
Study of the Photosynthesis Response during the Gradual Lack of Water for 14 Olea europaea L. subsp europaea Cultivars and Their Adaptation to Climate Change


 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Curve Fitting
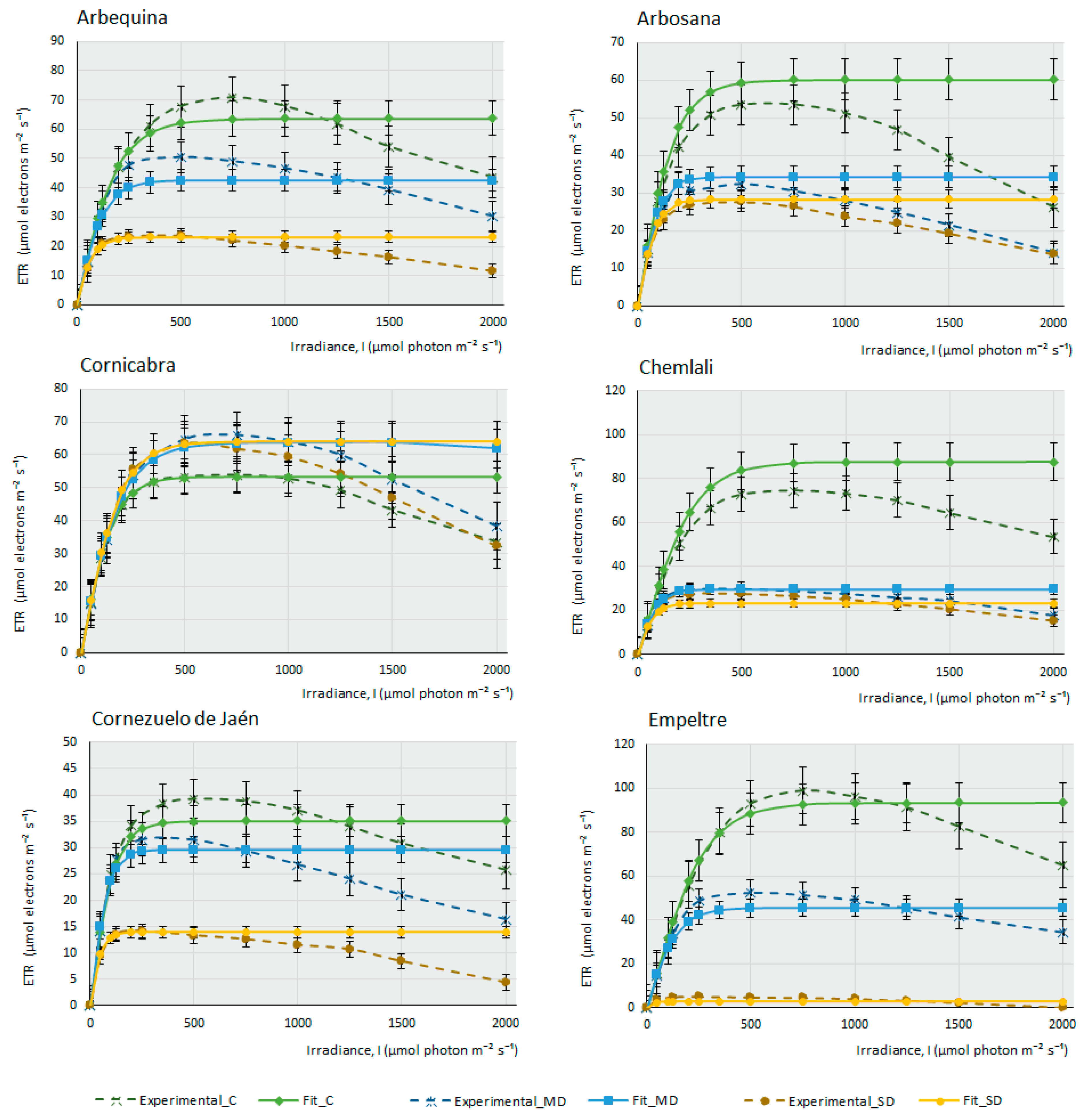
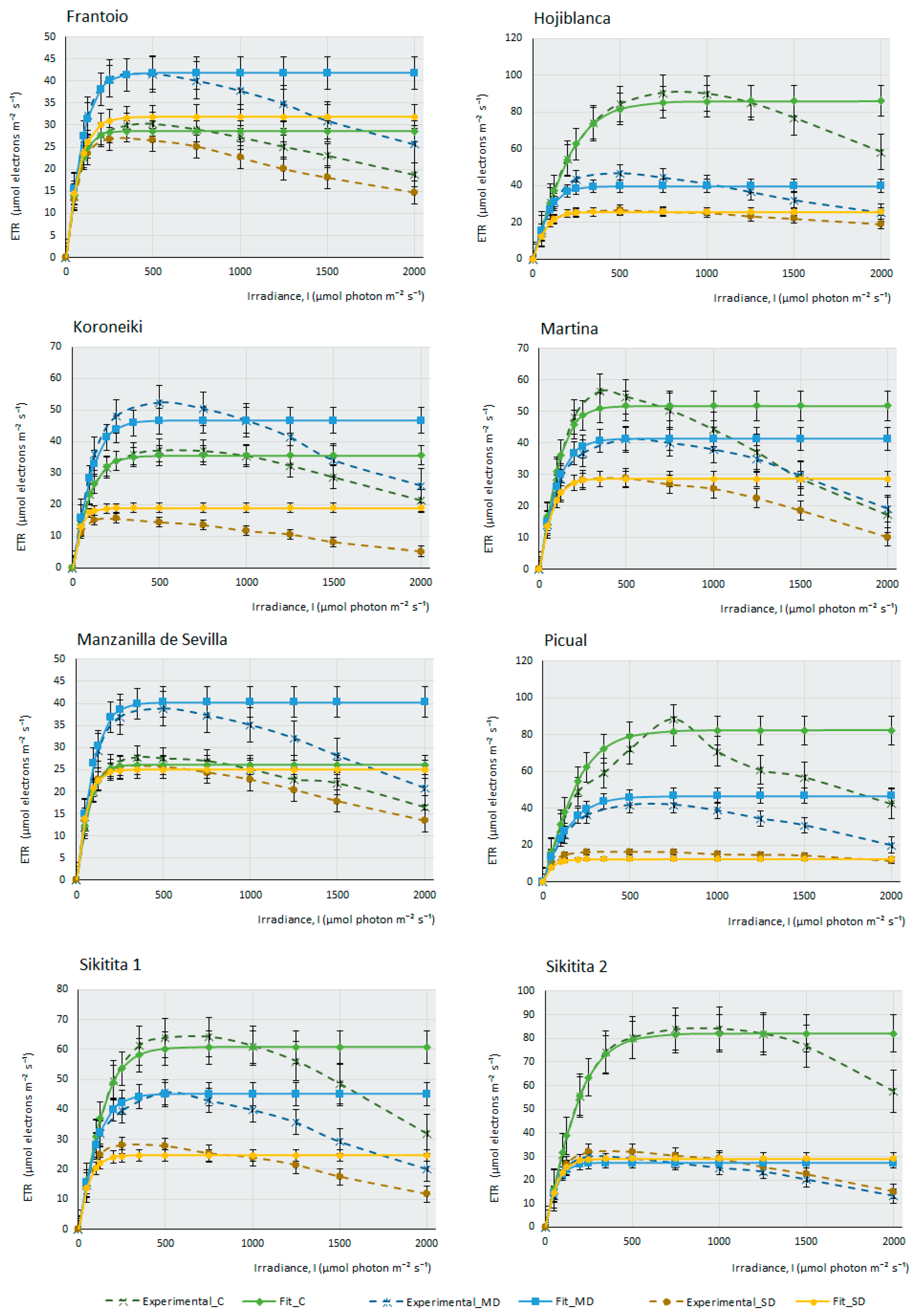
2.2. Photosynthesis-Light Response and ETR-Light Curves of the 14 Olive Cultivars
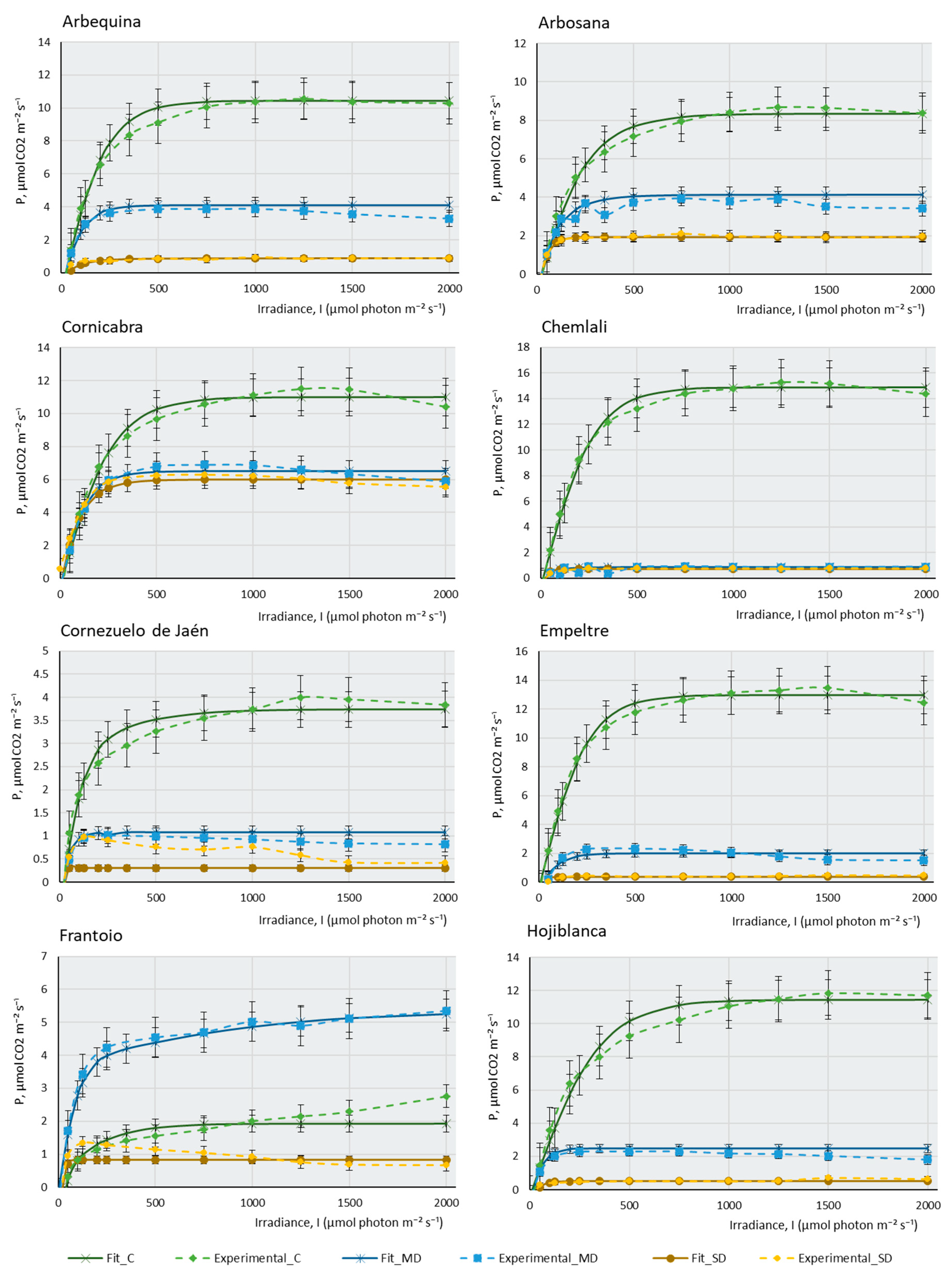
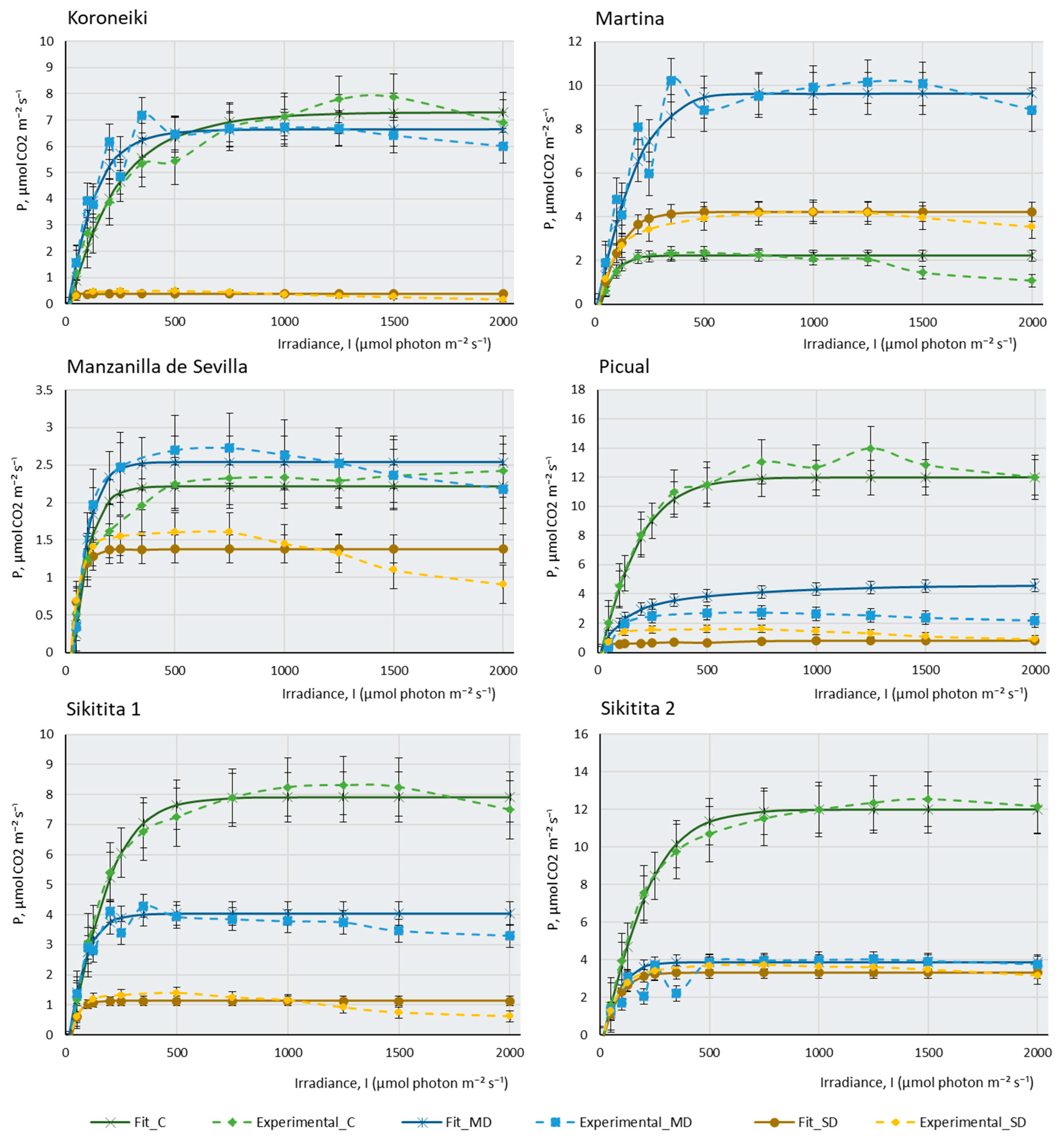
3. Materials and Methods
3.1. Plant Material
3.2. Curve Fitting
3.3. Photosynthesis and Fluorescence versus Light Response Curves
3.4. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Convention on Biological Diversity; Treaty Collection: 1992; United Nations Environment Programme: Nairobi, Kenya, 1992; Available online: https://wedocs.unep.org/20.500.11822/8340 (accessed on 27 November 2023).
- Vijay, V.; Pimm, S.L.; Jenkins, C.N.; Smith, S.J. The Impacts of Oil Palm on Recent Deforestation and Biodiversity Loss. PLoS ONE 2016, 11, e0159668. [Google Scholar] [CrossRef] [PubMed]
- Leclère, D.; Obersteiner, M.; Barrett, M.; Butchart, S.H.; Chaudhary, A.; De Palma, A.; DeClerck, F.A.J.; Di Marco, M.; Doelman, J.C.; Dürauer, M.; et al. Bending the curve of terrestrial biodiversity needs an integrated strategy. Nature 2020, 585, 551–556. [Google Scholar] [CrossRef]
- Pyšek, P.; Hulme, P.E.; Simberloff, D.; Bacher, S.; Blackburn, T.M.; Carlton, J.T.; Dawson, W.; Essl, F.; Foxcroft, L.C.; Genovesi, P.; et al. Scientists’ warning on invasive alien species. Biol. Rev. 2020, 95, 1511–1534. [Google Scholar] [CrossRef] [PubMed]
- Belaj, A.; Gurbuz Veral, M.; Sikaoui, H.; Moukhli, A.; Khadari, B.; Mariotti, R.; Baldoni, L. Olive genetic resources. In The Olive Tree Genome; Springer: Cham, Switzerland, 2016; pp. 27–54. [Google Scholar]
- Bito, I.; Doron-Faigenboim, A.; Jamwal, M.; Mani, Y.; Eshed, R.; Rosen, A.; Sherman, A.; Ophir, R.; Lavee, S.; Avidan, B.; et al. Development of a large set of SNP markers for assessing phylogenetic relationships between the olive cultivars composing the Israeli olive germplasm collection. Mol. Breed. 2015, 35, 107. [Google Scholar] [CrossRef]
- Fanelli, V.; Mascio, I.; Falek, W.; Miazzi, M.M.; Montemurro, C. Current Status of Biodiversity Assessment and Conservation of Wild Olive (Olea europaea L. subsp europaea var. sylvestris). Plants 2022, 11, 480. [Google Scholar] [CrossRef] [PubMed]
- León, L.; De la Rosa, R.; Velasco, L.; Belaj, A. Using wild olives in breeding programs: Implications on oil quality composition. Front. Plant Sci. 2018, 9, 232. [Google Scholar] [CrossRef] [PubMed]
- Rugini, E.; De Pace, C. Olive Breeding with Classical and Modern Approaches. In The Olive Tree Genome. Compendium of Plant Genomes; Rugini, E., Baldoni, L., Muleo, R., Sebastiani, L., Eds.; Springer: Cham, Switzerland, 2016; pp. 163–193. [Google Scholar] [CrossRef]
- Colella, C.; Miacola, C.; Amenduni, M.; D’Amico, M.; Bubici, G.; Cirulli, M. Sources of verticillium wilt resistance in wild olive germplasm from the Mediterranean region. Plant Pathol. 2008, 57, 533–539. [Google Scholar] [CrossRef]
- Arias-Calderon, R.; Rodriguez-Jurado, D.; Leon, L.; Bejarano-Alcazar, J.; De la Rosa, R.; Belaj, A. Pre-breeding for resistance to Verticillium wilt in olive: Fishing in the wild relative gene pool. Crop Prot. 2015, 75, 25–33. [Google Scholar] [CrossRef]
- Aranda-Barranco, S.; Kowalski, A.S.; Serrano-Ortiz, P.; Sanchez-Canete, E.P. Net CO2 Assimilation and Transpiration of Olive Trees: Effect of Weed Cover at the Leaf Scale. AGU Fall Meet. Abstr. 2020, 2020, B002-0003. Available online: https://ui.adsabs.harvard.edu/abs/2020AGUFMB002.0003A (accessed on 10 June 2023).
- Aranda-Barranco, S.; Kowalski, A.S.; Serrano-Ortiz, P.; Sánchez-Cañete, E.P. Influence of weed cover on leaf-level CO2 and H2O fluxes in an olive grove. In Proceedings of the EGU General Assembly Conference Abstracts, Online, 4–8 May 2020; p. 5181. [Google Scholar] [CrossRef]
- Barranco Navero, D.; Cimato, A.; Fiorino, P.; Rallo Romero, L.; Touzani, A.; Castañeda, C.; Serafini, F.; Trujillo Navas, I. Catálogo Mundial de Variedades de Olivo; Ediciones Mundi-Prensa: Madrid, Spain, 2004; ISBN 8484761924. [Google Scholar]
- Cardona, T.; Shao, S.; Nixon, P.J. Enhancing photosynthesis in plants: The light reactions. Essays Biochem. 2018, 62, 85–94. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development, 6th ed.; Sinauer Associates Incorporated: Sunderland, MA, USA, 2015; p. 761. [Google Scholar]
- Blankenship, R.E. Molecular Mechanisms of Photosynthesis; John Wiley & Sons: Hoboken, NJ, USA, 2021. [Google Scholar]
- Mathur, S.; Jain, L.; Jajoo, A. Photosynthetic efficiency in sun and shade plants. Photosynthetica 2018, 56, 354–365. [Google Scholar] [CrossRef]
- Yang, X.; Lu, M.; Wang, Y.; Wang, Y.; Liu, Z.; Chen, S. Response mechanism of plants to drought stress. Horticulturae 2021, 7, 50. [Google Scholar] [CrossRef]
- Tholen, D.; Ethier, G.; Genty, B.; Pepin, S.; Zhu, X.G. Variable mesophyll conductance revisited: Theoretical background and experimental implications. Plant Cell Environ. 2012, 35, 2087–2103. [Google Scholar] [CrossRef]
- Loreto, F.; Centritto, M.; Chartzoulakis, K. Photosynthetic limitations in olive cultivars with different sensitivity to salt stress. Plant Cell Environ. 2003, 26, 595–601. [Google Scholar] [CrossRef]
- Bhusal, N.; Han, S.G.; Yoon, T.M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh.). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Rico, E.I.; de la Fuente, G.C.M.; Morillas, A.O.; Ocaña, A.M.F. Physiological and biochemical study of the drought tolerance of 14 main olive cultivars in the Mediterranean basin. Photosynth. Res. 2023; Online ahead of print. [Google Scholar] [CrossRef]
- Brito, C.; Dinis, L.T.; Moutinho-Pereira, J.; Correia, C.M. Drought stress effects and olive tree acclimation under a changing climate. Plants 2019, 8, 232. [Google Scholar] [CrossRef] [PubMed]
- Baccari, S.; Elloumi, O.; Chaari-Rkhis, A.; Fenollosa, E.; Morales, M.; Drira, N.; Ben Abdallah, F.; Fki, L.; Munné-Bosch, S. Linking leaf water potential, photosynthesis and chlorophyll loss with mechanisms of photo-and antioxidant protection in juvenile olive trees subjected to severe drought. Front. Plant Sci. 2020, 11, 614144. [Google Scholar] [CrossRef]
- Fernández, J.E.; Díaz-Espejo, A.; Infante, J.M.; Durán, P.; Palomo, M.J.; Chamorro, V.; Girón, I.F.; Villagarcía, L. Water relations and gas exchange in olive trees under regulated deficit irrigation and partial rootzone drying. Plant Soil 2006, 284, 273–291. [Google Scholar] [CrossRef]
- Sofo, A.; Dichio, B.; Montanaro, G.; Xiloyannis, C. Shade effect on photosynthesis and photoinhibition in olive during drought and rewatering. Agric. Water Manag. 2009, 96, 1201–1206. [Google Scholar] [CrossRef]
- Diaz-Espejo, A.; Nicolas, E.; Fernandez, J.E. Seasonal evolution of diffusional limitations and photosynthetic capacity in olive under drought. Plant Cell Environ. 2007, 30, 922–933. [Google Scholar] [CrossRef]
- Bacelar, E.A.; Moutinho-Pereira, J.M.; Gonçalves, B.C.; Lopes, J.I.; Correia, C.M. Physiological responses of different olive genotypes to drought conditions. Acta Physiol. Plant. 2009, 31, 611–621. [Google Scholar] [CrossRef]
- Zhu, X.G.; Long, S.P.; Ort, D.R. Improving photosynthetic efficiency for greater yield. Annu. Rev. Plant Biol. 2010, 61, 235–261. [Google Scholar] [CrossRef] [PubMed]
- Nayak, N.; Mehrotra, R.; Mehrotra, S. Carbon biosequestration strategies: A review. Carbon Capture Sci. Technol. 2022, 4, 100065. [Google Scholar] [CrossRef]
- Galan-Martin, A.; del Mar Contreras, M.; Romero, I.; Ruiz, E.; Bueno-Rodríguez, S.; Eliche-Quesada, D.; Castro-Galiano, E. The potential role of olive groves to deliver carbon dioxide removal in a carbon-neutral Europe: Opportunities and challenges. Renew. Sustain. Energy Rev. 2022, 165, 112609. [Google Scholar] [CrossRef]
- Ministerio de Agricultura, Pesca y Alimentación. La Política Agrícola Común 2023–2027 y el Plan Estratégico de la Política Agrícola Común de España (PEPAC). 2023. Available online: https://www.mapa.gob.es/es/pac/pac-2023-2027/) (accessed on 21 May 2023).
- Farquhar, G.D.; von Caemmerer, S.V.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef]
- Boussadia, O.; Mariem, F.B.; Mechri, B.; Boussetta, W.; Braham, M.; El Hadj, S.B. Response to drought of two olive tree cultivars (cv Koroneki and Meski). Sci. Hortic. 2008, 116, 388–393. [Google Scholar] [CrossRef]
- Boughalleb, F.; Hajlaoui, H. Physiological and anatomical changes induced by drought in two olive cultivars (cv Zalmati and Chemlali). Acta Physiol. Plant. 2011, 33, 53–65. [Google Scholar] [CrossRef]
- Prioul, J.L.; Chartier, P. Partitioning of transfer and carboxylation components of intracellular resistance to photosynthetic CO2 fixation: A critical analysis of the methods used. Ann. Bot. 1977, 41, 789–800. [Google Scholar] [CrossRef]
- Dias, M.C.; Correia, S.; Serôdio, J.; Silva, A.M.S.; Freitas, H.; Santos, C. Chlorophyll fluorescence and oxidative stress endpoints to discriminate olive cultivars tolerance to drought and heat episodes. Sci. Hortic. 2018, 231, 31–35. [Google Scholar] [CrossRef]
- Faraloni, C.; Cutino, I.; Petruccelli, R.; Leva, A.R.; Lazzeri, S.; Torzillo, G. Chlorophyll fluorescence technique as a rapid tool for in vitro screening of olive cultivars (Olea europaea L.) tolerant to drought stress. Environ. Exp. Bot. 2011, 73, 49–56. [Google Scholar] [CrossRef]
- Hyams, D.G. CurveExpert Software. 2010. Available online: http://www.curveexpert.net (accessed on 12 May 2023).
- Jassby, A.D.; Platt, T. Mathematical formulation of the relationship between photosynthesis and light for phytoplankton. Limnol. Oceanogr. 1976, 21, 540–547. [Google Scholar] [CrossRef]
- de A. Lobo, F.; de Barros, M.P.; Dalmagro, H.J.; Dalmolin, A.C.; Pereira, W.E.; de Souza, E.C.; Vourlitis, G.L.; Rodriguez-Ortiz, C.E. Fitting net photosynthetic light-response curves with Microsoft Excel—A critical look at the models. Photosynthetica 2013, 51, 445–456. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Wong, S.C. An Empirical Model of Stomatal Conductance. Funct. Plant Biol. 1984, 11, 191–210. [Google Scholar] [CrossRef]
- Schreiber, U.; Schliwa, U.; Bilger, W. Continuous recording of photochemical and non-photochemical chlorophyll fluorescence quenching with a new type of modulation fluorometer. Photosynth. Res. 1986, 10, 51–62. [Google Scholar] [CrossRef]
- Eichelmann, H.; Oja, V.; Rasulov, B.; Padu, E.; Bichele, I.; Pettai, H.; Niinemets, U.; Laisk, A. Development of leaf photosynthetic parameters in Betula pendula Roth leaves: Correlations with photosystem I density. Plant Biol. 2004, 6, 307–318. [Google Scholar] [CrossRef]
- Fernández-Solas, Á.; Fernández-Ocaña, A.M.; Almonacid, F.; Fernández, E.F. Potential of agrivoltaics systems into olive groves in the Mediterranean region. Appl. Energy 2023, 352, 121988. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Condition | Amax | φ_PSII | Rd | ISAT_A50 | ISAT_A99.9 | Jmax | Tetha | Fv/Fm |
|---|---|---|---|---|---|---|---|---|---|
| Name | Symbol | µmol CO₂ m−²·s−¹ | µmol CO₂/µmol photons | µmol CO₂ m−²·s−¹ | µmol photons m−²·s−¹ | µmol photons m−²·s−¹ | µmol CO₂ m−²·s−¹ | - | - |
| AE | Control | 11.15 a ± 1.72 ABC | 0.0497 a ± 0.0035 AB | 0.91 a ± 0.05 A | 145.0 a ± 7.3 A | 925.1 a ± 46.3 A | 205.5 a ± 31.08 B | 0.246 a ± 0.012 B | 0.827 a ± 0.041 A |
| MD | 4.87 b ± 1.54 BCD’ | 0.0464 a ± 0.0121 AB’ | 1.16 a ± 0.06 AB’ | 80.5 b ± 5.8 A’ | 425.7 b ± 27.9 A’ | 86.9 a ± 38.3 B’ | 0.213 a ± 0.01 A’ | 0.830 a ± 0.041 A’ | |
| SD | 1.55 c ± 0.40 C’’ | 0.0340 a ± 0.0166 C’’ | 0.96 a ± 0.08 A’’ | 66.7 b ± 11.0 A’’ | 289.8 b ± 51.6 A’’ | 27.7 a ± 6.9 A’’ | 0.350 a ± 0.055 AB’’ | 0.801 a ± 0.030 A’’ | |
| Ab | Control | 9.16 a ± 2.01 BC | 0.0332 a ± 0.0087 CD | 0.80 a ± 0.04 A | 114.3 a ± 5.7 A | 888.9 a ± 44.5 A | 222.4 a ± 46.3 B | 0.189 a ± 0.009 B | 0.829 a ± 0.041 A |
| MD | 3.87 b ± 0.95 CD’ | 0.0452 a ± 0.0151 ABCD’ | 0.85 a ± 0.07 AB’ | 72.0 a ± 6.4 A’ | 415.7 a ± 42.3 A’ | 75.7 a ± 37.6 B’ | 0.146 a ± 0.008 A’ | 0.775 a ± 0.038 A’ | |
| SD | 2.60 c ± 0.68 BC’’ | 0.0454 a ± 0.0076 C’’ | 0.89 a ± 0.04 A’’ | 96.8 a ± 20.5 A’’ | 459.0 a ± 84.2 A’’ | 47.9 a ± 18.5 A’’ | 0.192 a ± 0.005 B’’ | 0.751 a ± 0.036 A’’ | |
| C | Control | 10.92 a ± 2.09 ABC | 0.0438 a ± 0.0062 ABC | 0.77 a ± 0.04 A | 143.8 a ± 7.2 A | 921.9 a ± 46.1 A | 193.9 a ± 44.4 B | 0.118 a ± 0.006 B | 0.836 a ± 0.042 A |
| MD | 6.92 b ± 1.88 AB’ | 0.0512 a ± 0.0114 ABCD’ | 0.59 a ± 0.07 B’ | 95.2 a ± 36.4 A’ | 611.3 a ± 255.7 A’ | 86.7 b ± 15.6 B’ | 0.154 a ± 0.285 A’ | 0.806 a ± 0.039 A’ | |
| SD | 6.45 b ± 1.98 A’’ | 0.0436 a ± 0.0066 C’’ | 0.11 a ± 0.01 A’’ | 86.8 a ± 8.5 A’’ | 611.1 a ± 89.6 A’’ | 81.5 b ± 26.5 A’’ | 0.348 a ± 0.085 AB’’ | 0.818 a ± 0.039 A’’ | |
| Ch | Control | 14.82 a ± 1.72 A | 0.0562 a ± 0.0023 A | 0.58 a ± 0.03 A | 180.8 a ± 9.0 A | 1203.0 a ± 60.2 A | 200.5 a ± 37.9 B | 0.266 a ± 0.013 B | 0.804 a ± 0.040 A |
| MD | 1.50 b ± 0.49 D’ | 0.0336 a ± 0.0074 DCD’ | 0.71 a ± 0.02 B’ | 54.5 b ± 6.4 A’ | 273.7 b ± 37.4 A’ | 56.9 b ± 26.1 B’ | 0.170 a ± 0.012 A’ | 0.701 b ± 0.038 A’ | |
| SD | 1.43 b ± 0.17 C’’ | 0.0301 a ± 0.0081 D’’ | 0.72 a ± 0.02 A’’ | 32.9 b ± 2.1 A’’ | 148.4 b ± 10.8 A’’ | 42.2 b ± 11.3 A’’ | 0.113 a ± 0.012 B’’ | 0.723 ab ± 0.031 A’’ | |
| CJ | Control | 3.69 a ± 1.83 DE | 0.0400 a ± 0.0066 ABC | 0.61 a ± 0.04 A | 161.6 a ± 11.7 A | 810.9 a ± 57.5 A | 90.1 a ± 11.0 B | 0.059 a ± 0.009 B | 0.800 a ± 0.037 A |
| MD | 1.80 b ± 0.45 D’ | 0.0469 b ± 0.0072 CD’ | 0.73 a ± 0.06 B’ | 51.9 b ± 4.1 A’ | 273.2 ab ± 27.7 A’ | 63.1 a ± 19.9 B’ | 0.340 a ± 0.034 A’ | 0.781 a ± 0.706 A’ | |
| SD | 1.05 b ± 0.08 C’’ | 0.0451 b ± 0.0048 BC’’ | 0.79 a ± 0.04 A’’ | 6.4 b ± 1.4 A’’ | 33.1 b ± 7.2 A’’ | 57.2 a ± 6.4 A’’ | 0.179 a ± 0.037 B’’ | 0.648 a ± 0.028 A’’ | |
| E | Control | 12.99 a ± 2.40 AB | 0.0510 a ± 0.0067 AB | 0.56 a ± 0.03 A | 126.1 a ± 5.2 A | 825.0 a ± 33.1 A | 181.3 a ± 28.3 B | 0.268 a ± 0.013 AB | 0.826 a ± 0.000 A |
| MD | 2.40 b ± 1.66 D’ | 0.0613 a ± 0.0067 A’ | 1.10 a ± 0.08 AB’ | 56.5 ab ± 3.7 A’ | 319.0 ab ± 22.9 A’ | 132.5 a ± 51.9 B’ | -0.025 a ± 0.010 A’ | 0.754 a ± 0.029 A’ | |
| SD | 1.37 b ± 0.34 C’’ | 0.0680 a ± 0.0070 A’’ | 0.89 a ± 0.04 A’’ | 23.1 b ± 3.2 A’’ | 90.4 b ± 12.5 A’’ | 82.6 a ± 2.8 A’’ | 0.394 a ± 0.03 AB’’ | 0.738 a ± 0.030 A’’ | |
| F | Control | 2.15 a ± 0.81 E | 0.0479 a ± 0.0103 ABC | 0.97 a ± 0.06 A | 180.7 a ± 7.7 A | 553.9 a ± 51.1 A | 232.9 a ± 46.3 A | 0.060 a ± 0.006 B | 0.820 a ± 0.000 A |
| MD | 5.22 b ± 1.00 ABC’ | 0.0540 a ± 0.0103 ABC’ | 0.32 a ± 0.04 B’ | 136.6 a ± 24.4 A’ | 951.7 a ± 187.5 A’ | 69.5 b ± 16.2 AB’ | 0.398 a ± 0.049 A’ | 0.781 a ± 0.000 A’ | |
| SD | 1.69 a ± 0.48 BC’’ | 0.0639 a ± 0.0053 A’’ | 0.59 a ± 0.05 A’’ | 100.7 a ± 2.0 A’’ | 695.2 a ± 178.6 A’’ | 1.2 b ± 0.6 A’’ | 0.603 a ± 0.040 AB’’ | 0.714 a ± 0.018 A’’ | |
| H | Control | 11.35 a ± 1.66 ABC | 0.0354 a ± 0.0040 BCD | 0.58 a ± 0.03 A | 65.9 a ± 5.8 A | 405.9 a ± 40.6 A | 260.4 a ± 36.3 B | -0.019 a ± 0.001 B | 0.808 a ± 0.039 A |
| MD | 2.74 b ± 0.81 D’ | 0.0507 a ± 0.0134 BCD’ | 0.67 a ± 0.05 AB’ | 59.8 a ± 6.9 A’ | 348.7 a ± 50.5 A’ | 51.5 a ± 24.8 B’ | 0.363 a ± 0.033 A’ | 0.804 a ± 0.037 A’ | |
| SD | 1.22 b ± 0.33 C’’ | 0.0388 a ± 0.0099 CD’’ | 0.68 a ± 0.05 A’’ | 50.5 a ± 3.4 A’’ | 253.1 a ± 30.6 A’’ | 35.7 a ± 9.7 A’’ | -0.177 a ± 0.076 AB’’ | 0.804 a ± 0.037 A’’ | |
| K | Control | 7.57 a ± 2.00 CD | 0.0173 a ± 0.0061 D | 0.55 a ± 0.06 A | 212.4 a ± 10.5 A | 1294.8 a ± 71.8 A | 345.3 a ± 55.7 B | 0.093 a ± 0.008 B | 0.799 a ± 0.039 A |
| MD | 6.82 a ± 1.25 ABCD’ | 0.0449 ab ± 0.0105 ABCD’ | 0.63 a ± 0.03 AB’ | 79.9 b ± 5.6 A’ | 509.3 b ± 39.4 A’ | 89.6 b ± 39.4 B’ | 0.151 a ± 0.016 A’ | 0.696 b ± 0.038 A’ | |
| SD | 1.05 b ± 0.09 C’’ | 0.0481 b ± 0.0174 C’’ | 0.78 a ± 0.02 A’’ | 27.6 b ± 2.3 A’’ | 157.9 b ± 13.5 A’’ | 30.2 b ± 9.2 A’’ | 0.295 a ± 0.029 AB’’ | 0.680 b ± 0.039 A’’ | |
| M | Control | 2.96 a ± 0.97 DE | 0.0471 a ± 0.0050 ABC | 0.65 a ± 0.03 A | 118.4 a ± 7.6 A | 702.9 a ± 50.7 A | 47.8 a ± 17.9 B | 1.614 a ± 0.159 A | 0.803 a ± 0.040 A |
| MD | 9.55 b ± 2.20 A’ | 0.0460 a ± 0.077 BCD’ | 0.66 a ± 0.02 B’ | 112.9 a ± 3.9 A’ | 699.7 a ± 42.5 A’ | 127.6 b ± 36.5 B’ | -0.012 b ± 0.006 A’ | 0.772 a ± 0.002 A’ | |
| SD | 4.43 a ± 1.29 AB’’ | 0.0385 a ± 0.0081 C’’ | 0.94 a ± 0.05 A’’ | 78.1 a ± 4.4 A’’ | 414.5 a ± 24.0 A’’ | 95.9 ab ± 29.4 A’’ | 0.124 b ± 0.007 B’’ | 0.784 a ± 0.038 A’’ | |
| MS | Control | 3.04 a ± 0.10 DE | 0.0384 a ± 0.0110 BC | 1.30 a ± 0.07 A | 89.2 a ± 4.7 A | 445.2 a ± 18.3 A | 105.0 a ± 9.5 B | 0.161 a ± 0.009 B | 0.804 a ± 0.040 A |
| MD | 3.93 a ± 0.79 BCD’ | 0.0475 a ± 0.0087 ABCD’ | 1.63 a ± 0.11 A’ | 92.4 a ± 5.3 A’ | 452.3 a ± 30.5 A’ | 116.5 a ± 31.1 B’ | 0.222 a ± 0.021 A’ | 0.804 a ± 0.000 A’ | |
| SD | 1.96 a ± 0.93 BC’’ | 0.0537 a ± 0.0103 AB’’ | 1.05 a ± 0.07 A’’ | 45.1 a ± 2.6 A’’ | 232.4 a ± 18.0 A’’ | 81.8 a ± 27.6 A’’ | 0.254 a ± 0.018 B’’ | 0.782 a ± 0.036 A’’ | |
| P | Control | 12.65 a ± 3.06 AB | 0.0540 a ± 0.0068 A | 0.94 a ± 0.04 A | 127.1 a ± 6.1 A | 809.7 a ± 39.2 A | 132.2 a ± 54.6 B | 0.258 a ± 0.012 B | 0.800 a ± 0.040 A |
| MD | 5.06 b ± 2.95 BCD’ | 0.0412 a ± 0.0117 D’ | 0.59 a ± 0.06 B’ | 94.8 a ± 18.3 A’ | 620.9 a ± 153.3 A’ | 64.6 a ± 14.5 A’ | 0.160 a ± 0.021 A’ | 0.787 a ± 0.034 A’ | |
| SD | 1.18 b ± 0.16 C’’ | 0.0456 a ± 0.0167 CD’’ | 0.76 a ± 0.04 A’’ | 58.1 a ± 18.2 A’’ | 284.7 a ± 93.3 A’’ | 46.7 a ± 3.5 A’’ | 1.755 a ± 0.383 A’’ | 0.623 a ± 0.023 A’’ | |
| S1 | Control | 8.08 a ± 1.69 CD | 0.0353 a ± 0.0098 CD | 0.67 a ± 0.04 A | 107.5 a ± 5.5 A | 640.2 a ± 34.6 A | 186.1 a ± 25.6 B | 0.213 a ± 0.010 B | 0.835 a ± 0.042 A |
| MD | 4.10 b ± 1.07 BCD’ | 0.0491 a ± 0.0090 ABCD’ | 0.76 a ± 0.06 AB’ | 52.8 b ± 3.1 A’ | 303.9 b ± 19.8 A’ | 35.0 a ± 18.4 B’ | 0.191 a ± 0.009 A’ | 0.733 a ± 0.036 A’ | |
| SD | 2.20 c ± 1.03 C’’ | 0.0466 a ± 0.0079 BC’’ | 0.84 a ± 0.04 A’’ | 32.3 b ± 2.0 A’’ | 173.7 b ± 13.9 A’’ | 27.9 a ± 4.4 A’’ | 0.329 a ± 0.024 AB’’ | 0.724 a ± 0.034 A’’ | |
| S2 | Control | 12.15 a ± 1.82 ABC | 0.0454 a ± 0.0034 ABC | 0.75 a ± 0.04 A | 157.1 a ± 8.1 A | 879.8 a ± 39.2 A | 253.3 a ± 52.4 B | 0.209 a ± 0.011 B | 0.820 a ± 0.000 A |
| MD | 4.18 b ± 0.70 BCD’ | 0.0459 a ± 0.0063 BCD’ | 0.73 a ± 0.04 B’ | 71.2 b ± 3.6 A’ | 415.7 b ± 21.0 A’ | 83.9 b ± 19.1 B’ | 0.226 a ± 0.010 A’ | 0.764 a ± 0.038 A’ | |
| SD | 3.54 b ± 0.90 BC’’ | 0.0457 a ± 0.0071 BC’’ | 0.76 a ± 0.03 A’’ | 64.2 b ± 3.2 A’’ | 364.2 b ± 17.8 A’’ | 54.2 b ± 16.0 A’’ | 0.242 a ± 0.013 B’’ | 0.778 a ± 0.038 A’’ |
| Cultivar | Condition | ETR_max | Alpha-ETR (α) | ETRd | ISAT_A50 | ISAT_A75 | ISAT_A99.9 |
|---|---|---|---|---|---|---|---|
| Name | Symbol | µmol e− m−²·s−¹ | µmol e−/µmol photons | µmol e− m−²·s−¹ | µmol photons m−²·s−¹ | µmol photons m−²·s−¹ | µmol photons m−²·s−¹ |
| AE | Control | 72.380 a ± 3.62 AB | 0.3400 a ± 0.0278 A | 0.87 a ± 0.04 ABCD | 125.0 a ± 6.3 AB | 219.5 a ± 11.0 AB | 851.8 a ± 42.6 AB |
| MD | 42.38 b ± 1.78 AB’ | 0.3367 a ± 0.0270 A’ | 0.50 a ± 0.04 AB’ | 69.3 a ± 2.9 AB’ | 121.9 a ± 5.2 AB’ | 474.2 a ± 20.2 AB’ | |
| SD | 25.02 b ± 1.33 B’’ | 0.2953 a ± 0.0557 A’’ | 0.03 b ± 0.01 BC’’ | 276.6 a ± 30.5 A’’ | 485.1 a ± 53.3 A’’ | 1881.4 a ± 206.4 A’’ | |
| Ab | Control | 55.03 a ± 2.75 BCD | 0.3324 a ± 0.0278 A | 0.72 a ± 0.04 ABCD | 91.6 a ± 4.6 AB | 161.7 a ± 8.1 AB | 629.7 a ± 31.5 AB |
| MD | 32.99 b ± 1.70 B’ | 0.3158 ab ± 0.0184 A’ | 0.14 b ± 0.01 B’ | 57.7 ab ± 2.8 B’ | 101.8 ab ± 5.0 B’ | 396.5 ab ± 19.3 B’ | |
| SD | 26.88 b ± 1.29 B’’ | 0.2947 b ± 0.0249 A’’ | 0.07 b ± 0.01 BC’’ | 36.3 b ± 2.3 B’’ | 64.4 b ± 3.9 B’’ | 251.8 b ± 15.6 B’’ | |
| C | Control | 69.24 a ± 3.46 ABC | 0.3405 a ± 0.0090 A | 0.96 a ± 0.05 ABC | 113.1 a ± 5.7 AB | 198.8 a ± 9.9 AB | 772.6 a ± 38.6 AB |
| MD | 68.21 a ± 3.75 A’ | 0.3217 a ± 0.0227 A’ | 0.49 a ± 0.03 AB’ | 117.8 a ± 6.4 A’ | 207.7 a ± 11.2 A’ | 808.9 a ± 43.6 A’ | |
| SD | 74.26 a ± 4.32 A’’ | 0.3310 a ± 0.0241 A’’ | 0.59 a ± 0.04 A’’ | 83.8 a ± 4.8 B’’ | 147.4 a ± 8.5 B’’ | 573.0 a ± 33.2 B’’ | |
| Ch | Control | 88.94 a ± 4.45 QB | 0.3340 a ± 0.0162 A | 1.25 a ± 0.06 AB | 133.4 a ± 6.7 AB | 234.5 a ± 11.7 AB | 910.9 a ± 45.5 AB |
| MD | 22.96 b ± 1.11 B’ | 0.3161 a ± 0.0298 A’ | 0.08 b ± 0.00 B’ | 45.93 b ± 2.4 B’ | 81.0 b ± 4.1 B’ | 315.5 b ± 16.0 B’ | |
| SD | 26.17 b ± 1.51 B’’ | 0.3051 a ± 0.0439 A’’ | 0.24 b ± 0.02 B’’ | 47.5 b ± 2.4 B’’ | 83.7 b ± 4.2 B’’ | 325.5 b ± 16.4 B’’ | |
| CJ | Control | 36.10 a ± 2.12 CD | 0.3209 a ± 0.0238 A | 0.34 ab ± 0.02 CDE | 63.7 a ± 4.5 AB | 113.0 a ± 8.1 AB | 441.7 a ± 31.7 AB |
| MD | 37.68 a ± 3.18 B’ | 0.3452 a ± 0.0299 A’ | 0.44 a ± 0.06 AB’ | 59.0 a ± 5.4 B’ | 103.7 a ± 9.5 B’ | 403.2 a ± 36.8 B’ | |
| SD | 19.89 a ± 1.14 BC’’ | 0.3010 a ± 0.0441 A’’ | 0.03 b ± 0.00 BC’’ | 37.3 a ± 1.8 B’’ | 65.9 a ± 3.1 B’’ | 256.6 a ± 12.2 B’’ | |
| E | Control | 94.48 a ± 4.92 A | 0.3479 a ± 0.0080 A | 1.26 a ± 0.06 A | 138.6 a ± 6.1 A | 243.3 a ± 10.7 A | 944.0 a ± 41.5 A |
| MD | 39.42 b ± 1.70 B’ | 0.3346 a ± 0.0317 A’ | 0.38 b ± 0.03 AB’ | 37.0 b ± 3.6 B’ | 64.7 b ± 6.5 B’ | 250.5 b ± 25.3 B’ | |
| SD | 2.09 c ± 0.11 C’’ | 0.2960 b ± 0.0420 A’’ | 0.17 b ± 0.01 BC’’ | 10.5 b ± 0.9 B’’ | 18.6 b ± 1.6 B’’ | 72.6 b ± 6.4 B’’ | |
| F | Control | 31.33 a ± 1.78 CD | 0.3151 a ± 0.0300 A | 0.44 ab ± 0.02 BCDE | 55.3 a ± 3.1 AB | 97.5 a ± 5.3 AB | 379.5 a ± 20.7 AB |
| MD | 58.06 b ± 3.75 AB’ | 0.3403 a ± 0.0261 A’ | 0.66 a ± 0.05 A’ | 81.8 a ± 4.3 AB’ | 143.8 a ± 7.5 AB’ | 558.8 a ± 29.0 AB’ | |
| SD | 31.45 a ± 1.63 B’’ | 0.3336 a ± 0.0291 A’’ | 0.13 b ± 0.03 BC’’ | 35.6 a ± 2.5 B’’ | 63.0 a ± 4.4 B’’ | 245.7 a ± 17.2 B’’ | |
| H | Control | 86.03 a ± 4.05 AB | 0.3276 a ± 0.0141 A | 0.34 a ± 0.03 CDE | 132.3 a ± 5.9 AB | 233.5 a ± 10.3 AB | 909.4 a ± 40.2 AB |
| MD | 51.05 ab ± 4.14 AB’ | 0.3153 a ± 0.0301 A’ | 0.24 a ± 0.03 AB’ | 89.0 a ± 7.9 AB’ | 157.0 a ± 14.0 AB’ | 611.5 a ± 54.7 AB’ | |
| SD | 24.58 b ± 1.34 BC’’ | 0.2968 a ± 0.0345 A’’ | 0.22 a ± 0.02 BC’’ | 69.2 a ± 6.1 B’’ | 122.7 a ± 10.8 B’’ | 479.1 a ± 41.8 B’’ | |
| K | Control | 36.33 a ± 1.91 CD | 0.2925 a ± 0.0471 A | 0.17 a ± 0.03 E | 40.5 a ± 3.7 AB | 71.9 a ± 6.6 AB | 280.9 a ± 26.0 AB |
| MD | 44.38 a ± 2.24 AB’ | 0.3215 a ± 0.0283 A’ | 0.02 a ± 0.01 B’ | 62.9 a ± 2.8 AB’ | 111.0 a ± 4.9 AB’ | 432.6 a ± 19.4 AB’ | |
| SD | 27.02 a ± 1.76 B’’ | 0.3248 a ± 0.0202 A’’ | 0.05 a ± 0.01 BC’’ | 45.6 a ± 3.1 B’’ | 80.5 a ± 5.4 B’’ | 313.9 a ± 21.2 B’’ | |
| M | Control | 52.98 a ± 2.65 BCD | 0.3581 a ± 0.0277 A | 0.98 a ± 0.04 ABC | 115.6 a ± 7.8 AB | 203.1 a ± 13.7 AB | 788.3 a ± 53.2 AB |
| MD | 49.11 ab ± 3.41 AB’ | 0.3198 b ± 0.0252 A’ | 0.30 b ± 0.04 AB’ | 94.1 a ± 6.2 AB’ | 165.9 a ± 10.9 AB’ | 646.0 a ± 42.5 AB’ | |
| SD | 30.13 b ± 1.68 B’’ | 0.3029 b ± 0.0212 A’’ | 0.02 b ± 0.00 BC’’ | 58.9 a ± 5.0 B’’ | 104.4 a ± 8.8 B’’ | 407.7 a ± 34.4 B’’ | |
| MS | Control | 26.06 a ± 1.22 D | 0.3052 a ± 0.0372 A | 0.11 ab ± 0.01 DE | 14.3 a ± 2.8 B | 24.6 a ± 5.1 B | 94.0 a ± 20.0 B |
| MD | 40.67 b ± 2.29 AB’ | 0.3200 a ± 0.0155 A’ | 0.23 a ± 0.01 AB’ | 70.5 b ± 4.2 AB’ | 124.4 b ± 7.5 AB’ | 484.7 b ± 29.4 AB’ | |
| SD | 25.17 a ± 1.49 B’’ | 0.3233 a ± 0.0197 A’’ | 0.07 b ± 0.01 BC’’ | 50.5 ab ± 3.2 B’’ | 89.4 ab ± 5.7 B’’ | 348.6 ab ± 22.2 B’’ | |
| P | Control | 79.79 a ± 4.10 AB | 0.3357 a ± 0.0146 A | 0.99 a ± 0.06 ABC | 123.6 a ± 5.5 AB | 217.9 a ± 9.7 AB | 848.1 a ± 37.5 AB |
| MD | 45.69 b ± 1.92 AB’ | 0.2925 a ± 0.0341 A’ | 0.23 b ± 0.01 AB’ | 81.9 a ± 3.3 AB’ | 145.6 a ± 6.8 AB’ | 569.7 a ± 26.6 AB’ | |
| SD | 8.38 c ± 0.51 C’’ | 0.2634 b ± 0.0372 B’’ | 0.09 c ± 0.02 C’’ | 27.9 b ± 2.6 B’’ | 49.7 b ± 4.7 B’’ | 195.2 b ± 18.6 B’’ | |
| S1 | Control | 71.01 a ± 4.03 AB | 0.3560 a ± 0.0146 A | 1.10 a ± 0.05 ABC | 112.1 a ± 6.6 AB | 196.7 a ± 11.6 AB | 762.8 a ± 45.1 AB |
| MD | 36.11 b ± 1.71 B’ | 0.3358 a ± 0.0244 A’ | 0.47 b ± 0.03 AB’ | 64.4 b ± 3.9 AB’ | 113.3 b ± 6.8 AB’ | 440.0 b ± 26.5 AB’ | |
| SD | 29.27 b ± 1.68 B’’ | 0.3291 a ± 0.0237 A’’ | 0.24 b ± 0.02 B’’ | 41.2 b ± 2.4 B’’ | 72.7 b ± 4.3 B’’ | 283.2 b ± 16.8 B’’ | |
| S2 | Control | 69.06 a ± 3.34 ABC | 0.3443 a ± 0.0056 A | 0.69 a ± 0.04 ABCD | 111.2 a ± 5.3 AB | 195.8 a ± 9.3 AB | 761.3 a ± 36.0 AB |
| MD | 28.51 b ± 1.59 B’ | 0.3183 b ± 0.0129 A’ | 0.22 b ± 0.01 AB’ | 49.7 b ± 2.5 B’ | 87.6 b ± 4.4 B’ | 341.0 b ± 17.0 B’ | |
| SD | 29.73 b ± 1.61 B’’ | 0.3284 ab ± 0.01659 A’’ | 0.17 b ± 0.01 BC’’ | 50.0 b ± 3.1 B’’ | 88.2 b ± 5.4 B’’ | 343.7 b ± 21.3 B’’ |
| Cultivar | Condition | Fv/Fm | ΔF/Fm′ | NPQ | qP | qN | mol photons/mol e− | mol photons/mol CO₂ | mol e−/mol CO₂ |
|---|---|---|---|---|---|---|---|---|---|
| Name | Symbol | - | - | - | - | - | µmol photons/µmol e− | µmol photons/µmol CO₂ | µmol e−/µmol CO₂ |
| AE | Control | 0.827 a ± 0.041 AB | 0.583 a ± 0.029 AB | 2.47 a ± 0.12 AB | 0.388 a ± 0.019 AB | 0.803 a ± 0.04 A | 2.94 a ± 0.03 B | 20.18 a ± 1.49 A | 6.86 a ± 0.04 A |
| MD | 0.830 a ± 0.041 A’ | 0.599 a ± 0.026 ABC’ | 2.34 a ± 0.14 ABC’ | 0.366 a ± 0.019 BCDE’ | 0.788 a ± 0.038 A’ | 2.97 a ± 0.03 B’ | 23.51 a ± 7.37 CD’ | 7.92 a ± 0.20 B’ | |
| SD | 0.832 a ± 0.000 A’’ | 0.561 a ± 0.029 AB’’ | 2.86 a ± 0.13 ABC’’ | 0.346 a ± 0.023 AB’’ | 0.830 a ± 0.040 A’’ | 3.39 a ± 0.06 B’’ | 30.66 a ± 8.17 BC’’ | 9.06 a ± 0.46 B’’ | |
| Ab | Control | 0.829 a ± 0.041 AB | 0.552 a ± 0.028 ABC | 3.10 a ± 0.15 A | 0.400 a ± 0.020 AB | 0.841 a ± 0.042 A | 3.01 a ± 0.03 B | 31.41 a ± 6.71 A | 10.44 a ± 0.19 A |
| MD | 0.769 ab ± 0.038 ABC’ | 0.533 a ± 0.028 BCD’ | 2.33 b ± 0.14 ABC’ | 0.412 a ± 0.021 ABCD’ | 0.777 a ± 0.038 A’ | 3.17 ab ± 0.02 AB’ | 23.87 a ± 7.38 CD’ | 7.54 ab ± 0.14 AB’ | |
| SD | 0.742 b ± 0.036 AB’’ | 0.505 a ± 0.023 AB’’ | 1.83 b ± 0.11 DE’’ | 0.434 a ± 0.025 A’’ | 0.774 a ± 0.037 AB’’ | 3.39 b ± 0.03 B’’ | 22.78 a ± 3.59 BC’’ | 6.71 b ± 0.09 B’’ | |
| C | Control | 0.836 a ± 0.042 A | 0.631 a ± 0.032 A | 2.42 a ± 0.12 AB | 0.352 a ± 0.018 AB | 0.790 a ± 0.040 A | 2.94 a ± 0.01 B | 23.193 a ± 3.3 A | 7.90 a ± 0.03 A |
| MD | 0.804 a ± 0.039 ABC’ | 0.603 a ± 0.032 AB’ | 1.60 a ± 0.09 CD’ | 0.313 a ± 0.017 DE’ | 0.716 a ± 0.036 A’ | 3.11 a ± 0.02 AB’ | 20.42 a ± 4.61 CD’ | 6.57 a ± 0.11 AB’ | |
| SD | 0.809 a ± 0.039 AB’’ | 0.576 a ± 0.030 A’’ | 2.06 a ± 0.11 CDE’’ | 0.373 a ± 0.023 A’’ | 0.779 a ± 0.038 AB’’ | 3.02 a ± 0.02 B’’ | 23.80 a ± 3.52 B’’ | 7.88 a ± 0.09 B’’ | |
| Ch | Control | 0.804 a ± 0.040 AB | 0.549 a ± 0.027 ABC | 2.38 a ± 0.12 AB | 0.336 a ± 0.017 B | 0.812 a ± 0.041 A | 2.99 a ± 0.02 B | 18.10 a ± 1.46 A | 6.05 a ± 0.02 A |
| MD | 0.715 a ± 0.037 C’ | 0.536 a ± 0.027 BCD’ | 1.90 ab ± 0.08 BCD’ | 0.406 a ± 0.022 ABCDE’ | 0.753 a ± 0.038 A’ | 3.16 a ± 0.03 AB’ | 31.02 a ± 6.20 A’ | 9.81 a ± 0.19 AB’ | |
| SD | 0.709 a ± 0.034 AB’’ | 0.519 a ± 0.025 AB’’ | 1.16 b ± 0.06 E’’ | 0.414 a ± 0.030 A’’ | 0.681 a ± 0.030 B’’ | 3.28 a ± 0.04 B’’ | 34.72 a ± 6.22 A’’ | 10.59 a ± 0.27 B’’ | |
| CJ | Control | 0.824 a ± 0.000 AB | 0.551 a ± 0.027 ABC | 2.80 a ± 0.16 AB | 0.392 a ± 0.020 AB | 0.831 a ± 0.042 A | 3.12 a ± 0.02 B | 25.64 a ± 3.35 A | 8.23 a ± 0.08 A |
| MD | 0.808 ab ± 0.038 ABC’ | 0.556 a ± 0.025 BCD’ | 2.39 ab ± 0.11 ABC’ | 0.516 b ± 0.027 A’ | 0.808 a ± 0.040 A’ | 2.90 ab ± 0.03 B’ | 22.27 a ± 3.25 ABC’ | 7.69 ab ± 0.10 B’ | |
| SD | 0.770 b ± 0.039 AB’’ | 0.506 a ± 0.026 AB’’ | 2.03 b ± 0.10 CD E’’ | 0.361 a ± 0.015 AB’’ | 0.809 a ± 0.040 AB’’ | 3.33 b ± 0.04 B’’ | 22.13 a ± 1.46 BCD’’ | 6.65 b ± 0.07 B’’ | |
| E | Control | 0.826 a ± 0.000 AB | 0.565 a ± 0.028 ABC | 2.51 a ± 0.13 AB | 0.393 a ± 0.020 AB | 0.821 a ± 0.041 A | 2.88 a ± 0.01 B | 19.86 a ± 2.43 A | 6.91 a ± 0.02 A |
| MD | 0.814 a ± 0.037 AB’ | 0.539 a ± 0.028 BCD’ | 3.05 ab ± 0.15 A’ | 0.464 a ± 0.025 ABC’ | 0.846 ab ± 0.042 A’ | 3.00 a ± 0.03 B’ | 16.52 a ± 1.79 D’ | 5.51 a ± 0.06 B’ | |
| SD | 0.462 a ± 0.029 AB’’ | 0.516 a ± 0.026 AB’’ | 3.44 b ± 0.17 A’’ | 0.450 a ± 0.023 A’’ | 0.869 b ± 0.043 A’’ | 3.38 a ± 0.04 B’’ | 14.78 a ± 1.52 D’’ | 4.37 a ± 0.07 B’’ | |
| F | Control | 0.820 a ± 0.000 AB | 0.539 a ± 0.027 ABC | 2.71 a ± 0.13 AB | 0.451 a ± 0.025 AB | 0.840 a ± 0.042 A | 3.17 a ± 0.03 B | 21.58 a ± 3.28 A | 6.80 a ± 0.10 A |
| MD | 0.814 a ± 0.039 AB’ | 0.586 a ± 0.030 ABCD’ | 2.19 b ± 0.11 BC’ | 0.374 a ± 0.021 CDE’ | 0.784 a ± 0.039 A’ | 2.94 a ± 0.03 B’ | 19.04 a ± 3.46 CD’ | 6.48 a ± 0.09 B’ | |
| SD | 0.803 a ± 0.038 AB’’ | 0.574 a ± 0.028 AB’’’ | 2.23 b ± 0.10 BCD’’ | 0.438 a ± 0.029 A’’ | 0.783 a ± 0.037 AB’’ | 3.00 a ± 0.03 B’’ | 16.11 a ± 2.17 D’’ | 5.37 a ± 0.06 B’’ | |
| H | Control | 0.825 a ± 0.000 AB | 0.562 a ± 0.029 ABC | 2.88 a ± 0.14 AB | 0.438 a ± 0.022 AB | 0.819 a ± 0.040 A | 3.05 a ± 0.01 B | 28.53 a ± 3.22 A | 9.35 a ± 0.05 A |
| MD | 0.825 a ± 0.000 AB’ | 0.580 a ± 0.031 ABCD’ | 2.39 a ± 0.10 ABC’ | 0.433 a ± 0.022 ABCD’ | 0.797 a ± 0.037 A’ | 3.17 a ± 0.03 AB’ | 20.93 a ± 5.70 ABCD’ | 6.60 a ± 0.17 AB’ | |
| SD | 0.825 a ± 0.000 AB’’ | 0.567 a ± 0.029 AB’’ | 2.58 a ± 0.13 ABCD’’ | 0.397 a ± 0.022 A’’ | 0.818 a ± 0.041 AB’’ | 3.37 a ± 0.03 B’’ | 35.98 a ± 10.75 AB’’ | 10.68 a ± 0.37 B’’ | |
| K | Control | 0.798 a ± 0.039 B | 0.487 a ± 0.024 C | 2.98 a ± 0.18 A | 0.411 a ± 0.015 AB | 0.865 a ± 0.044 A | 3.42 a ± 0.05 A | 65.16 a ± 24.13 A | 19.06 a ± 1.14 A |
| MD | 0.731 a ± 0.036 BC’ | 0.480 a ± 0.024 D’ | 2.10 a ± 0.11 BC’ | 0.452 a ± 0.025 ABCD’ | 0.806 a ± 0.040 A’ | 3.11 a ± 0.03 AB’ | 23.92 b ± 8.55 CD’ | 7.69 a ± 0.24 AB’ | |
| SD | 0.713 a ± 0.037 AB’’ | 0.467 a ± 0.024 B’’ | 1.94 a ± 0.11 CDE’’ | 0.436 a ± 0.021 A’’ | 0.793 a ± 0.039 AB’’ | 3.08 a ± 0.02 B’’ | 21.80 b ± 6.17 BC’’ | 7.08 a ± 0.12 B’’ | |
| M | Control | 0.803 a ± 0.040 AB | 0.569 a ± 0.028 ABC | 2.11 a ± 0.10 BC | 0.474 a ± 0.024 A | 0.787 a ± 0.039 AB | 2.79 a ± 0.03 B | 21.57 a ± 2.08 A | 7.73 a ± 0.06 A |
| MD | 0.770 a ± 0.039 ABC’ | 0.646 a ± 0.033 A’ | 0.76 b ± 0.03 D’ | 0.344 a ± 0.024 CDE’ | 0.457 b ± 0.012 B’ | 3.13 ab ± 0.03 AB’ | 22.39 a ± 3.94 BCD’ | 7.15 ab ± 0.10 AB’ | |
| SD | 0.790 a ± 0.057 AB’’ | 0.596 a ± 0.037 A’’ | 1.97 a ± 0.12 CDE’’ | 0.379 a ± 0.027 A’’ | 0.718 ab ± 0.048 AB’’ | 3.30 b ± 0.021 B’’ | 27.43 a ± 7.96 B’’ | 8.31 b ± 0.17 B’’ | |
| MS | Control | 0.792 a ± 0.039 B | 0.515 a ± 0.026 BC | 2.87 a ± 0.10 AB | 0.363 a ± 0.018 AB | 0.848 a ± 0.042 A | 3.28 a ± 0.04 B | 28.01 a ± 9.30 A | 8.55 a ± 0.34 A |
| MD | 0.804 a ± 0.000 ABC’ | 0.571 b ± 0.027 ABCD’ | 2.07 b ± 0.12 BC’ | 0.396 a ± 0.023 BCDE’ | 0.780 b ± 0.041 A’ | 3.13 a ± 0.02 AB’ | 21.54 ab ± 3.49 BCD’ | 6.89 a ± 0.05 AB’ | |
| SD | 0.804 a ± 0.000 AB’’ | 0.580 b ± 0.029 A’’ | 1.96 b ± 0.10 CDE’’ | 0.409 a ± 0.025 A’’ | 0.770 b ± 0.039 AB’’ | 3.09 a ± 0.02 B’’ | 19.30 b ± 4.13 CD’’ | 6.24 a ± 0.08 B’’ | |
| P | Control | 0.800 a ± 0.040 B | 0.612 a ± 0.031 A | 1.44 a ± 0.07 C | 0.374 a ± 0.020 AB | 0.711 a ± 0.037 B | 2.98 a ± 0.02 B | 18.80 a ± 2.64 A | 6.31 a ± 0.04 A |
| MD | 0.819 a ± 0.000 AB’ | 0.593 a ± 0.029 ABC’ | 2.11 a ± 0.13 BC’ | 0.263 ab ± 0.009 E’ | 0.759 ab ± 0.039 A’ | 3.42 a ± 0.03 A’ | 25.46 a ± 6.09 AB’ | 7.45 a ± 0.21 A’ | |
| SD | 0.601 a ± 0.020 B’’ | 0.554 a ± 0.029 AB’’ | 3.01 b ± 0.15 AB’’ | 0.213 b ± 0.008 B’’ | 0.858 b ± 0.046 A’’ | 3.80 a ± 0.03 A’’ | 27.52 a ± 8.88 AB’’ | 7.24 a ± 0.26 A’’ | |
| S1 | Control | 0.835 a ± 0.042 A | 0.525 a ± 0.024 BC | 3.16 a ± 0.16 A | 0.389 a ± 0.019 AB | 0.866 a ± 0.045 A | 2.81 a ± 0.02 B | 29.68 a ± 7.52 A | 10.57 a ± 0.11 A |
| MD | 0.772 a ± 0.035 ABC’ | 0.494 a ± 0.027 D’ | 2.79 a ± 0.20 AB’ | 0.441 a ± 0.026 ABCD’ | 0.836 a ± 0.043 A’ | 2.98 a ± 0.02 B’ | 21.04 ab ± 3.59 BCD’ | 7.06 a ± 0.09 B’ | |
| SD | 0.794 a ± 0.040 AB’’ | 0.521 a ± 0.026 AB’’ | 2.59 a ± 0.13 ABCD’’ | 0.478 a ± 0.024 A’’ | 0.825 a ± 0.041 A’’ | 3.04 a ± 0.02 B’’ | 21.97 b ± 3.75 BCD’’ | 7.24 a ± 0.09 B’’ | |
| S2 | Control | 0.820 a ± 0.000 AB | 0.549 a ± 0.028 ABC | 2.74 a ± 0.14 AB | 0.411 a ± 0.021 AB | 0.833 a ± 0.041 A | 2.90 a ± 0.01 B | 22.12 a ± 1.62 A | 7.62 a ± 0.01 A |
| MD | 0.764 a ± 0.038 ABC’ | 0.513 a ± 0.027 CD’ | 2.05 a ± 0.12 BC’ | 0.492 a ± 0.019 AB’ | 0.817 a ± 0.045 A’ | 3.14 b ± 0.01 AB’ | 22.13 a ± 2.81 BCD’ | 7.043 b ± 0.04 AB’ | |
| SD | 0.778 a ± 0.038 AB’’ | 0.522 a ± 0.025 AB’’ | 2.32 a ± 0.14 ABCD’’ | 0.470 a ± 0.0230 A’’ | 0.808 a ± 0.041 AB’’ | 3.05 ab ± 0.02 B’’ | 22.34 a ± 3.41 BCD’’ | 7.34 ab ± 0.05 B’’ |
| Arid Areas 200–400 mm/Year | Moderately Tolerant 350–500 mm/Year | Moderately Sensitive 400–650 mm/Year | Non-Arid Areas 500–800 mm/Year | |
|---|---|---|---|---|
| Low irradiances | - | F | MS | CJ |
| Medium irradiances | AO | S1 | K | |
| High irradiances | M, S2, C | CH, P | AE, H | E |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martos de la Fuente, G.C.; Viñegla, B.; Illana Rico, E.; Fernández Ocaña, A.M. Study of the Photosynthesis Response during the Gradual Lack of Water for 14 Olea europaea L. subsp europaea Cultivars and Their Adaptation to Climate Change. Plants 2023, 12, 4136. https://doi.org/10.3390/plants12244136
Martos de la Fuente GC, Viñegla B, Illana Rico E, Fernández Ocaña AM. Study of the Photosynthesis Response during the Gradual Lack of Water for 14 Olea europaea L. subsp europaea Cultivars and Their Adaptation to Climate Change. Plants. 2023; 12(24):4136. https://doi.org/10.3390/plants12244136
Chicago/Turabian StyleMartos de la Fuente, Genoveva Carmen, Benjamín Viñegla, Elena Illana Rico, and Ana Maria Fernández Ocaña. 2023. "Study of the Photosynthesis Response during the Gradual Lack of Water for 14 Olea europaea L. subsp europaea Cultivars and Their Adaptation to Climate Change" Plants 12, no. 24: 4136. https://doi.org/10.3390/plants12244136
APA StyleMartos de la Fuente, G. C., Viñegla, B., Illana Rico, E., & Fernández Ocaña, A. M. (2023). Study of the Photosynthesis Response during the Gradual Lack of Water for 14 Olea europaea L. subsp europaea Cultivars and Their Adaptation to Climate Change. Plants, 12(24), 4136. https://doi.org/10.3390/plants12244136