.; Berjak, P.; Wesley-Smith, J. Cryopreservation of embryonic axes of selected amaryllid species. CryoLetters 2007, 28, 387–399. [Google Scholar] [PubMed]
- Wesley-Smith, J.; Berjak, P.; Pammenter, N.W.; Walters, C. Intracellular ice and cell survival in cryo-exposed embryonic axes of recalcitrant seeds of Acer saccharinum: An ultrastructural study of factors affecting cell and ice structures. Ann. Bot. 2014, 113, 695–709. [Google Scholar] [CrossRef] [PubMed]
- Corredoira, E.; San-José, M.C.; Ballester, A.; Vieitez, A.M. Cryopreservation of zygotic embryo axes and somatic embryos of European chestnut. CryoLetters 2004, 25, 33–42. [Google Scholar]
- Ballesteros, D.; Fanega-Sleziak, N.; Davies, R.M. Cryopreservation of seeds and seed embryos in orthodox-, intermediate-, and recalcitrant-seeded species. In Cryopreservation and Freeze-Drying Protocols; Wolkers, W.F., Oldenhof, H., Eds.; Humana: New York, NY, USA, 2021; pp. 663–682. [Google Scholar]
- Berjak, P.; Varghese, B.; Pammenter, N.W. Cathodic amelioration of the adverse effects of oxidative stress accompanying procedures necessary for cryopreservation of embryonic axes of recalcitrant-seeded species. Seed Sci. Res. 2011, 21, 187–203. [Google Scholar] [CrossRef]
- Ballesteros, D.; Naidoo, S.; Varghese, B.; Berjak, P.; Pammenter, N.W. Towards understanding the protection afforded by cryoprotectants commonly used in the cryopreservation of zygotic explants of recalcitrant seeds. Cryobiology 2014, 69, 518. [Google Scholar] [CrossRef]
- Xia, K.; Hill, L.M.; Li, D.Z.; Walters, C. Factors affecting stress tolerance in recalcitrant embryonic axes from seeds of four Quercus (Fagaceae) species native to the USA or China. Ann. Bot. 2014, 114, 1747–1759. [Google Scholar] [CrossRef]
- Volk, G.M.; Walters, C. Plant vitrification solution 2 lowers water content and alters freezing behavior in shoot tips during cryoprotection. Cryobiology 2006, 52, 48–61. [Google Scholar] [CrossRef]
- Benson, E.E. Cryopreservation theory. In Plant Cryopreservation: A Practical Guide; Reed, B., Ed.; Springer: New York, NY, USA, 2008; pp. 15–32. [Google Scholar]
- Elliott, G.D.; Wang, S.; Fuller, B.J. Cryoprotectants: A review of the actions and applications of cryoprotective solutes that modulate cell recovery from ultra-low temperatures. Cryobiology 2017, 76, 74–91. [Google Scholar] [CrossRef]
- Varghese, B.; Pammenter, N.W.; Berjak, P. Cryo-tolerance of zygotic embryos from recalcitrant seeds in relation to oxidative stress—A case study on two amaryllid species. J. Plant Phys. 2012, 169, 999–1011. [Google Scholar]
- Funnekotter, B.; Mancera, R.L.; Bunn, E. Advances in understanding the fundamental aspects required for successful cryopreservation of Australian flora. In Vitro Cell. Dev. Biol. Plant 2017, 53, 289–298. [Google Scholar] [CrossRef]
- Pritchard, H.W. The rise of plant cryobiotechnology and demise of plant cryopreservation? Cryobiology 2018, 85, 160–161. [Google Scholar] [CrossRef]
- Ballesteros, D.; Pritchard, H.W. The cryobiotechnology of oaks: An integration of approaches for the long-term ex situ conservation of Quercus Species. Forests 2020, 11, 1281. [Google Scholar] [CrossRef]
- Abdalla, F.H.; Roberts, E.H. The effects of temperature, moisture, and oxygen on the induction of chromosome damage in seeds of barley, broad beans, and peas during storage. Ann. Bot. 1968, 32, 119–136. [Google Scholar] [CrossRef]
- Abdalla, F.H.; Roberts, E.H. The effects of temperature and moisture on the induction of genetic changes in seeds of barley, broad beans, and peas during storage. Ann. Bot. 1969, 33, 153–167. [Google Scholar] [CrossRef]
- Roberts, E.H.; Abdalla, F.H. The influence of temperature, moisture, and oxygen on period of seed viability in barley, broad beans, and peas. Ann. Bot. 1968, 32, 97–117. [Google Scholar] [CrossRef]
- TeKrony, D.M. Accelerated aging test. J. Seed Tech. 1993, 8, 55. [Google Scholar]
- Floris, C. Ageing in Triticum durum seeds: Behaviour of embryos and endosperms from aged seeds as revealed by the embryo-transplantation technique. J. Exp. Bot. 1970, 21, 462–468. [Google Scholar] [CrossRef]
- Powell, A.A.; Matthews, S. Towards the validation of the controlled deterioration vigour test for small-seeded vegetables. Seed Test. Int. 2005, 129, 21–24. [Google Scholar]
- Murthy, U.M.N.; Kumar, P.P.; Sun, W.Q. Mechanisms of seed ageing under different storage conditions for Vigna radiata (L.) Wilczek: Lipid peroxidation, sugar hydrolysis, Maillard reactions and their relationship to glass state transition. J. Exp. Bot. 2003, 54, 1057–1067. [Google Scholar] [CrossRef]
- Schwember, A.R.; Bradford, K.J. Quantitative trait loci associated with longevity of lettuce seeds under conventional and controlled deterioration storage conditions. J. Exp. Bot. 2010, 61, 4423–4436. [Google Scholar] [CrossRef] [PubMed]
- Kranner, I.; Birtić, S.; Anderson, K.M.; Pritchard, H.W. Glutathione half-cell reduction potential: A universal stress marker and modulator of programmed cell death? Free Rad. Biol. Med. 2006, 40, 2155–2165. [Google Scholar] [CrossRef] [PubMed]
- Seal, C.E.; Zammit, R.; Scott, P.; Flowers, T.J.; Kranner, I. Glutathione half-cell reduction potential and α-tocopherol as viability markers during the prolonged storage of Suaeda maritima seeds. Seed Sci. Res. 2010, 20, 47–53. [Google Scholar] [CrossRef]
- Seal, C.E.; Zammit, R.; Scott, P.; Nyamongo, D.O.; Daws, M.I.; Kranner, I. Glutathione half-cell reduction potential as a seed viability marker of the potential oilseed crop Vernonia galamensis. Ind. Crops Prod. 2010, 32, 687–691. [Google Scholar] [CrossRef]
- Birtic, S.; Colville, L.; Pritchard, H.W.; Pearce, S.R.; Kranner, I. Mathematically combined half-cell reduction potentials of low-molecular-weight thiols as markers of seed ageing. Free Rad. Res. 2011, 45, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Schafer, F.Q.; Buettner, G.R. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Rad. Biol. Med. 2001, 30, 1191–1212. [Google Scholar] [CrossRef]
- Nagel, M.; Seal, C.E.; Colville, L.; Rodenstein, A.; Un, S.; Richter, J.; Pritchard, H.W.; Börner, A.; Kranner, I. Wheat seed ageing viewed through the cellular redox environment and changes in pH. Free Rad. Res. 2019, 53, 641–654. [Google Scholar] [CrossRef]
- Colville, L.; Kranner, I. Desiccation tolerant plants as model systems to study redox regulation of protein thiols. Plant Growth Regul. 2010, 62, 241–255. [Google Scholar] [CrossRef]
- Job, C.; Rajjou, L.; Lovigny, Y.; Belghazi, M.; Job, D. Patterns of protein oxidation in Arabidopsis seeds and during germination. Plant Phys. 2005, 138, 790–802. [Google Scholar] [CrossRef]
- Rajjou, L.; Lovigny, Y.; Groot, S.P.; Belghazi, M.; Job, C.; Job, D. Proteome-wide characterization of seed aging in Arabidopsis: A comparison between artificial and natural aging protocols. Plant Phys. 2008, 148, 620–641. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.P.; Cueff, G.; Hegedus, D.D.; Rajjou, L.; Bentsink, L. A role for seed storage proteins in Arabidopsis seed longevity. J. Exp. Bot. 2015, 66, 6399–6413. [Google Scholar] [CrossRef] [PubMed]
- Pearce, R.S.; Samad, I.A. Change in fatty acid content of polar lipids during ageing of seeds of peanut (Arachis hypogea L.). J. Exp. Bot. 1980, 31, 1283–1980. [Google Scholar] [CrossRef]
- Stewart, R.R.; Bewley, J.D. Lipid peroxidation associated with accelerated aging of soybean axes. Plant Phys. 1980, 65, 245–248. [Google Scholar] [CrossRef]
- Priestley, D.A.; Leopold, A.C. Lipid changes during natural aging of soybean seeds. Phys. Plant. 1983, 59, 467–470. [Google Scholar] [CrossRef]
- Pukacka, S.; Ratajczak, E. Age-related biochemical changes during storage of beech (Fagus sylvatica L.) seeds. Seed Sci. Res. 2007, 17, 45–53. [Google Scholar] [CrossRef]
- Mira, S.; González-Benito, M.E.; Hill, L.M.; Walters, C. Characterization of volatile production during storage of lettuce (Lactuca sativa) seed. J. Exp. Bot. 2010, 61, 3915–3924. [Google Scholar] [CrossRef]
- Wiebach, J.; Nagel, M.; Börner, A.; Altmann, T.; Riewe, D. Age-dependent loss of seed viability is associated with increased lipid oxidation and hydrolysis. Plant Cell Env. 2020, 43, 303–314. [Google Scholar] [CrossRef]
- Waterworth, W.M.; Bray, C.M.; West, C.E. Seeds and the art of genome maintenance. Front. Plant Sci. 2019, 10, 706. [Google Scholar] [CrossRef]
- Waterworth, W.M.; Footitt, S.; Bray, C.M.; Finch-Savage, W.E.; West, C.E. DNA damage checkpoint kinase ATM regulates germination and maintains genome stability in seeds. Proc. Natl. Acad. Sci. USA 2016, 113, 9647–9652. [Google Scholar] [CrossRef]
- Osborne, D.J.; Boubriak, I. Telomeres and their relevance to the life and death of seeds. Crit. Rev. Plant Sci. 2002, 21, 127–141. [Google Scholar] [CrossRef]
- FAO. The Second Report on the State of the World’s Plant Genetic Resources for Food and Agriculture; Food and Agricultural Organization of the United Nations: Rome, Italy, 2010. [Google Scholar]
- Fu, Y.B.; Ahmed, Z.; Diederichsen, A. Towards a better monitoring of seed ageing under ex situ seed conservation. Conserv. Physiol. 2015, 3, cov026. [Google Scholar] [CrossRef] [PubMed]
- Edelstein, M.; Bradford, K.J.; Burger, D.W. Metabolic heat and CO2 production rates during germination of melon (Cucumis melo L.) seeds measured by microcalorimetry. Seed Sci. Res. 2001, 11, 265–272. [Google Scholar]
- Dalziell, E.L.; Tomlinson, S. Reduced metabolic rate indicates declining viability in seed collections: An experimental proof-of-concept. Conserv. Physiol. 2017, 5, cox058. [Google Scholar] [CrossRef]
- Bello, P.; Bradford, K. Single-seed oxygen consumption measurements and population-based threshold models link respiration and germination rates under diverse conditions. Seed Sci. Res. 2016, 26, 199–221. [Google Scholar] [CrossRef]
- Powell, A.A. A Review of the Principles and Use of the Q2 Seed Analyser; International Seed Testing Association (ISTA): Bassersdof, Switzerland, 2017. Available online: https://www.seedtest.org/api/rm/9DD4DNNYFVH4ZV6/areviewoftheprinciplesanduseoftheq2seedanalyser1.pdf (accessed on 7 December 2022).
- Xin, X.; Wan, Y.; Wang, W.; Yin, G.; McLamore, E.S.; Lu, X. A real-time, non-invasive, micro-optrode technique for detecting seed viability by using oxygen influx. Sci. Rep. 2013, 3, 1–6. [Google Scholar] [CrossRef]
- Ambrose, A.; Lohumi, S.; Lee, W.H.; Cho, B.K. Comparative nondestructive measurement of corn seed viability using Fourier transform near-infrared (FT-NIR) and Raman spectroscopy. Sens. Actuators B Chem. 2016, 224, 500–506. [Google Scholar] [CrossRef]
- Kusumaningrum, D.; Lee, H.; Lohumi, S.; Mo, C.; Kim, M.S.; Cho, B.K. Non-destructive technique for determining the viability of soybean (Glycine max) seeds using FT-NIR spectroscopy. J. Sci. Food Agr. 2018, 98, 1734–1742. [Google Scholar] [CrossRef]
- Qiu, G.; Lü, E.; Lu, H.; Xu, S.; Zeng, F.; Shui, Q. Single-kernel FT-NIR spectroscopy for detecting supersweet corn (Zea mays L. saccharata sturt) seed viability with multivariate data analysis. Sensors 2018, 18, 1010. [Google Scholar] [CrossRef]
- Ma, T.; Tsuchikawa, S.; Inagaki, T. Rapid and non-destructive seed viability prediction using near-infrared hyperspectral imaging coupled with a deep learning approach. Comput. Electron. Agric. 2020, 177, 105683. [Google Scholar] [CrossRef]
- Zhang, M.; Yajima, H.; Umezawa, Y.; Nakagawa, Y.; Esashi, Y. GC-MS identification of volatile compounds evolved by dry seeds in relation to storage conditions. Seed Sci. Tech. 1995, 23, 59–68. [Google Scholar]
- Taylor, A.G.; Lee, P.C.; Zhang, M. Volatile compounds as indicators of seed quality and their influence on seed aging. Seed Tech. 1999, 21, 57–65. [Google Scholar]
- Han, B.; Fernández, V.; Pritchard, H.W.; Colville, L. Gaseous environment modulates volatile emission and viability loss during seed artificial ageing. Planta 2021, 253, 1–16. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nadarajan, J.; Walters, C.; Pritchard, H.W.; Ballesteros, D.; Colville, L. Seed Longevity—The Evolution of Knowledge and a Conceptual Framework. Plants 2023, 12, 471. https://doi.org/10.3390/plants12030471
Nadarajan J, Walters C, Pritchard HW, Ballesteros D, Colville L. Seed Longevity—The Evolution of Knowledge and a Conceptual Framework. Plants. 2023; 12(3):471. https://doi.org/10.3390/plants12030471
Chicago/Turabian StyleNadarajan, Jayanthi, Christina Walters, Hugh W. Pritchard, Daniel Ballesteros, and Louise Colville. 2023. "Seed Longevity—The Evolution of Knowledge and a Conceptual Framework" Plants 12, no. 3: 471. https://doi.org/10.3390/plants12030471
APA StyleNadarajan, J., Walters, C., Pritchard, H. W., Ballesteros, D., & Colville, L. (2023). Seed Longevity—The Evolution of Knowledge and a Conceptual Framework. Plants, 12(3), 471. https://doi.org/10.3390/plants12030471