
Breeding of Modern Rose Cultivars Decreases the Content of Important Biochemical Compounds in Rose Hips



Abstract
:1. Introduction
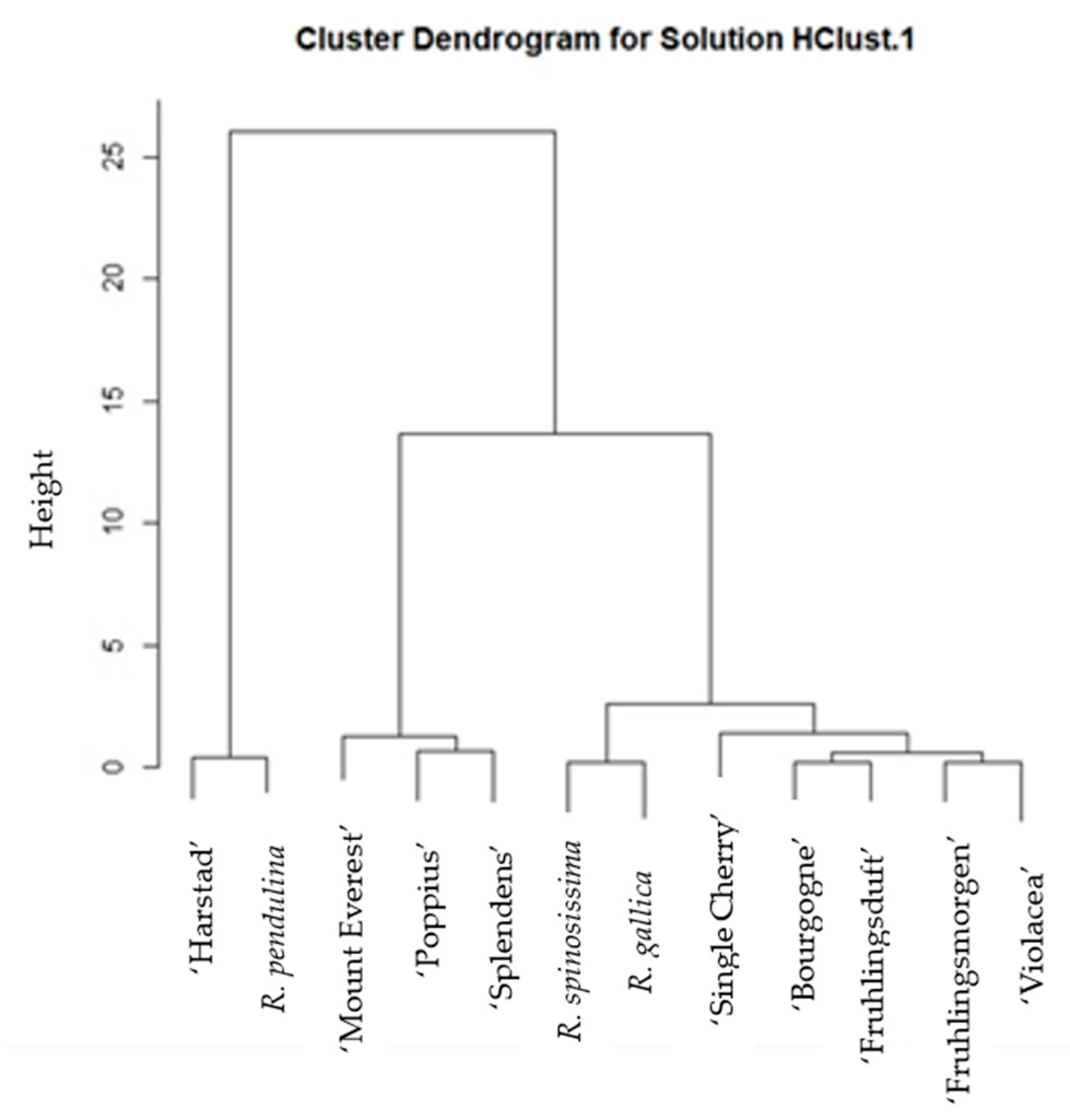
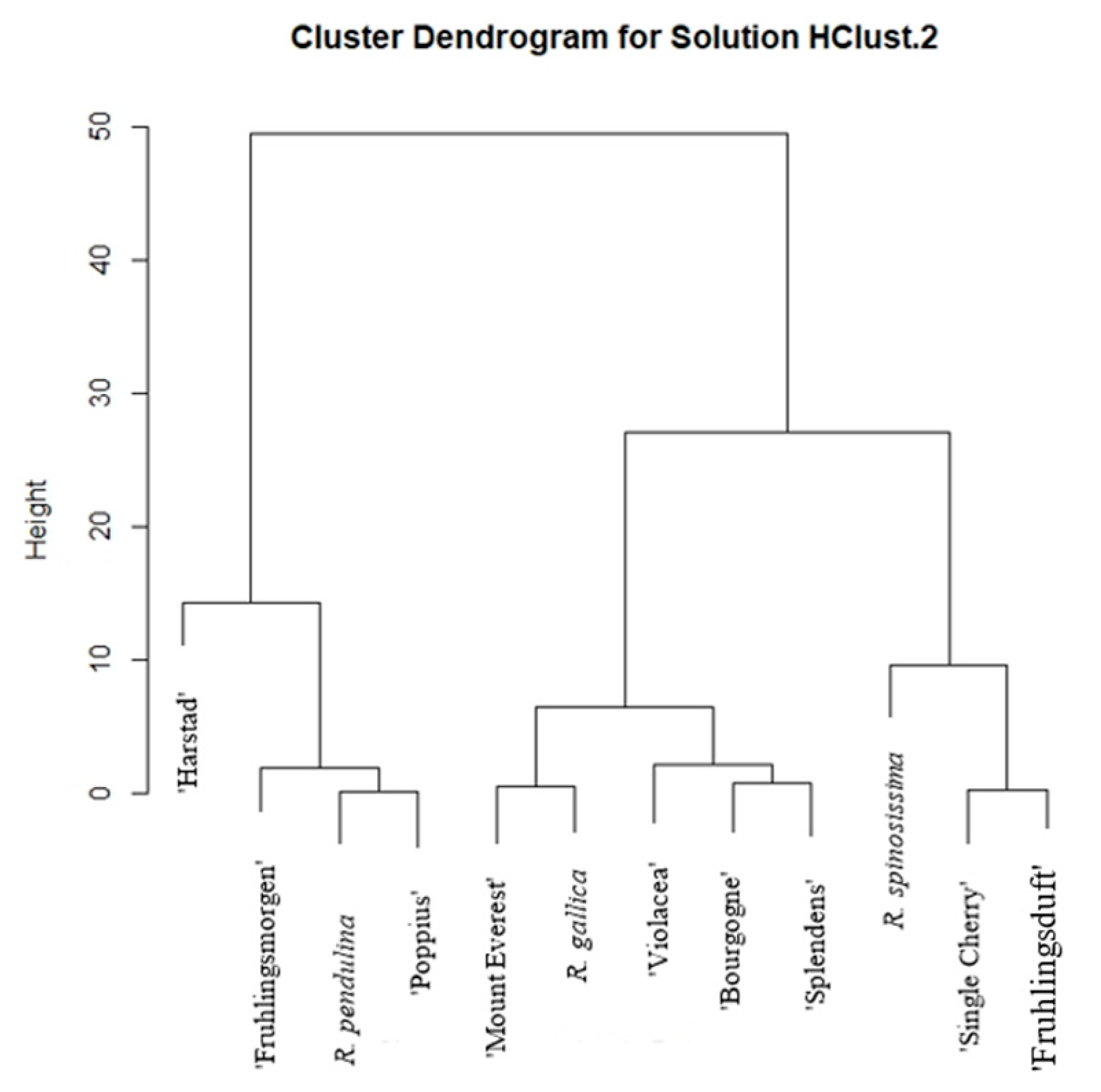
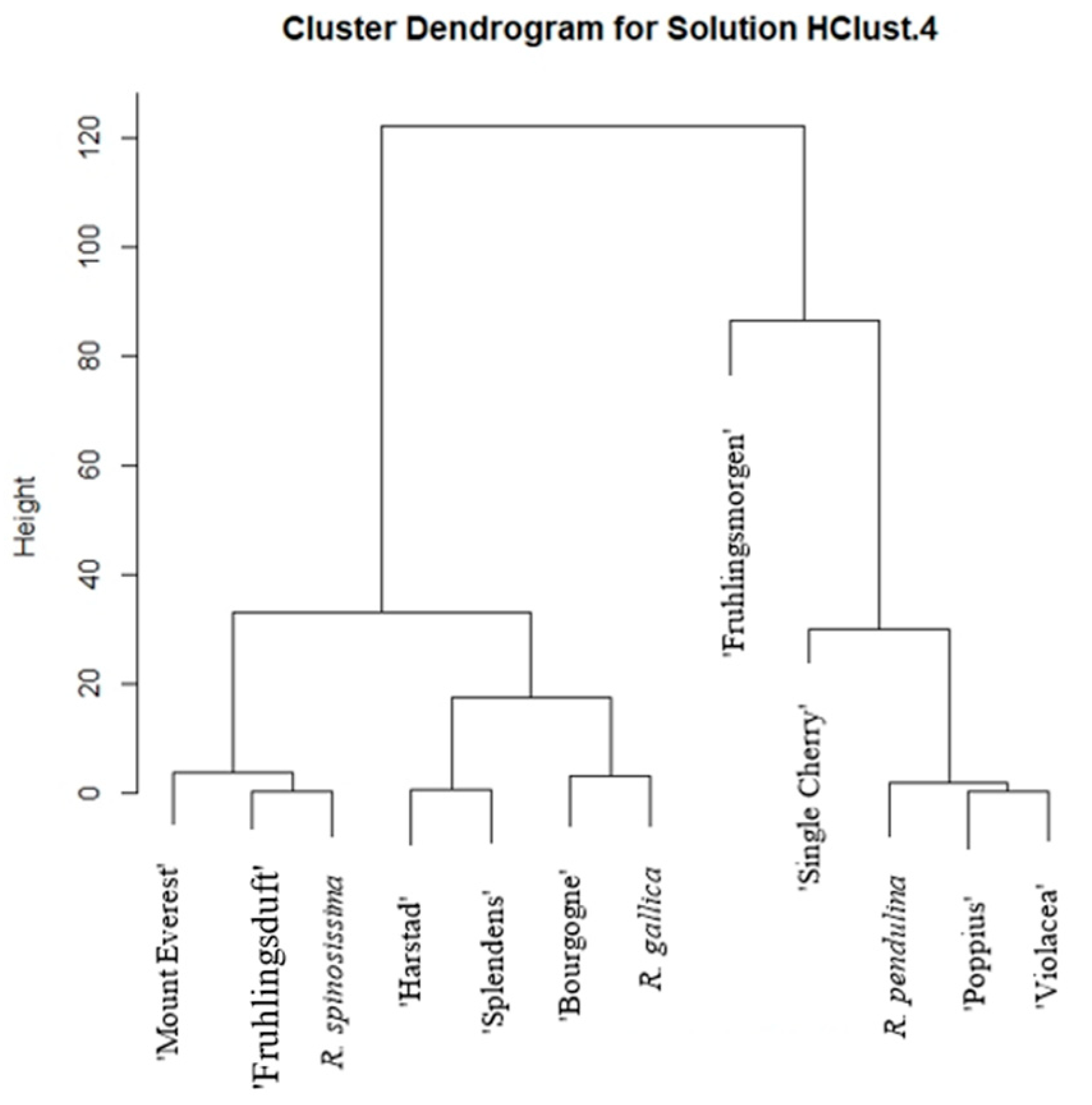
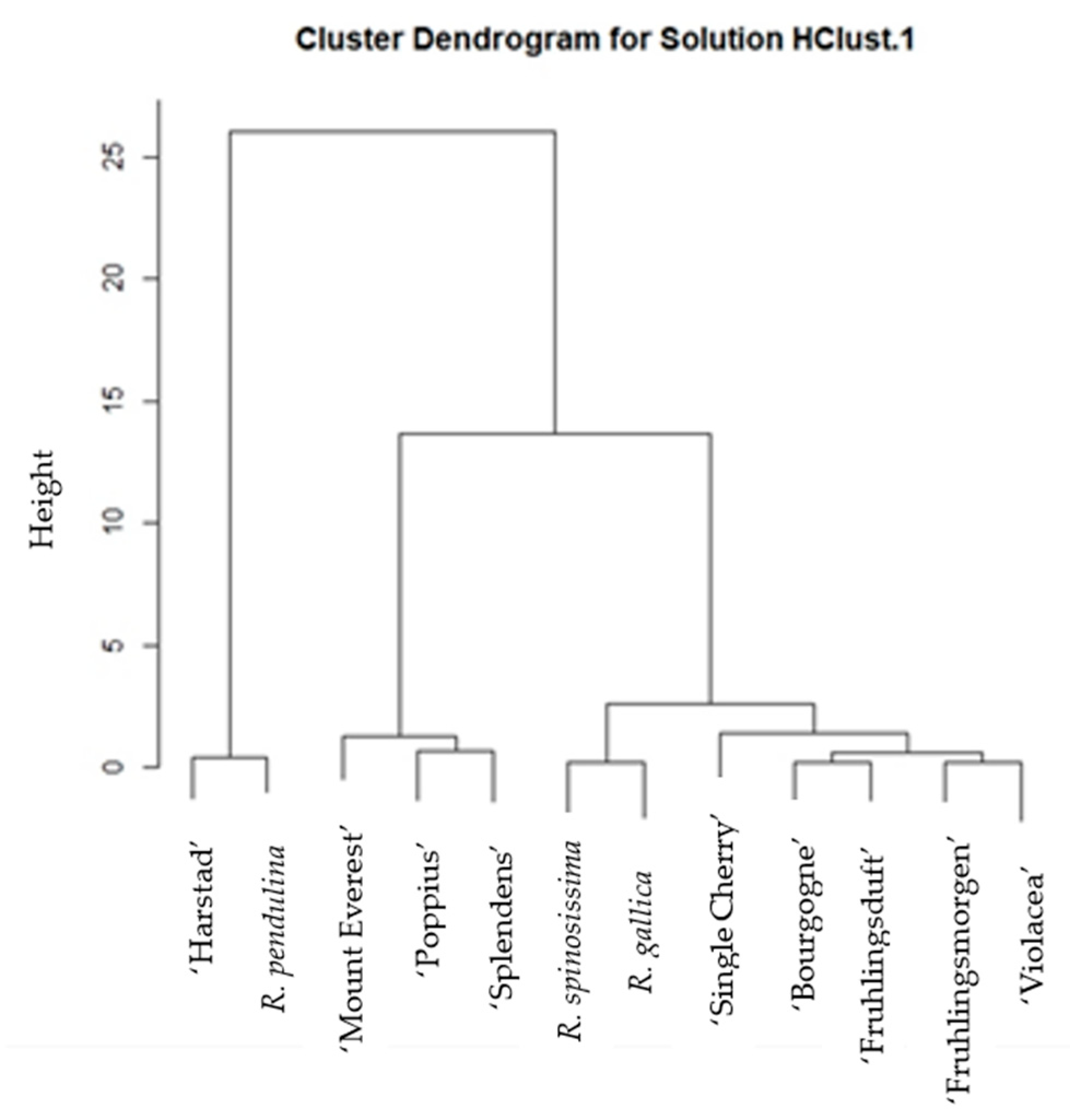
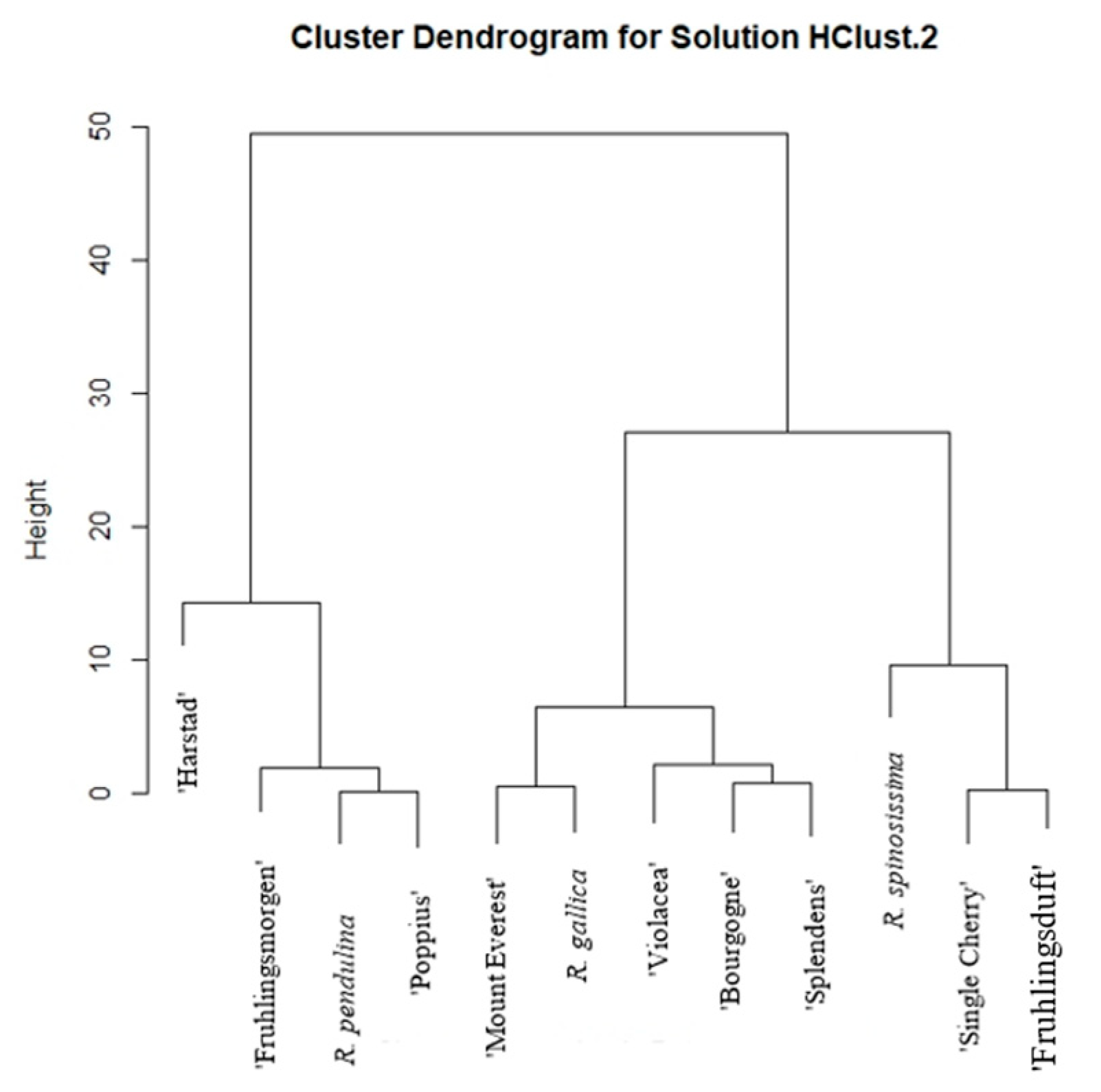
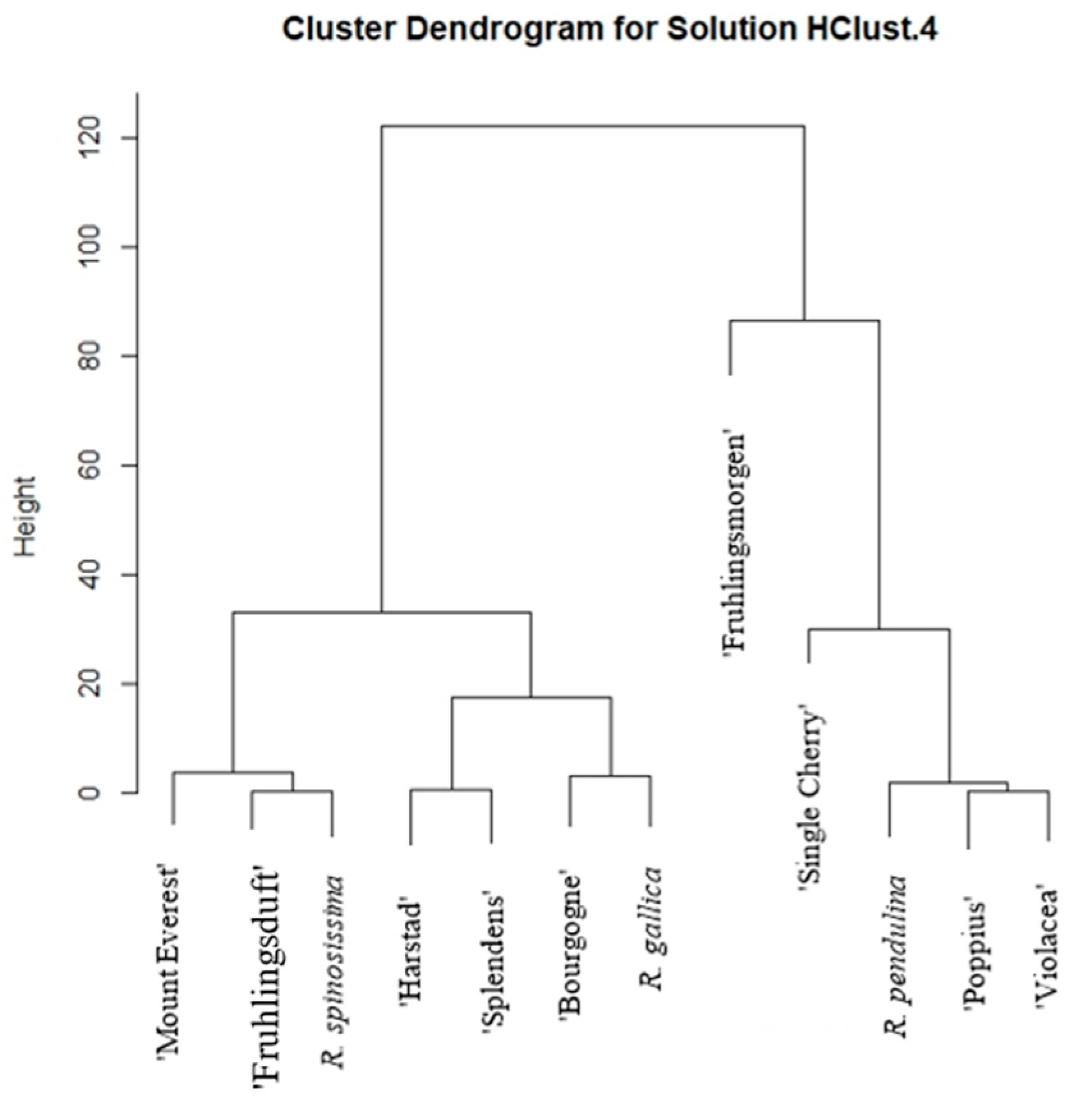
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Extraction and Analysis of Ascorbic Acid
4.3. Extraction and Analysis of Organic Acids
4.4. Extraction and Analysis of Carotenoids
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meier, U.; Bleiholder, H.; Brumme, H.; Bruns, E.; Mehring, B.; Proll, T.; Wiegand, J. Phenological growth stages of roses (Rosa sp.): Codification and description according to the BBCH scale. Ann. Appl. Biol. 2009, 154, 231–238. [Google Scholar] [CrossRef]
- Jones, P. Clinical nutrition: 7. Functional foods—More than just nutrition. CMAJ 2002, 166, 1555–1563. [Google Scholar] [PubMed]
- Kunc, N.; Hudina, M.; Bavcon, J.; Vreš, B.; Luthar, Z.; Gostinčar, K.; Mikulic-Petkovsek, M.; Osterc, G.; Ravnjak, B. Characterization of the Slovene autochthonous Rose hybrid Rosa pendulina × spinosissima (Rosa reversa Waldst. and Kit) using biochemical patterns of the plant blossoms. Plants 2023, 12, 505. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.; Pacier, C.; Martirosyan, D. Rosehip (Rosa canina L.): A functional food perspective. Funct. Foods Health Dis. 2014, 4, 493–509. [Google Scholar] [CrossRef]
- Martínez-Valverde, I.; Periago, M.J.; Ros, G. Significado nutricional de los compuestos fenólicos de la dieta [Nutritional importance of phenolic compounds in the diet]. Arch. Latinoam. Nutr. 2000, 50, 5–18. [Google Scholar] [PubMed]
- Tumbas Šaponjac, V.; Čanadanović-Brunet, J.; Četojević Simin, D.; Ćetković, G.; Ðilas, S.; Gille, L. Effect of rosehip (Rosa canina L.) phytochemicals on stable free radicals and human cancer cells. J. Sci. Food Agric. 2012, 92, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Ashtiyani, C.S.; Najafi, H.; Jalalvandi, S.; Hosseinei, F. Protective effects of Rosa canina L. fruit extracts on renal disturbances induced by reperfusion injury in rats. Iran. J. Kidney Dis. 2013, 7, 290–298. [Google Scholar]
- Paciolla, C.; Fortunato, S.; Dipierro, N.; Paradiso, A.; De Leonardis, S.; Mastropasqua, L.; de Pinto, M.C. Vitamin C in plants: From functions to biofortification. Antioxidants 2019, 8, 519. [Google Scholar] [CrossRef]
- Fenech, M.; Amaya, I.; Valpuesta, V.; Botella, M.A. Vitamin C content in fruits: Biosynthesis and regulation. Front. Plant. Sci. 2019, 9, 2006. [Google Scholar] [CrossRef]
- Hoyle, S. Study shows vitamin C is essential for plant growth. Plant J. 2007, 1. Available online: https://www.eurekalert.org/news-releases/561248 (accessed on 5 August 2023).
- Li, Y.; Schellhorn, H.E. New developments and novel therapeutic perspectives for vitamin C. J. Nutr. 2007, 137, 2171–2184. [Google Scholar] [CrossRef] [PubMed]
- Locato, V.; Cimini, S.; De Gara, L. Strategies to increase Vitamin C in plants: From plant defense perspective to food biofortification. Front. Plant. Sci. 2013, 4, 152. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.C.; Rowe, S. Factors affecting Vitamin C status and prevalence of deficiency: A global health perspective. Nutrients 2020, 12, 1963. [Google Scholar] [CrossRef] [PubMed]
- Kunc, N.; Mikulič-Petkovšek, M.; Hudina, M.; Bavcon, J.; Vreš, B.; Osterc, G.; Ravnjak, B. Autochthonous rose hybrid Rosa pendulina × spinosissima overshines main genotype Rosa pendulina in the biochemical characteristics of their hips. Horticulturae 2022, 8, 669. [Google Scholar] [CrossRef]
- Darlington, C.D. Vitamin C and chromosome number in Rosa. Nature 1942, 150, 404. [Google Scholar] [CrossRef]
- Uggla, M.; Gustavsson, K.E.; Olsson, M.E.; Nybom, H. Changes in colour and sugar content in rose hips (Rosa dumalis L. and R. rubiginosa L.) during ripening. J. Hortic. Sci. Biotechnol. 2005, 80, 204–208. [Google Scholar] [CrossRef]
- Babis, A.; Kucharska, A. Przydatność owoców Rosa spinossima i Rosa hybrida do produkcji wysokowitaminowych soków mȩtnych. Biul. Wydz. Farm. AMW 2004, 3, 1–24. [Google Scholar] [CrossRef]
- Javanmard, M.; Asadi-Gharneh, H.A.; Nikneshan, P. Characterization of biochemical traits of dog rose (Rosa canina L.) ecotypes in the central part of Iran. Nat. Prod. Res. 2018, 32, 1738–1743. [Google Scholar] [CrossRef]
- Kaushik, P.; Andújar, I.; Vilanova, S.; Plazas, M.; Gramazio, P.; Herraiz, F.J.; Brar, N.S.; Prohens, J. Breeding Vegetables with Increased Content in Bioactive Phenolic Acids. Molecules 2015, 20, 18464–18481. [Google Scholar] [CrossRef]
- Khanizadeh, S.; Ehsani-Moghaddam, B.; Tsao, R.; Desjardins, Y.; Gosselin, A. Strategy for breeding fruits with higher content in bioactive compounds. Acta Hortic. 2007, 744, 225–232. [Google Scholar] [CrossRef]
- Igamberdiev, A.U.; Eprintsev, A.T. Organic acids: The pools of fixed carbon involved in redox regulation and energy balance in higher plants. Front. Plant Sci. 2016, 7, 1042. [Google Scholar] [CrossRef] [PubMed]
- Panchal, P.; Miller, A.; Giri, J. Organic acids: Versatile stress response roles in plants. J. Exp. Bot. 2021, 72, 4038–4052. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.Y.; Wang, C.K.; Zhao, Y.W.; Sun, C.H.; Hu, D.G. Mechanisms and regulation of organic acid accumulation in plant vacuoles. Hortic. Res. 2021, 8, 227. [Google Scholar] [CrossRef] [PubMed]
- Drincovich, M.F.; Voll, L.M.; Maurino, V.G. Editorial: On the diversity of roles of organic acids. Front. Plant Sci. 2016, 7, 1592. [Google Scholar] [CrossRef] [PubMed]
- Qamar, A.; Braunwald, E. Treatment of Hypertension: Addressing a Global Health Problem. JAMA 2018, 320, 1751–1752. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Zhang, L.Y. Enhancing effects of organic acids on saltiness of low-sodium salt. Sci. Technol. Food Ind. 2012, 33, 370–373. [Google Scholar]
- Cunja, V.; Mikulič Petkovšek, M.; Zupan, A.; Štampar, F.; Schmitzer, V. Frost decreases content of sugars, ascorbic acid and some quercetin glycosides but stimulates selected carotenes in Rosa canina hips. J. Plant Physiol. 2015, 178, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Demir, N.; Yildiz, O.; Alpaslan, M.; Hayaloglu, A. Evaluation of volatiles, phenolic compounds and antioxidant activities of rose hip (Rosa L.) fruits in Turkey. LWT Food Sci. Technol. 2014, 57, 126–133. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W.; Barker, D.H.; Logan, B.A.; Bowling, D.R.; Verhoeven, A.S. Using chlorophyll fluorescence to assess the fraction of absorbed light allocated to thermal dissipation of excess excitation. Physiol. Plant. 1996, 98, 253–264. [Google Scholar] [CrossRef]
- Kovačević, J.; Popović, L.Č.; Dimitrijević, M.S. Analysis of optical Fe II emission in a sample of active galactic nucleus spectra. ApJS 2010, 189, 15–36. [Google Scholar] [CrossRef]
- Kim, J.; DellaPenna, D. Defining the primary route for lutein synthesis in plants: The role of Arabidopsis carotenoid beta-ring hydroxylase CYP97A3. Proc. Natl. Acad. Sci. USA 2006, 103, 3474–3479. [Google Scholar] [CrossRef]
- Šircelj, H. Karotenoidi v fotosinteznem aparatu in odziv na stres. Acta Agric. Slov. 2008, 91, 271–282. [Google Scholar]
- Andersson, S.; Rumpunen, K.; Johansson, E.; Olsson, M. Carotenoid content and composition in rose hips (Rosa spp.) during ripening, determination of suitable maturity marker and implications for health promoting food products. Food Chem. 2011, 128, 689–696. [Google Scholar] [CrossRef]
- Olsson, M.E.; Andersson, S.; Werlemark, G.; Uggla, M.; Gustavsson, K.E. Carotenoids and phenolics in rose hips. Acta Hortic. 2005, 690, 249–252. [Google Scholar] [CrossRef]
- Hornero-Méndez, D.; Mínguez-Mosquera, M. Carotenoid pigments in Rosa mosqueta hips, an alternative carotenoid source for foods. J. Agric. Food Chem. 2000, 48, 825–828. [Google Scholar] [CrossRef] [PubMed]
- Jayram, N.P. A Comparison of Vitamin C in Modern and Older Rose Varieties; Project Number J0406; California State Science Fair: Riverside, CA, USA, 2010; 1p. [Google Scholar]
- Tabaszewska, M.; Najgebauer-Lejko, D. The content of selected phytochemicals and in vitro antioxidant properties of rose hip (Rosa canina L.) tinctures. NFS J. 2020, 21, 50–56. [Google Scholar] [CrossRef]
- Fascella, G.; D’Angiolillo, F.; Mammano, M.M.; Amenta, M.; Romeo, F.V.; Rapisarda, P.; Ballistreri, G. Bioactive compounds and antioxidant activity of four rose hip species from spontaneous Sicilian flora. Food Chem. 2019, 289, 56–64. [Google Scholar] [CrossRef]
- Roman, I.; Stănilă, A.; Stănilă, S. Bioactive compounds and antioxidant activity of Rosa canina L. biotypes from spontaneous flora of Transylvania. Chem. Cent. J. 2013, 7, 73. [Google Scholar] [CrossRef]
- Skrypnik, K.; Bogdański, P.; Sobieska, M.; Suliburska, J. The effect of multistrain probiotic supplementation in two doses on iron metabolism in obese postmenopausal women: A randomized trial. Food Funct. 2019, 10, 5228–5238. [Google Scholar] [CrossRef]
- Adamczak, A.; Buchwald, W.; Zielinski, J.; Mielcarek, S. Flavonoid and organic acid content in rose hips (Rosa L., SECT. Caninae DC. EM. Christ.). Acta Biol. Crac. Ser. Bot. 2012, 54, 105–112. [Google Scholar] [CrossRef]
- Alp, Ş.; Jurikova, T.; Çakir, Ö.; Gozlekci, S. Bioactive Content of rose hips of different wildly grown Rosa dumalis genotypes. Not. Bot. Horti. Agrobot. 2016, 44, 472–476. [Google Scholar] [CrossRef]
- Kunc, N.; Hudina, M.; Mikulič-Petkovšek, M.; Bavcon, J.; Ravnjak, B.; Osterc, G. Detailed Metabolic Characterization of Flowers and Hips of Rosa gallica L. Grown in Open Nature. Plants 2023, 12, 2979. [Google Scholar] [CrossRef] [PubMed]
- Medveckiene, B.; Kulaitiene, J.; Jariene, E.; Vaitkeviciene, N.; Hallman, E. Carotenoids, polyphenols, and ascorbic acid in organic rosehips (Rosa spp.) cultivated in Lithuania. Appl. Sci. 2020, 10, 5337. [Google Scholar] [CrossRef]
- Turkben, C.; Uylaser, V.; Incedayi, B.; Çelikkol, I. Effects of different maturity periods and processes on nutritional components of rose hip (Rosa canina L.). J. Food Agric. Environ. 2010, 8, 26–30. [Google Scholar]
- Baranski, R.; Baranska, M.; Schulz, H. Changes in carotenoid content and distribution in living plant tissue can be observed and mapped in situ using NIR-FT-Raman spectroscopy. Plants 2005, 222, 448–457. [Google Scholar] [CrossRef]
- Rodriguez-Amaya, D.B.; Kimura, M.; Godoy, H.T.; Amaya-Farfan, J. Updated Brazilian database on food carotenoids: Factors affecting carotenoid composition. J. Food Compos. Anal. 2008, 21, 445–463. [Google Scholar] [CrossRef]
- Rubtsova, O.; Kolesnichenko, O.; Shumyk, M.; Shynder, O.; Chyzhankova, V.; Dzyba, A.; Hrysiuk, S. Scots roses in Northern Ukraine. Hort. Sci. 2021, 48, 144–148. [Google Scholar] [CrossRef]
- Kunc, N.; Hudina, M.; Osterc, G.; Bavcon, J.; Ravnjak, B.; Mikulič-Petkovšek, M. Phenolic compounds of rose hips of some rosa species and their hybrids native grown in the South-West of Slovenia during a two-year period (2020–2021). Foods 2023, 12, 1952. [Google Scholar] [CrossRef]
- Mikulic-Petkovsek, M.; Slatnar, A.; Schmitzer, V.; Stampar, F.; Veberic, R.; Koron, D. Chemical profile of black currant fruit modified by different degree of infection with black currant leaf spot. Sci. Hortic. 2013, 150, 399–409. [Google Scholar] [CrossRef]
- Mikulic-Petkovsek, M.; Stampar, F.; Veberic, R.; Sircelj, H. Wild Prunus fruit species as a rich source of bioactive compounds. J. Food Sci. 2016, 81, C1928–C1937. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species and Cultivars | Content |
|---|---|
| R. gallica | 3.16 ± 0.32 a |
| ‘Violacea’ | 2.17 ± 0.49 a |
| ‘Splendens’ | 6.32 ± 0.77 b |
| R. spinosissima | 3.39 ± 0.44 bc |
| ‘Poppius’ | 5.67 ± 0.49 c |
| ‘Fruhlingsduft’ | 2.57 ± 0.27 ab |
| ‘Single cherry’ | 1.24 ± 0.66 a |
| ‘Fruhlingsmorgen’ | 1.93 ± 0.16 a |
| R. pendulina | 12.36 ± 0.77 c |
| ‘Harstad’ | 12.79 ± 0.44 c |
| ‘Bourgogne’ | 2.38 ± 0.13 a |
| ‘Mount Everest’ | 7.11 ± 0.47 b |
| Species and Cultivars | Citric Acid | Malic Acid | Quinic Acid | Shikimic Acid | Fumaric Acid | Total |
|---|---|---|---|---|---|---|
| R. gallica | 5.14 ± 1.41 b | 2.05 ± 0.85 ab | 10.87 ± 4.50 a | 0.05 ± 0.01 b | 0.01 ± 0.001 a | 18.12 ± 6.77 a |
| ‘Violacea’ | 3.68 ± 1.23 a | 1.49 ± 0.63 a | 8.59 ± 3.01 a | 0.02 ± 0.008 a | 0.008 ± 0.004 b | 13.79 ± 4.88 a |
| ‘Splendens’ | 2.69 ± 0.99 a | 3.28 ± 1.58 b | 9.28 ± 3.42 a | 0.04 ± 0.007 ab | 0.01 ± 0.002 ab | 15.3 ± 5.99 a |
| R. spinosissima | 0.42 ± 0.22 a | 0.25 ± 0.11 a | 0.89 ± 0.34 a | 0.01 ± 0.004 a | 0.002 ± 0.001 a | 1.57 ± 0.68 a |
| ‘Poppius’ | 4.18 ± 1.75 b | 3.34 ± 1.19 b | 16.36 ± 8.29 c | 0.04 ± 0.01 b | 0.008 ± 0.002 b | 23.93 ± 11.24 a |
| ‘Fruhlingsduft’ | 4.23 ± 1.19 b | 3.24 ± 1.27 b | 4.14 ± 2.09 ab | 0.11 ± 0.06 c | 0.007 ± 0.002 b | 8.73 ± 4.61 a |
| ‘Single cherry’ | 2.95 ± 1.17 ab | 1.52 ± 0.56 ab | 4.57 ± 1.62 ab | 0.05 ± 0.03 b | 0.006 ± 0.004 ab | 9.10 ± 3.38 a |
| ‘Fruhlingsmorgen’ | 12.09 ±5.23 c | 6.89 ± 2.17 c | 6.29 ± 3.10 b | 0.18 ± 0.05 d | 0.006 ± 0.001 ab | 25.46 ± 10.55 a |
| R. pendulina | 1.67 ± 0.46 a | 0.66 ± 0.38 a | 21.68 ± 13.53 b | 0.05 ± 0.02 b | 0.003 ± 0.001 a | 24.06 ± 14.39 a |
| ‘Harstad’ | 4.47 ± 2.09 c | 3.31 ± 1.36 c | 26.52 ± 12.97 b | 0.08 ± 0.02 c | 0.008 ± 0.003 b | 34.39 ± 16.44 a |
| ‘Bourgogne’ | 3.75 ± 1.20 c | 2.41 ± 0.92 bc | 9.86 ± 3.46 a | 0.04 ± 0.03 ab | 0.006 ± 0.001 b | 16.07 ± 5.61 a |
| ‘Mount Everest’ | 2.71 ± 1.29 b | 1.75 ± 0.96 ab | 11.43 ± 6.58 a | 0.03 ± 0.01 a | 0.006 ± 0.003 b | 18.64 ± 8.87 a |
| Species and Cultivars | Lutein | Zeaxanthin | Lycopene | α-Carotene | β-Carotene | Total |
|---|---|---|---|---|---|---|
| R. gallica | - | 0.005 ± 0.004 a | 0.66 ± 0.35 ab | 0.16 ± 0.09 a | 25.25 ± 11.73 a | 26.25 ± 12.17 a |
| ‘Violacea’ | 0.88 ± 0.26 b | 0.03 ± 0.01 a | 0.24 ± 0.15 a | 0.20 ± 0.09 a | 36.88 ± 13.09 a | 38.23 ± 13.60 a |
| ‘Splendens’ | 0.55 ± 0.12 a | 0.02 ± 0.01 a | 0.15 ± 0.05 b | 0.18 ± 0.04 a | 17.00 ± 6.61 a | 17.90 ± 6.83 a |
| R. spinosissima | 0.99 ± 0.11 a | 0.05 ± 0.02 b | 0.02 ± 0.01 a | - | 8.20 ± 2.04 a | 9.26 ± 2.18 a |
| ‘Poppius’ | 1.23 ± 0.07 a | 0.05 ± 0.02 b | 0.07 ± 0.04 ab | 0.37 ± 0.10 b | 36.34 ± 12.16 b | 38.06 ± 12.39 a |
| ‘Fruhlingsduft’ | 1.56 ± 0.14 a | 0.01 ± 0.005 a | 0.03 ± 0.02 a | 0.32 ± 0.16 a | 7.04 ± 2.16 a | 8.96 ± 2.49 a |
| ‘Single cherry’ | 0.24 ± 0.11 a | - | 0.13 ± 0.03 b | - | 57.69 ± 21.45 b | 58.06 ± 21.59 a |
| ‘Fruhlingsmorgen’ | 1.55 ± 0.71 a | 0.008 ± 0.003 a | 0.08 ± 0.04 ab | 0.33 ± 0.18 a | 98.87 ± 39.34 c | 100.84 ± 40.27 a |
| R. pendulina | 0.84 ± 0.32 a | 0.04 ± 0.02 a | 4.07 ± 1.77 b | 6.14 ± 2.02 b | 25.68 ± 13.06 b | 36.77 ± 17.19 a |
| ‘Harstad’ | 0.66 ± 0.21 a | 0.02 ± 0.01 a | 4.11 ± 1.64 b | 4.16 ± 1.85 b | 9.52 ± 4.31 a | 18.47 ± 8.02 a |
| ‘Bourgogne’ | 0.66 ± 0.15 a | 0.05 ± 0.02 a | 2.35 ± 0.72 b | 3.86 ± 1.43 b | 22.34 ± 11.04 b | 29.26 ± 13.36 a |
| ‘Mount Everest’ | 2.57 ± 0.32 b | 0.04 ± 0.03 a | 0.11 ± 0.05 a | 0.13 ± 0.07 a | 9.06 ± 3.78 a | 11.91 ± 4.25 a |
| Cultivar | Species of the Origin | Breeding Company | Year of Origin |
|---|---|---|---|
| ‘Violacea’ | R. gallica | unknown (The Netherlands) | 1795 |
| ‘Splendens’ | R. gallica | unknown | 1583 |
| ‘Poppius’ | R. spinosissima | Stenberg (Sweden) | 1872 |
| ‘Fruhlingsduft’ | R. spinosissima | Kordes (Germany) | 1941 |
| ‘Fruhlingsmorgen’ | R. spinosissima | Kordes (Germany) | 1941 |
| ‘Single cherry’ | R. spinosissima | unknown | 1962 |
| ‘Harstad’ | R. pendulina | unknown | unknown (old variety) |
| ‘Mount Everest’ | R. pendulina | Interplant (The Netherlands) | 1956 |
| ‘Bourgogne’ | R. pendulina | Interplant (The Netherlands) | 1983 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kunc, N.; Hudina, M.; Mikulic-Petkovsek, M.; Osterc, G. Breeding of Modern Rose Cultivars Decreases the Content of Important Biochemical Compounds in Rose Hips. Plants 2023, 12, 3734. https://doi.org/10.3390/plants12213734
Kunc N, Hudina M, Mikulic-Petkovsek M, Osterc G. Breeding of Modern Rose Cultivars Decreases the Content of Important Biochemical Compounds in Rose Hips. Plants. 2023; 12(21):3734. https://doi.org/10.3390/plants12213734
Chicago/Turabian StyleKunc, Nina, Metka Hudina, Maja Mikulic-Petkovsek, and Gregor Osterc. 2023. "Breeding of Modern Rose Cultivars Decreases the Content of Important Biochemical Compounds in Rose Hips" Plants 12, no. 21: 3734. https://doi.org/10.3390/plants12213734
APA StyleKunc, N., Hudina, M., Mikulic-Petkovsek, M., & Osterc, G. (2023). Breeding of Modern Rose Cultivars Decreases the Content of Important Biochemical Compounds in Rose Hips. Plants, 12(21), 3734. https://doi.org/10.3390/plants12213734