
PacBio Full-Length Transcriptome Sequencing Reveals the Mechanism of Salt Stress Response in Sonneratia apetala


Abstract
:1. Introduction
2. Results
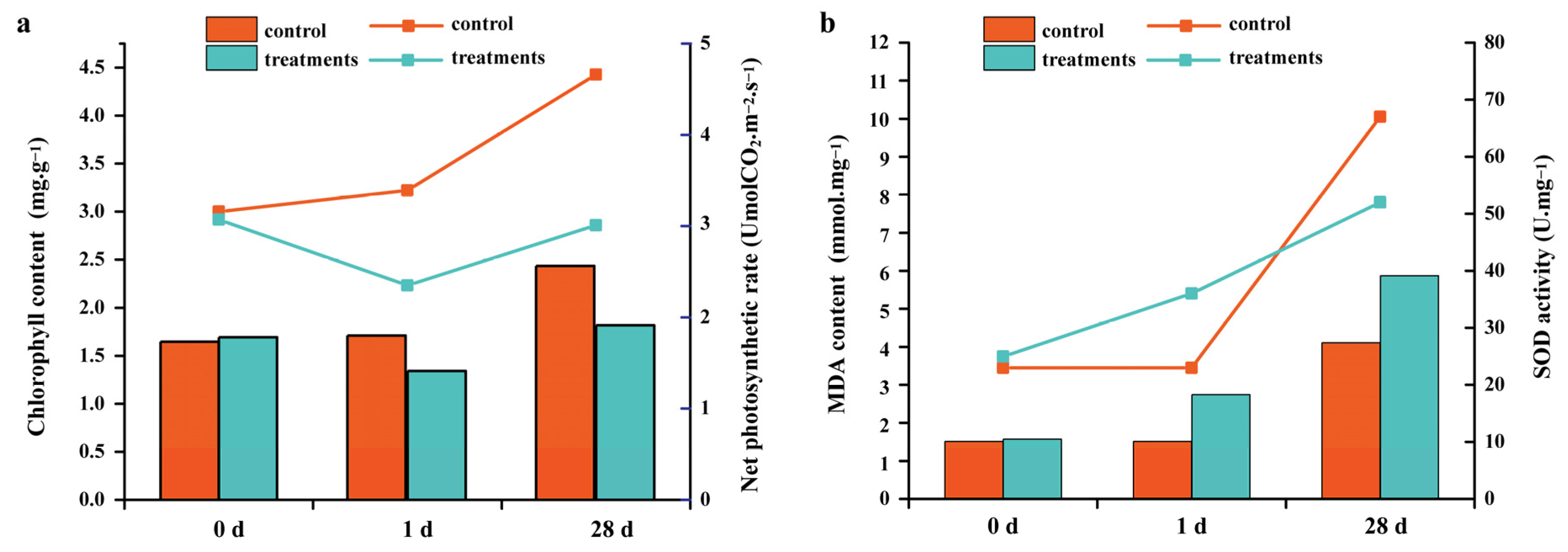
2.1. Identification of Growth and Physiological Indexes after Salt Stress
2.2. An Overview of Analysis of Sequencing Data
2.3. Analysis of Differentially Expressed Genes (DEGs) and Co-DEGs Cluster
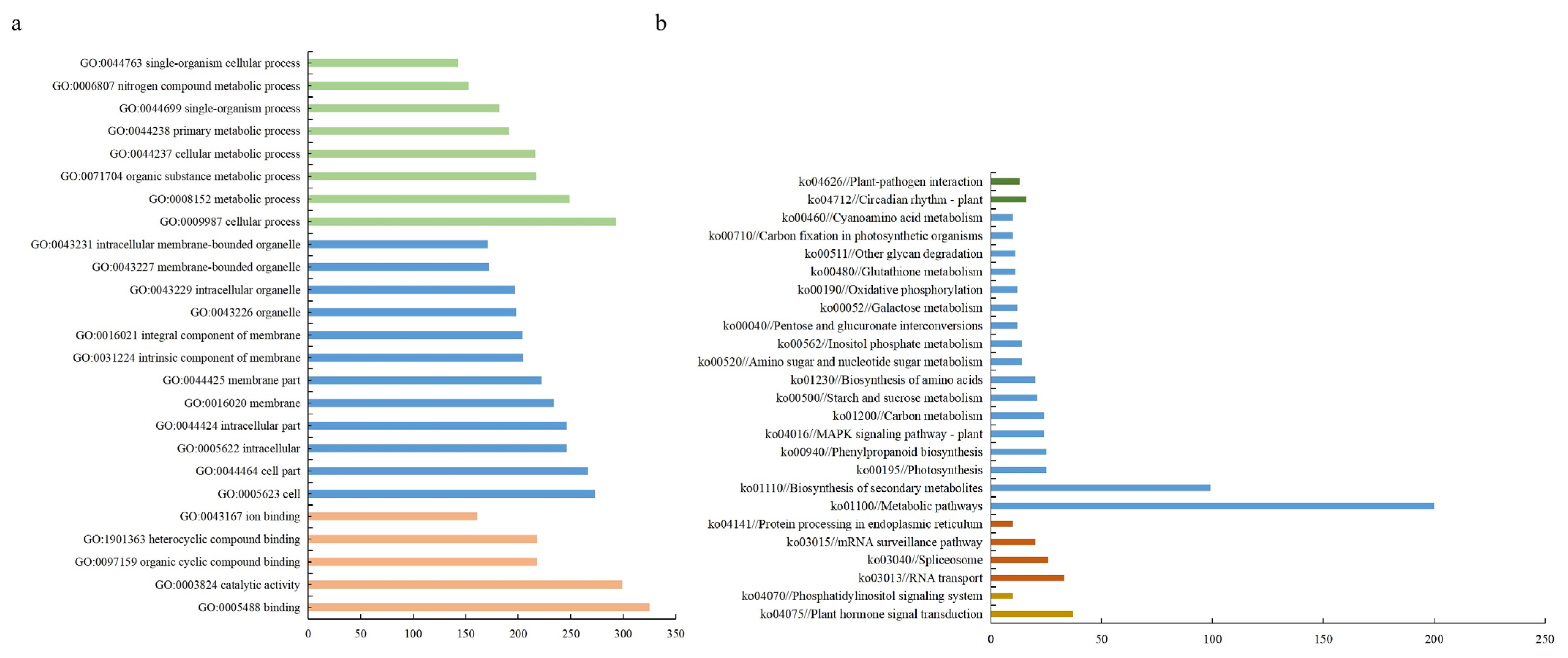
2.4. Functional Categorization of Deferentially Expressed Genes
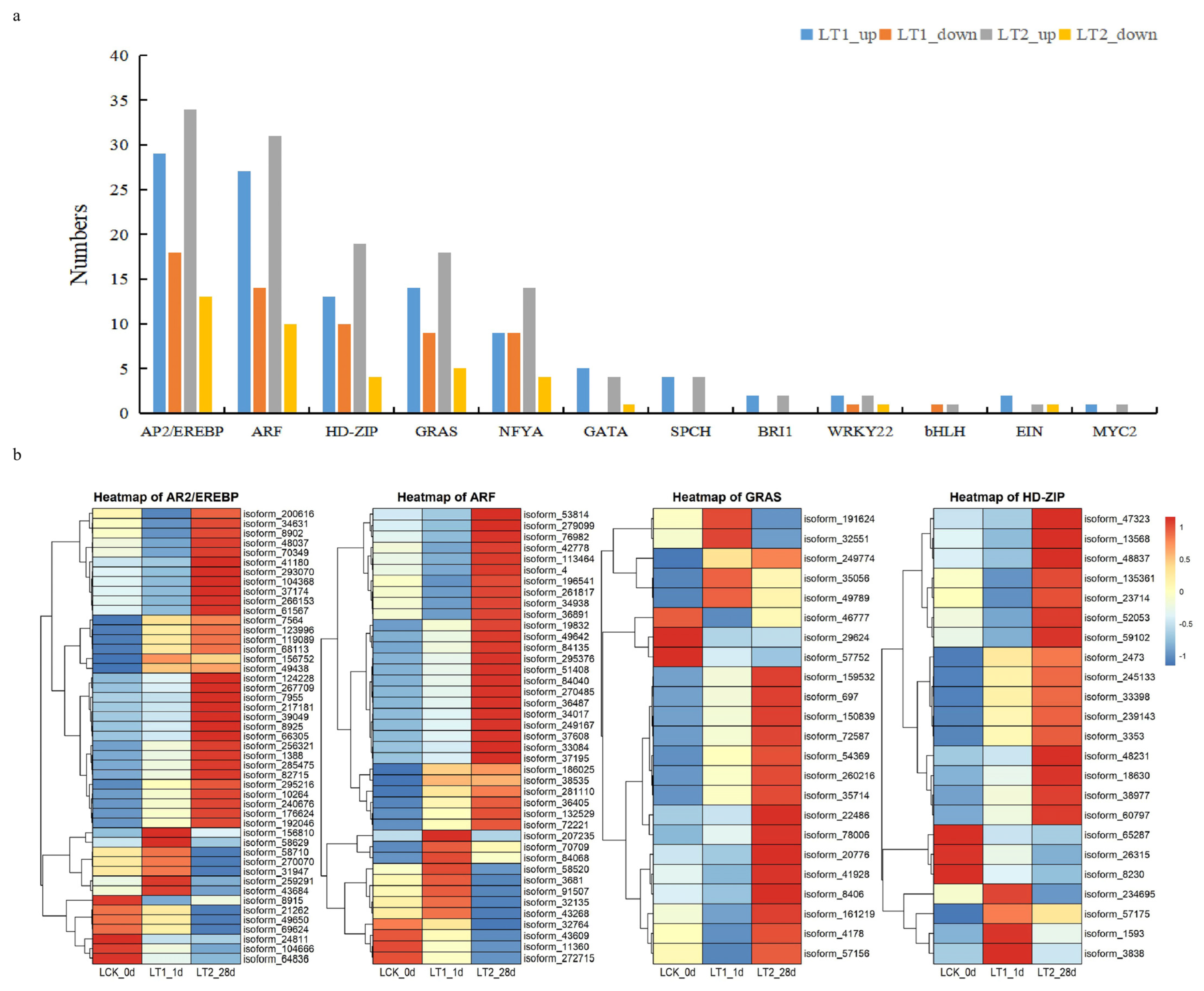
2.4.1. Annotation of Transcription Factors in Co-Expressed DEGS
2.4.2. Co-Expressed DEGs Involved in Signal Transduction under Salt Stress
2.4.3. Co-Expressed DEGs Involved in Ionic Balance and Water Transport
2.4.4. Co-Expressed DEGs Involved in Production and Scavenging of Reactive Oxygen Species
2.4.5. Co-Expressed DEGs Involved in Hormone Response
2.4.6. Co-Expressed DEGs Involved in Cell Wall Synthesis or Modification
3. Discussion
4. Materials and Methods
4.1. Experimental Materials
4.2. RNA Preparation, Library Construction and Sequencing
4.3. RNA-Seq and Differentially Expressed Genes Identification
4.4. GO and KEGG Enrichment Analysis of Co-Expressed DEGs
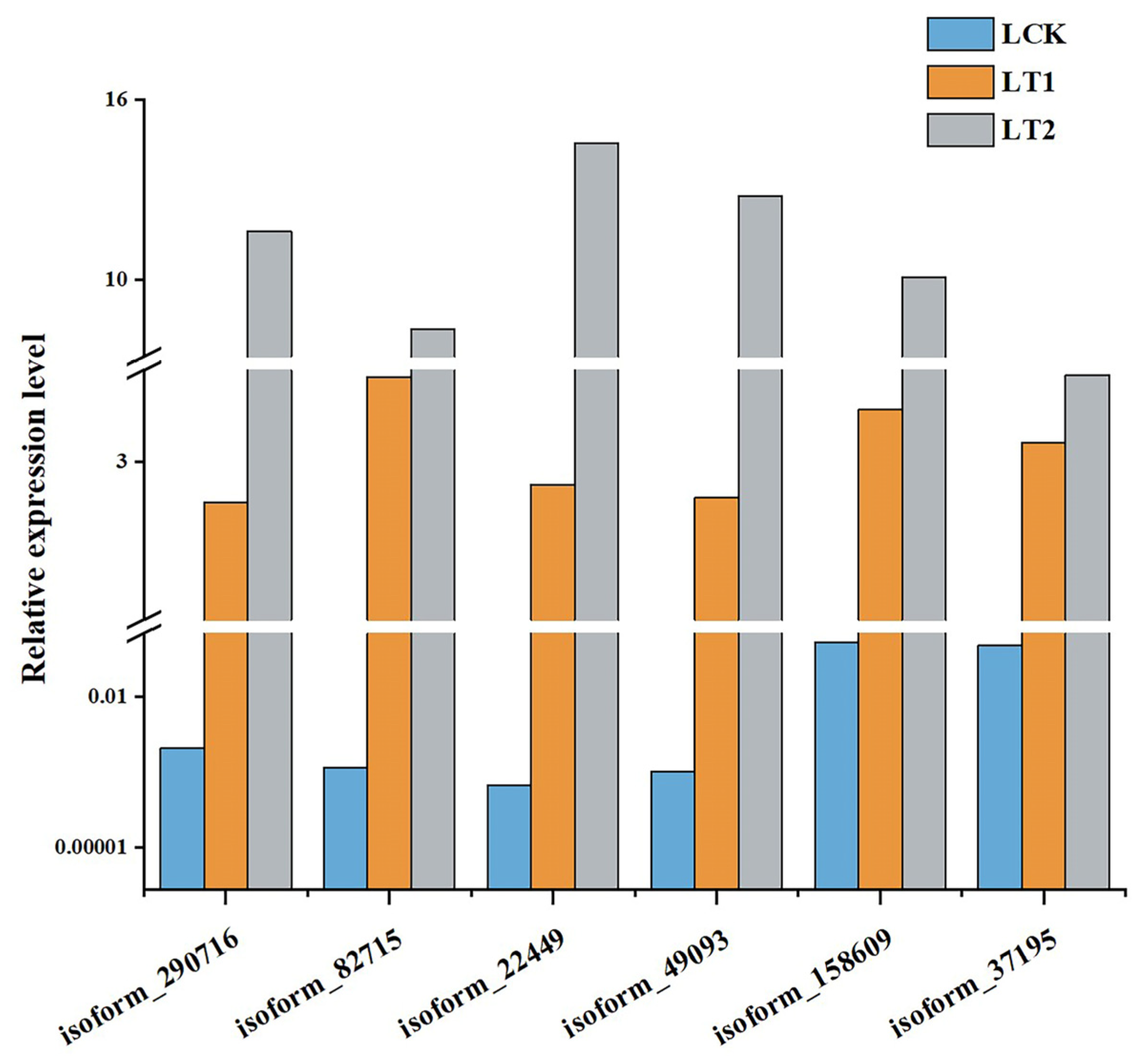
4.5. Quantification of Transcript Levels via Quantitative Real-Time PCR (qRT-PCR)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant salt-tolerance mechanism: A review. Biochem. Biophys. Res. Commun. 2018, 495, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Møller, I.S.; Tester, M. Salinity tolerance of Arabidopsis: A good model for cereals? Trends Plant Sci. 2007, 12, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Mishra, S.; Heckathorn, S.A.; Frantz, J.M.; Krause, C. Proteomic analysis of leaves in response to acute boron deficiency and toxicity reveals effects on photosynthesis, carbohydrate metabolism, and protein synthesis. J. Plant Physiol. 2014, 171, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Zhao, L.; Zhang, K.; Lu, H.; Bhanbhro, N.; Yang, C. Comparative genomics and transcriptomics of the extreme halophyte Puccinellia tenuiflora provides insights into salinity tolerance differentiation between Halophytes and Glycophytes. Front. Plant Sci. 2021, 12, 649001. [Google Scholar] [CrossRef]
- Flowers, T.J.; Galal, H.K.; Bromham, L. Evolution of halophytes: Multiple origins of salt tolerance in land plants. Funct. Plant Biol. 2010, 37, 604–612. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, Q.; Gao, Z.; Wang, Y.; Liu, Y.; Ma, Z.; Chen, Y.; Zhang, Y.; Yan, F.; Li, J. Analysis of Phytohormone Signal Transduction in under Salt Stress. Int. J. Mol. Sci. 2021, 22, 7313. [Google Scholar] [CrossRef]
- Madhu; Kaur, A.; Tyagi, S.; Shumayla; Singh, K.; Upadhyay, S.K. Exploration of glutathione reductase for abiotic stress response in bread wheat (Triticum aestivum L.). Plant Cell Rep. 2022, 41, 639–654. [Google Scholar] [CrossRef]
- Yoshida, T.; Mogami, J.; Yamaguchi-Shinozaki, K. ABA-dependent and ABA-independent signaling in response to osmotic stress in plants. Curr. Opin. Plant Biol. 2014, 21, 133–139. [Google Scholar] [CrossRef]
- Qi, N.; Wang, N.; Hou, X.; Li, Y.; Liao, W. Involvement of calcium and calmodulin in NO-alleviated salt stress in tomato seedlings. Plants 2022, 11, 2479. [Google Scholar] [CrossRef]
- Ni, L.; Wang, Z.; Liu, X.; Wu, S.; Hua, J.; Yin, Y.; Li, H.; Gu, C. Transcriptome Analysis of Salt Stress in Hibiscus hamabo Sieb. et Zucc Based on Pacbio Full-Length Transcriptome Sequencing. Int. J. Mol. Sci. 2021, 23, 138. [Google Scholar] [CrossRef]
- Forlani, G.; Bertazzini, M.; Cagnano, G. Stress-driven increase in proline levels, and not proline levels themselves, correlates with the ability to withstand excess salt in a group of 17 Italian rice genotypes. Plant Biol. 2019, 21, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Naidoo, G. The mangroves of Africa: A review. Mar. Pollut. Bull. 2023, 190, 114859. [Google Scholar] [PubMed]
- Banzai, T.; Hershkovits, G.; Katcoff, D.J.; Hanagata, N.; Dubinsky, Z.; Karube, I. Identification and characterization of mRNA transcripts differentially expressed in response to high salinity by means of differential display in the mangrove, Bruguiera gymnorrhiza. Plant Sci. 2002, 162, 499–505. [Google Scholar] [CrossRef]
- Yamada, A.; Saitoh, T.; Mimura, T.; Ozeki, Y. Expression of mangrove Allene Oxide Cyclase enhances salt tolerance in escherichia coli, yeast, and tobacco Cells. Plant Cell Physiol. 2002, 43, 903–910. [Google Scholar] [CrossRef] [PubMed]
- Parida, A.K.; Das, A.B.; Mohanty, P. Defense potentials to NaCl in a mangrove, Bruguiera parviflora: Differential changes of isoforms of some antioxidative enzymes. J. Plant Physiol. 2004, 161, 531–542. [Google Scholar]
- Shen, Z.; Qin, Y.; Luo, M.; Li, Z.; Ma, D.; Wang, W.; Zheng, H. Proteome analysis reveals a systematic response of cold-acclimated seedlings of an exotic mangrove plant Sonneratia apetala to chilling stress. J. Proteomics. 2021, 248, 104349. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, J.; Yang, Z.; Chen, E.; Zhang, C.; Zhang, X.; Li, F. Genome-wide analysis of GRAS transcription factor gene family in Gossypium hirsutum L. BMC Genom. 2018, 19, 1–12. [Google Scholar] [CrossRef]
- Ernst, J.; Bar-Joseph, Z. STEM: A tool for the analysis of short time series gene expression data. BMC Bioinform. 2006, 7, 191. [Google Scholar] [CrossRef]
- Yang, Y.; Guo, Y. Unraveling salt stress signaling in plants. J. Integr. Plant. Biol. 2018, 60, 796–804. [Google Scholar] [CrossRef]
- Peng, Z.; He, S.; Gong, W.; Sun, J.; Pan, Z.; Xu, F.; Lu, Y.; Du, X. Comprehensive analysis of differentially expressed genes and transcriptional regulation induced by salt stress in two contrasting cotton genotypes. BMC Genom. 2014, 15, 760. [Google Scholar] [CrossRef]
- Lennicke, C.; Cochemé, H.M. Redox metabolism: ROS as specific molecular regulators of cell signaling and function. Mol. Cell 2021, 81, 3691–3707. [Google Scholar] [CrossRef] [PubMed]
- Yousefirad, S.; Soltanloo, H.; Ramezanpour, S.S.; Nezhad, K.Z.; Shariati, V. The RNA-seq transcriptomic analysis reveals genes mediating salt tolerance through rapid triggering of ion transporters in a mutant barley. PLoS ONE 2020, 15, e0229513. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Ding, Z.; Zhou, X.; Wang, Y.; Huang, F.; Sun, J.; Chen, J.; Han, W. Integrated full-length transcriptome and microRNA sequencing approaches provide insights into salt tolerance in mangrove (Sonneratia apetala Buch.-Ham.). Front. Genet. 2022, 13, 932832. [Google Scholar] [CrossRef]
- Okamuro, J.K.; Caster, B.; Villarroel, R.; Van Montagu, M.; Jofuku, K.D. The AP2 domain of APETALA2 defines a large new family of DNA binding proteins in Arabidopsis. Proc. Natl. Acad. Sci. USA 1997, 94, 7076–7081. [Google Scholar] [CrossRef]
- Dietz, K.; Vogel, M.O.; Viehhauser, A. AP2/EREBP transcription factors are part of gene regulatory networks and integrate metabolic, hormonal and environmental signals in stress acclimation and retrograde signalling. Protoplasma 2010, 245, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Frosi, G.; Ferreira-Neto, J.R.C.; Bezerra-Neto, J.P.; Pandolfi, V.; da Silva, M.D.; de Lima Morais, D.A.; Benko-Iseppon, A.M.; Santos, M.G. Transcriptome of Cenostigma pyramidale roots, a woody legume, under different salt stress times. Physiol. Plant. 2021, 173, 1463–1480. [Google Scholar] [CrossRef]
- van Zelm, E.; Zhang, Y.; Testerink, C. Salt Tolerance Mechanisms of Plants. Annu. Rev. Plant Biol. 2020, 71, 403–433. [Google Scholar] [CrossRef]
- Park, H.J.; Kim, W.Y.; Yun, D.J. A New Insight of Salt Stress Signaling in Plant. Mol. Cells. 2016, 39, 447–459. [Google Scholar] [CrossRef]
- Tyagi, S.; Sharma, Y.; Sharma, A.; Pandey, A.; Singh, K.; Upadhyay, S.K. Expression of TaNCL2-A ameliorates cadmium toxicity by increasing calcium and enzymatic antioxidants activities in Arabidopsis. Chemosphere 2023, 329, 138636. [Google Scholar]
- Steinhorst, L.; He, G.; Moore, L.K.; Schültke, S.; Schmitz-Thom, I.; Cao, Y.; Hashimoto, K.; Andrés, Z.; Piepenburg, K.; Ragel, P.; et al. A Ca2+-sensor switch for tolerance to elevated salt stress in Arabidopsis. Dev. Cell. 2022, 57, 2081–2094. [Google Scholar] [CrossRef]
- Bacha, H.; Rodenas, R.; Lopez-Gomez, E.; Garcia-Legaz, M.F.; Nieves-Cordones, M.; Rivero, R.M.; Martinez, V.; Botella, M.A.; Rubio, F. High Ca2+ reverts the repression of high-affinity K+ uptake produced by Na+ in Solanum lycopersycum L. (var. microtom) plants. J. Plant Physiol. 2015, 180, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Perochon, A.; Aldon, D.; Galaud, J.P.; Ranty, B. Calmodulin and calmodulin-like proteins in plant calcium signaling. Biochimie 2011, 93, 2048–2053. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Chen, N.; Song, L.; Wang, D.; Cai, H.; Yao, L.; Li, X.; Guo, C. Alfalfa (Medicago sativa L.) MsCML46 gene encoding calmodulin-like protein confers tolerance to abiotic stress in tobacco. Plant Cell Rep. 2021, 40, 1907–1922. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Aharon, G.S.; Sottosanto, J.B.; Blumwald, E. Vacuolar Na+/H+ antiporter cation selectivity is regulated by calmodulin from within the vacuole in a Ca2+ and pH-dependent manner. Proc. Natl. Acad. Sci. USA 2005, 102, 16107–16112. [Google Scholar] [CrossRef]
- Acharya, B.R.; Zhao, C.; Reyes, L.A.R.; Ferreira, J.F.S.; Sandhu, D. Understanding the salt overly sensitive pathway in Prunus: Identification and characterization of NHX, CIPK, and CBL genes. Plant Genome 2023, e20371. [Google Scholar] [CrossRef]
- Sanyal, S.K.; Pandey, A.; Pandey, G.K. The CBL-CIPK signaling module in plants: A mechanistic perspective. Physiol. Plant. 2015, 155, 89–108. [Google Scholar] [CrossRef]
- McAinsh, M.R.; Pittman, J.K. Shaping the calcium signature. New Phytol. 2009, 181, 275–294. [Google Scholar] [CrossRef]
- Costa, A.; Resentini, F.; Buratti, S.; Bonza, M.C. Plant Ca2+-ATPases: From biochemistry to signalling. BBA-Mol. Cell. Res. 2023, 1870, 119508. [Google Scholar] [CrossRef]
- Lee, H.J.; Seo, P.J. Ca2+ talyzing initial responses to environmental stresses. Trends Plant Sci. 2021, 26, 849–870. [Google Scholar] [CrossRef]
- Isom, D.G.; Sridharan, V.; Baker, R.; Clement, S.T.; Smalley, D.M.; Dohlman, H.G. Protons as second messenger regulators of G protein signaling. Mol. Cell. 2013, 51, 531–538. [Google Scholar] [CrossRef]
- Liu, Y.; Dai, X.; Zhao, L.; Huo, K.; Jin, P.; Zhao, D.; Zhou, Z.; Tang, J.; Xiao, S.; Cao, Q. RNA-seq reveals the salt tolerance of Ipomoea pes-caprae, a wild relative of sweet potato. J. Plant Physiol. 2020, 255, 153276. [Google Scholar] [CrossRef]
- Xu, H.; Zhang, Q.; Cui, W.; Zhang, X.; Liu, W.; Zhang, L.; Islam, M.N.; Baek, K.; Wang, Y. AbSte7 a MAPKK gene of alternaria brassicicola, is involved in conidiation, salt/oxidative stress, and pathogenicity. J. Microbiol. Biotechn. 2016, 26, 1311–1319. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.D.; Cho, Y.H.; Tena, G.; Xiong, Y.; Sheen, J. Dual control of nuclear EIN3 by bifurcate MAPK cascades in CH signalling. Nature 2008, 451, 789–795. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Li, B.; Li, S.; Liang, Y.; Wu, X.; Ma, M.; Wang, J.; Gao, J.; Cai, Y.; Zhang, Y.; et al. Mitogen-activated protein kinase cascade MKK7-MPK6 plays important roles in plant development and regulates shoot branching by phosphorylating PIN1 in Arabidopsis. PLoS Biol. 2016, 14, e1002550. [Google Scholar] [CrossRef] [PubMed]
- Son, S.; Kim, J.; An, C.S.; Kim, S.L.; Lee, H.; Im, J.H. GmMPK6 positively regulates salt tolerance through induction of GmRbohI1 in soybean. Antioxidants 2023, 12, 601. [Google Scholar] [CrossRef]
- Tang, H.; Yu, Q.; Li, Z.; Liu, F.; Su, W.; Zhang, C.; Ling, H.; Luo, J.; Su, Y.; Que, Y. A PIP-mediated osmotic stress signaling cascade plays a positive role in the salt tolerance of sugarcane. BMC Plant Biol. 2021, 21, 589. [Google Scholar] [CrossRef]
- Kumar, R.S.; Ji, G.; Guo, H.; Zhao, L.; Zheng, B. Over-expression of a grafting-responsive gene from hickory increases abiotic stress tolerance in Arabidopsis. Plant Cell Rep. 2018, 37, 541–552. [Google Scholar] [CrossRef]
- Vaziriyeganeh, M.; Khan, S.; Zwiazek, J.J. Analysis of aquaporins in northern grasses reveal functional importance of Puccinellia nuttalliana PIP2;2 in salt tolerance. Plant Cell Environ. 2023, 46, 2159–2173. [Google Scholar] [CrossRef]
- Dai, F.; Li, A.; Rao, S.; Chen, J. Potassium transporter LrKUP8 is essential for K+ preservation in Lycium ruthenicum, a salt-resistant desert shrub. Genes 2019, 10, 600. [Google Scholar] [CrossRef]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The Role of Na+ and K+ Transporters in Salt Stress Adaptation in Glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef]
- Sharma, H.; Taneja, M.; Upadhyay, S.K. Identification, characterization and expression profiling of cation-proton antiporter superfamily in Triticum aestivum L. and functional analysis of TaNHX4-B. Genomics 2020, 112, 356–370. [Google Scholar] [CrossRef]
- Tyagi, S.; Sharma, Y.; Sharma, A.; Pandey, A.; Singh, K.; Upadhyay, S.K. TaGPX1-D overexpression provides salinity and osmotic stress tolerance in Arabidopsis. Chemosphere 2023, 337, 111881. [Google Scholar] [CrossRef] [PubMed]
- Brosché, M.; Overmyer, K.; Wrzaczek, M.; Kangasjärvi, J.; Kangasjärvi, S. Stress Signaling III: Reactive Oxygen Species (ROS); Pareek, A., Sopory, S., Bohnert, H., Eds.; Abiotic Stress Adaptation in Plants; Springer: Dordrecht, The Netherlands, 2009; pp. 91–102. [Google Scholar]
- Chapman, J.M.; Muhlemann, J.K.; Gayomba, S.R.; Muday, G.K. RBOH-dependent ROS synthesis and ROS scavenging by plant specialized metabolites to modulate plant development and stress Responses. Chem. Res. Toxicol. 2019, 32, 370–396. [Google Scholar] [CrossRef] [PubMed]
- Soliman, A.; Adam, L.R.; Rehal, P.K.; Daayf, F. Overexpression of Solanum tuberosum respiratory burst oxidase homolog A (StRbohA) promotes potato tolerance to phytophthora infestans. Phytopathology 2021, 111, 1410–1419. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Wang, W.; Bittner, F.; Schmidt, N.; Berkey, R.; Zhang, L.; King, H.; Zhang, Y.; Feng, J.; Wen, Y.; et al. Dual and opposing roles of xanthine dehydrogenase in defense-associated reactive oxygen species metabolism in Arabidopsis. Plant Cell 2016, 28, 1108–1126. [Google Scholar] [CrossRef] [PubMed]
- Gournas, C.; Papageorgiou, I.; Diallinas, G. The nucleobase–ascorbate transporter (NAT) family: Genomics, evolution, structure–function relationships and physiological role. Mol. Biosyst. 2008, 4, 404–416. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yuan, J.; Qin, L.; Shi, W.; Xia, G.; Liu, S. TaCYP81D5, one member in a wheat cytochrome P450 gene cluster, confers salinity tolerance via reactive oxygen species scavenging. Plant Biotechnol. J. 2020, 18, 791–804. [Google Scholar] [CrossRef]
- Meng, H.; Zhao, J.; Yang, Y.; Diao, K.; Zheng, G.; Li, T.; Dai, X.; Li, J. PeGSTU58, a Glutathione S-Transferase from Populus euphratica, enhances salt and drought stress tolerance in transgenic Arabidopsis. Int. J. Mol. Sci. 2023, 24, 9354. [Google Scholar] [CrossRef]
- Verma, S.; Negi, N.P.; Pareek, S.; Mudgal, G.; Kumar, D. Auxin response factors in plant adaptation to drought and salinity stress. Physiol. Plant. 2022, 174, e13714. [Google Scholar] [CrossRef]
- Shi, J.; Wang, N.; Zhou, H.; Xu, Q.; Yan, G. The role of gibberellin synthase gene GhGA2ox1 in upland cotton (Gossypium hirsutum L.) responses to drought and salt stress. Biotechnol. Appl. Biochem. 2019, 66, 298–308. [Google Scholar] [CrossRef]
- Yao, X.; Meng, L.; Zhao, W.; Mao, G. Changes in the morphology traits, anatomical structure of the leaves and transcriptome in Lycium barbarum L. under salt stress. Front. Plant Sci. 2023, 14, 1090366. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Li, Z.; Wang, Y.; Wang, J.; Xiao, M.; Liu, H.; Quan, R.; Zhang, H.; Huang, R.; Zhu, L.; et al. Cellulose synthase-like protein OsCSLD4 plays an important role in the response of rice to salt stress by mediating abscisic acid biosynthesis to regulate osmotic stress tolerance. Plant Biotechnol. J. 2022, 20, 468–484. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Cheng, X.; Li, X.; Deng, X.; Dong, X.; Wang, S.; Pu, X. Effects of silicon application on leaf structure and physiological characteristics of Glycyrrhiza uralensis Fisch. and Glycyrrhiza inflata Bat. under salt treatment. BMC Plant Biol. 2022, 22, 390. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Chen, Y.; Xiao, X.; Qu, Y.; Li, P.; Lu, Q.; Huang, J. Genome-wide analysis of the CalS gene family in cotton reveals their potential roles in fiber development and responses to stress. PeerJ 2021, 9, e12557. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Sun, L.; Dong, X.; Lu, S.; Tian, W.; Liu, J. Cellulose synthesis genes CESA6 and CSI1 are important for salt stress tolerance in Arabidopsis. J. Integr. Plant Biol. 2016, 58, 623–626. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; He, H.; Fang, L.; Zhang, A. Pectin methylesterase31 positively regulates salt stress tolerance in Arabidopsis. Biochem. Biophys. Res. Commun. 2018, 496, 497–501. [Google Scholar] [CrossRef]
- Ni, L.; Wang, Q.; Chen, C.; Wang, S.; Shen, T.; Jiang, J.; Cui, Z.; Li, K.; Yang, Q.; Jiang, M. OsDMI3-mediated OsUXS3 phosphorylation improves oxidative stress tolerance by modulating OsCATB protein abundance in rice. J. Integr. Plant Biol. 2022, 64, 1087–1101. [Google Scholar] [CrossRef]
- Li, W.; Xu, R.; Yan, X.; Liang, D.; Zhang, L.; Qin, X.; Caiyin, Q.; Zhao, G.; Xiao, W.; Hu, Z.; et al. De novo leaf and root transcriptome analysis to explore biosynthetic pathway of Celangulin V in Celastrus angulatus maxim. BMC Genom. 2019, 20, 7. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 1–16. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- BIG Data Center Members. Database resources of the BIG Data Center in 2018. Nucleic Acids Res. 2018, 46, D14–D20. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Increment of Root (cm) | Increment of Stem Length (cm) | Increment of Stem Base Diameter (cm) | Increment of Leaf Number (Piece) | Increment of Leaf Length (cm) | Increment of Leaf Width (cm) | Increment of Seedlings Fresh Weight (g) |
|---|---|---|---|---|---|---|---|
| Control | 8.01 ± 0.22 a | 1.42 ± 0.16 a | 0.45 ± 0.01 a | 2.43 ± 0.04 a | 0.68 ± 0.04 a | 0.18 ± 0.02 a | 0.26 ± 0.05 a |
| Salinity treatments | 7.33 ± 0.30 b | 1.13 ± 0.02 b | 0.37 ± 0.08 ab | 2.06 ± 0.11 b | 0.51 ± 0.03 b | 0.15 ± 0.06 ab | 0.18 ± 0.03 b |
| Sample | Total Raw Reads (M) | Total Clean Reads (M) | Total Clean Bases (Gb) | Clean Reads Q20 (%) | Clean Reads Q30 (%) | Clean Reads Ratio (%) | Total Mapping (%) |
|---|---|---|---|---|---|---|---|
| LCK_a | 43.78 | 43.16 | 6.47 | 97.13 | 92.33 | 98.58 | 89.77 |
| LCK_b | 43.78 | 43.27 | 6.49 | 97.21 | 92.65 | 98.84 | 89.37 |
| LCK_c | 43.78 | 43.21 | 6.48 | 97.26 | 92.38 | 98.7 | 90.11 |
| LT1_a | 43.78 | 43.02 | 6.46 | 96.86 | 92.31 | 98.26 | 88.43 |
| LT1_b | 43.78 | 43.15 | 6.47 | 97.29 | 92.46 | 98.56 | 88 |
| LT1_c | 43.78 | 43.26 | 6.49 | 97.07 | 92.29 | 98.81 | 88.37 |
| LT2_a | 43.78 | 43.11 | 6.47 | 97.23 | 92.46 | 98.47 | 88.96 |
| LT2_b | 43.78 | 43.06 | 6.46 | 97.07 | 92.39 | 98.36 | 88.62 |
| LT2_c | 43.82 | 43.21 | 6.48 | 96.96 | 92.16 | 98.61 | 89.07 |
| Sum | 394.06 | 388.45 | 58.27 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, B.; Liu, T.; Yang, Z.; Yang, S.; Chen, J. PacBio Full-Length Transcriptome Sequencing Reveals the Mechanism of Salt Stress Response in Sonneratia apetala. Plants 2023, 12, 3849. https://doi.org/10.3390/plants12223849
Chen B, Liu T, Yang Z, Yang S, Chen J. PacBio Full-Length Transcriptome Sequencing Reveals the Mechanism of Salt Stress Response in Sonneratia apetala. Plants. 2023; 12(22):3849. https://doi.org/10.3390/plants12223849
Chicago/Turabian StyleChen, Beibei, Tingting Liu, Zhuanying Yang, Shaoxia Yang, and Jinhui Chen. 2023. "PacBio Full-Length Transcriptome Sequencing Reveals the Mechanism of Salt Stress Response in Sonneratia apetala" Plants 12, no. 22: 3849. https://doi.org/10.3390/plants12223849
APA StyleChen, B., Liu, T., Yang, Z., Yang, S., & Chen, J. (2023). PacBio Full-Length Transcriptome Sequencing Reveals the Mechanism of Salt Stress Response in Sonneratia apetala. Plants, 12(22), 3849. https://doi.org/10.3390/plants12223849