

Potentiality of Sustainable Maize Production under Rainfed Conditions in the Tropics by Triggering Agro-Physio-Biochemical Traits Ascertained from a Greenhouse

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Results of the Greenhouse Experiment
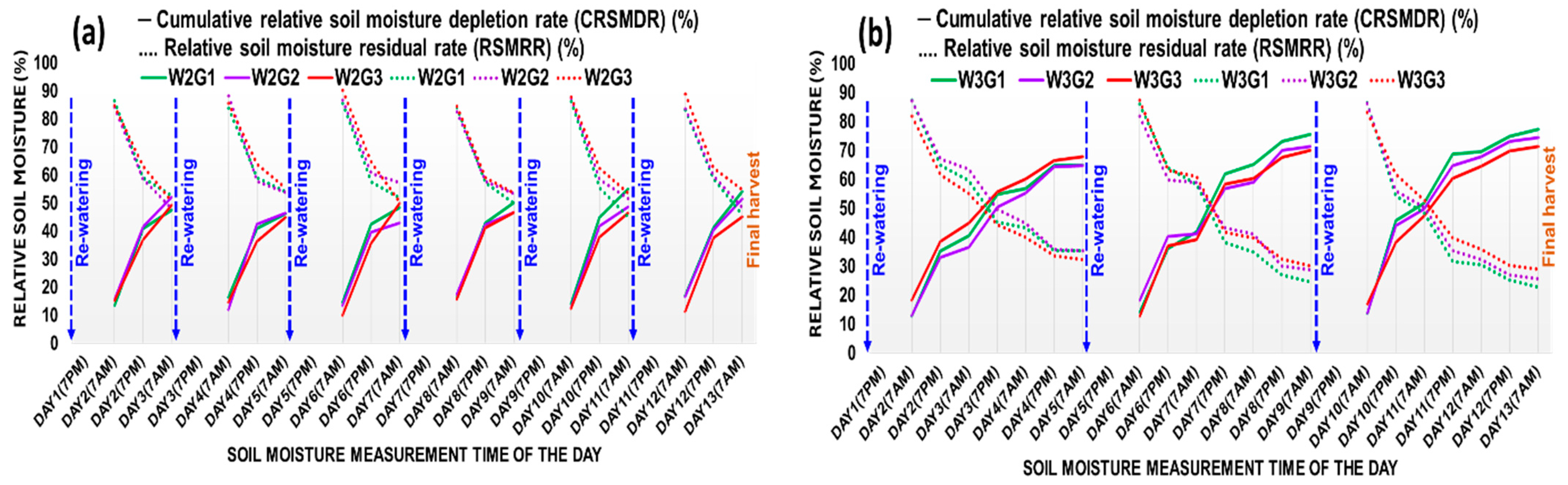
2.1.1. Relative Soil Moisture Dynamics in GH Experiment
2.1.2. Plant Growth Dynamics in GH Experiment
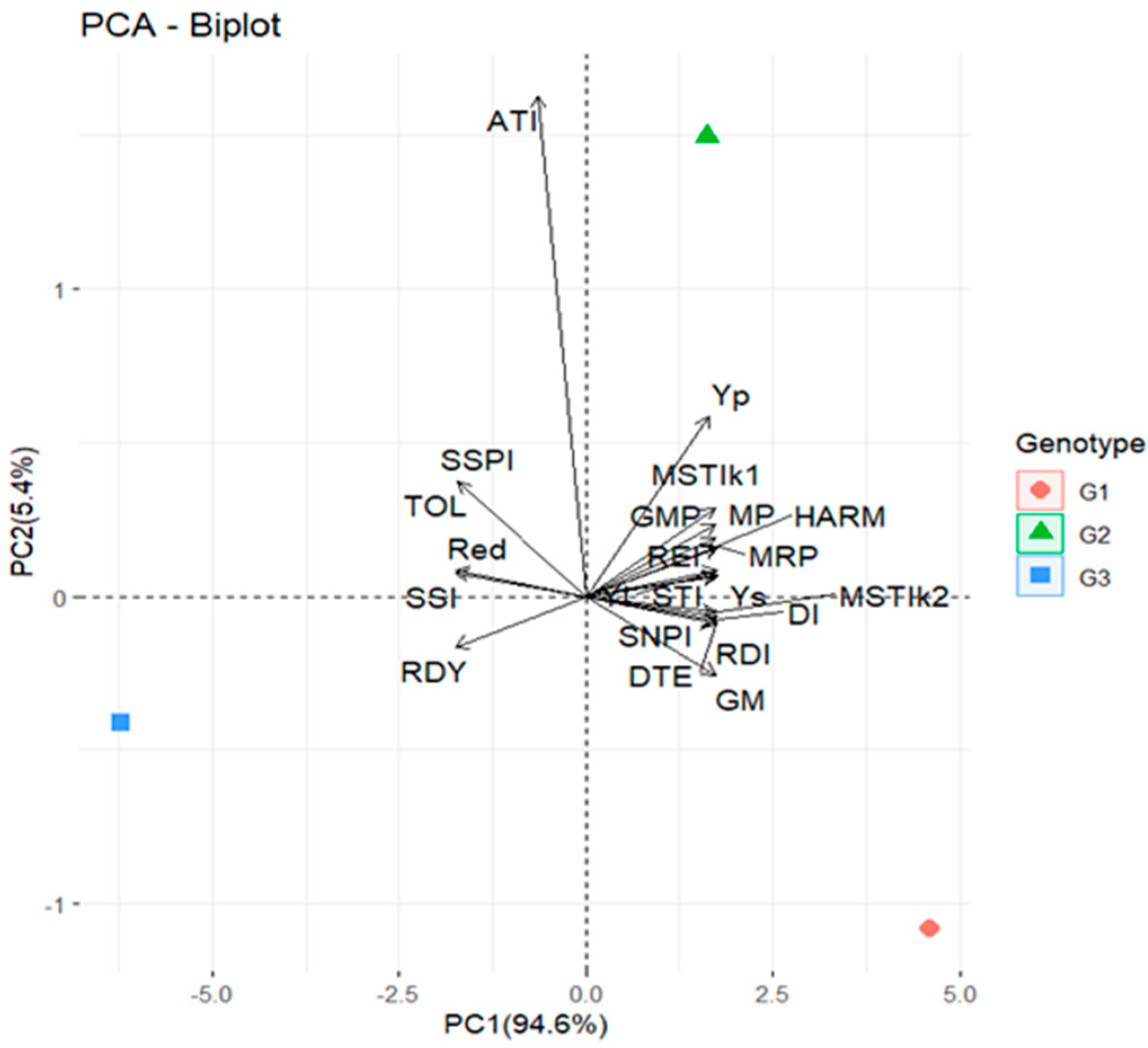
2.1.3. Principal Component Analysis (PCA) and Correlation Coefficients of Drought Tolerance Indices in GH
2.1.4. Selection of Traits Based on Yield Index (YI) in GH
2.2. Results of the Field Experiment
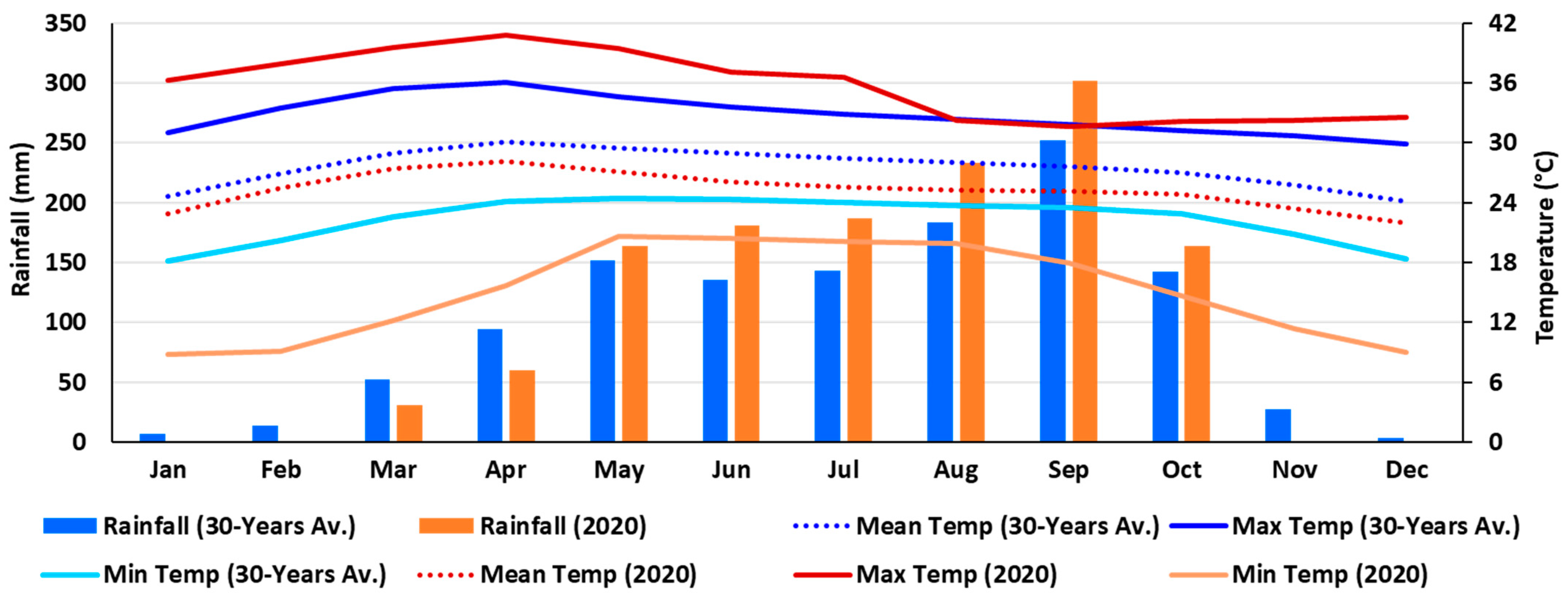
2.2.1. Environmental Conditions of the Field Experimental Location
2.2.2. Plant Growth Dynamics in the Field Experiment
2.2.3. Other Agronomic Traits of Maize as Influenced by Ethephon Application Tactics
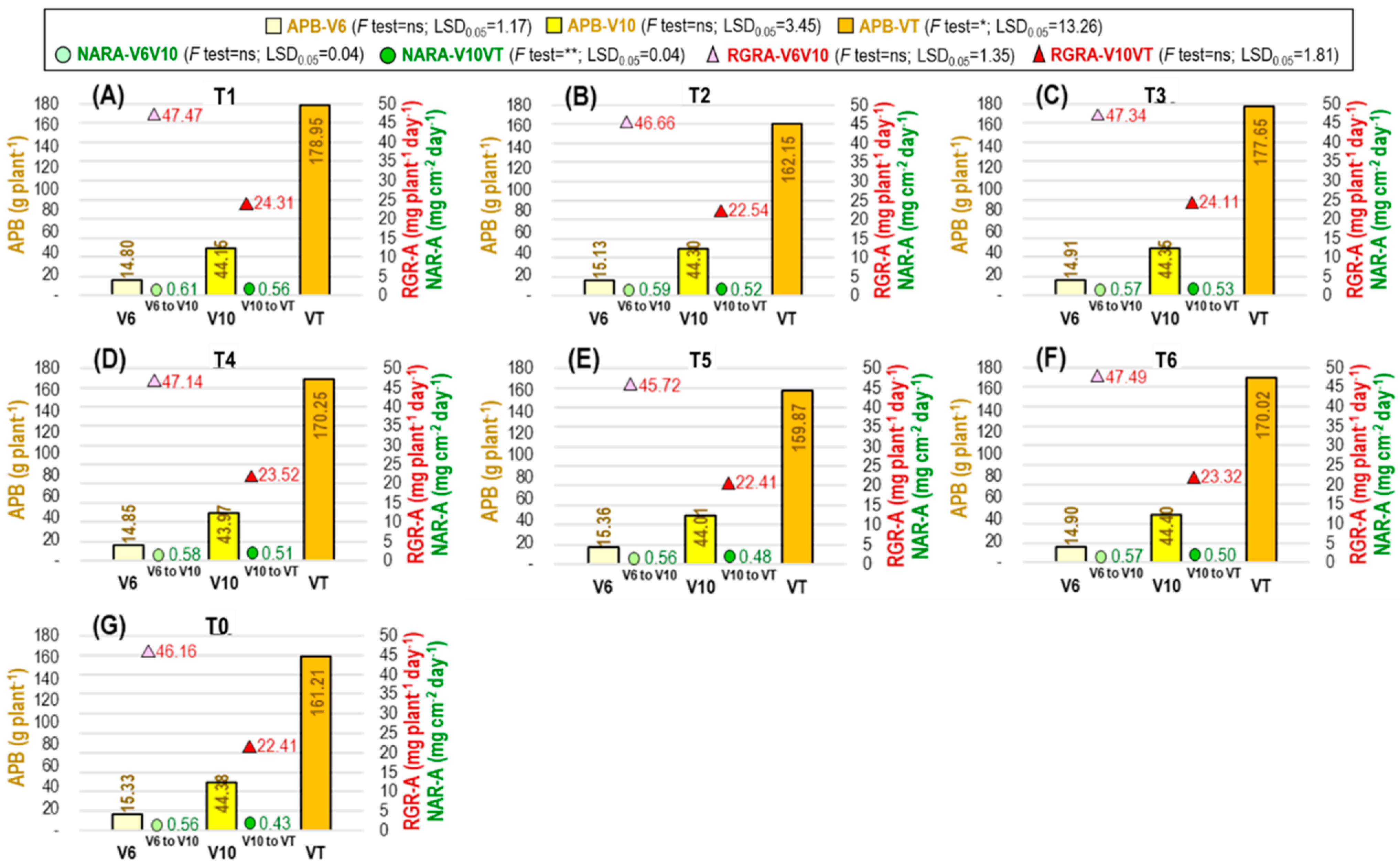
2.2.4. Agro-Physiological Traits of Maize as Influenced by Ethephon Application Tactics
2.2.5. Physio-Biochemical Traits of Maize as Influenced by Ethephon Application Tactics
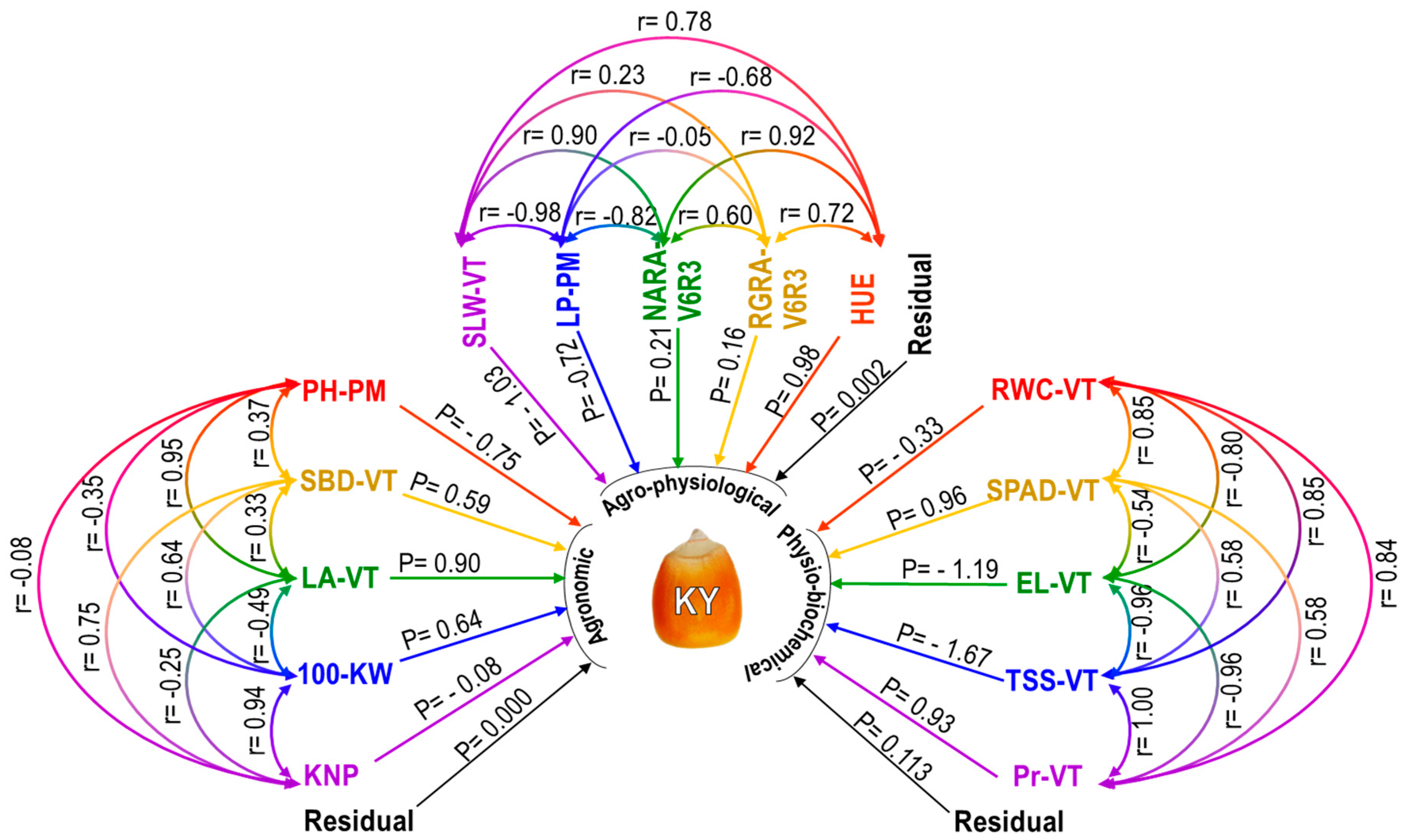
2.2.6. Analysis of Correlation and Path Coefficient Separately for Agronomic, Agro-Physiological, and Physio-Biochemical Traits
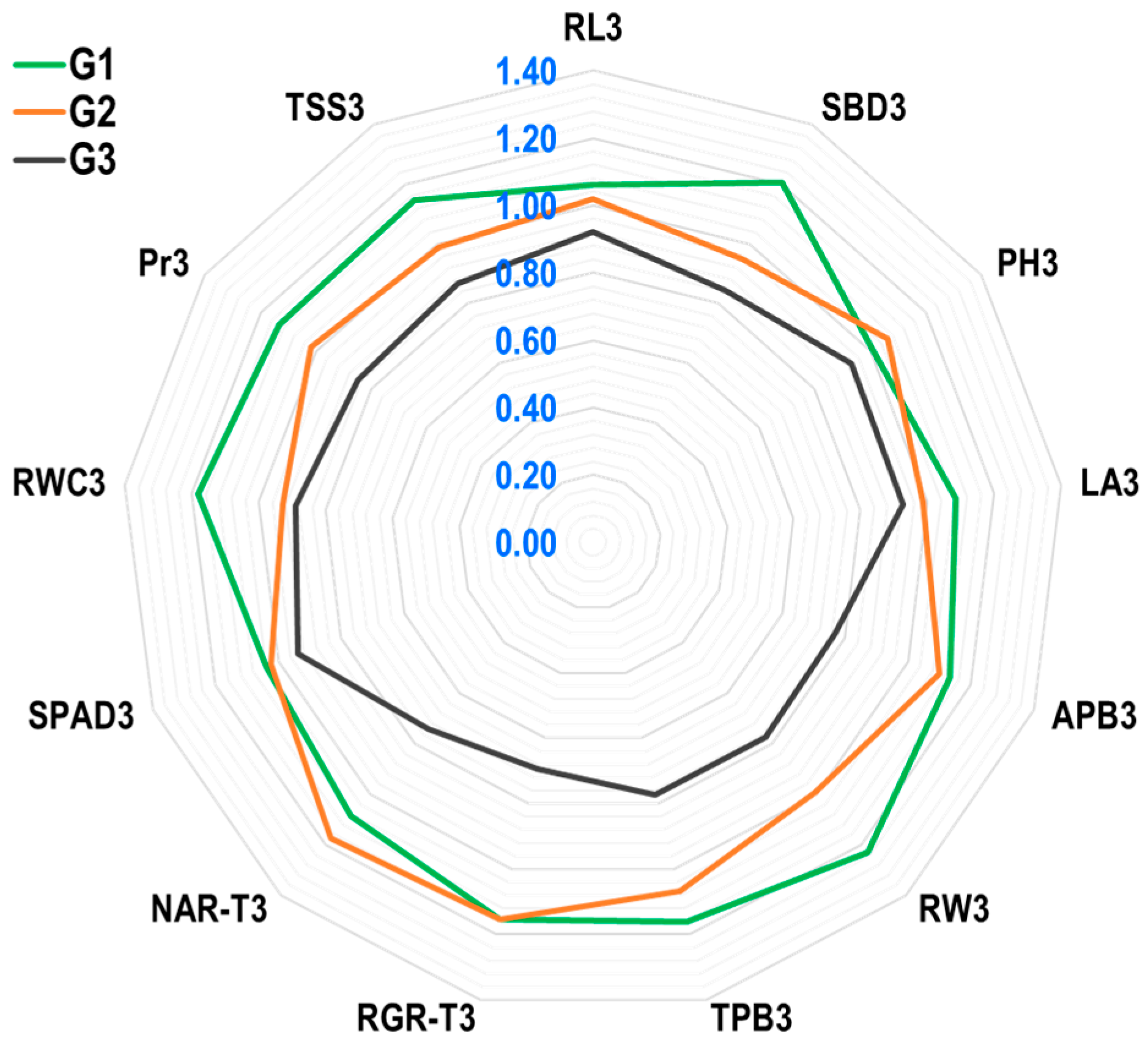
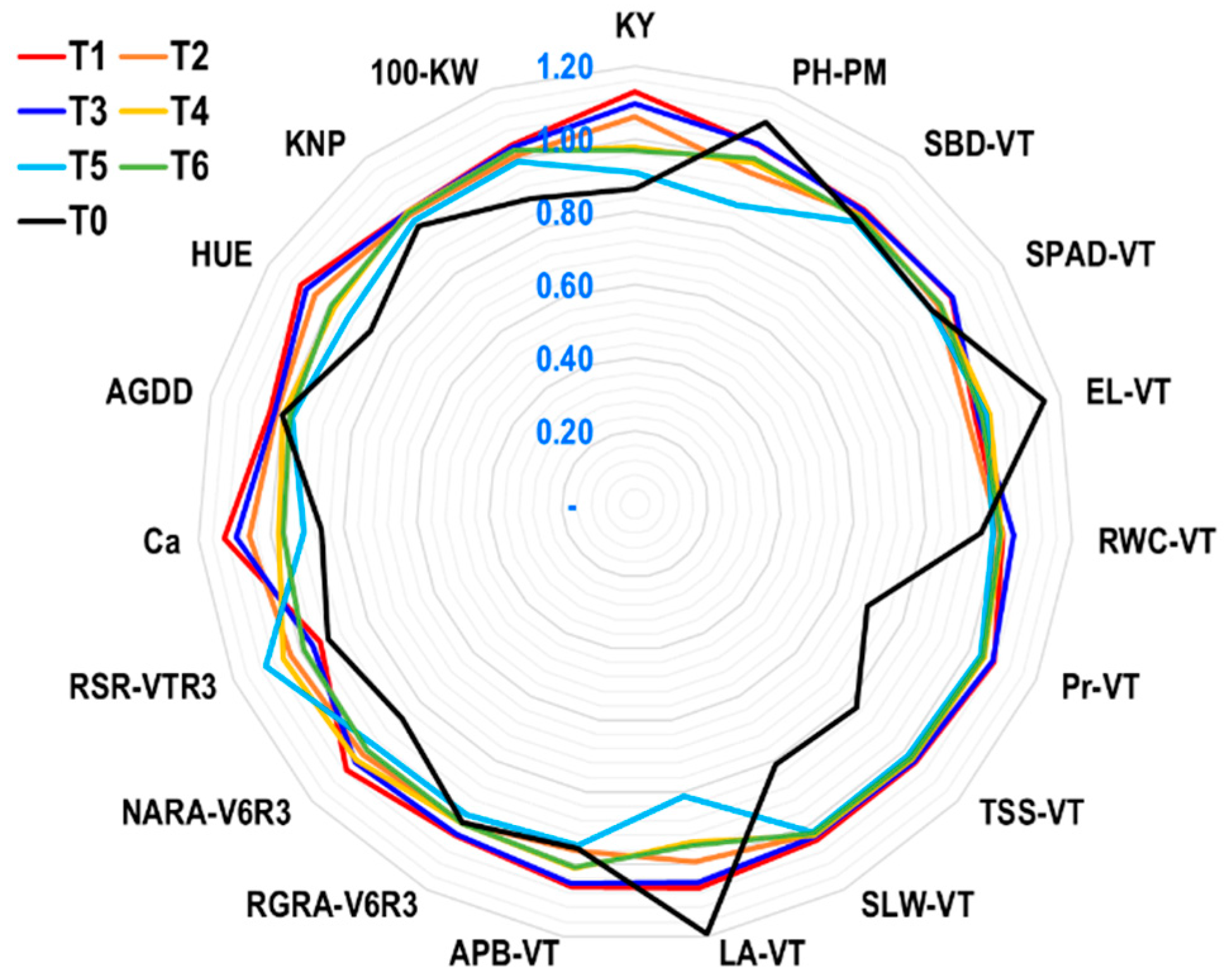
2.2.7. Yield Index (YI) of Agro-Physio-Biochemical Traits of Maize at Field Level
2.2.8. Sustainability
Accumulated Growing Degree Days (AGDDs) and Heat Use Efficiency (HUE)
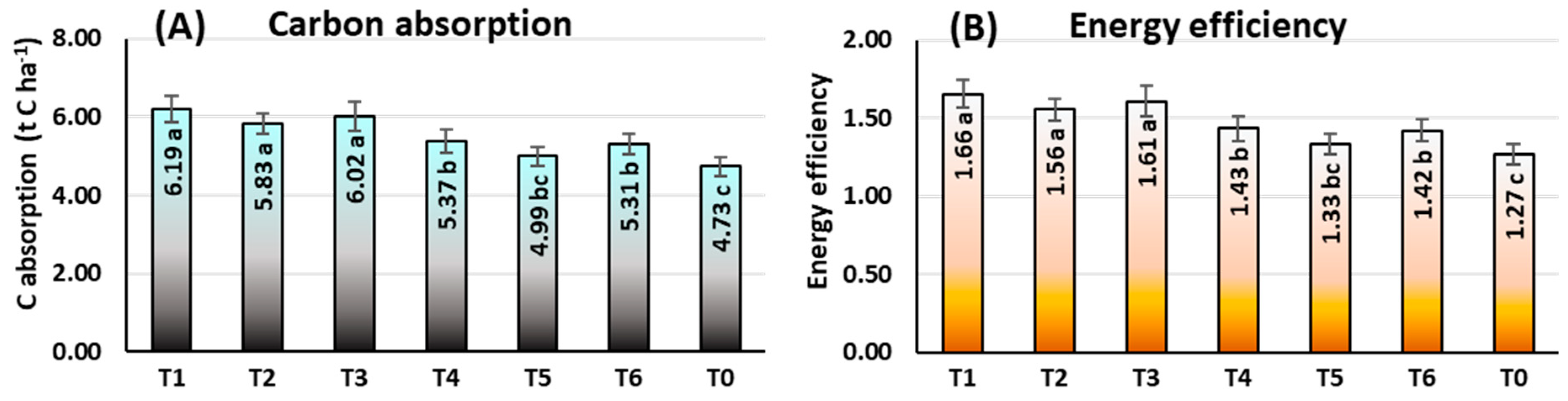
Carbon Absorption (Ca)
Energy Efficiency
Profitability: Gross Return (GR), Gross Margin (GM), and Marginal Benefit–Cost Ratio (MBCR)
3. Discussion
3.1. Climate and Weather Patterns, and Affected Crop Stages in the Field
3.2. Agro-Physio-Biochemical Performance of Maize
3.3. Agronomic Performance of Maize under Greenhouse and Field Experiments
3.4. Agro-Physiological Performance of Maize under Greenhouse and Field Experiments
3.5. Physio-Biochemical Performance of Maize under Greenhouse and Field Experiments
3.6. Sustainability of Maize Production Using Ethephon in Rainfed Conditions
4. Materials and Methods
4.1. Design with Treatments, Plant Materials, Cultural Management, Soil, Climate, and Weather
4.1.1. Greenhouse Experiment
4.1.2. Field Experiment
4.2. Measurements of Different Traits and Plant Sampling in Greenhouse and Field Experiments
4.3. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharma, B.R.; Rao, K.V.; Vittal, K.P.R.; Ramakrishna, Y.S.; Amarasinghe, U. Estimating the potential of rainfed agriculture in India: Prospects for water productivity improvements. Agric. Water Manag. 2010, 97, 23–30. [Google Scholar] [CrossRef]
- USDA-FAS. World Agricultural Outlook. Available online: https://www.fas.usda.gov/data/world-agricultural-production (accessed on 14 November 2022).
- Thakur, N.S.; Singh, M.K.; Bhayal, L.; Meena, K.; Choudhary, S.K.; Kumawat, N.; Singh, R.K.; Singh, U.P.; Singh, S.K.; Sanodiya, P. Sustainability in rainfed maize (Zea mays L.) production using choice of corn variety and nitrogen scheduling. Sustainability 2022, 14, 3116. [Google Scholar] [CrossRef]
- Phuphisith, S.; Gheewala, S.H.; Sampattagul, S. Assessing environmentally sustainable practices of smallholder highland farmers: A case study of maize production in Northern Thailand. Clean Technol. Environ. Policy 2022, 24, 1159–1172. [Google Scholar] [CrossRef]
- Soni, P.; Taewichit, C.; Salokhe, V.M. Energy consumption and CO2 emissions in rainfed agricultural production systems of Northeast Thailand. Agric. Syst. 2013, 116, 25–36. [Google Scholar] [CrossRef]
- Dwivedi, J.L.; Joshi, R.; Arya, V.; Babu, V.R. Rainfed agriculture: Giant strides of current farming practices. In Sustainable Agriculture for Food Security; Apple Academic Press: Point Pleasant, NJ, USA, 2021; pp. 47–74. [Google Scholar]
- Anapalli, S.; Fisher, D.; Reddy, K.; Pettigrew, W.; Sui, R.; Ahuja, L. Vulnerabilities and adapting irrigated and rainfed cotton to climate change in the lower Mississippi delta region. Climate 2016, 4, 55. [Google Scholar] [CrossRef]
- Scott, M. National Climate Assessment: Great Plains’ Ogallala Aquifer Drying Out; National Oceanic and Atmospheric Administration: Washington, DC, USA, 2019; Volume 19.
- Quevauviller, P.; Gemmer, M. EU and international policies for hydrometeorological risks: Operational aspects and link to climate action. Adv. Clim. Change Res. 2015, 6, 74–79. [Google Scholar] [CrossRef]
- Chen, J.; Brissette, F.P.; Lucas-Picher, P.; Caya, D. Impacts of weighting climate models for hydro-meteorological climate change studies. J. Hydrol. 2017, 549, 534–546. [Google Scholar] [CrossRef]
- Khadka, D.; Babel, M.S.; Collins, M.; Shrestha, S.; Virdis, S.G.; Chen, A.S. Projected changes in the near future mean climate and extreme climate events in northeast Thailand. Int. J. Climatol. 2022, 42, 2470–2492. [Google Scholar] [CrossRef]
- Hui, P.; Tang, J.; Wang, S.; Niu, X.; Zong, P.; Dong, X. Climate change projections over China using regional climate models forced by two CMIP5 global models. Part II: Projections of future climate. Int. J. Climatol. 2018, 38, e78–e94. [Google Scholar] [CrossRef]
- Adeyeri, O.E.; Lawin, A.E.; Laux, P.; Ishola, K.A.; Ige, S.O. Analysis of climate extreme indices over the Komadugu-Yobe basin, Lake Chad region: Past and future occurrences. Weather. Clim. Extrem. 2019, 23, 100194. [Google Scholar] [CrossRef]
- Ohba, M.; Sugimoto, S. Differences in climate change impacts between weather patterns: Possible effects on spatial heterogeneous changes in future extreme rainfall. Clim. Dyn. 2019, 52, 4177–4191. [Google Scholar] [CrossRef]
- Di Luca, A.; de Elía, R.; Bador, M.; Argüeso, D. Contribution of mean climate to hot temperature extremes for present and future climates. Weather. Clim. Extrem. 2020, 28, 100255. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Min, S.-K.; Zhang, X.; Sillmann, J.; Sandstad, M. Evaluation of the CMIP6 multi-model ensemble for climate extreme indices. Weather. Clim. Extrem. 2020, 29, 100269. [Google Scholar] [CrossRef]
- Bates, B.C.; Kundzewicz, Z.W.; Wu, S.; Palutikof, J.P. (Eds.) Climate Change and Water; Technical Paper of the Intergovernmental Panel on Climate Change; IPCC Secretariat: Geneva, Switzerland, 2008; 210p. [Google Scholar]
- Singh, D.; Tsiang, M.; Rajaratnam, B.; Diffenbaugh, N.S. Observed changes in extreme wet and dry spells during the South Asian summer monsoon season. Nat. Clim. Change 2014, 4, 456–461. [Google Scholar] [CrossRef]
- Limsakul, A.; Limjirakan, S.; Suthamanuswong, B. Spatio-temporal changes in total annual rainfall and the annual number of rainy days in Thailand. J. Environ. Res. 2007, 29, 1–21. [Google Scholar]
- Le, P.V.; Randerson, J.T.; Willett, R.; Wright, S.; Smyth, P.; Guilloteau, C.; Foufoula-Georgiou, E. Climate-driven changes in the predictability of seasonal precipitation. Nat. Commun. 2023, 14, 3822. [Google Scholar] [CrossRef]
- Pinnamaneni, S.R.; Lima, I.M.; Boone, S.A.; Beacorn, J.A.; Bellaloui, N. Effects of pelleted sugarcane biochar applications on growth, yield and economics of rainfed corn (Zea mays L.) in the Mississippi Delta. Ind. Crops Prod. 2023, 204, 117318. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis; Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013. [Google Scholar]
- IPCC. Global Warming of 1.5 °C; An IPCC Special Report on the Impacts of Global Warming of 1.5 °C above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change; IPCC: Geneva, Switzerland, 2018. [Google Scholar]
- Wangwacharakul, V.; Tangtham, N.; Vijchulata, P.; Liangchalernsit, V.; Towprayoon, S.; Jermsawatdipong, P.; Sumdin, M.S. Thailand’s Initial National Communication under the United Nations Framework Convention on Climate Change. 1991–2020. Available online: https://climateknowledgeportal.worldbank.org/country/Thailand/climate-data-historical (accessed on 25 September 2023).
- Amnuaylojaroen, T.; Chanvichit, P.; Janta, R.; Surapipith, V. Projection of Rice and Maize Productions in Northern Thailand under Climate Change Scenario RCP8. 5. Agriculture 2021, 11, 23. [Google Scholar] [CrossRef]
- Qi, Y.; Zhang, Q.; Hu, S.; Wang, R.; Wang, H.; Zhang, K.; Zhao, H.; Ren, S.; Yang, Y.; Zhao, F.; et al. Effects of High Temperature and Drought Stresses on Growth and Yield of Summer Maize during Grain Filling in North China. Agriculture 2022, 12, 1948. [Google Scholar] [CrossRef]
- Hama, B.M.; Mohammed, A.A. Physiological performance of maize (Zea mays L.) under stress conditions of water deficit and high temperature. Appl. Ecol. Environ. Res. 2019, 17, 1261–1278. [Google Scholar] [CrossRef]
- Boyer, J.S.; Westgate, M.E. Grain yield with limited water. J. Exp. Bot. 2004, 55, 2385–2394. [Google Scholar] [CrossRef]
- Britz, S.J.; Prasad, P.V.V.; Moreau, R.A.; Allen Jr, L.H.D.; Kremer, F.; Boote, K.J. Influence of growth temperature on amounts of tocopherols, tocotrienols and gamma-ozyzanol in brown rice. J. Agric. Food. Chem. 2007, 55, 7559–7565. [Google Scholar] [CrossRef]
- Athar, H.R.; Ashraf, M. Photosynthesis under drought stress. In Handbook of Photosynthesis; Pessarakli, M., Ed.; Taylor and Francis: New York, NY, USA, 2005; pp. 793–804. [Google Scholar]
- Unyayar, S.; Yuksel, K.; Unal, V. Proline and ABA levels in two sunflower genotypes subjected to water stress. Bulg. J. Plant Physiol. 2004, 30, 34–47. [Google Scholar]
- Lawlor, D.W.; Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef]
- Khaled, A.A.; Reda, O.I.; Yaser, H.M.; Esmail, S.M.; El Sabagh, A. Anatomical, Biochemical and Physiological Changes in Some Egyptian Wheat Cultivars Inoculated with Puccinia graminis f. sp. tritici. Fresenius Environ. Bull. 2018, 27, 296–305. [Google Scholar]
- Zhang, L.X.; Li, S.X.; Liang, Z.S. Differential plant growth and osmotic effects of two maize (Zea mays L.) cultivars to exogenous glycinebetaine application under drought stress. Plant Growth Regul. 2009, 58, 297–305. [Google Scholar] [CrossRef]
- Sabagh, A.E.; Hossain, A.; Barutçular, C.; Khaled, A.A.; Fahad, S.; Anjorin, F.B.; Konuskan, O. Sustainable maize (Zea mays L.) production under drought stress by understanding its adverse effect, survival mechanism and drought tolerance indices. J. Exp. Biol. Agric. Sci. 2018, 6, 282–295. [Google Scholar] [CrossRef]
- Bu, L.; Zhang, R.; Chang, Y.; Xue, J.; Han, M. Response of photosynthetic characteristics to water stress of maize leaf in seeding. Acta Ecol. Sin. 2010, 30, 1184–1191. [Google Scholar] [CrossRef]
- Chang, J.; Yang, D.; Tan, W.; Yueshang, L.U. Effects of water stress on maize leaf photosynthesis. J. Northeast Agric. Univ. 2008, 39, 1–5. [Google Scholar]
- Duan, B.; Yang, Y.; Lu, Y.; Korpelainen, H.; Berninger, F.; Li, C. Interactions between drought stress, ABA and genotypes in Picea asperata. J. Exp. Bot. 2007, 58, 3025–3036. [Google Scholar] [CrossRef]
- Chen, P.; Haboudane, D.; Tremblay, N.; Wang, J.; Vigneault, P.; Li, B. New spectral indicator assessing the efficiency of crop nitrogen treatment in corn and wheat. Remote Sens. Environ. 2010, 114, 1987–1997. [Google Scholar] [CrossRef]
- Koksal, E.S. Hyperspectral reflectance data processing through cluster and principal component analysis for estimating irrigation and yield related indicators. Agric. Water Manag. 2011, 98, 1317–1328. [Google Scholar] [CrossRef]
- Sah, R.P.; Chakraborty, M.; Prasad, K.; Pandit, M.; Tudu, V.K.; Chakravarty, M.K.; Narayan, S.C.; Rana, M.; Moharana, D. Impact of water deficit stress in maize: Phenology and yield components. Sci. Rep. 2020, 10, 2944. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Wen, R.; Tian, S.; Su, Y.; He, X.; Su, Y.; Cheng, W.; Huang, K.; Zhang, S. Effects of drought stress and re-watering on growth and yield of various maize varieties at tasseling stage. Agric. Sci. Technol. 2017, 18, 1145–1151. [Google Scholar]
- Ge, T.; Sui, F.; Bai, L.; Tong, C.; Sun, N. Effects of water stress on growth, biomass partitioning, and water use efficiency in summer maize (Zea mays L.) throughout the growth cycle. Acta Physiol. Plant. 2012, 34, 1043–1053. [Google Scholar] [CrossRef]
- Cakir, R. Effect of water stress at different development stages on vegetative and reproductive growth of corn. Field Crops Res. 2004, 89, 1–16. [Google Scholar] [CrossRef]
- Kumdee, O.; Molla, M.S.H.; Kanavittaya, K.; Romkaew, J.; Sarobol, E.; Nakasathien, S. Morpho-Physiological and Biochemical Responses of Maize Hybrids under Recurrent Water Stress at Early Vegetative Stage. Agriculture 2023, 13, 1795. [Google Scholar] [CrossRef]
- Cheng, M.; Wang, H.; Fan, J.; Zhang, F.; Wang, X. Effects of soil water deficit at different growth stages on maize growth, yield, and water use efficiency under alternate partial root-zone irrigation. Water 2021, 13, 148. [Google Scholar] [CrossRef]
- Sinay, H.; Karuwal, R.L. Proline and total soluble sugar content at the vegetative phase of six corn cultivars from Kisar Island Maluku, grown under drought stress conditions. Int. J. Adv. Agric. Res. 2014, 2, 77–82. [Google Scholar]
- Lei, L.; Shi, J.; Chen, J.; Zhang, M.; Sun, S.; Xie, S.; Li, X.; Zeng, B.; Peng, L.; Hauck, A. Ribosome profiling reveals dynamic translational landscape in maize seedlings under drought stress. Plant J. 2015, 84, 1206–1218. [Google Scholar] [CrossRef]
- Seki, M.; Umezawa, T.; Urano, K.; Shinozaki, K. Regulatory metabolic networks in drought stress responses. Curr. Opin. Plant Biol. 2007, 10, 296–302. [Google Scholar] [CrossRef]
- Kamali, B.; Jahanbakhshi, F.; Dogaru, D.; Dietrich, J.; Nendel, C.; Aghakouchak, A. Probabilistic modeling of crop-yield loss risk under drought: A spatial showcase for sub-Saharan Africa. Environ. Res. Lett. 2022, 17, 024028. [Google Scholar] [CrossRef]
- Qayyum, A.; Ahmad, S.; Liaqat, S.; Malik, W.; Noor, E.; Saeed, H.M.; Hanif, M. Screening for drought tolerance in maize (Zea mays L.) hybrids at an early seedling stage. Afr. J. Agric. Res. 2012, 7, 3594–3603. [Google Scholar] [CrossRef]
- Meeks, M.; Murray, S.C.; Hague, S.; Hays, D. Measuring Maize Seedling Drought Response in Search of Tolerant Germplasm. Agronomy 2013, 3, 135–147. [Google Scholar] [CrossRef]
- Hendgen, M.; Gunther, S.; Schubert, S.; Lohnertz, O. Ethephon activates the transcription of senescence-associated genes and nitrogen mobilization in grapevine leaves (Vitis vinifera cv. Riesling). Plants 2021, 10, 333. [Google Scholar] [CrossRef]
- Huang, G.M.; Liu, Y.R.; Guo, Y.L.; Peng, C.X.; Tan, W.M.; Zhang, M.C.; Li, Z.; Zhou, Y.; Duan, L. A novel plant growth regulator improves the grain yield of high-density maize crops by reducing stalk lodging and promoting a compact plant type. Field Crops Res. 2021, 260, 107982. [Google Scholar] [CrossRef]
- Shekoofa, A.; Emam, Y. Plant growth regulator (ethepon) alters maize (Zea mays L.) growth: Water use and grain yield under water stress. J. Agron. 2008, 7, 41–48. [Google Scholar] [CrossRef]
- Kasele, I.N.; Nyirenda, F.; Shanahan, J.F.; Nielsen, D.C.; D’Andria, R. Ethephon alters corn growth, water use, and grain yield under drought stress. Agron. J. 1994, 86, 283–288. [Google Scholar] [CrossRef]
- Peltonen-Sainio, P.; Peltonen, J. Breaking uniculm growth habit of spring cereals at high latitudes by crop management. I. Leaf area index and biomass accumulation. J. Agron. Crop Sci. 1997, 178, 79–86. [Google Scholar] [CrossRef]
- D’ Andria, R.; Chiaranda, F.Q.; Lavini, A.; Mori, M. Grain yield and water consumption of ethephon-treated corn under different irrigation regimes. Agron. J. 1997, 89, 104–112. [Google Scholar] [CrossRef]
- Iqbal, N.; Umar, S.; Per, T.S.; Khan, N.A. Ethephon increases photosynthetic-nitrogen use efficiency, proline and antioxidant metabolism to alleviate decrease in photosynthesis under salinity stress in mustard. Plant Signal. Behav. 2017, 12, e1297000. [Google Scholar] [CrossRef]
- Achard, P.; Cheng, H.; De Grauwe, L.; Decat, J.; Schoutteten, H.; Moritz, T.; Van Der Straeten, D.; Peng, J.; Harberd, N.P. Integration of plant responses to environmentally activated phytohormonal signals. Science 2006, 311, 91–94. [Google Scholar] [CrossRef]
- Lee, H.; Lee, B.R.; Islam, M.T.; La, V.H.; Park, S.H.; Bae, D.W.; Kim, T.H. Cultivar variation in hormone- and sugar-response reveals abscisic acid responsive sucrose phloem loading at the early regenerative stage is a significant determinant of seed yield in Brassica napus. Environ. Exp. Bot. 2020, 169, 103917. [Google Scholar] [CrossRef]
- Iqbal, N.; Nazar, R.; Syeed, S.; Masood, A.; Khan, N.A. Exogenously-sourced ethylene increases stomatal conductance, photosynthesis and growth under optimal and deficient nitrogen fertilization in mustard. J. Exp. Bot. 2011, 62, 4955–4963. [Google Scholar] [CrossRef]
- Monyo, E.S.; Ejeta, G.; Rhodes, D. Genotypic variation for glycinebetaine in sorghum and its relationship to agronomic and morphological traits. Media 1992, 37, 283–286. [Google Scholar]
- Mattioli, R.; Falasca, G.; Sabatini, S.; Altamura, M.M.; Costantino, P.; Trovato, M. The proline biosynthetic genes P5CS1 and P5CS2 play overlapping roles in Arabidopsis flower transition but not in embryo development. Physiol. Plant. 2009, 137, 72–85. [Google Scholar] [CrossRef]
- Ali, Q.; Ashraf, M. Exogenously applied glycinebetaine enhances seed and seed oil quality of maize (Zea mays L.) under water deficit conditions. Environ. Exp. Bot. 2011, 71, 249–259. [Google Scholar] [CrossRef]
- Kaya, C.; Sonmez, O.; Aydemir, S.; Dikilitaş, M. Mitigation effects of glycine betaine on oxidative stress and some key growth parameters of maize exposed to salt stress. Turk. J. Agric. For. 2013, 37, 188–194. [Google Scholar] [CrossRef]
- Reddy, K.R.; Henry, W.B.; Seepaul, R.; Lokhande, S.; Gajanayake, B.; Brand, D. Exogenous application of glycinebetaine facilitates maize (Zea mays L.) growth under water deficit conditions. Am. J. Exp. Agric. 2013, 3, 1–13. [Google Scholar] [CrossRef]
- Sun, Q.; Miao, C. Extreme rainfall (R20mm, RX5day) in Yangtze-Huai, China, in June–July 2016: The role of ENSO and anthropogenic climate change. Bull. Am. Meteorol. Soc. 2018, 99, S102–S106. [Google Scholar] [CrossRef]
- Tang, Q.; Ren, J.; Du, X.; Niu, S.; Liu, S.; Wei, D.; Zhang, Y.; Bian, D.; Cui, Y.; Gao, Z. Reduced stem nonstructural carbohydrates caused by plant growth retardant had adverse effects on maize yield under low density. Front. Plant Sci. 2022, 13, 1035254. [Google Scholar] [CrossRef]
- Gaska, J.M.; Oplinger, E.S. Yield, lodging and growth characteristics in sweet corn as influenced by ethephon timing and rate. Agron. J. 1988, 80, 722–726. [Google Scholar] [CrossRef]
- Norberg, O.S.; Mason, S.C.; Lowry, S.R. Ethephon influence on harvestable yield, grain quality, and lodging of corn. Agron. J. 1988, 80, 768–772. [Google Scholar] [CrossRef]
- Sanvicente, P.; Lazarevitch, S.; Blouet, A.; Guckert, A. Morphological and anatomical modifications in winter barley culm after late plant growth regulator treatment. Eur. J. Agron. 1999, 11, 45–51. [Google Scholar] [CrossRef]
- Gong, L.S.; Qu, S.J.; Huang, G.M.; Guo, Y.L.; Zhang, M.C.; Li, Z.H.; Zhou, Y.Y.; Duan, L.S. Improving maize grain yield by formulating plant growth regulator strategies in north China. J. Integr. Agric. 2021, 20, 622–632. [Google Scholar] [CrossRef]
- Molla, M.S.H.; Kumdee, O.; Worathongchai, N.; Khongchiu, P.; Ali, M.A.; Anwar, M.M.; Wongkaew, A.; Nakasathien, S. Efforts to Stimulate Morpho-Physio-Biochemical Traits of Maize for Efficient Production under Drought Stress in Tropics Field. Agronomy 2023, 13, 2673. [Google Scholar] [CrossRef]
- Ghorbani, R.; Mondani, F.; Amirmoradi, S.; Feizi, H.; Khorramdel, S.; Teimouri, M.; Sanjani, S.; Anvarkhah, S.; Aghel, H. A case study of energy use and economical analysis of irrigated and dryland wheat production systems. Appl. Energy 2011, 88, 283–288. [Google Scholar] [CrossRef]
- Bohra, C.P. A Multi-Criteria Decision Making (MCDM) Approach to Agricultural Production Planning with Energy and Environmental Considerations: The Case of a Central Indian Village. Ph.D. Thesis, Asian Institute of Technology, Bangkok, Thailand, 1998. No. AE-98-2; Unpublished. [Google Scholar]
- Lal, R. Carbon emission from farm operations. Environ. Int. 2004, 30, 981–990. [Google Scholar] [CrossRef]
- IPCC. IPCC Guidelines for National Greenhouse Gas Inventories; Eggleston, H.A.S., Biennia, L., Miwa, K., Negara, T., Tanabe, K., Eds.; Prepared by National Greenhouse Gas Inventories Programme; Eggleston Published; IGES: Hayama, Japan, 2006. [Google Scholar]
- Nakagawa, H.; Harada, T.; Ichinose, T.; Takeno, K.; Matsumoto, S.; Kobayashi, M.; Sakai, M. Biomethanol production and CO2 emission reduction from forage grasses, trees, and crop residues. JARQ 2007, 41, 173–180. [Google Scholar] [CrossRef]
- Ozkan, B.; Fert, C.; Karadeniz, F. Energy and cost analysis for greenhouse and open-field grape production. Energy 2007, 32, 1500–1504. [Google Scholar] [CrossRef]
- Van Ittersum, M.K.; Ewert, F.; Heckelei, T.; Wery, J.; Olsson, J.A.; Andersen, E.; Bezlepkina, I.; Brouwer, F.M.; Donatelli, M.; Flichman, G.; et al. Integrated assessment of agricultural systems—A component-based framework for the European Union (SEAMLESS). Agric. Syst. 2008, 96, 150–165. [Google Scholar] [CrossRef]
- Haarhoff, S.J.; Kotzé, T.N.; Swanepoel, P.A. A prospectus for sustainability of rainfed maize production systems in South Africa. Crop Sci. 2020, 60, 14–28. [Google Scholar] [CrossRef]
- Yan, W.; Rajcan, I. Biplot analysis of test sites and trait relations of soybean in Ontario. Crop Sci. 2002, 42, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, T.C. Maize. In Crop Yield Response to Water; FAO Irrigation & Drainage paper No. 66; Steduto, P., Hsiao, T.C., Fereres, E., Raes, D., Eds.; FAO: Rome, Italy, 2012. [Google Scholar]
- Lievens, E. Parameterization and Testing of the FAO Aquacrop Model to Simulate Yield Response to Water in North-Eastern Thailand. Ph.D., Thesis, Ghent University, Ghent, Belgium, 2014.
- Wongpila, W. Drought Tolerance of Tropical Maize (Zea mays L.) as Affected by Pre-Anthesis Drought. Ph.D. Thesis, Kasetsart University, Bangkok, Thailand, 2008. [Google Scholar]
- Molla, M.S.H. Agro-Physiological and Anatomical Characteristics Associated with Drought and Nitrogen Stress in Maize. Ph.D. Thesis, Kasetsart University, Bangkok, Thailand, 2013. [Google Scholar]
- Jain, M.; Kataria, S.; Hirve, M.; Prajapati, R. Water Deficit Stress Effects and Responses in Maize. In Plant Abiotic Stress Tolerance; Hasanuzzaman, M., Hakeem, K., Nahar, K., Alharby, H., Eds.; Springer: Cham, Switzerland, 2019; pp. 129–151. [Google Scholar] [CrossRef]
- Alam, M.R.; Nakasathien, S.; Molla, M.S.H.; Islam, M.A.; Maniruzzaman, M.; Ali, M.A.; Hossain, A. Kernel water relations and kernel filling traits in maize (Zea mays L.) are influenced by water-deficit condition in a tropical environment. Front. Plant Sci. 2021, 12, 717178. [Google Scholar] [CrossRef] [PubMed]
- Bograd, J.H.; Sarobol, E.; Nakasathien, S. Isogenic genetically-modified and non-genetically-modified corn yields and response to drought and N rates. Agric. Nat. Resour. 2021, 55, 1086–1097. [Google Scholar] [CrossRef]
- Fernandez, G.C. Effective selection criteria for assessing plant stress tolerance. In Proceedings of the International Symposium on Adaptation of Vegetables and other Food Crops in Temperature and Water Stress, Shanhua, Taiwan, 13–18 August 1992; pp. 257–270. [Google Scholar]
- Kaya, Y.; Palta, C.; Taner, S. Additive main effects and multiplicative interactions analysis of yield performances in bread wheat genotypes across environments. Turk. J. Agric. For. 2002, 26, 275–279. [Google Scholar]
- Esmaeilpour, A.; Van Labeke, M.C.; Samson, R.; Ghaffaripour, S.; Van Damme, P. Comparison of biomass production-based drought tolerance indices of pistachio (Pistacia vera L.) seedlings in drought stress conditions. Int. J. Agron. Agric. Res. 2015, 7, 36–44. [Google Scholar]
- Golabadi, M.; Arzani, A.; Maibody, M. Assessment of drought tolerance in segregating populations in durum wheat. Afr. J. Agric. Res. 2006, 1, 162–171. [Google Scholar]
- Zare, M. Evaluation of drought tolerance indices for the selection of Iranian barley (Hordeum vulgare L.) cultivars. Afr. J. Biotechnol. 2012, 11, 15975–15981. [Google Scholar] [CrossRef]
- Najaphy, A.; Geravandi, M. Assessment of indices to identify wheat genotypes adapted to irrigated and rain -fed environments. Adv. Environ. Biol. 2011, 5, 3212–3218. [Google Scholar] [CrossRef]
- Abd El-Mohsen, A.A.; Abd El-Shafi, M.A.; Gheith, E.M.S.; Suleiman, H.S. Using different statistical procedures for evaluating drought tolerance indices of bread wheat genotypes. Adv. Agric. Biol. 2015, 4, 19–30. [Google Scholar]
- Yan, W.; Kang, M.S. Biplot Analysis: A Graphical Tool for Breeders, Geneticists and Agronomist; CRC Press: Boca Raton, FL, USA, 2003; 313p. [Google Scholar]
- Duan, W.; He, B.; Nover, D.; Fan, J.; Yang, G.; Chen, W.; Meng, H.; Liu, C. Floods and Associated Socioeconomic Damages in China over the Last Century. Nat. Hazards 2016, 82, 401–413. [Google Scholar] [CrossRef]
- World Bank Group. Climate Change Knowledge Portal, Thailand. Available online: https://climateknowledgeportal.worldbank.org/country/Thailand/climate-data-historical) (accessed on 15 August 2023).
- ESCAP. Ready for the Dry Years: Building Resilience to Drought in South-East Asia; ESCAP: Bangkok, Thailand, 2020; Available online: https://www.unescap.org/sites/default/files/publications/Ready%20for%20the%20Dry%20Years.pdf (accessed on 14 November 2022).
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought–from genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef] [PubMed]
- Jaleel, C.A.; Gopi, R.; Sankar, B.; Gomathinayagam, M.; Panneerselvam, R. Differential responses in water use efficiency in two varieties of Catharanthus roseus under drought stress. Comptes Rendus Biol. 2008, 331, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Hayano-Kanashiro, C.; Calderon-Vazquez, C.; Ibarra-Laclette, E.; Herrera-Estrella, L.; Simpson, J. Analysis of Gene Expression and Physiological Responses in Three Mexican Maize Landraces under Drought Stress and Recovery Irrigation. PLoS ONE 2009, 4, e7531. [Google Scholar] [CrossRef]
- Molla, M.S.H.; Nakasathien, S.; Ali, M.A.; Khan, A.S.M.M.R.; Alam, M.R.; Hossain, A.; Farooq, M.; El Sabagh, A. Influence of nitrogen application on dry biomass allocation and translocation in two maize varieties under short pre-anthesis and prolonged bracketing flowering periods of drought. Arch. Agron. Soil Sci. 2019, 65, 928–944. [Google Scholar] [CrossRef]
- Laskari, M.; Menexes, G.; Kalfas, I.; Gatzolis, I.; Dordas, C. Water stress effects on the morphological, physiological characteristics of maize (Zea mays L.), and on environmental cost. Agronomy 2022, 12, 2386. [Google Scholar] [CrossRef]
- Salifu, M. Effects of water deficit on the growth and yield formation of maize (Zea mays L.). Acta Agrar. Debreceniensis 2017, 72, 143–148. [Google Scholar] [CrossRef]
- Ali, M.A.; Sarwar, A.K.M.G.; Prodhan, A.K.M.A. Effect of water stresses on the growth features of different maize (Zea mays L.) cultivars. Pak. J. Bot. 1999, 31, 455–654. [Google Scholar]
- Molla, M.S.H.; Nakasathien, S.; Sarobol, E.; Vichukit, V. Growth and physiological responses to supra-optimal nitrogen and pre-anthesis drought stress in maize. Agric. Nat. Resour. 2014, 48, 676–688. [Google Scholar]
- Anjum, S.A.; Xie, X.; Wang, L.C.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar]
- Lee, B.R.; Zaman, R.; La, V.H.; Bae, D.W.; Kim, T.H. Ethephon-induced ethylene enhances starch degradation and sucrose transport with an interactive abscisic acid-mediated manner in mature leaves of oilseed rape (Brassica napus L.). Plants 2021, 10, 1670. [Google Scholar] [CrossRef] [PubMed]
- Li, L.L.; Gu, W.R.; Li, C.F.; Li, W.H.; Chen, X.C.; Zhang, L.G.; Wei, S. Dual application of ethephon and DCPTA increases maize yield and stalk strength. Agron J. 2019, 111, 612–627. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Liu, C.; Ye, D.; Ren, D.; Li, Z.; Zhang, M. Ethephon Reduces Maize Nitrogen Uptake but Improves Ni-trogen Utilization in Zea mays L. Front. Plant Sci. 2022, 12, 762736. [Google Scholar] [CrossRef] [PubMed]
- Karimi, A.; Amirnia, R.; Tajbakhsh, M.; Eivaz, A.R.; Karimi, K. Effect of plant growth inducers on morpho-physiological traits of corn (Zea mays L). Life Sci. 2012, 9, 1683–1688. [Google Scholar] [CrossRef]
- Kepczynska, E.; Zielinska, S. Regulation of Medicago sativa L. Somatic Embryos Regeneration by Gibberellins GA3 and Abscisic Acid in Relation to Starch Content and Alpha-Amylase Activity. Plant Growth Regul. 2006, 29, 209–217. [Google Scholar] [CrossRef]
- Chen, Y.L.; Xiao, C.X.; Wu, D.L.; Xia, T.T.; Chen, Q.W.; Chen, F.J. Effects of nitrogen application rate on grain yield and grain nitrogen concentration in two maize hybrids with contrasting nitrogen remobilization efficiency. Eur. J. Agron. 2015, 62, 79–89. [Google Scholar] [CrossRef]
- Huo, Y.; Wang, M.; Wei, Y.; Xia, Z. Overexpression of the Maize psbA Gene Enhances Drought Tolerance Through Regulating Antioxidant System, Photosynthetic Capability, and Stress Defense Gene Expression in Tobacco. Front. Plant Sci. 2016, 6, 1223. [Google Scholar] [CrossRef]
- Alam, M.R.; Nakasathien, S.; Sarobol, E.; Vichukit, V. Responses of physiological traits of maize to water deficit induced at different phenological stages. Agric. Nat. Resour. 2014, 48, 183–196. [Google Scholar]
- Kasele, I.N.; Shanahan, J.F.; Nielsen, D.C. Impact of growth retardants on corn leaf morphology and gas exchange traits. Crop Sci. 1995, 35, 190–194. [Google Scholar] [CrossRef]
- Horvath, E.; Gombos, B.; Szeles, A. Evaluation phenology, yield and quality of maize genotypes in drought stress and non-stress environments. Agron. Res. 2021, 19, 408–422. [Google Scholar] [CrossRef]
- Malo, M.; Ghosh, A. Studies on different agrometeorological indices and thermal use efficiencies of rice in New Alluvial Zone of West Bengal. Bull. Environ. Pharmacol. Life Sci. 2018, 7, 72–78. [Google Scholar]
- Singh, J.; Hadda, M.S. Phenology and thermal indices of maize (Zea mays L.) influenced by subsoil compaction and nitrogen fertilization under semi-arid irrigated conditions. J. Appl. Nat. Sci. 2014, 6, 349–355. [Google Scholar] [CrossRef]
- Misheck, C.; Fanuel, C. Effect of ethephon and planting density on lodged plant percentage and crop yield in maize (Zea mays L.). Afr. J. Plant Sci. 2014, 8, 113–117. [Google Scholar] [CrossRef]
- Ren, J.; Tang, Q.; Niu, S.; Liu, S.; Wei, D.; Zhang, Y.; Gao, Z. High dose of plant growth regulator enhanced lodging resistance without grain yield reduction of maize under high density. Int. J. Plant Prod. 2022, 16, 329–339. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Ye, D.; Wang, W.; Qiu, X.; Duan, L.; Zhang, M. Ethephon improved stalk strength of maize (Zea mays L.) mainly through altering internode morphological traits to modulate mechanical properties under field conditions. Agronomy 2019, 9, 186. [Google Scholar] [CrossRef]
- Campos, H.; Cooper, M.; Habben, J.E.; Edmeades, G.O.; Schussler, J.R. Improving drought tolerance in maize: A view from industry. Field Crops Res. 2004, 90, 19–34. [Google Scholar] [CrossRef]
- Zlatev, Z.S.; Yordanov, I.T. Effects of soil drought on photosynthesis and chlorophyll fluorescence in bean plants. Bulg. J. Plant Physiol. 2004, 30, 3–18. [Google Scholar]
- Li, R.; Guo, P.; Michael, B.; Stefania, G. Evaluation of Chlorophyll Content and Fluorescence Parameters as Indicators of Drought Tolerance in Barley. Agric. Sci. China 2006, 5, 751–757. [Google Scholar] [CrossRef]
- Ekmekçi, Y.; Bohms, A.; Thomson, J.A.; Mundree, S.G. Photochemica and Antioxidant Responses in the Leaves of Xerophyta viscosa Baker and Digitaria sanguinalis L. under Water Deficit. J. Biosci. Z. Naturforschung C 2005, 60, 435–443. [Google Scholar] [CrossRef]
- Mohsenzadeh, S.; Malboobi, M.A.; Razavi, K.; Farrahi-Aschtiani, S. Physiological and molecular responses of Aeluropus lagopoides (Poaceae) to water deficit. Environ. Exp. Bot. 2006, 56, 314–322. [Google Scholar] [CrossRef]
- Lima, A.L.S.; DaMatta, F.M.; Pinheiro, H.A.; Totola, M.R.; Loureiro, M.E. Photochemical responses and oxidative stress in two clones of Coffea canephora under water deficit conditions. Environ. Exp. Bot. 2002, 47, 239–247. [Google Scholar] [CrossRef]
- Nayyar, H.; Gupta, D. Differential sensitivity of C3 and C4 plants to water deficit stress: Association with oxidative stress and antioxidants. Environ. Exp. Bot. 2006, 58, 106–113. [Google Scholar] [CrossRef]
- Molla, M.S.H.; Nakasathien, S.; Sarobol, E.; Vichukit, V. Anthesis and silking dynamics of maize under contrasting nitrogen and water levels. Agric. Nat. Resour. 2014, 48, 837–850. [Google Scholar]
- Reddy, M.S.B.; Ponnamma, D.; Choudhary, R.; Sadasivuni, K.K. A comparative review of natural and synthetic biopolymer composite scaffolds. Polymers 2021, 13, 1105. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, Y.; Xie, Y.; Wang, Y.; Duan, L.; Zhang, M.; Li, Z. Ethephon improved drought tolerance in maize seedlings by modulating cuticular wax biosynthesis and membrane stability. J. Plant Physiol. 2017, 214, 123–133. [Google Scholar] [CrossRef]
- Zhang, J.; Kirkham, M.B. Water status of drought-resistant and drought-sensitive sorghum treated with ethephon. J. Plant Growth Regulat. 1990, 9, 189–194. [Google Scholar] [CrossRef]
- Shen, S.; Li, B.B.; Deng, T.; Xiao, Z.D.; Chen, X.M.; Hu, H.; Zhang, B.C.; Wu, G.; Li, F.; Zhao, X.; et al. The equilibrium between sugars and ethylene is involved in shading-and drought-induced kernel abortion in maize. Plant Growth Regul. 2020, 91, 101–111. [Google Scholar] [CrossRef]
- Begum, N.; Ahanger, M.A.; Su, Y.; Lei, Y.; Mustafa, N.S.A.; Ahmad, P.; Zhang, L. Improved Drought Tolerance by AMF Inoculation in Maize (Zea mays) Involves Physiological and Biochemical Implications. Plants 2019, 8, 579. [Google Scholar] [CrossRef]
- Feng, Q.; Xu, J.; Zhang, Y.; Li, X.; Xu, J.; Han, H.; Ning, T.; Lal, R.; Li, Z. CO2 fixation in above-ground biomass of summer maize under different tillage and straw management treatments. Sci. Rep. 2017, 7, 16888. Available online: https://www.nature.com/articles/s41598-017-17247-8 (accessed on 3 October 2023). [CrossRef]
- Busari, M.A.; Kukal, S.S.; Kaur, A.; Bhatt, R.; Dulazi, A.A. Conservation tillage impacts on soil, crop and the environment Busaria. Int. Soil Water Conserv. Res. 2015, 3, 119–129. [Google Scholar] [CrossRef]
- Abebe, A.; Pathak, H.; Singh, S.D.; Bhatia, A.; Harit, R.C.; Kumar, V. Growth, yield and quality of maize with elevated atmospheric carbon dioxide and temperature in north–west India. Agric. Ecosyst. Environ. 2016, 218, 66–72. [Google Scholar] [CrossRef]
- Poudel, S.; Bhattarai, S.; Sherpa, T.; Karki, A.; Hyun, D.; Kafle, S. The energy input-output analysis of maize production in Sundarharaincha Municipality, Morang district, Nepal. IOP Conf. Ser. Earth Environ. Sci. 2019, 301, 012027. [Google Scholar] [CrossRef]
- Zhang, W.; Yu, C.; Zhang, K.; Zhou, Y.; Tan, W.; Zhang, L. Plant growth regulator and its interactions with environment and genotype affect maize optimal plant density and yield. Eur. J. Agron. 2017, 91, 34–43. [Google Scholar] [CrossRef]
- Hussain, T.; Gollany, H.T.; Hussain, N.; Ahmed, M.; Tahir, M.; Duangpan, S. Synchronizing nitrogen fertilization and planting date to improve resource use efficiency, productivity, and profitability of upland rice. Front. Plant Sci. 2022, 13, 895811. [Google Scholar] [CrossRef]
- Kanavittaya, K.; Nakasathien, S.; Sarobol, E.; Kumdee, O. Growth and development of 3 corn cultivars under different water deficit intervals during early vegetative growth stages. In Proceedings of the 56th Kasetsart University Annual Conference: Plants, Animals, Veterinary Medicine, Agricultural Extension and Home Economics, Bangkok, Thailand, 30 January–2 February 2018; pp. 282–288. [Google Scholar] [CrossRef]
- Farahani, A.; Lebaschi, H.; Hussein, M.; Hussein, S.A.; Reza, V.A.; Jahanfar, D. Effects of arbuscular mycorrhizal fungi, different levels of phosphorus and drought stress on water use efficiency, relative water content and proline accumulation rate of Coriander (Coriandrum sativum L.). J. Med. Plant Res. 2008, 2, 125–131. [Google Scholar]
- Molla, M.S.H.; Nakasathien, S.; Sarobol, E.; Vichukit, V. Effects of nitrogen and water on maize varieties under short and prolonged drought. J. Dev Sustain Agric. 2014, 9, 97–110. [Google Scholar] [CrossRef]
- Diaz-Lopez, E.; Aguilar-Luna, J.M.; Loeza-Corte, J.M. Net assimilation rate and agronomic efficiency of nitrogen in tartago (Ricinus communis L.) (Euphorbiaceae) in dry climate. Scientifica 2020, 2020, 7064745. [Google Scholar] [CrossRef]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998, 135, 1–9. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Emerich, D.W.; Sanchez-Diaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Rosielle, A.A.; Hamblin, J. Theoretical aspects of selection for yield in stress and non-stress environments. Crop Sci. 1981, 21, 943–946. [Google Scholar] [CrossRef]
- Gavuzzi, P.; Rizza, F.; Palumbo, M.; Campaline, R.G.; Ricciardi, G.L.; Borghi, B. Evaluation of field and laboratory predictors of drought and heat tolerance in winter cereals. Can. J. Plant Sci. 1997, 77, 523–531. [Google Scholar] [CrossRef]
- Lin, C.S.; Binns, M.R.; Lefkovitch, L.P. Stability analysis: Where do we stand? Crop Sci. 1986, 26, 894–900. [Google Scholar] [CrossRef]
- Hossain, A.B.S.; Sears, A.G.; Cox, T.S.; Paulsen, G.M. Desiccation tolerance and its relationship to assimilate partitioning in winter wheat. Crop Sci. 1999, 30, 622–627. [Google Scholar] [CrossRef]
- Farshadfar, E.; Javadinia, J. Evaluation of chickpea (Cicer arietinum L.) genotypes for drought tolerance. Seed Plant Improv. J. 2011, 27, 517–537. [Google Scholar]
- Fischer, R.A.; Maurer, R. Drought resistance in spring wheat cultivars. Aust. J. Agric. Res. 1978, 29, 897–912. [Google Scholar] [CrossRef]
- Fischer, R.A.; Wood, J.T. Drought resistance in spring wheat cultivars III. Yield association with morphological traits. Aust. J. Agric. Res. 1979, 30, 1001–1020. [Google Scholar] [CrossRef]
- Lan, J. Comparison of evaluating methods for agronomic drought resistance in crops. Acta Agric Boreali-Occident. Sin. 1998, 7, 85–87. [Google Scholar]
- Moradi, H.; Akbari, G.A.; Khorasani, S.K.; Ramshini, H.A. Evaluation of drought tolerance in corn (Zea mays L.) new hybrids with using stress tolerance indices. Eur. J. Sustain. Dev. 2012, 1, 543–560. [Google Scholar] [CrossRef]
- Moosavi, S.S.; Samadi, Y.B.; Naghavi, M.R.; Zali, A.A.; Dashti, H.; Pourshahbazi, A. Introduction of new indices to identify relative drought tolerance and resistance in wheat genotypes. Desert 2008, 12, 165–178. [Google Scholar] [CrossRef]
- Farshadfar, E.; Sutka, J. Multivariate analysis of drought tolerance in wheat substitution lines. Cereal Res. Commun. 2002, 31, 33–39. [Google Scholar] [CrossRef]
- Farshadfar, E.; Pour Siahbidi, M.M.; Pour Aboughadareh, A.R. Repeatability of drought tolerance indices in bread wheat genotypes. Int. J. Agric. Crop Sci. 2012, 4, 891–903. [Google Scholar]
- Dadbakhsh, A.; Yazdansepas, A.; Ahmadizadeh, M. Study drought stress on yield of wheat (Triticum aestivum L.) genotypes by drought tolerance indices. Adv. Environ. Biol. 2011, 5, 1804–1810. [Google Scholar]
- Fischer, K.S.; Wood, G. Breeding and selection for drought tolerance in tropical maize. In Symposium on Principles and Methods in Crop Improvement for Drought Resistance with Emphasis on Rice; IRRI: Los Baños, Philippines, 1981. [Google Scholar]
- Li, K.R. Land Use Change and Net Emissions of Greenhouse Gases and Terrestrial Ecosystem Carbon Cycle; Weather Press: Beijing, China, 2002. [Google Scholar]
- Lorzadeh, S.H.; Mahdavidamghani, A.; Enayatgholizadeh, M.R.; Yousefi, M. Agrochemical input application and energy use efficiency of maize production systems in Dezful, Iran. Middle-East J. Sci. Res. 2011, 9, 153–156. [Google Scholar]
- Vural, H.; Efecan, I. An analysis of energy use and input costs for maize production in Turkey. J. Food Agric. Environ. 2012, 10, 613–616. [Google Scholar]
- Erdal, G.; Esengun, K.; Erdal, H.; Gunduz, O. Energy use and economical analysis of sugar beet production in Tokat province of Turkey. Energy 2007, 32, 35–41. [Google Scholar] [CrossRef]
- Zhelyazkova, T. Nutritive value and energy efficiency of winter pea production, treated with plant growth regulators. Trakia J. Sci. 2010, 8, 44–51. [Google Scholar]
- Shahin, R.; Mousavi-Avval, S.; Mohammadi, A. Modeling and sensitivity analysis of energy inputs for apple production in Iran. Energy 2010, 35, 3301–3306. [Google Scholar] [CrossRef]
- Chowdhury, A.K.; Zahan, T.; Anowar, M.M.; Islam, M.K.; Molla, S.H.; Khan, A.R. Improvement of potato-jute-t. aman cropping pattern through inclusion of mungbean. Bangladesh Agron. J. 2018, 20, 87–95. [Google Scholar] [CrossRef]
- CIMMYT. From Agronomic Data to Farmer Recommendation: An Economics Training Manual; CIMMYT: Veracruz, Mexico, 1988. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yp | Ys | MP | MRP | SSI | TOL | GMP | REI | STI | MSTIk1 | MSTIk2 | HARM | YI | Red | RDI | DI | GM | ATI | SSPI | SNPI | RDY | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| G1 | 20.16 | 12.72 | 16.44 | 2.19 | 0.84 | 7.44 | 16.02 | 1.19 | 0.69 | 0.70 | 0.90 | 15.60 | 1.16 | 36.90 | 1.13 | 0.73 | 4.42 | 66.53 | 18.96 | 4.03 | 97.43 |
| G2 | 20.31 | 11.70 | 16.00 | 2.10 | 0.96 | 8.61 | 15.41 | 1.10 | 0.62 | 0.66 | 0.70 | 14.85 | 1.07 | 42.41 | 1.03 | 0.61 | 3.72 | 71.12 | 21.94 | 3.74 | 97.62 |
| G3 | 18.40 | 8.45 | 13.43 | 1.71 | 1.22 | 9.95 | 12.47 | 0.72 | 0.40 | 0.35 | 0.24 | 11.58 | 0.77 | 54.08 | 0.82 | 0.35 | 2.70 | 69.27 | 25.35 | 3.09 | 98.45 |
| Yp | Ys | MP | MRP | SSI | TOL | GMP | REI | STI | MSTIk1 | MSTIk2 | HARM | YI | Red | RDI | DI | GM | ATI | SSPI | SNPI | RDY | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ys | 0.95 * | ||||||||||||||||||||
| MP | 0.98 * | 1.00 * | |||||||||||||||||||
| MRP | 0.97 * | 1.00 * | 1.00 * | ||||||||||||||||||
| SSI | −0.93 * | −1.00 * | −0.98 * | −0.99 * | |||||||||||||||||
| TOL | −0.85 * | −0.97 * | −0.94 * | −0.95 * | 0.99 * | ||||||||||||||||
| GMP | 0.97 * | 1.00 * | 1.00 * | 1.00 * | −0.99 * | −0.95 * | |||||||||||||||
| REI | 0.97 * | 1.00 * | 1.00 * | 1.00 * | −0.99 * | −0.95 * | 1.00 * | ||||||||||||||
| STI | 0.95 * | 1.00 * | 1.00 * | 1.00 * | −1.00 * | −0.97 * | 1.00 * | 1.00 * | |||||||||||||
| MSTIk1 | 0.98 * | 0.99 * | 1.00 * | 1.00 * | −0.98 * | −0.93 * | 1.00 * | 1.00 * | 0.99 * | ||||||||||||
| MSTIk2 | 0.93 * | 1.00 * | 0.99 * | 0.99 * | −1.00 * | −0.98 * | 0.99 * | 0.99 * | 1.00 * | 0.98 * | |||||||||||
| HARM | 0.97 * | 1.00 * | 1.00 * | 1.00 * | −0.99 * | −0.95 * | 1.00 * | 1.00 * | 1.00 * | 1.00 * | 0.99 * | ||||||||||
| YI | 0.96 * | 1.00 * | 1.00 * | 1.00 * | −1.00 * | −0.97 * | 1.00 * | 1.00 * | 1.00 * | 0.99 * | 1.00 * | 1.00 * | |||||||||
| Red | −0.92 * | −1.00 * | −0.98 * | −0.99 * | 1.00 * | 0.99 * | −0.99 * | −0.99 * | −1.00 * | −0.98 * | −1.00 * | −0.99 * | −1.00 * | ||||||||
| RDI | 0.92 * | 1.00 * | 0.98 * | 0.99 * | −1.00 * | −0.99 * | 0.99 * | 0.99 * | 1.00 * | 0.98 * | 1.00 * | 0.99 * | 1.00 * | −1.00 * | |||||||
| DI | 0.93 * | 1.00 * | 0.98 * | 0.99 * | −1.00 * | −0.99 * | 0.99 * | 0.99 * | 1.00 * | 0.98 * | 1.00 * | 0.99 * | 1.00 * | −1.00 * | 1.00 * | ||||||
| GM | 0.88 * | 0.98 * | 0.96 * | 0.97 * | −0.99 * | −1.00 * | 0.97 * | 0.97 * | 0.98 * | 0.95 * | 0.99 * | 0.97 * | 0.98 * | −1.00 * | 1.00 * | 0.99 * | |||||
| ATI | −0.04 ns | −0.34 ns | −0.24 ns | −0.28 ns | 0.41 ns | 0.56 * | −0.27 ns | −0.29 ns | −0.34 ns | −0.21 ns | −0.40 ns | −0.28 ns | −0.33 ns | 0.42 ns | −0.42 ns | −0.41 ns | −0.50 * | ||||
| SSPI | −0.85 * | −0.97 * | −0.94 * | −0.95 * | 0.99 * | 1.00 * | −0.95 * | −0.95 * | −0.97 * | −0.93 * | −0.98 * | −0.95 * | −0.97 * | 0.99 * | −0.99 * | −0.99 * | −1.00 * | 0.56 * | |||
| SNPI | 0.93 * | 1.00 * | 0.99 * | 0.99 * | −1.00 * | −0.98 * | 0.99 * | 0.99 * | 1.00 * | 0.98 * | 1.00 * | 0.99 * | 1.00 * | −1.00 * | 1.00 * | 1.00 * | 0.99 * | −0.41 ns | −0.98 * | ||
| RDY | −0.97 * | −1.00 * | −1.00 * | −1.00 * | 0.99 * | 0.95 * | −1.00 * | −1.00 * | −1.00 * | −1.00 * | −0.99 * | −1.00 * | −1.00 * | 0.99 * | −0.99 * | −0.99 * | −0.97 * | 0.28 ns | 0.95 * | −0.99 * | |
| DTE | 0.92 * | 1.00 * | 0.98 * | 0.99 * | −1.00 * | −0.99 * | 0.99 * | 0.99 * | 1.00 * | 0.98 * | 1.00 * | 0.99 * | 1.00 * | −1.00 * | 1.00 * | 1.00 * | 1.00 * | −0.42 ns | −0.99 * | 1.00 * | −0.99 * |
| Treatment | PH-PM (cm) | EH-PM (cm) | EH:PH Ratio (%) | SBD-VT (mm) | LA-V6 (cm2 Plant−1) | LA-V10 (cm2 Plant−1) | LA-VT (cm2 Plant−1) | LA-R3 (cm2 Plant−1) | APB-R3 (g Plant−1) | KNP (No. Plant−1) | 100-KW (g) | KY (t ha−1) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T1 | 192.90 abc | 73.58 b | 38.15 b | 24.49 | 1247.15 | 3203.74 b | 6107.50 b | 4270.00 b | 271.34 a | 352.25 | 25.31 a | 5.26 a |
| T2 | 178.35 c | 65.13 c | 36.52 bc | 24.13 | 1298.87 | 3272.50 b | 5687.50 bc | 3885.00 c | 247.14 bc | 348.12 | 24.57 a | 4.95 a |
| T3 | 193.50 ab | 73.01 b | 37.73 b | 24.42 | 1278.00 | 3632.48 a | 6020.00 b | 4532.50 b | 270.35 ab | 351.14 | 25.15 a | 5.11 a |
| T4 | 183.65 bc | 66.45 c | 36.18 bc | 23.95 | 1246.52 | 2697.77 c | 5372.50 c | 3797.50 c | 245.12 c | 352.11 | 24.95 a | 4.56 b |
| T5 | 160.42 d | 55.20 d | 34.41 c | 23.53 | 1262.96 | 2724.45 c | 4637.50 d | 3412.50 d | 235.21 c | 340.87 | 24.12 a | 4.24 bc |
| T6 | 185.50 bc | 65.25 c | 35.18 bc | 23.90 | 1269.92 | 3628.98 a | 5425.00 c | 3832.50 c | 244.68 c | 350.14 | 24.92 a | 4.51 b |
| T0 | 205.14 a | 99.87 a | 48.68 a | 23.66 | 1260.00 | 3650.22 a | 6825.00 a | 5075.00 a | 251.12 abc | 334.13 | 21.56 b | 4.02 c |
| LSD0.05 | 14.57 | 5.64 | 2.98 | ns | ns | 254.99 | 448.31 | 324.38 | 23.66 | ns | 1.91 | 0.37 |
| Treatment | SLW-VT (g m−2) | LP-PM (%) | RGRA-V6R3 (mg Plant−1 Day−1) | NAR-V6R3 (mg cm−2 Day−1) | RWC-VT (%) | SPAD-VT | RSR-VTR3 (%) | EL-VT (%) | TSS-VT (mg g−1 FW) | Pr-VT (µmol g−1 FW) |
|---|---|---|---|---|---|---|---|---|---|---|
| T1 | 51.71 a | 3.02 c | 25.26 a | 0.91 a | 90.38 ab | 42.75 | 1.67 d | 4.91 b | 1629.58 a | 24.79 a |
| T2 | 50.87 a | 2.11 d | 24.26 b | 0.85 ab | 89.74 ab | 41.02 | 1.83 abc | 4.80 b | 1594.23 a | 23.98 a |
| T3 | 51.27 a | 3.80 b | 25.16 a | 0.88 ab | 93.12 a | 42.96 | 1.71 cd | 4.95 b | 1627.78 a | 24.75 a |
| T4 | 51.04 a | 3.07 c | 24.35 b | 0.87 ab | 90.11 ab | 41.32 | 1.87 ab | 5.11 b | 1610.12 a | 24.15 a |
| T5 | 50.64 a | 1.89 d | 23.70 c | 0.82 b | 88.12 ab | 40.10 | 1.96 a | 5.06 b | 1590.21 a | 23.90 a |
| T6 | 50.79 a | 3.75 b | 24.30 b | 0.84 b | 90.04 ab | 41.28 | 1.76 bcd | 5.01 b | 1605.84 a | 24.03 a |
| T0 | 40.07 b | 14.86 a | 24.28 b | 0.73 c | 85.14 b | 40.12 | 1.63 d | 5.90 a | 1287.05 b | 16.07 b |
| LSD0.05 | 3.89 | 0.48 | 0.14 | 0.07 | 7.01 | ns | 0.14 | 0.40 | 122.98 | 1.83 |
| Indirect Effect via Agronomic Traits | ||||||
| Trait | PH-PM | SBD-VT | LA-VT | 100-KW | KNP | Total Correlation with KY |
| PH-PM | 0.22 | 0.85 | −0.22 | 0.01 | 0.11 | |
| SBD-VT | −0.28 | 0.30 | 0.41 | −0.06 | 0.96 | |
| LA-VT | −0.71 | 0.19 | −0.31 | 0.02 | 0.10 | |
| 100-KW | 0.26 | 0.38 | −0.44 | −0.08 | 0.76 | |
| KNP | 0.06 | 0.44 | −0.22 | 0.60 | 0.80 | |
| Indirect effect via agro-physiological traits | ||||||
| Trait | SLW-VT | LP-PM | NARA-V6R3 | RGRA-V6R3 | HUE | Total correlation with KY |
| SLW-VT | 0.71 | 0.19 | 0.04 | 0.76 | 0.67 | |
| LP-PM | 1.00 | −0.17 | −0.01 | −0.67 | −0.57 | |
| NAR-V6R3 | −0.92 | 0.59 | 0.09 | 0.90 | 0.87 | |
| RGR-V6R3 | −0.24 | 0.04 | 0.13 | 0.71 | 0.79 | |
| HUE | −0.80 | 0.49 | 0.19 | 0.11 | 0.98 | |
| Indirect effect via physio-biochemical traits | ||||||
| Trait | RWC-VT | SPAD-VT | EL-VT | TSS-VT | Pr-VT | Total correlation with KY |
| RWC-VT | 0.81 | 0.95 | −1.41 | 0.78 | 0.81 | |
| SPAD-VT | −0.28 | 0.64 | −0.97 | 0.54 | 0.89 | |
| EL-VT | 0.26 | −0.52 | 1.59 | −0.89 | −0.75 | |
| TSS-VT | −0.28 | 0.56 | 1.14 | 0.93 | 0.68 | |
| Pr-VT | −0.27 | 0.56 | 1.14 | −1.67 | 0.69 | |
| Ethephon | Kernel Yield (t ha−1) | Gross Return (USD ha−1) | Total Variable Cost (USD ha−1) | Gross Margin (USD ha−1) | MBCR (Over No Ethephon T0) |
|---|---|---|---|---|---|
| T1 | 5.26 | 1446.50 | 764.48 | 682.02 | 3.32 |
| T2 | 4.95 | 1361.25 | 867.27 | 493.98 | 1.24 |
| T3 | 5.11 | 1405.25 | 764.48 | 640.77 | 2.92 |
| T4 | 4.56 | 1254.00 | 841.77 | 412.23 | 0.83 |
| T5 | 4.24 | 1166.00 | 1021.77 | 144.23 | 0.17 |
| T6 | 4.51 | 1240.25 | 841.77 | 398.48 | 0.75 |
| T0 | 4.02 | 1105.50 | 661.77 | 443.73 | - |
| Sl. | Index | Equation | Reference | Sl. | Index | Equation | Reference |
|---|---|---|---|---|---|---|---|
| 1. | MP | [152] | 11. | YI | [153,154] | ||
| 2. | MRP | [155] | 12. | Red | [156] | ||
| 3. | SSI | [157] | 13. | RDI | [158] | ||
| 4. | TOL | [152] | 14. | DI | [159] | ||
| 5. | GMP | [91] | 15. | GM | [160] | ||
| 6. | REI | [155] | 16. | ATI | [161] | ||
| 7. | STI | [91] | 17. | SSPI | [161] | ||
| 8. | MSTIk1 | [162] | 18. | SNPI | [161] | ||
| 9. | MSTIk2 | [162] | 19. | RDY | [163] | ||
| 10. | HARM | [164] | 20. | DTE | [165] |
| Equipment/Inputs | Unit | Energy Equivalent Coefficient (MJ/Unit) | Reference | Quantity per Hectare |
|---|---|---|---|---|
| A. Inputs | ||||
| 1. Human labor | H | 1.96 | [142,167] | T0 = 378.9; T1, T3, T4, T6 = 402.9; T2, T5 = 426.9 |
| 2. Machinery | H | 64.8 | [168] | 30.4 |
| 3. Nitrogen (N) | kg | 66.14 | [167,169] | 194 |
| 4. Phosphorus (P) | kg | 12.44 | [167,169] | 50 |
| 5. Potassium (K) | kg | 11.15 | [169] | 50 |
| 6. Pesticides | kg | 120 | [142] | 0.31 |
| 7. Plant growth regulator (PGR) | kg | 85 | [170] | T0 = 0.0; T1, T3 = 0.281; T2, T4 T6 = 0.562; T5 = 1.124 |
| 8. Diesel | L | 56.31 | [171] | 524.2 |
| 9. Seed of maize | kg | 14.7 | [142] | 25 |
| B. Output | ||||
| 1. Maize | kg | 14.7 | [167] | Treatment-wise grain yield |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molla, M.S.H.; Kumdee, O.; Wongkaew, A.; Khongchiu, P.; Worathongchai, N.; Alam, M.R.; Mahmud, A.-A.; Nakasathien, S. Potentiality of Sustainable Maize Production under Rainfed Conditions in the Tropics by Triggering Agro-Physio-Biochemical Traits Ascertained from a Greenhouse. Plants 2023, 12, 4192. https://doi.org/10.3390/plants12244192
Molla MSH, Kumdee O, Wongkaew A, Khongchiu P, Worathongchai N, Alam MR, Mahmud A-A, Nakasathien S. Potentiality of Sustainable Maize Production under Rainfed Conditions in the Tropics by Triggering Agro-Physio-Biochemical Traits Ascertained from a Greenhouse. Plants. 2023; 12(24):4192. https://doi.org/10.3390/plants12244192
Chicago/Turabian StyleMolla, Md. Samim Hossain, Orawan Kumdee, Arunee Wongkaew, Phanuphong Khongchiu, Nattaporn Worathongchai, Md. Robiul Alam, Abdullah-Al Mahmud, and Sutkhet Nakasathien. 2023. "Potentiality of Sustainable Maize Production under Rainfed Conditions in the Tropics by Triggering Agro-Physio-Biochemical Traits Ascertained from a Greenhouse" Plants 12, no. 24: 4192. https://doi.org/10.3390/plants12244192
APA StyleMolla, M. S. H., Kumdee, O., Wongkaew, A., Khongchiu, P., Worathongchai, N., Alam, M. R., Mahmud, A.-A., & Nakasathien, S. (2023). Potentiality of Sustainable Maize Production under Rainfed Conditions in the Tropics by Triggering Agro-Physio-Biochemical Traits Ascertained from a Greenhouse. Plants, 12(24), 4192. https://doi.org/10.3390/plants12244192