1. Introduction
Water deficit is considered one of the principal environmental stresses for crops, and many regions of the world already face substantial water scarcity, with a consequent reduction in agricultural production [
1]. Historically, most rice breeding efforts have been devoted to increasing yield potential, while currently, due to the problem of climate change, rice breeders are focusing on developing cultivars that are more resistant to abiotic stresses [
2]. The initial plant physiological responses to water deficit include a decrease in leaf area, stimulation of leaf abscission, directional root growth towards moister soil regions, and induction of stomatal closure. In this context, the abscisic acid hormone (ABA) prevents excessive water loss during transpiration [
3]. Stomata are essential plant gas exchange structures that regulate photosynthesis, respiration, transpiration, and temperature [
4]. Furthermore, stomata are also the water-regulating organs of plants and are essential for water conservation, a decisive factor for the survival of plants under conditions of water deficit [
5]. Water deficit can also induce a physiological imbalance due to an excess of reactive oxygen species (ROS), resulting in oxidative stress. In response, plants produce a variety of antioxidant enzymes, such as superoxide dismutase (SOD), peroxidase (POD), and antioxidant enzymes [
6], to maintain their metabolic stability under situations of environmental stress.
Plant genes associated with water deficit tolerance encode proteins for cellular adaptation, including chaperones, transcription factors, or enzymes involved in signal transduction, such as protein kinases [
7]. Numerous studies have been conducted in rice to elucidate the roles of different genes in water stress tolerance. Specific genes that have led to enhanced water deficit tolerance in rice include
OsEPF1 [
8],
EDT1 [
9],
OsRab7 [
10],
OsMYB6 [
11],
OsTF1L [
12],
OsbZIP42 [
13], and
OsNAR2.1 [
14]. Interestingly, several of these mentioned genes have significantly improved rice grain yield and plant biomass accumulation. However, the complexity of responses to water deficit suggests the involvement of a substantial number of genes that require further investigation for a more comprehensive understanding of stress tolerance mechanisms.
An increase in intracellular calcium concentration occurs in response to various stimuli, such as the accumulation of the hormone abscisic acid (ABA) in leaves during water deficit conditions [
15]. Upon reaching the plasma membrane, ABA binds to membrane receptors, leading to an increase in cytosolic calcium that can occur either via the transient entry of Ca
2+ ions into the intracellular environment or via the release of these ions from internal reservoirs, such as the endoplasmic reticulum and the vacuole. Such variation is essential in triggering signaling cascades in response to plant acclimatization. This process involves calcium-binding proteins that transmit the calcium signal, thereby inducing specific cellular and physiological responses [
16].
Three main classes of Ca
2+-binding proteins have been characterized in plants, namely calcium-dependent protein kinases (CDPKs), calmodulins, and B-like calcineurin proteins [
17]. Among these classes, calcium-dependent protein kinases, commonly referred to as CPKs or CDPKs, are of particular significance due to their capacity to bind and transmit intracellular Ca
2+ signals via a single gene product [
18]. Such a combination may have originated in ancestral organisms through the fusion of protein kinase and calmodulin genes, wherein the former can modify other proteins by adding phosphate groups, while the latter can bind calcium [
19]. Thus, CPKs can directly activate and regulate target proteins containing serine and threonine (Ser/Thr) residues upon detecting intracellular Ca
2+ signals, facilitated via specific domains within their structural composition.
In plants, CPKs are widely distributed across various tissues, including the roots, stems, leaves, and flowers [
20]. At the cellular level, they are abundant in the meristem, xylem, pollen, guard cells, and embryonic cells [
21]. Subcellularly, they are found in the cytosol, nucleus, tonoplasts, mitochondria, chloroplasts, and peroxisomes, with the plasma membrane being the location where most CPKs are located in the model plant
Arabidopsis. The diverse presence of CPKs in different parts of plants suggests their involvement in various signal transduction pathways [
22]. It is well known that CPKs play multiple roles in plant biology, including senescence and cell death, hormone signal transduction, stress and defense responses, growth and development, carbon and nitrogen metabolism, cytoskeletal formation, and regulation of ion channels [
23]. Protein kinases are species-specific and are encoded by a multi-gene family [
24]. For instance, through genomic analysis, 34 CPK genes have been identified in
Arabidopsis [
25], and 31 CPK genes have been identified in rice [
26].
Some CPKs have been functionally characterized in relation to water deficit tolerance in
Arabidopsis. Transgenic plants overexpressing the
AtCPK6 gene have been found to exhibit increased tolerance to water and salt deficits, along with elevated transcription levels of this gene under stress conditions [
27]. Several CPK genes appear to play a role in regulating stomatal opening and closing in
Arabidopsis, a phenomenon also observed in maize with the
ZmCPK4 gene, which was found to increase water deficit tolerance by influencing stomatal closure through abscisic acid (ABA)-mediated pathways [
28]. In
Arabidopsis, the overexpression of
CPK10 was shown to lead to enhanced water deficit tolerance through the participation of
CPK10 in ABA- and calcium-mediated stomatal movements [
29]. It is important to note that stomatal closure, despite reducing transpiration, limits gas exchange. This decrease in carbon dioxide assimilation generally results in reduced biomass and productivity in environments subjected to water deficits.
Functional studies involving different CPKs in relation to water deficit tolerance have already been conducted in rice. The
OsCPK4 gene has been associated with enhanced water retention capacity in plants overexpressing this gene [
30]. Under water deficit conditions, the
OsCPK10 gene increased the hydrogen peroxide detoxification capacity in rice plants [
31]. Overexpression of the
OsCPK9 gene led to improved stomatal closure and plant osmotic adjustment capacity, along with enhanced pollen viability and increased spikelet fertility [
32]. The functions of CPK genes in rice, particularly in relation to water deficit tolerance, have not been yet fully understood, and studies involving gene overexpression in genetically modified plants (GMOs) compared to non-genetically modified plants can contribute to a better comprehension of the roles that these proteins play in plant adaptation to adverse water conditions. Such research also has the potential to aid in the development of commercial cultivars with increased drought tolerance. The objective of this work was to study the effect of the overexpression of
OsCPK5 in the genetically modified (GM) BRSMG Curinga upland rice plants in comparison to the non-genetically modified (NGM) BRSMG Curinga plants subjected to water deficit.
2. Results
The performance of the GM and NGM plants was compared in relation to two irrigation treatments. When comparing the harvest index (HI) of the GM and NGM plants, we observed that the GM plants subjected to the control treatment exhibited a HI 11.1% higher than that of the NGM plants in the same treatment (
p < 0.05;
Table 1). Conversely, no significant difference between the genotypes was observed in the water deficit treatment. Statistical differences (
p < 0.05) were observed for each genotype (GM and NGM plants) across different irrigation treatments for the following traits: grain yield, number of filled grains, number of empty grains, and fresh mass (the latter only for the GM genotype). On the other hand, no significant difference was observed for the following traits: tiller number, panicle number, flag leaf length and width, and dry mass (
Table 1).
The GM plants subjected to water deficit showed a drought susceptibility index (DSI) of 0.97 and were determined to be relatively tolerant to drought stress, according to the criteria established in [
33]. Hence, the overexpression of the
OsCPK5 gene in GM plants was found to confer relative tolerance to water deficit, indicating that there would be a reduced drought impact on the performance of these GM upland rice plants. The DSI of 1.08 for BRSMG Curinga (NGM) indicates that this cultivar straddles a delicate balance between tolerance and sensitivity to water deficit, as observed by the authors of [
34].
After 14 days of water deficit in the reproductive phase (stage R6), the transpiration rate (E) of GM plants was 42.1% lower than that of the NGM plants (
p < 0.05;
Table 2). At this stage, both the GM and NGM plants exposed to water deficit exhibited 44.5% and 23.2% lower transpiration rates than their corresponding control plants, respectively (
p < 0.05). At the R6 stage, the decline in the transpiration rate (E) was the result of the reduction in stomatal conductance (gs), reaching 51.2 and 33.0% for the GM and NGM plants, respectively (
Table 2). It is worth mentioning that the GM plants exhibited a stomatal conductance 46.7% lower than that of the NGM plants subjected to water deficit. Regarding carboxylation efficiency (A/Ci), no differences were evident between the GM and NGM genotypes under any irrigation treatment at any collection period. Moreover, it was observed that the GM plants maintained a consistent A/Ci across all collection periods in both irrigation treatments (
Table 2).
Regarding the intrinsic water-use efficiency (iWUE), under the control treatment, the GM plants had an iWUE 40.7% higher than that of the NGM plants under the second collection period (R6 stage) (
Table 2). Comparing the GM and NGM plants grown under the water deficit and control treatments at each data collection stage, it was observed that in the third collection period (7 days after the return of regular irrigation), the iWUE of the GM plants under water deficit was not significantly different from that of the corresponding plants in the control treatment. The NGM plants cultivated under water deficit had a 41.4% lower iWUE than the corresponding plants in the control treatment.
The GM plants also exhibited a notable intrinsic water-use efficiency (WUEintr). In the second collection period (stage R6), the GM plants under the control treatment displayed a WUEintr that was 40.2% higher than that of the NGM plants under the same irrigation conditions (
Table 2). The higher WUEintr value in the GM plants occurred under the second collection period (stage R6) across irrigation treatments. Both genotypes showed higher WUEintr values 14 days after the irrigation cut-off relative to the collection periods. However, the GM plants were 40.2% more efficient in using water compared to their corresponding NGM plants under water deficit.
Discrepancies in the chlorophyll a and b proportions were observed between the GM and NGM genotypes during the first collection period (79 DAS, stage R3). In this period, the GM plants exhibited a chlorophyll a/b ratio that was 25% and 23.1% higher than that of the NGM plants under control and water deficit conditions, respectively (
Table 3). In the second collection period (93 DAS, stage R6), no significant differences between the genotypes were observed in either irrigation treatment. However, during the third collection period (101 DAS, stage R8), the GM plants displayed chlorophyll a/b ratios that were 10.7% and 12% higher than those of the NGM plants under the control and water deficit treatments, respectively. No significant differences were observed in the chlorophyll a/b ratio during the first and second collection periods between plants of the same genotype grown under control conditions and those grown under water deficit treatment. However, during the third collection period, NGM plants exhibited a notable difference of 13.7% in the chlorophyll a/b ratio between plants grown under the control and water deficit treatments.
Through visual comparisons of the general appearance of the GM and NGM plants subjected to water deficit (
Figure 1), it was observed that in the second collection period (93 DAS, stage R6), 14 days after irrigation cut-off, the GM plants sustained a green coloration of their leaves. Conversely, the NGM plants exhibited numerous senescent leaves from the second collection period, a condition that persisted into the third collection period (101 DAS, stage R8), even following the restoration of irrigation.
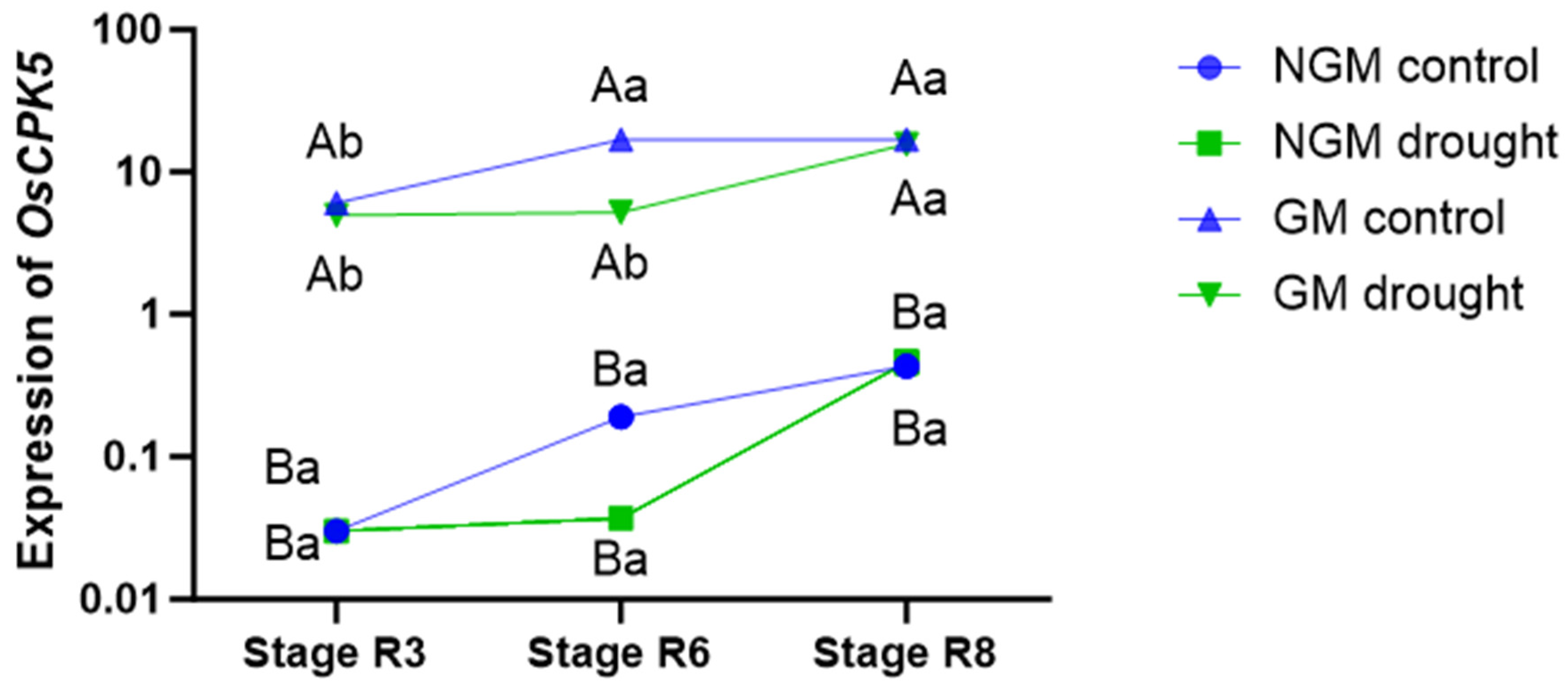
The GM plants exhibited higher
OsCPK5 gene expression levels compared to the NGM plants under all collection periods and in both irrigation treatments (
Figure 2). In the control treatment, the GM plants showed the highest level of
OsCPK5 gene expression during period 2 (stage R6), and this pattern persisted in period 3 (stage R8). For the GM plants subjected to the water deficit treatment, the highest expression of the
OsCPK5 gene was observed in period 3 (stage R8), 7 days after the return of regular irrigation. No significant difference in
OsCPK5 gene expression was observed between NGM plants under the control irrigation treatment and those under the water deficit treatment under any of the collection periods. Furthermore, in the NGM plants, there was no significant difference in the expression of the
OsCPK5 gene among the three collection periods under either irrigation treatment.
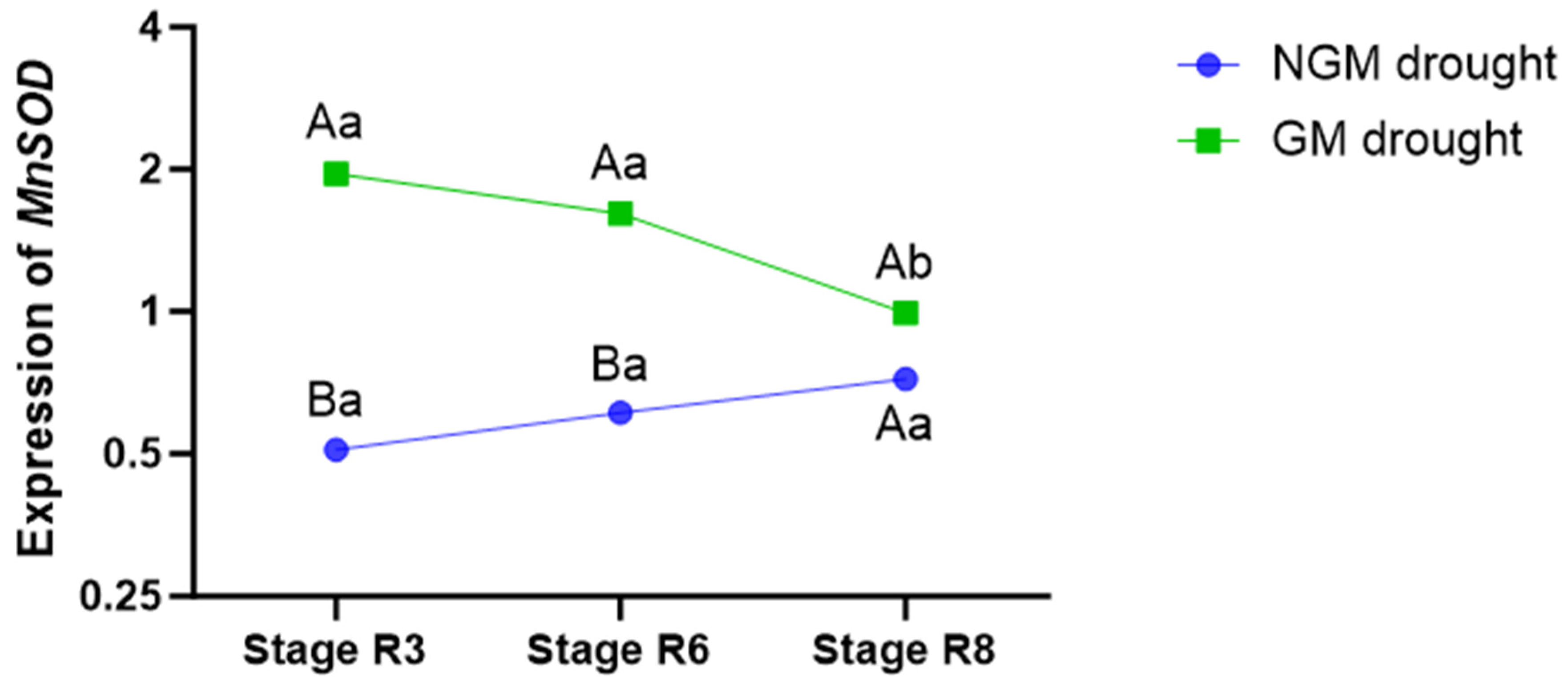
In order to investigate the underlying factors contributing to the delayed leaf senescence in the GM plants, we examined the gene expression of
MnSOD, an essential component in the defense against oxidative stress in rice plants subjected to water deficit conditions. Compared to the NGM plants, the GM plants exhibited higher transcription levels of this gene starting from the first collection period (stage R3), and this elevated expression persisted (
p < 0.05) until the end of the water deficit phase, corresponding to the second collection period (stage R6). Seven days after the return of irrigation (stage R8), the
MnSOD expression level of the GM plants cultivated under water deficit conditions equaled that of the NGM plants under the same irrigation treatment (
Figure 3).
RNA-seq generated an average of 43.6 Gbp of clean reads, of which 41.7 Gb (95.6%) were aligned to the rice reference genome (
Table 4). Five billion reads were aligned to exons (89.3%), with the highest alignment achieved in well-annotated reference genomes. The 10 most expressed genes, determined via the abundance of transcripts mapped to the genome, are described in
Table 5. For illustrative purposes, the transcript of the
OsCPK5 gene used in the transformation (Os02g0685900) has been added to
Table 5.
Gene expression level analysis stands as a central task in a RNA-seq experiment, with the gene expression level being computed through the count of mapped reads. When contrasting the libraries of the GM and NGM plants exposed to drought conditions, the FPKM count for the Os02g0685900 transcript (OsCPK5) reached 233,193,538,410,951 (upregulated) for the GM plants, whereas for the NGM plants, it amounted to only 0.52.
A functional classification of GM vs. NGM sequences from drought libraries based on a gene ontology (GO) analysis revealed two groups of significantly upregulated genes: those related to biological process (BP), encompassing 13 classes, with “response to the oxygen-containing compound” showing the highest gene count at 22, and those related to molecular function (MF), representing four categories, with the category “unfolded protein binding” hosting 10 genes. Regarding significantly downregulated genes, three GO groups were identified: BP, with 89 classes, where “defense response” featured 43 genes; MF comprising 19 classes, with “carbohydrate binding” harboring 34 genes; and cellular component (CC) with one class, where “region extracellular” comprised 45 genes.
Examining the upregulated genes within the BP category, the classes most relevant to water deficit included “response to hydrogen peroxide” (GO:0042542), with six heat shock protein (hsp) genes, “response to reactive oxygen species” (GO:0000302) comprising five hsp genes, two peroxidases, and one aminotransferase, “osmotic stress response” (GO:0006970, see
Table 6), and “oxidative stress response” (GO:0006979) with five hsp genes, two peroxidases, one aminotransferase, one HLH protein, and two unknown proteins). Notably, the last two classes were absent in the GO analysis of the GM × NGM control libraries. Among the downregulated genes in the BP category, the most directly related was “stress response” (GO:0080134, see
Table 7). This class also did not appear in the GO analysis of the GM × NGM control libraries.
The enrichment analysis identified two statistically significant KEGG pathways associated with differentially expressed genes between the GM and NGM transcripts (the libraries from the drought treatment) (
Table 8). None of these pathways were identified in the GM versus NGM libraries for the control treatment, but “Ribosome”, “Photosynthesis—antenna protein”, “Ribosome biogenesis in eukaryotes”, and “Phenylpropanoid biosynthesis” were found.
3. Discussion
This study highlighted how
OsCPK5 gene overexpression in GM plants impacts the regulation of stomatal closure, resulting in a decrease in stomatal conductance (gs) under WD conditions. Stomatal closure serves as the primary leaf response to drought conditions, minimizing water loss by decreasing the transpiration rate (E) and increasing the water-use efficiency (WUE) [
35]. Notably, significant differences in gs and E were observed between the GM and NGM plants during the second collection period (stage R6), a critical phase when water loss intensifies due to grain filling in the panicles. Despite the identification of this important physiological difference between the two genotypes, with the GM plants being more efficient in their water use, both the GM and NGM plants presented the same grain yield. Certainly, this outcome arises from the intricate nature of grain productivity, influenced as it is via an array of metabolic pathways and genetic interactions.
To address water deficit, plants close their stomata to maintain cell turgor and metabolism, thereby affecting the photosynthetic rate [
2]. Photosynthesis is highly sensitive to drought stress and is the foremost process that is altered by such conditions. Decreased production of photoassimilates reduces leaf growth and crop yield [
36]. The enhancement of stomatal regulation, probably due to the overexpression of
OsCPK5, has also been reported in studies involving other genes of the CPK family. For instance, the authors of [
28] investigated the impact of overexpressing the corn
ZmCPK4 gene in
Arabidopsis thaliana and found an amplified sensitivity of the plant to the hormone abscisic acid (ABA) that increased stomatal closure in response to water deficit. The authors of [
32] observed that compared to control plants, transgenic rice plants overexpressing
OsCPK9 under normal growth conditions exhibited no significant differences in stomatal opening; however, following a period of water deficit, a higher proportion of fully closed stomata was observed in the transformed plants, in contrast to what was observed in the control plants, suggesting that the
OsCPK9 gene influences stomatal movement under water deficit conditions.
Water deficit-tolerant plants respond to drought mainly through fine control of stomatal and mesophilic conductance [
37], and under these conditions impairment of CO
2 assimilation may occur, as well as restrictions on growth and metabolism [
38]. However, in this study, it was observed that the improvement in stomatal closure in the GM plants at stage R6 (the grain-filling phase), following a 14-day water deficit, did not lead to a reduction in the productivity of this genotype when compared to the NGM plants cultivated under similar irrigation treatments. Several factors may have contributed to decreasing the impact of water restriction on the productivity of GM plants relative to NGM plants, even after a substantial reduction in stomatal conductance and transpiration rate during the grain-filling phase. These factors include greater water-use efficiency during the grain-filling phase, delayed leaf senescence, and higher levels of chlorophyll a and b in the GM plants than in the NGM plants.
The stability in carboxylation efficiency observed in the GM plants under the three collection stages in both irrigation treatments could potentially be linked to enhanced protection against the degradation of Rubisco (ribulose-1,5-bisphosphate carboxylase/oxygenase). Another probable reason was that ribulose activase, the enzyme that regulates Rubisco activation [
39], was the most expressed enzyme identified via RNA-seq, with the GM plants under water deficit having 9% more transcripts of this enzyme than the NGM plants under the same irrigation conditions and 18% more transcripts than the NGM plants in the control treatment. Furthermore, the elevated protection against the degradation of this enzyme after re-irrigation in the GM plants may have contributed to a better cellular redox balance, thereby creating a more favorable cellular environment for its preservation and activity. The carboxylation efficiency was also noted by the authors of [
40] when evaluating the effect of water deficit on Kentucky bluegrass (
Poa pratensis L.), a C3 perennial grass. In that study, the drought-tolerant genotype demonstrated an enhanced activity and state of activation of Rubisco following re-irrigation, restoring metabolic activity to levels comparable to those of control plants. This phenomenon was considered a plausible cause for the recovery of photosynthetic activity.
Water-use efficiency (WUE) is associated with the ability of plants to deal with varying degrees of water deficit and plays a pivotal role in maintaining productivity even when water availability is restricted [
41]. During the critical water deficit period (R6 stage), the GM plants overexpressing the
OsCPK5 gene exhibited higher iWUE levels than the NGM plants. The iWUE level observed after the water deficit period suggested that the NGM plants were unable to restore their CO
2 assimilation machinery, leading to malfunction of the photosynthetic apparatus and degradation of pigments, compromising the different stages of photosynthesis [
42]. The correlation between gs reduction and greater water-use efficiency has been observed by the authors of [
43,
44], who evaluated C3 plants (such as rice) under moderate water deficit scenarios.
Reducing the gs per amount of CO
2 assimilated and increasing the rate of CO
2 assimilation can improve the WUEintr [
45]. In the R6 stage, the GM plants overexpressing the
OsCPK5 gene showed higher WUEintr levels than the NGM plants under both irrigation treatments. In fact, in contrast to the NGM plants under water deficit, the GM plants exhibited a notable decrease in gs and E at 14 days after cutting off irrigation (stage R6), concomitant with an augmentation in WUEintr at this particular stage. In wheat, negative correlations among photosynthesis, transpiration, and stomatal conductance on water-use efficiency have been reported, suggesting that stomatal characteristics are factors responsible for regulating water-use efficiency in these plants [
43]. Increasing the water-use efficiency of rice is critical, as rice grown under the lowland system require more than 2.5 kg of water for each grain of rice produced [
2].
Visually, the GM rice plants overexpressing the
OsCPK5 gene showed a greener leaf appearance than the NGM plants during the grain-filling phase after 14 days of irrigation restriction, while the leaves of the NGM plants turned yellow and senescent. The correlation between the overexpression of a calcium-dependent protein kinase and the green appearance of leaves was also observed by the authors of [
30] when examining the overexpression of
OsCPK4 to increase tolerance to water deficit in rice plants of the cultivar Nipponbare (
Oryza sativa ssp.
japonica). Visual differences between plants overexpressing the
OsCPK4 gene and control plants were observed from the 14th day without irrigation, when the control plants (wild type) showed symptoms of damage induced via water deficit, such as leaf wilting, which remained until the last day of cutting irrigation (17th day). On the other hand, transgenic plants overexpressing
OsCPK4 remained healthy and green. These authors verified that after rehydration of the plants, approximately 90% of the transgenic plants recovered from the stress, while none of the control plants survived the water deficit treatment. In our experiment, a similar result occurred, as the return of irrigation for 7 days (stage R8) failed to rejuvenate the NGM plants after the applied water deficit, increasing the senescent appearance of their leaves in comparison with those of the GM plants, where senescence was delayed. Plants with the stay-green trait are important in the case of drought, as the extension of the photosynthesis period leads to greater grain productivity [
46].
According to the authors of [
47], the relationship between the expression of CPK family genes and the senescence process in rice leaves was further underscored through the functional validation of the
OsCPK12 gene. In this context, they observed that mutant plants, in which the gene was knocked out, exhibited yellowing and senescent leaves during the grain-filling stage, while wild-type plants exhibited a greener appearance. Investigations into the possible reasons for this difference revealed that mutant plants had a greater accumulation of H
2O
2 and superoxide radicals in leaves, in addition to more cell death and apoptosis, than wild-type plants. On the other hand,
OsCPK12 overexpression led to higher photosynthetic rates, increased chlorophyll a and b contents, delayed leaf senescence, and a delayed plant growth period, providing positive effects for crop productivity.
There is a positive correlation between increased
SOD gene expression and increased plant tolerance to environmental stresses [
3]. In this work, the evaluation of the expression of
MnSOD, important in preventing oxidative stress [
48], revealed that the GM plants overexpressing
OsCPK5 grown under water deficit treatment presented a rapid and strong activation of this defense response from the R3 stage that persisted until the end of the irrigation cut during the reproductive phase. Levels of antioxidant metabolites and enzymes that regulate the cellular redox state increase under different stresses, i.e., a greater antioxidant activity confers greater tolerance to these stresses in plants [
49]. Increased expression/activity of total superoxide dismutases (SODs) due to water deficit was observed in rice [
50]. Furthermore, the authors of [
51], when evaluating the enzymatic activity and gene expression levels of eight SOD isoforms in rice, observed that more SOD genes were expressed in the vegetative phase than under the reproductive phase. However, the enzymatic activity of SOD only increased during the reproductive phase of the plants.
In the current study, although quantification in the greenhouse experiment revealed that the values of the chlorophyll a/b ratio of the GM and NGM plants were not significantly different, the GM plants presented 35% more transcripts of the chlorophyll a/b binding protein-encoding gene than the NGM plants under water deficit, while the GM plants had 75% more transcripts than the NGM plants under control irrigated treatment. Light-harvesting chlorophyll a/b binding proteins, according to the authors of [
52], play a role in the efficiency of photosynthesis, in addition to participating in the response to abiotic stresses. As pointed out by the authors of [
3], an increased proportion of chlorophyll a and b is indicative of greater drought tolerance, which provides further evidence that the GM plants have a better response to drought. The relationship between overexpression of a calcium-dependent protein kinase and an increase in chlorophyll content was also observed by the authors of [
32] following a water deficit treatment in rice plants overexpressing the
OsCPK9 gene (
Oryza sativa, cultivar Nipponbare) in contrast to plants with silencing of this gene and control plants. The authors of [
47], when investigating possible reasons for differences in chlorophyll content from overexpression or silencing of CPK family genes in
Oryza sativa ssp.
indica, noted that mutant plants of genes of the CPK family presented negative regulation of genes involved in the synthesis of chlorophylls and positive regulation of genes involved in the degradation of these pigments. This fact may indicate that overexpression of
OsCPK5 also acts on the synthesis or degradation pathways of these pigments in upland rice, paving the way for new investigations into which genes and metabolic pathways related to the synthesis or degradation of chlorophylls were altered from overexpression of the
OsCPK5 gene.
The leaves of the GM plants overexpressing the endogenous and exogenous
OsCPK5 gene showed higher transcription levels of this gene than the NGM plants under both irrigation treatments at the three evaluated collection periods (stages R3, R6, and R8). A similar result was obtained by the authors of [
27] when examining the transcription levels of the
AtCPK6 gene, an orthologue of
OsCPK5, in
A. thaliana, in which the levels of the
AtCPK6 gene in transformed plants were higher than those in wild-type plants and associated with increased drought tolerance. The
OsCPK5 gene was poorly expressed in relation to the other transcripts identified via RNA-seq analysis. Even so, the transcription level of the
OsCPK5 gene was slightly higher in the GM plants than in the NGM plants, which indicates that the best alternative to monitor the presence of specific genes is quantitative PCR. The authors of [
26], when evaluating CPK family genes in
Oryza sativa ssp.
indica using microarrays and RT-qPCR, observed that fourteen CPK genes were upregulated during the panicle development stage and six during the seed development stage, indicating that CPK expression is also regulated via the rice development stage.
The majority of the upregulated transcripts in the GM drought libraries identified via the GO analysis were “heat shock proteins” (HSPs), which have important roles in transducing cell signaling, regulating apoptosis, and protecting cells against biotic and abiotic stresses [
53]. Conversely, the downregulated transcripts were mainly TIFY genes, which exert a regulatory function in plant development and responses to biotic and abiotic stresses [
54], exhibiting a similar function to HSPs. The WRKY transcription factor, which plays a key role in transmitting and responding to drought stress signals, was also found to be downregulated [
3]; that is, WRKY expression was higher in the NGM plants. According to the KEGG analysis, TIFY proteins were identified in the “plant hormone signal transduction” pathway, showing higher expression levels in the NGM plants than in the GM plants under water deficit. The other pathway identified via KEGG analysis was the “MAPK signalling pathway”, associated with the mitogen-activated protein kinase (MAPK) cascade, a defense mechanism induced against abiotic stresses in plants [
55]. Most of the genes in this pathway were upregulated in the NGM plants, supporting the hypothesis that these plants are more adversely affected by the consequences of water deficit than GM plants.



 ,
, 

{kind=link}
{kind=link}
{kind=link}