Abstract
Bacterial canker caused by Pseudomonas syringae pv. syringae (Pss) is responsible for substantial loss to the production of sweet cherry in Chile. To date, the molecular mechanisms of the Pss–sweet cherry interaction and the disease-related genes in the plant are poorly understood. In order to gain insight into these aspects, a transcriptomic analysis of the sweet cherry cultivar ‘Lapins’ for differentially expressed genes (DEGs) in response to Pss inoculation was conducted. Three Pss strains, A1M3, A1M197, and 11116_b1, were inoculated in young twigs, and RNA was extracted from tissue samples at the inoculation site and distal sections. RNA sequencing and transcriptomic expression analysis revealed that the three strains induced different patterns of responses in local and distal tissues. In the local tissues, A1M3 triggered a much more extensive response than the other two strains, enriching DEGs especially involved in photosynthesis. In the distal tissues, the three strains triggered a comparable extent of responses, among which 11116_b1 induced a group of DEGs involved in defense responses. Furthermore, tissues from various inoculations exhibited an enrichment of DEGs related to carbohydrate metabolism, terpene metabolism, and cell wall biogenesis. This study opened doors to future research on the Pss–sweet cherry interaction, immunity responses, and disease control.
1. Introduction
Sweet cherry (Prunus avium L.) ranks first in fruit tree cultivation and fruit production in Chile [1]. During the 2021–2022 season, a total of 259,202.5 tons of fruit were exported to the international market, making Chile the top exporting country in the world [2]. With the increasing cultivation, diseases have emerged, devastating the crop trees and impairing fruit production. Among these diseases, bacterial canker has become a main cause of yield loss [3]. The pathovars P. syringae pv. syringae and P. amygdali pv. morsprunorum have been isolated from the symptomatic trees and confirmed to be the causal pathogens [4,5]. Between them, P. syringae pv. syringae (Pss) is the major cause of the disease, resulting in up to 40% of tree losses [6].
To control the disease and minimize yield loss, studies have been conducted to comprehend the interactions between the pathogen and the host during the development of bacterial canker disease. So far, most research in this field has focused on the pathogenicity of P. syringae pvs., leaving a significant knowledge gap concerning disease-related genes in the host. A similar lack of knowledge is also evident in other Prunus species affected by bacterial canker, even though phenotype screenings have identified cultivars displaying differential susceptibilities to the disease [7,8,9,10]. In a recent study involving apricot (P. armeniaca L.), genome-wide single-nucleotide polymorphism markers revealed associations between seven chromosomal loci and plant resistance to Pss [11]. However, no candidate genes for resistance to bacterial canker were identified within these loci.
The identification of disease-related genes in sweet cherry necessitates whole-genome sequencing and gene annotation. Currently, GenBank provides access to the genomes of four P. avium cultivars: ‘Satonishiki’, the leading variety in Japan [12]; ‘Big Star’, a self-crossing progeny of ‘Lapins’ generated by the University of Bologna [13,14]; ‘Tieton’, a hybrid of ‘Stella’ and ‘Early Burlat’ created by Washington State University [15]; and ‘Hongmanao’, the most extensively cultivated sweet cherry variety in north China (GenBank BioProject PRJNA489346). Gene prediction and annotation have been performed on ‘Satonishiki’ and ‘Tieton’, the former being available in GenBank. Additionally, a basic transcriptome analysis of three cultivars, ‘Bing’, ‘Lapins’ and ‘Rainier’, has been conducted using the annotation data [16].
This study presents the first comparative transcriptome analysis of sweet cherry infected with P. syringae pv. syringae (Pss). Three Pss strains of varying virulence were used to inoculate sweet cherry through wounding, and RNA was extracted from the tissue near and away from the wound. RNA sequencing revealed differentially expressed genes post-infection with each strain, as well as the genes induced by two or three individual strains. Gene ontology enrichment analysis unveiled unique and shared responses elicited by the three strains. These datasets address the scarcity of disease-related gene candidates in sweet cherry and open the way for extensive research into the defense responses of sweet cherry against this bacterium.
2. Results
2.1. Disease Development
Three Pss strains, A1M3, A1M197, and 11116_b1, were inoculated into twigs of sweet cherry cv. ‘Lapins’. Forty days after Pss inoculation, wounds with either an opening or filled irregular cork structure with gum, typical of bacterial canker, were observed for all three strains. By contrast, the mock-treated twigs displayed a closed wound covered with compact cork, giving it a clean appearance (Supplementary Figure S1A). Lesions induced by Pss were observed up to 5 cm around the inoculation zone. To confirm the effectiveness of Pss infection in the twigs, the presence of viable Pss in the local samples near the wound was assessed (Supplementary Figure S1A). The colonies analyzed from the infected twigs exhibited UV-induced fluorescence with a LOPAT profile consistent with Pss [17] (Supplementary Figure S1B). DNA was extracted from both local and distal samples, and Pss DNA was quantified using Real-time PCR (Supplementary Table S1, Supplementary Figure S1C,D). In the local samples, Pss was detected in all Pss-inoculated twigs, whereas in the mock-treated samples, only one sample tested positive for Pss, with an amount of bacterial DNA lower than that of the Pss-inoculated samples (Supplementary Table S2). The A1M197-inoculated samples exhibited the highest Pss DNA content, nearly 50 times higher than that of the A1M3-inoculated samples. The 11116_b1-inoculated samples presented an amount of bacterial DNA that ranged between the values of the other two samples. Intriguingly, Pss was also detected in one of the three replicates of the distal samples from both A1M197- and A1M3-inoculated twigs (Supplementary Table S2). These results confirmed that the three Pss strains were capable of establishing in the cv. ‘Lapins’ twigs around the inoculation zone, and their infection was effective 40 days after inoculation.
2.2. The Transcriptome Datasets of Sweet Cherry cv. ‘Lapins’
The RNA sequencing of each of the 24 samples generated 53,215,158 to 69,961,046 raw reads, each with a length of 151 nt (Table 1). Within each dataset, 92.03% to 96.77% of reads reached the quality of Q20, and 86.79% to 92.50% of reads reached the quality of Q30. After quality trimming, 46,916,794 to 66,028,852 trimmed reads were obtained, accounting for 85.89% to 95.40% of the raw reads and resulting in average read lengths of 126.38 to 134.25 nt.

Table 1.
Statistics of the Prunus avium cv. Lapins transcriptome datasets.
For 22 out of 24 datasets, 21.66% to 39.59% of the trimmed reads were mapped to the reference transcriptome of Prunus avium cv. ‘Satonishiki’, covering 67.01% to 75.29% of the total of 35,009 transcripts. Two datasets corresponding to an A1M3-inoculated local sample and an A1M3-inoculated distal sample exhibited significantly lower percentage of mapped reads compared to the others, with values of 6.64% and 9.18%, respectively. However, the low percentage was not related to the quality of the reads.
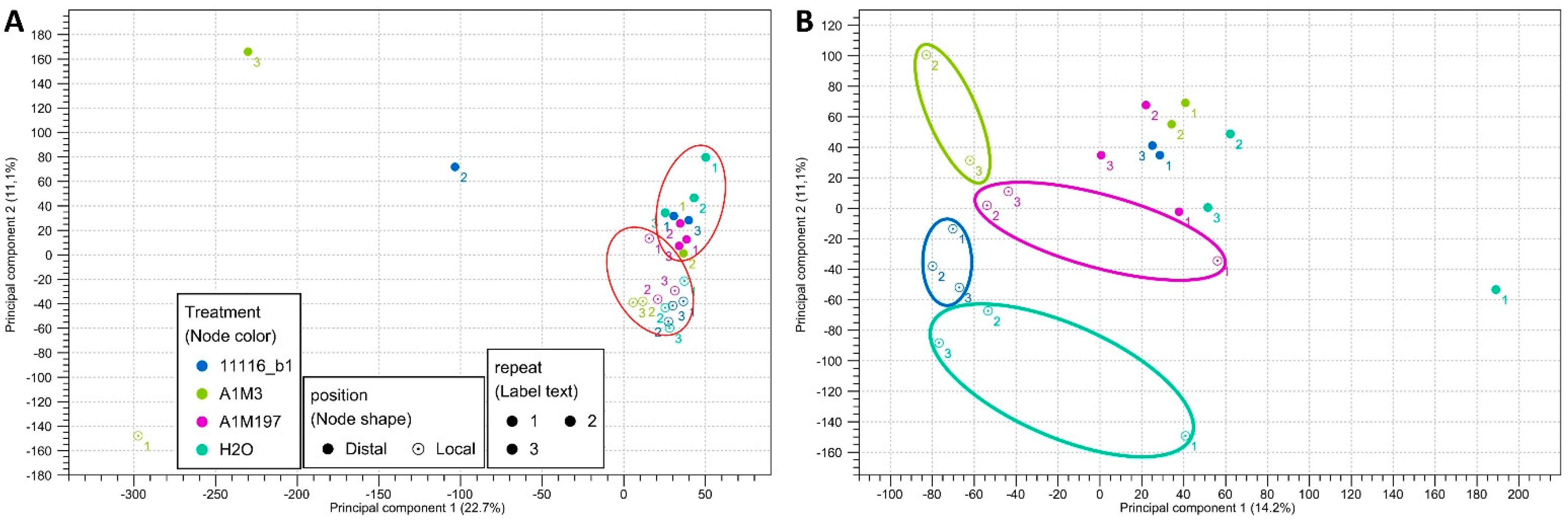
Principal component analysis (PCA) of the mapped transcripts from all the datasets revealed two clusters with three outliers (Figure 1A, Supplementary Figure S2). The two clusters included 11 local samples and 10 distal samples, respectively, indicating that they exhibited similar expression profiles for the majority of the transcripts. The three outliers corresponded to the datasets with the shortest average read lengths after trimming and the lowest percentages of reads mapped to the reference transcriptome (Table 1). This observation suggests that the difference in clustering might be attributed to the length and number of mapped reads. In order to further separate the clusters, the outliers were removed, and the remaining 21 datasets were reanalyzed. The new scatter plot revealed that, for the local samples, the datasets clustered distinctly based on different treatments, whereas for the distal samples, the datasets from various treatments did not exhibit significant clustering patterns (Figure 1B). These findings imply that inoculation with different Pss strains resulted in more diverse local responses. However, removing the three outliers did not alter the results of the expression analysis; therefore, they were retained for the subsequent analyses.
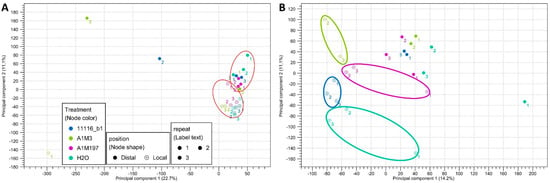
Figure 1.
Principal component scatter plot (2D) of the transcriptome datasets from (A) the 24 samples of sweet cherry inoculated with three Pss strains and (B) the 21 samples circled in (A). The red circles in (A) indicate the clustering of local and distal datasets. The four circles in (B) indicate the clustering of datasets from different treatments.
2.3. Different Pss Strains Trigger Different Local and Distal Responses
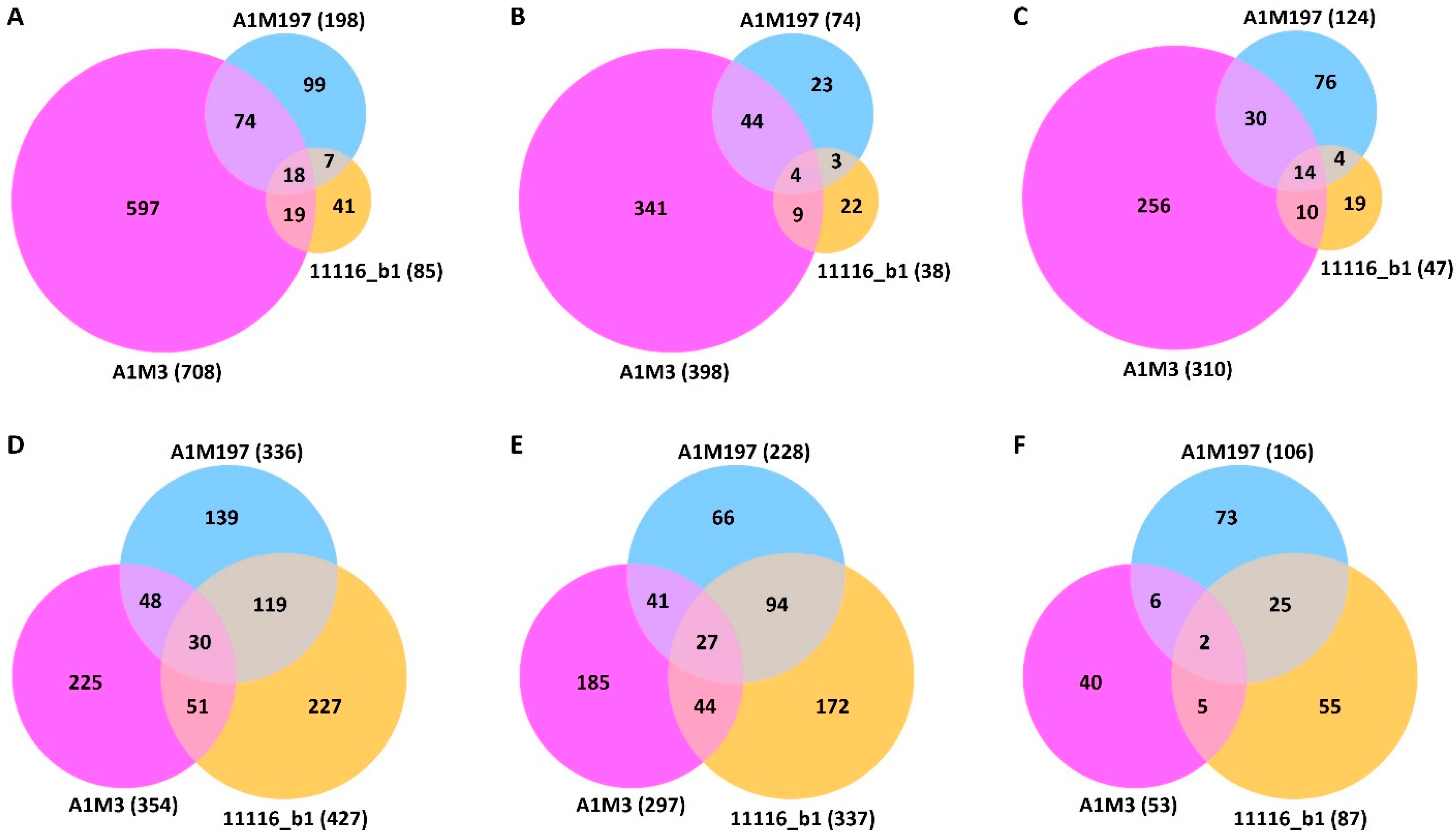
Differential expression analysis revealed that the three Pss strains elicited different local and distal responses in sweet cherry (Supplementary Figure S3). In the local samples, A1M3 inoculation caused the differential expression of 708 genes (Supplementary Table S3, Figure 2A). In contrast, A1M197 caused the differential expression of 198 genes, whereas 11116_b1 only caused the differential expression of 85 genes (Supplementary Tables S4 and S5, Figure 2A). This difference indicates that A1M3 triggered a more significant local response in sweet cherry than the other two Pss strains. A1M3 and A1M197 inoculations shared 92 DEGs, accounting for 46.5% of the total DEGs caused by A1M197 but only 13.0% of that of A1M3 (Figure 2A). Similarly, A1M3 and 11116_b1 inoculations shared 37 DEGs, representing 43.5% of the total DEGs caused by 11116_b1 but only 5.2% of that of A1M3 (Figure 2A). In the distal samples, however, inoculation with A1M3, A1M197, and 11116_b1 caused the differential expression of 354, 336, and 427 genes, respectively (Supplementary Tables S6–S8, Figure 2D), suggesting that the three strains triggered a comparable extent of distal response in the plant.
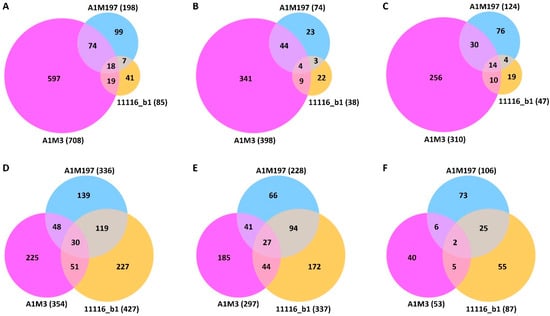
Figure 2.
Venn diagrams showing differentially expressed sweet cherry genes post-inoculation of the three Pss strains. (A–C) Local samples. (D–F) Distal samples. (A,D) show the total numbers of DEG. (B,E) show the numbers of up-regulated genes. (C,F) show the numbers of down-regulated genes.
The ratios of up- and down-regulated genes in response to the inoculation showed different patterns in the local and distal datasets. In the local samples, the numbers of up- and down-regulated genes were comparable: 398 and 310 for A1M3, 73 and 125 for A1M197, and 38 and 47 for 11116_b1, respectively (Figure 2B,C, Supplementary Tables S3–S5). By contrast, in the distal samples, the number of up-regulated genes was significantly higher than the number of down-regulated genes for all three Pss strains, with the former being two to six times higher than the latter: 297 and 53 for A1M3, 228 and 106 for A1M197, and 337 and 87 for 11116_b1, respectively (Figure 2E,F, Supplementary Tables S6–S8). These data suggest that the response to the inoculation in the distal tissue could be more specific to disease resistance than in the local tissue.
Four DEGs included in Figure 2D were not presented in Figure 2E,F due to their different responses in different treatments (Supplementary Tables S6–S8; Figure 3A). XM_021971224.1, which encodes a homolog of an Arabidopsis G-type lectin S-receptor-like serine/threonine-protein kinase, was up-regulated by A1M3 inoculation but down-regulated by A1M197 inoculation. XM_021954392.1, which encodes a probable aquaporin, and XM_021974777.1, which encodes a probable L-cysteine desulfhydrase in the chloroplast, were up-regulated by A1M3 inoculation but down-regulated by 11116_b1 inoculation. XM_021953649.1, which encodes a (3S,6E)-nerolidol synthase 1-like protein, was up-regulated by A1M3 inoculation but down-regulated by A1M197 and 11116_b1 inoculation.
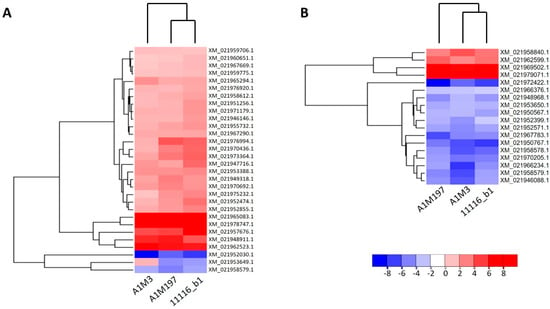
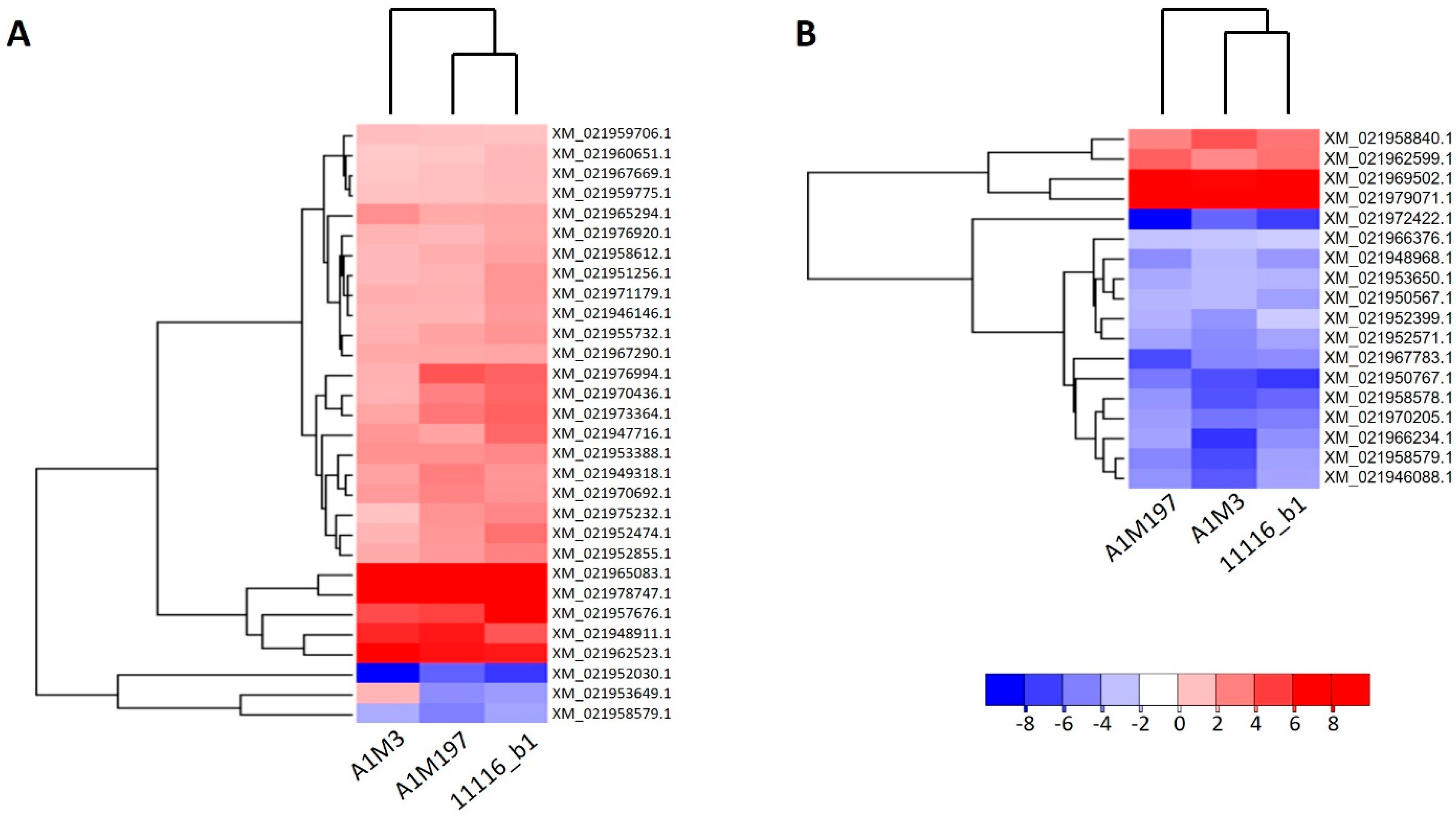
Figure 3.
Heat maps showing the expression patterns of the DEGs after inoculation of all three Pss strains in sweet cherry. (A) Distal samples. (B) Local samples. Genes and datasets are clustered in hierarchical order according to their average Euclidean distances. The color gradient represents the log2 fold change in gene expression.
The fold change in genes also varied among the inoculations with the three Pss strains. In the local samples from A1M3 inoculation, DEGs that were regulated by 2- to 16-fold constituted over 80% of all DEGs, whereas the percentages for A1M197 and 11116_b1 were significantly lower (Supplementary Figure S4). This suggests that the local response triggered by A1M3 was more extensive, rather than stronger, than that of the other two strains. On the other hand, genes up- or down-regulated more than 256-fold accounted for a small percentage of the total DEGs from all three inoculations (8.3%, 7.6%, and 5.9% for A1M3, A1M197, and 11116_b1, respectively; Supplementary Figure S4). Notably, two genes, XM_021969502.1 and XM_021979071.1, were up-regulated more than 184-fold in response to all three inoculations (Figure 3B). XM_021969502.1 encoded an isoform of E3 ubiquitin-protein ligase PRT6, whereas XM_021979071.1 encoded an isoform of golgin subfamily A member 4 (Supplementary Tables S3–S5).
In the distal samples from A1M3 inoculation, a higher percentage of DEGs were up-regulated over 16-fold (Supplementary Figure S4), suggesting a stronger response to the pathogen. In contrast, the percentages of DEGs up-regulated over 16-fold in the distal samples from A1M197 and 11116_b1 inoculations were smaller than those in the corresponding local samples, indicating different patterns of response to these three strains.
2.4. Functional Categories and GO Enrichment Analysis
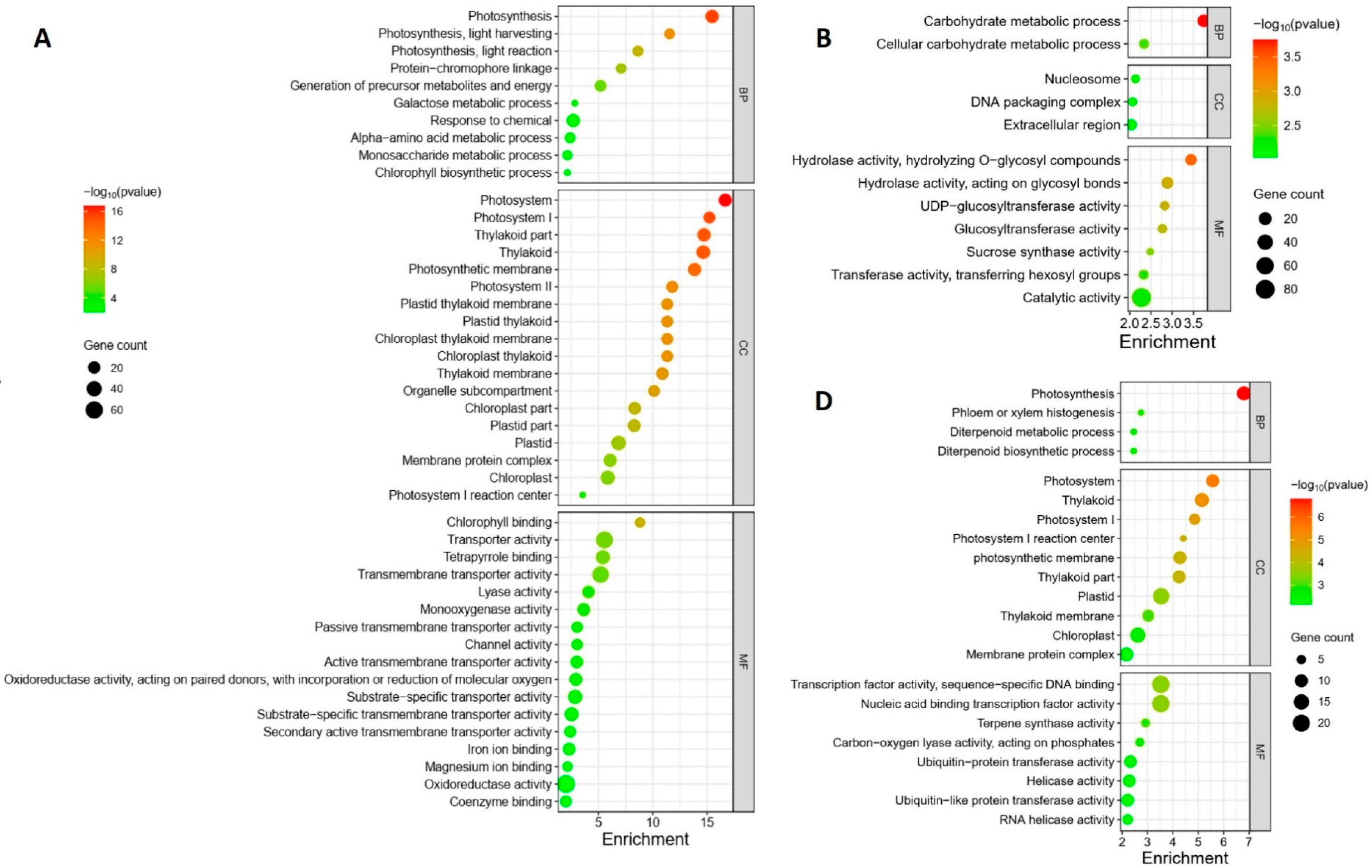
The GO enrichment analysis revealed distinct regulation patterns in response to different Pss inoculations. The local samples from plants inoculated with the three Pss strains exhibited unique enrichment profiles; there was no overlap among the enrichment patterns of the three inoculations except for the term “Magnesium ion binding”, which was shared by A1M3 and 11116_b1 (Figure 4A–C). Specifically, in the samples from A1M3 inoculation, the most highly enriched GO terms were related to chloroplast structure and photosynthesis (Figure 4A); in the samples from A1M197 inoculation, the most highly enriched GO terms were associated with carbohydrate metabolism (Figure 4B); and in the samples from 11116_b1 inoculation, the most highly enriched GO terms were linked to terpene metabolism (Figure 4C). Notably, all the enriched DEGs in the three categories were down-regulated, indicating that infection with A1M3, A1M197, and 1111b_b1 could locally hinder the normal processes of photosynthesis, carbohydrate metabolism, and terpene metabolism, respectively, in sweet cherry. Additionally, A1M3 inoculation also caused the enrichment of DEGs encoding a group of transporters and oxidoreductase (Figure 4C), suggesting that this Pss strain might interfere with multiple aspects of cellular metabolism as well as the redox state of the cell.
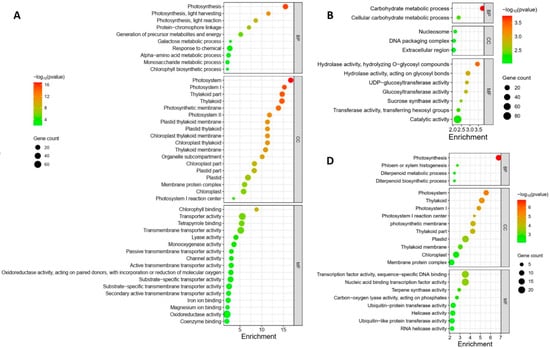
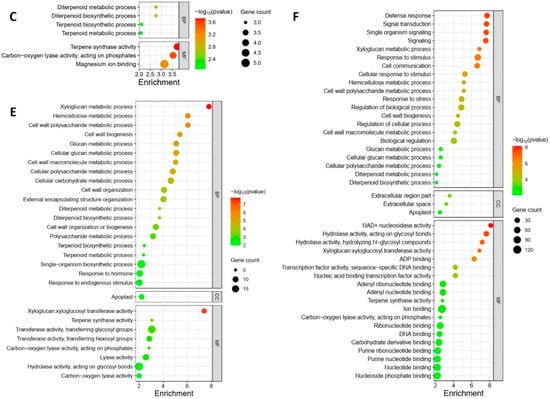
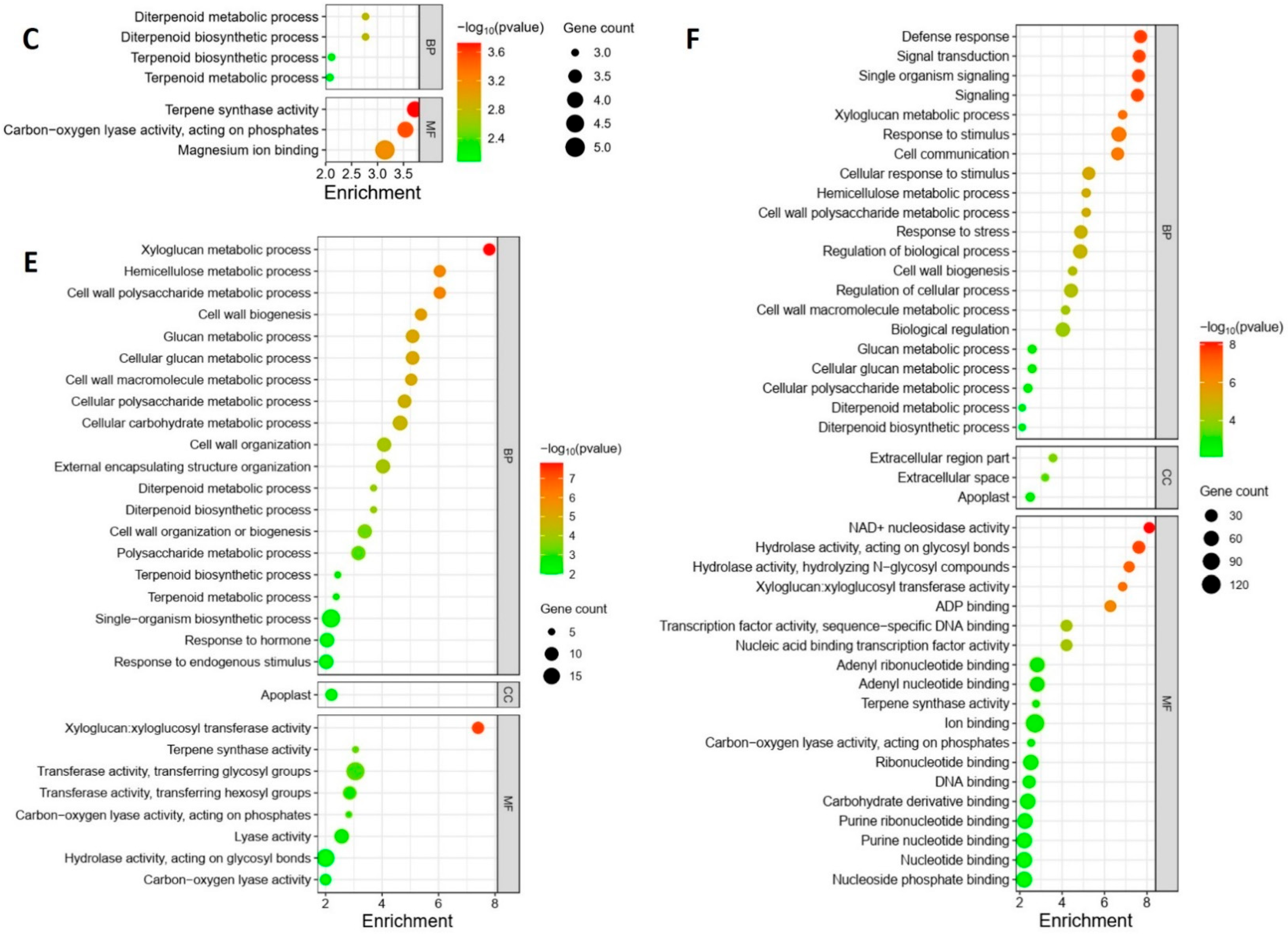
Figure 4.
GO enrichment of DEGs in response to Pss inoculation. (A) Local samples inoculated with A1M3. (B) Local samples inoculated with A1M197. (C) Local samples inoculated with 11116_b1. (D) Distal samples inoculated with A1M3. (E) Distal samples inoculated with A1M197. (F) Distal samples inoculated with 11116_b1. Y axis, GO pathways. X axis, enrichment factors. BP, biological process. CC, cellular component. MF, molecular function.
The distal samples from plants inoculated with the three Pss strains showed more complicated enrichment patterns. On the one hand, the samples from A1M197 and 11116_b1 inoculations exhibited similarities in highly enriched GO terms, including “xyloglucan metabolic process”, “glucan metabolic process”, “cell wall polysaccharide metabolic process”, “cell wall biogenesis”, “hemicellulose metabolic process”, “cell wall macromolecule metabolic process”, and “xyloglucan: xyloglucosyl transferase activity” (Figure 4E,F). All the DEGs in these categories were up-regulated, suggesting that A1M197 and 11116_b1 infection could promote primary cell wall biogenesis in the distal tissues of sweet cherry. By contrast, in the samples from A1M3 inoculation, the highly enriched GO terms were related to photosynthesis (Figure 4D), and all the enriched DEGs in this category were down-regulated. This feature, together with Figure 4A, suggests that A1M3 infection could interfere with photosynthesis both locally and distally in sweet cherry. On the other hand, all three Pss strains caused the enrichment of the GO terms “terpene synthase activity” and “carbon-oxygen lyase activity” (Figure 4D–F), suggesting a certain extent of similar distal response triggered by the three strains. Most intriguingly, the GO term “defense response” was highly enriched in the samples from 11116_b1 inoculation, but not for the other two strains (Figure 4F). Also enriched for 11116_b1 was a group of GO terms related to ribonucleotide binding and transcription factor activity. This peculiar feature suggests that 11116_b1 could trigger defense mechanisms in the distal tissues of sweet cherry.
2.5. Validation of Expression Changes by Quantitative RT-PCR
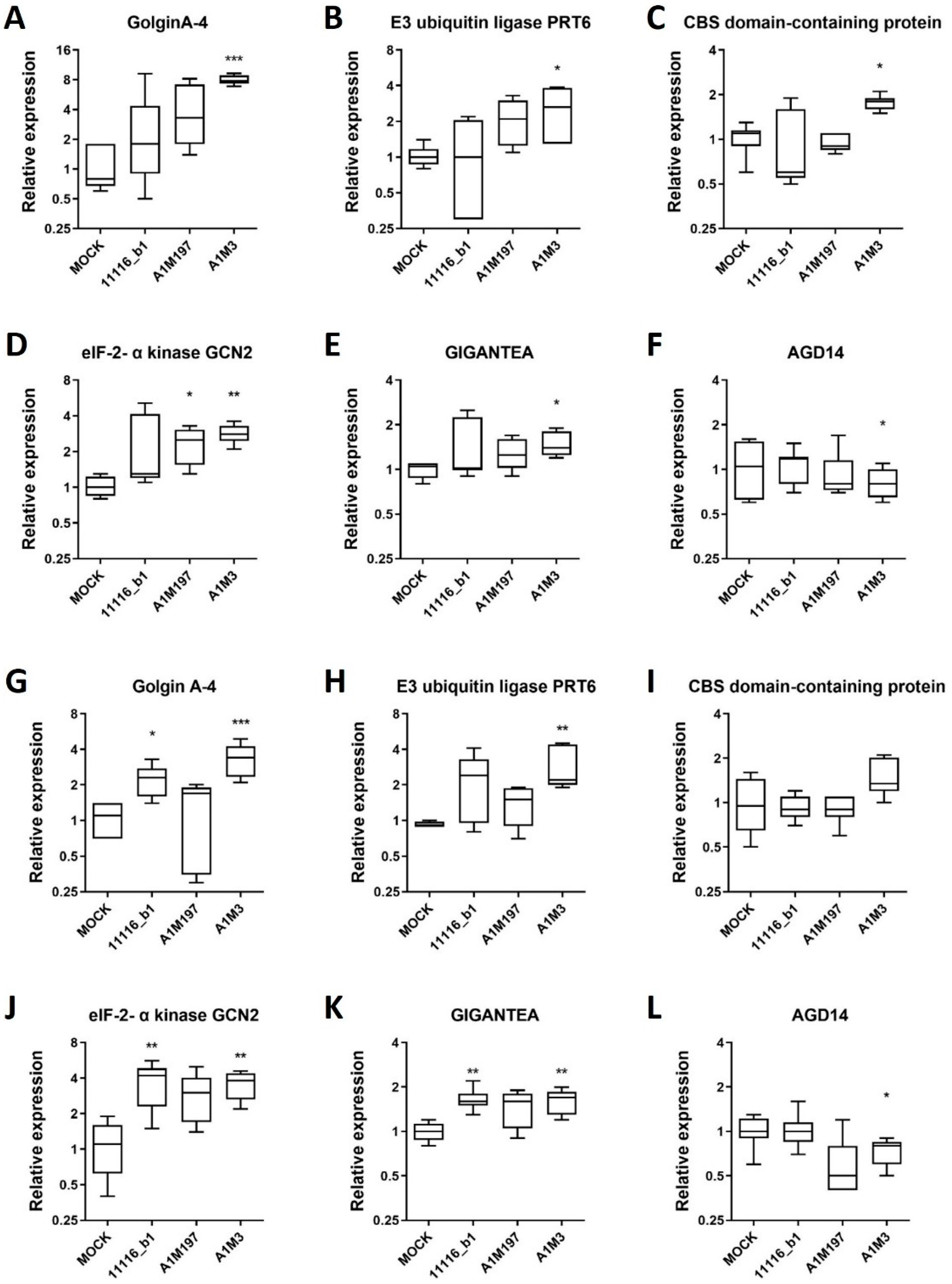
To validate the expression profile of RNA-seq data, six genes were selected by comparing their transcript levels in samples infected with each Pss strain versus the Mock control using RNA-seq data (Supplementary Tables S9 and S10). Specifically, three genes (GolginA-4, XM_021979071.1; E3 ubiquitin-protein ligase PRT6, XM_021969502.1; and CBS domain-containing protein, XM_021962599.1) were significantly up-regulated in local samples inoculated with each of the three Pss strains. Two genes (eIF-2-alpha kinase GCN2, XM_021957676.1 and GIGANTEA, XM_021962523.1) were significantly up-regulated in the distal samples infected with each Pss strain, and one gene (AGD14, XM_021952030.1) was significantly down-regulated in all the distal Pss-infected samples.
The expression of these genes was independently verified in the local and distal samples previously inoculated with each Pss strain by qPCR. In the local samples, GolginA-4 showed an increase in transcript level in samples infected with all three Pss strains compared to the mock control, but only the A1M3 strain-infected samples showed a significant increase (Figure 5A). Similarly, the transcript levels of E3 ubiquitin-protein ligase PRT6 and CBS domain-containing protein were significantly increased in A1M3-infected samples (Figure 5B,C). In the distal samples, the selected gene eIF-2-alpha kinase GCN2 presented increased transcript levels for all three Pss strains, whereas only A1M197- and A1M3-infected samples showed a significant increase compared to the mock condition (Figure 5D). GIGANTEA transcript levels were augmented, whereas AGD14 was down-regulated in A1M3-infected samples (Figure 5E,F).
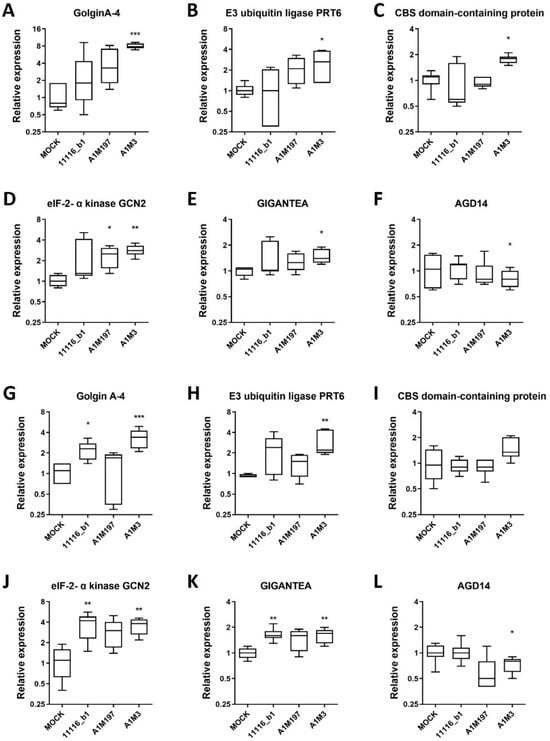
Figure 5.
Expression profile of selected DEGs in local and distal samples from plants inoculated with Pss. Relative expression analysis of six genes from local (A–F) and distal (G–L) samples inoculated with 11116_b1, A1M197, and A1M3 Pss strains in sweet cherry twigs. The expression of these selected genes was measured by qPCR and normalized to the mock condition. Peptidyl-prolyl cis-trans isomerase CYP21-1 and splicing factor 3B subunit 1 were used as reference genes. Kruskal–Wallis test with Dunn’s multiple comparisons test between inoculated and mock conditions was performed: ***, p < 0.0005; **, p < 0.005; *, p < 0.05; n = 3 (three independent biological replicates).
Interestingly, GolginA-4 and E3 ubiquitin-protein ligase PRT6 also showed an increase in transcript level in the distal samples for all three Pss strains compared to the mock control. Although GolginA-4 levels were significantly increased in 11116_b1-infected samples, both transcript levels were increased in A1M3-infected samples (Figure 5G,H). However, the changes in the transcript levels of the genes GIGANTEA, GolginA-4, and E3 ubiquitin-protein ligase PRT6 were less pronounced in the distal samples than in the local samples. Additionally, eIF-2-alpha kinase GCN2 and GIGANTEA genes were up-regulated in the distal samples from all three Pss-infected twigs (Figure 5J,K). In the case of AGD14, down-regulation in A1M3-infected samples was detected as expected (Figure 5L).
Moreover, qPCR data from the selected genes and Actin7, a housekeeping gene, in local or distal infected samples were linearly, positively, and strongly correlated with RNA-seq data from the same genes (r = 0.7414; p < 0.0001; Supplementary Figure S5). These results are consistent with the observed trend in RNA-seq data, validating the expression change analysis in local and distal Pss-infected conditions.
3. Discussion
3.1. Disease-Related Gene Candidates in Sweet Cherry Induced by Pss Inoculation
Heretofore, very little is understood about the molecular mechanism of defense responses in sweet cherry. Through conducting a comparative transcriptome analysis, a list of DEGs in response to Pss infection was identified, among which potential disease resistance genes were discovered. The GO enrichment analysis revealed that 33 DEGs caused by 11116_b1 inoculation were related to the defense response (Figure 4F), including XM_021974586.1, which encodes an RPM1-like protein; XM_021947717.1, which encodes an RPS6-like protein; XM_021977557.1, which encodes a TAO1-like protein; XM_021978626.1, which encodes an NDR1-like protein; XM_021961241.1, which encodes an SRC2-like protein; XM_021948923.1, which encodes an RPP13-like protein; XM_021976406.1, which encodes RGA3; XM_021976429.1, which encodes RGA4; XM_021969778.1, which encodes SNC1; XM_021977313.1, which encodes nematode resistance protein-like HSPRO2; XM_021953798.1, which encodes a homolog of probable disease resistance protein At5g66900 in Arabidopsis; XM_021946277.1, which encodes an uncharacterized protein; XM_021961188.1, which encodes NDR1/HIN1-like protein 3; XM_021961402.1 and XM_021961341.1, both of which encode BAP2-like proteins; and 18 genes encoding TMV resistance protein N-like proteins. With the exception of the uncharacterized protein, homologs of all these proteins have been studied in other plant systems [18,19,20,21,22,23,24,25,26,27,28,29], providing a starting point for elucidating the defense network in sweet cherry.
Interestingly, initial experiments on sweet cherry leaves revealed that 11116_b1 displayed the highest level of virulence when inoculated into leaf tissue (Supplementary Figure S6). Therefore, this strain exhibited markedly distinct behaviors in local and distal tissues. In the local, inoculated tissues, it demonstrated the greatest virulence while triggering the fewest DEGs (Figure 2A). However, in the distal tissues, which were not in direct contact with the bacteria, it induced robust resistance responses, likely mediated through long-distance signal transduction pathways. All three Pss strains belong to subgroup 2d of P. syringae phylogroup 2; however, they produce different collections of type III secreted effectors (T3SEs) [30]. It can be speculated that the specific T3SEs of 11116_b1 contributed to the high virulence of this strain in the local tissue by suppressing plant resistance responses. On the other hand, signals specifically triggered by these effectors could travel to distal tissues, where they induced a cohort of resistance genes. Understanding the interaction between the T3SEs and the resistance genes would aid in protecting sweet cherry against pathogens. For instance, this knowledge could be utilized to trigger resistance genes and thwart Pss infection.
Notably, TAO1 was also up-regulated 727- and 1390-fold in the local and distal samples from sweet cherry inoculated with A1M3 (Supplementary Tables S3 and S6), despite no enrichment of defense-related DEGs being observed. TAO1 was found to contribute to disease resistance induced by AvrB, an avirulence factor of P. syringae pv. tomato DC3000, in Arabidopsis [26]. The up-regulation of TAO1 by A1M3 suggests that this TIR-NBS-LRR disease resistance protein might play a role outside of the known defense pathway. Another noteworthy finding is that two genes encoding EDR2-like proteins, XM_021963532.1 and XM_021953322.1, were up-regulated 31- and 30-fold in distal samples from sweet cherry inoculated with A1M3 and 11116_b1, respectively. EDR2 is a negative regulator of salicylic acid-based defense in Arabidopsis, suppressing the hypersensitive response triggered by avirulent pathogens [31]. The up-regulation of EDR2 by A1M3 infection also suggests its unknown function in the plant–pathogen interaction.
3.2. Pss Infection Triggers a Variety of Biological Processes
Monoterpenes, including α-pinene and β-pinene, have been reported to play multiple roles in plant defense responses [32,33]. Both are produced at the pathogen infection site and could activate the salicylic acid (SA) signaling pathway, initiating systemic acquired resistance (SAR). Additionally, both are volatile and could induce defense responses in neighboring plants. This study revealed the down-regulation of genes encoding a group of (-)-α-pinene synthase-like proteins in both local samples from 11116_b1 inoculation and distal samples from all three inoculations (Figure 4C–F, Supplementary Tables S5–S8). These results suggest that 11116_b1 could suppress the production of α-pinene at the infection site, inhibiting the activation of the SA signaling pathway and SAR. It may also block the interplant priming by volatility. Moreover, all three Pss strains could suppress α-pinene production in distal tissues, possibly blocking the systemic signal.
Chloroplasts play a pivotal role in plant defense by generating a burst of reactive oxygen species (ROS) upon the recognition of pathogen-associated molecular patterns (PAMPs), which triggers the basal immune program known as PAMP-triggered immunity (PTI) [34]. Successful pathogens may secrete effectors that reduce the production of ROS or enhance their scavenging, thereby suppressing the ROS burst and PTI [34,35]. This study showed that A1M3 infection caused the down-regulation of a large number of genes involved in chloroplast structure and photosynthesis (Figure 4A,C). Given that A1M3 is the least virulent strain among the three (Supplementary Figure S6), it is likely that upon the infection of this strain, the regular function of chloroplasts was diverged to produce more ROS, leading to a ROS burst and subsequent immune response in sweet cherry, whereas the other two strains could suppress the ROS burst without down-regulating chloroplast-related genes. This speculation is supported by the enrichment of GO terms related to oxidoreductase activity in the local samples from A1M3 inoculation (Figure 4A).
One peculiar feature of the distal samples from A1M197 and 11116_b1 inoculations was the enrichment of GO terms related to primary cell wall biogenesis, especially hemicellulose metabolism (Figure 4E,F). The cell wall plays multiple roles in plant defense. At the infection site, it serves as a physical barrier against pathogens and as a source of signaling molecules [36]. On the other hand, systemic signals such as SA lead to the strengthening of the cell wall by depositing secondary components such as callose and lignin [37]. The altered metabolism of hemicellulose, a component of the primary cell wall, in the tissues distant from the infection site has not been previously reported. However, it could be speculated that certain signals triggered by the infection of A1M197 and 11116_b1 had travelled systemically to induce such a response in non-infected parts of the plant. Increased deposition of hemicellulose might strengthen the cell wall, albeit not as efficient as callose and lignin. It would be interesting to identify the signal and to study the function of hemicellulose in SAR.
Pathogen infection breaks the homeostasis of the host, forcing the plant to redistribute its resources to balance growth and defense [38]. To achieve the redistribution, contents of soluble carbohydrates such as glucose and sucrose are elevated, reflecting the up-regulation of related enzyme activity [39]. Interestingly, infection of A1M197 caused the down-regulation of genes involved in carbohydrate metabolism at the inoculation site, especially those encoding glucosyl hydrolases, glucosyltransferases, and sucrose synthases (Figure 4B, Supplemental Table S4), suggesting a reduced turnover rate of sucrose and other cellular carbohydrates. One possible explanation for this unexpected result might be that, at 40 days post-inoculation, with the establishment of bacterial population, carbohydrate synthesis was down-regulated to prevent the pathogen from exploiting its resources. This unique effect of A1M197, however, is worth exploring further.
3.3. Detection of Pss in Uninfected Samples
Using primers for the glutathione-dependent formaldehyde-activating enzyme 1 gene, Pss presence was detected in one mock-treated local sample and two distal samples (Supplementary Table S2). The primer pair was specifically designed to target a conserved sequence unique to Pss, ensuring that the positive result was not caused by the existence of other known Pseudomonas species or other P. syringae pathovars. The positive result, therefore, could be due to either experimental contamination of Pss or the existence of unknown Pseudomonas species. All the mock-treated twigs showed a healthy appearance at the time of sampling, and all the distal sections were kept intact and showed no symptoms at the time of sampling, indicating that Pss contamination, if it ever existed, did not cause infection and did not affect the results. On the other hand, there are Pseudomonas species existing in the environment and as endophytes [40] that might interfere with the detection Pss. Moreover, although most P. syringae strains are known as phytopathogens, some can exist in the environment without causing disease [41]. The Pss detected in the non-infected samples in this study could be a false positive result caused by other Pseudomonas species.
4. Materials and Methods
4.1. Plant Material and Growth Conditions
Two-year-old sweet cherry cv. ‘Lapins’ trees were purchased at a commercial plant nursery (34°28′12.0″ S 70°58′48.0″ W). These trees were transplanted to 20 L plastic pots filled with a mixture of peat moss, compost, and perlite (1:1:1, v/v/v) as substrate and supplemented with Basacote® plus 6M (Combo expert, 3 g/L substrate) at dormant stage. Trees were acclimated into a side-open shade house placed in the experimental station at the Instituto de Ciencias Agroalimentarias Animales y Ambientales (ICA3) of the Universidad de O’Higgins, San Fernando, Chile (34°36′36.0″ S 70°59′24.0″ W). Each tree was irrigated twice per day by an automatic drip irrigation system until full pot capacity (approximately 0.85 L per irrigation event).
4.2. Bacterial Inoculation, Plant Sampling, Bacterial Re-Isolation, and Biochemical Characterization
Three Pss strains isolated from commercial sweet cherry orchards, A1M3, A1M197 [42], and 11116_b1 (this report), were grown for 20 h at 26 °C on KB agar medium, and the colonies were resuspended in sterile saline buffer (0.8% NaCl, 0.04% NaH2PO4 and 0.27% Na2HPO4), adjusted to 2 × 108 CFU/mL (OD600 = 0.03). Six plants were inoculated per Pss strain, one twig per plant. Actively growing summer twigs were wounded at 10 cm from the apex with a sterile scalpel. The wound was immediately inoculated with 50 µL of one of the Pss suspensions or a sterile saline buffer as mock control, and immediately covered with sterile glycerol and Parafilm. Plants were maintained in the same shade house with the same irrigation as the acclimation period throughout the assay. Forty days after inoculation, twigs were cut at 30 cm from the apex, and disease symptoms were inspected around the wound. Green tissue beneath the epidermis was sampled (50–100 mg) next to the wound and 15–20 cm below the point of Pss inoculation for each inoculated twig. These samples were named local and distal samples, respectively.
Local samples were macerated in a sterile saline buffer, streaked on KB agar medium and incubated at 26 °C for 20 h. Two isolated colonies per sample were evaluated by LOPAT test [43] with minimum changes and by UV-induced fluorescence [17]. Oxidase assay was tested on oxidase strips (Sigma-Aldrich, Burlington, MA, USA) following the manufacturer’s instructions.
4.3. Nucleic Acid Sampling
Total RNA and DNA were extracted from the local and distal samples using the Spectrum™ Plant Total RNA Kit (Sigma) and FavorPrep™ Plant Genomic DNA Extraction Mini Kit (Favorgen, Wien, Austria), respectively. DNA and RNA quantity and RNA integrity were assessed with QUBIT 4 Fluorometer (Thermo Fisher Scientific, Waltham, MA, USA). For RNA samples, an IQ over 6 was considered acceptable in order to proceed sequencing.
4.4. Pss DNA Quantification in Sweet Cherry Samples
Calibration curves for DNA quantification were performed by qPCR using seven 10-fold serial dilutions of DNA extracted from A1M3 Pss strain cultivated in KB-Agar or total DNA from sweet cherry cv. ‘Lapins’ samples in an AriaMx Real-time PCR System. The qPCR reaction was performed using Brilliant III SYBR® Green qPCR Master Mix (Agilent Technologies, Santa Clara, CA, USA) following the manufacturer’s instructions, with 200 nM of each primer and 1 μL of each dilution, supplemented with Molecular Biology Grade Water (Corning®, Corning, NY, USA) to reach the final volume of 15 µL. Glutathione-dependent formaldehyde-activating enzyme 1 gene primers were used to quantify the DNA of Pss, and SAR DEFICIENT 1 gene primers were used to quantify the DNA of sweet cherry in each sample (Supplementary Table S1). Each qPCR reaction was performed in duplicate. The obtained Pss calibration curve equation was: log10 [Pss DNA (ng/μL)] = (Cq − 13.6263)/(−3.302), p-value < 0.00001, r2 × 100 = 99.947%; the sweet cherry calibration curve formula was: log10 [sweet cherry DNA (ng/μL)] = (Cq − 20.7584)/(−2.50357), p-value = 0.0038, r2 × 100 = 99.243% (Supplementary Figure S1C,D). Pss DNA quantity in each sample was defined as the ratio between the DNA quantification from Pss and sweet cherry. Purified DNA from A1M3 Pss strain and purified DNA from an uninoculated Prunus avium twig were used as positive controls. Plant DNA extraction was diluted 20-fold with Molecular Biology Grade Water (Corning®) before qPCR reaction.
4.5. Library Construction and RNA Sequencing
RNA samples were treated with DNase and library was constructed using the TruSeq stranded total RNA with Ribo-Zero Plant Kit (Illumina, San Diego, CA, USA), which generated paired-end reads of 151 nt. Sequencing was performed using the Illumina platform. DNase treatment, library construction, and RNA sequencing were carried out by Psomagen (Rockville, MD, USA).
4.6. RNA Sequence Curation and Mapping to the Reference Transcriptome
The predicted transcripts from the protein-coding genes of Prunus avium cultivar ‘Satonishiki’ [12] were used as a reference to map the reads (GenBank GCA_002207925.1). The reference transcriptome contained 35,009 transcripts, corresponding to 25,841 predicted protein-coding genes. The raw data of RNA-seq were loaded onto the CLC Genomic Workbench software, version 21.0.5 (Qiagen, Hong Kong) for the following analyses. The raw reads were trimmed to remove the adapters and nucleotides of quality scores lower than 20. The trimmed reads from each individual sample were mapped to the reference transcriptome using the following parameters: similarity fraction = 0.95, length fraction = 0.8, insertion/deletion cost = 3, mismatch cost = 2, and unspecific match limit = 10. Principal component analysis (PCA) of all the datasets was conducted to validate the technical repeats of each treatment.
4.7. Differential Gene Expression Analysis
The differential expression analysis was performed with the CLC Genomic Workbench software by comparing the mapping results of the group of samples from the trees inoculated with each Pss strain to the mock-treated control group. The samples were normalized using the trimmed mean of M values (TMM) method [44]. A false discovery rate (FDR) of 0.05 was applied to the multiple sample testing. Differential expression between two sample sets was determined based on the transcripts with absolute fold change ≥ 2.0 and FDR-adjusted p-value ≤ 0.05. Heat maps were generated with the NG-CHM builder online (https://build.ngchm.net/NGCHM-web-builder/ (accessed on 28 September 2022)) [45,46].
4.8. Gene Ontology Enrichment Analysis
The lists of differentially expressed genes (DEGs) were entered into Database for Annotation, Visualization and Integrated Discovery (DAVID, https://david.ncifcrf.gov/home.jsp (accessed on 26 June 2023)) for gene ontology (GO) annotation with p-value ≤ 0.01 [47]. The enrichment bubble plots were generated by SRplot web server (http://www.bioinformatics.com.cn/srplot (accessed on 26 June 2023).
4.9. Validation of DEGs
RNA was treated with RNase-Free Dnase I (Thermo Fisher Scientific). cDNA was synthesized using an M-MLV Reverse Transcriptase (Invitrogen™, Waltham, MA, USA) with the oligo dT primer (5′-TTTTTTTTTTTTTTTTVN-3′) according to the manufacturer’s instructions. Six DEGs between mock samples and each Pss strain-inoculated sample were selected based on a significant and similar trend of changes in their expression (Supplementary Tables S1, S9 and S10). qPCR reactions were performed using Brilliant III Ultra-Fast SYBR Green qRT-PCR Master Mix (Agilent Technologies, CA, USA) in an AriaMx Real-time PCR System (Agilent Technologies, CA, USA) following the manufacturer’s instructions (5 ng of cDNA, 200 nM of each primer, 15 μL of total volume of reaction). Each qPCR reaction was performed in triplicate. Relative expression was calculated using the 2−ΔΔCT with a primer efficiency correction [48]. Based on the RNA-seq data, two reference genes were used: peptidyl-prolyl cis-trans isomerase CYP21-1 gene and Splicing factor 3B subunit 1 gene (Supplementary Table S1). Simple linear regression analysis and coefficient determination between qPCR results and RNA-seq of the same genes were performed using Statgraphics Centurion XV, version 15.2.06.
5. Conclusions
The inoculation of Pseudomonas syringae pv. syringae strains induced differential gene expression in both local and distal tissues of sweet cherry cv. ‘Lapins’. The least virulent strain, A1M3, triggered the most extensive responses in local tissues, whereas the most virulent strain, 11116_b1, induced the strongest defense responses in distal tissues. DEGs involved in defense responses, terpene metabolism, photosynthesis, cell wall biogenesis, and carbohydrate metabolism were enriched in tissues from plants inoculated with different strains. These data serve as valuable groundwork for future research into sweet cherry defense mechanisms and the induction of immunity against Pss infection.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants12213718/s1, Figure S1. Evaluation of Pss infection on sweet cherry and calibration curves to quantify Pss DNA in plant samples. (A) Upper panels: twigs from sweet cherry cv. Lapins were inoculated by wounding with three Pss strains, 11116_b1, A1M197, and A1M3, in a 2 × 108 CFU/mL suspension. Sterile saline buffer was used as mock control. After 40 days, open wounds on twigs with Pss infection were observed (red arrows), whereas the mock control twigs showed closed wounds (blue arrows). Lower panels: viable bacteria were isolated from local samples inoculated with Pss. Scale bars, 1 cm. (B) Isolated colonies from two inoculated local samples per Pss strain were analyzed by UV fluorescence and LOPAT test. All colonies exhibited fluorescence under UV exposure, tested positive for levan production, negative for oxidase production, negative for pectinolytic activity in potato, negative for arginine hydrolase production, and triggered hypersensitive response in tobacco leaves. Two colonies per plant sample were evaluated. (C)–(D) Calibration curves for Pss and Prunus avium DNA quantification were obtained by detecting the glutathione-dependent formaldehyde-activating enzyme 1 gene in purified A1M3 strain DNA (C) and the protein SAR DEFICIENT 1 gene in DNAs purified from Prunus Avium twig (D). The equation between Cq and DNA concentration is shown. Cq, quantification cycle. Figure S2. Principal component (PC) analysis of the transcriptome datasets from the 24 samples of sweet cherry inoculated with Pss strains. (A) Proportions of variances. The red rectangle indicates the first six PCs, the sum of proportions of which exceeds 60%. (B)–(E) 3D scatter plots showing the first two PCs and one of the following PCs. Figure S3. Volcano plots showing the differential expression profiles of the datasets from the samples of sweet cherry inoculated with three Pss strains in comparison with mock-treated samples. (A)–(C) Samples taken locally to the inoculation sites. (D)–(F) Samples taken distally to the inoculation sites. Figure S4. Fold change distribution of differentially expressed genes in local and distal samples from sweet cherry inoculated with three Pss strains. Figure S5. Comparison between RNA-seq and qPCR expression data from selected DEGs. Relative expression data of genes from local samples (GolginA-4, E3 ubiquitin ligase PRT6, CBS domain-containing protein, and Actin7) and from distal samples (elF-2- α kinase GCN2, AGD14, GIGANTEA, and Actin7) obtained by qPCR were compared with their RNA-Seq results using simple linear regression. A scatter plot highlights a simple linear regression and shows the coefficient of determination (R2) and its significance between relative expression obtained by qPCR and its corresponding RNA-seq values. Figure S6. Pathogenicity assays of Pss strains in sweet cherry cv. ‘Lapins’. (A)–(C) Spray assay. (A) Pss strains (11116_b1, A1M197 and A1M3) were suspended in sterile distilled water and the concentration was adjusted to 108 CFU/mL. 50 mL of suspension was sprayed on 30 leaves per plant. The inoculated branches were sealed in plastic bags for two days. (B) Necrotic area on each leaf was measured using ImageJ. (C) Symptoms were evaluated 30 days post inoculation. Symptom incidence, percentage of leaves showing symptoms. Necrotic area, percentage of leaf area that was necrotic. The three Pss strains exhibited varying performance in both parameters. (D) Leaf disk assay adapted from Lienqueo et al. [49]. Freshly picked leaves, approximately 1 to 2 weeks old, were used in the assay. Pss suspension of 2 × 108 CFU/mL (OD600 = 0.2) was freshly prepared. Leaves were washed with tap water for 5 min and then superficially disinfected with 70% ethanol for 1 min, followed by three rinses in sterile distilled water. The disinfected leaves were dried with sterile paper towel, and circular leaf disks were obtained using a 17-mm diameter cork borer that had been pre-soaked in the bacterial suspension with 0.01% Silwet L-77 or a mock solution containing 0.8% KCl and 0.01% Silwet L-77. Sixteen leaf disks were evenly placed on each Petri dish containing 10 g/L agar, with the abaxial side facing up. Two Petri dishes were used for each strain or mock inoculation. The plates were then incubated in the dark at 26 °C for 10 days before photos were taken. Table S1. Primers for DNA quantification and DEG validation. Supplementary Table S2. DNA quantification in Prunus avium samples inoculated with different Pss strains. DNA quantification of Pss and Prunus avium was performed in three local and distal samples from twigs inoculated with mock control and 11116_b1, A1M197, and A1M3 Pss strains. Mean Cq value (performed in duplicate at least) and DNA concentrations are shown for each sample. Pss DNA presents the ratio between DNA quantification of Pss and Prunus avium in each biological replicate. ND, not detected; -, not determined. C+ Pss, purified A1M3 strain DNA; C+ P. avium, purified DNA from sweet cherry twig. C-, sterile water. Table S3. The differentially expressed genes from the local samples of sweet cherry inoculated with Pss strain A1M3. Table S4. The differentially expressed genes from the local samples of sweet cherry inoculated with Pss strain A1M197. Table S5. The differentially expressed genes from the local samples of sweet cherry inoculated with Pss strain 11116_b1. Table S6. The differentially expressed genes from the distal samples of sweet cherry inoculated with Pss strain A1M3. Table S7. The differentially expressed genes from the distal samples of sweet cherry inoculated with Pss strain A1M197. Table S8. The differentially expressed genes from the distal samples of sweet cherry inoculated with Pss strain 11116_b1. Table S9. The differentially expressed genes shared after inoculation with all three Pss strains from the local samples of sweet cherry. Table S10. The differentially expressed genes shared after inoculation with all three Pss strains from the distal samples of sweet cherry. The differentially expressed genes shared after inoculation with all three Pss strains from the distal samples of sweet cherry.
Author Contributions
Conceptualization, W.C., N.F., A.Z., P.P. and S.P.; methodology, C.R., W.C., A.Z. and L.P.; software, W.C. and F.F.; validation, F.F. and L.P.; formal analysis, W.C. and L.P.; investigation, C.R., W.C. and F.F.; resources, M.P., P.P. and N.F.; data curation, W.C.; writing—original draft preparation, W.C.; writing—review and editing, M.F.B., C.C., A.Z. and N.F.; supervision, A.Z.; project administration, M.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Proyecto Anillo ACTO 190001, Programa de Investigación Asociativa ANID, and supported by ANID FONDECYT N° 11200934, both by the Chilean government.
Data Availability Statement
The RNA-Seq data was deposited in the sequence read archive of the National Center for Biotechnology Information under accession number PRJNA1014457.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Catastro Frutícola CIREN-Odepa. Available online: https://www.odepa.gob.cl/estadisticas-del-sector/catastros-fruticolas/catastro-fruticola-ciren-odepa (accessed on 19 May 2023).
- iQonsulting. Available online: http://www.iqonsulting.com/site/ (accessed on 19 May 2023).
- Lemus, G.; Osorio, V.; France, A. Cáncer Bacteriano en Cerezos: Opciones de Control de Cancros; Informativo; Instituto de Investigaciones Agropearias: Rengo, Chile, 2019; No. 67. [Google Scholar]
- Latorre, B.A.; Gonza, J.A.; Cox, J.E.; Vial, F. Isolation of Pseudomonas syringae pv. syringae from cankers and effect of free moisture on its epiphytic populations on sweet cherry trees. Plant Dis. 1985, 69, 409–412. [Google Scholar] [CrossRef]
- Garcia, H.M.; Miranda, E.M.; Lopez, M.A.; Parra, S.J.; Rubilar, C.F.; Silva-Moreno, E.D.C.; Rubio, J.M.; Ramos, C.B. First report of bacterial canker caused by Pseudomonas syringae pv. morsprunorum race 1 on sweet cherry in Chile. Plant Dis. 2021, 105, 3287. [Google Scholar] [CrossRef] [PubMed]
- Correa, F.; Beltrán, M.F.; Millas, P.; Moreno, Z.; Hinrichsen, P.; Meza, P.; Sagredo, B. Genome sequence resources of Pseudomonas syringae strains isolated from sweet cherry orchards in Chile. MPMI 2022, 35, 933–937. [Google Scholar] [CrossRef]
- Garrett, C.M.E. Screening Prunus rootstocks for resistance to bacterial canker, caused by Pseudomonas morsprunorum. J. Hortic. Sci. 1979, 54, 189–193. [Google Scholar] [CrossRef]
- Hulin, M.T.; Vadillo Dieguez, A.; Cossu, F.; Lynn, S.; Russell, K.; Neale, H.C.; Jackson, R.W.; Arnold, D.L.; Mansfield, J.W.; Harrison, R.J. Identifying resistance in wild and ornamental cherry towards bacterial canker caused by Pseudomonas syringae. Plant Pathol. 2022, 71, 949–965. [Google Scholar] [CrossRef]
- Li, B.; Hulin, M.T.; Brain, P.; Mansfield, J.W.; Jackson, R.W.; Harrison, R.J. Rapid, automated detection of stem canker symptoms in woody perennials using artificial neural network analysis. Plant Methods 2015, 11, 57. [Google Scholar] [CrossRef]
- Mgbechi-Ezeri, J.; Porter, L.; Johnson, K.B.; Oraguzie, N. Assessment of sweet cherry (Prunus avium L.) genotypes for response to bacterial canker disease. Euphytica 2017, 213, 145. [Google Scholar] [CrossRef]
- Omrani, M.; Roth, M.; Roch, G.; Blanc, A.; Morris, C.E.; Audergon, J.M. Genome-wide association multi-locus and multi-variate linear mixed models reveal two linked loci with major effects on partial resistance of apricot to bacterial canker. BMC Plant Biol. 2019, 19, 31. [Google Scholar] [CrossRef]
- Shirasawa, K.; Isuzugawa, K.; Ikenaga, M.; Saito, Y.; Yamamoto, T.; Hirakawa, H.; Isobe, S. The genome sequence of sweet cherry (Prunus avium) for use in genomics-assisted breeding. DNA Res. 2017, 24, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Sansavini, S.; Lugli, S.; Lugli, A.; Pancaldi, M. Breeding sweet cherry for self-fertile, compact/spur tree habit and high quality fruits: Trait segregation. Acta Hortic. 1998, 468, 45–52. [Google Scholar] [CrossRef]
- Pinosio, S.; Marroni, F.; Zuccolo, A.; Vitulo, N.; Mariette, S.; Sonnante, G.; Aravanopoulos, F.A.; Ganopoulos, I.; Palasciano, M.; Vidotto, M.; et al. A draft genome of sweet cherry (Prunus avium L.) reveals genome-wide and local effects of domestication. Plant J. 2020, 103, 1420–1432. [Google Scholar] [CrossRef]
- Wang, J.; Liu, W.; Zhu, D.; Hong, P.; Zhang, S.; Xiao, S.; Tan, Y.; Chen, X.; Xu, L.; Zong, X.; et al. Chromosome-scale genome assembly of sweet cherry (Prunus avium L.) cv. Tieton obtained using long-read and Hi-C sequencing. Hortic. Res. 2020, 7, 122. [Google Scholar] [CrossRef]
- Maldonado, J.; Dhingra, A.; Carrasco, B.; Meisel, L.; Silva, H. Transcriptome datasets from leaves and fruits of the sweet cherry cultivars ‘Bing’ ‘Lapins’ and ‘Rainier’. Data Brief 2019, 23, 103696. [Google Scholar] [CrossRef] [PubMed]
- Bultreys, A.; Kaluzna, M. Bacterial cankers caused by Pseudomonas syringae on stone fruit species with special emphasis on the pathovars syringae and morsprunorum race 1 and race 2. J. Plant Pathol. 2010, 92, S1.21–S1.33. [Google Scholar]
- Tornero, P.; Chao, R.A.; Luthin, W.N.; Goff, S.A.; Dangl, J.L. Large-scale structure-function analysis of the Arabidopsis RPM1 disease resistance protein. Plant Cell 2002, 14, 435–450. [Google Scholar] [CrossRef]
- Kim, S.H.; Kwon, S.I.L.; Saha, D.; Anyanwu, N.C.; Gassmann, W. Resistance to the Pseudomonas syringae effector HopA1 Is governed by the TIR-NBS-LRR protein Rps6 and is enhanced by mutations in Srfr1. Plant Physiol. 2009, 150, 1723–1732. [Google Scholar] [CrossRef] [PubMed]
- Eitas, T.K.; Nimchuk, Z.L.; Dangl, J.L. Arabidopsis TAO1 is a TIR-NB-LRR protein that contributes to disease resistance induced by the Pseudomonas syringae effector AvrB. Proc. Natl. Acad. Sci. USA 2008, 105, 6475–6480. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Peang, H.; Li, X.; Liu, C.; Lv, X.; Wei, X.; Zou, A.; Zhang, J.; Fan, G.; Ma, G.; et al. Genome-wide analysis of NDR1/HIN1-like genes in pepper (Capsicum annuum L.) and functional characterization of CaNHL4 under biotic and abiotic stresses. Hortic. Res. 2020, 7, 93. [Google Scholar] [CrossRef]
- Kim, Y.C.; Kim, S.Y.; Choi, D.; Ryu, C.M.; Park, J.M. Molecular characterization of a pepper C2 domain-containing SRC2 protein implicated in resistance against host and non-host pathogens and abiotic stresses. Planta 2008, 227, 1169–1179. [Google Scholar] [CrossRef]
- Bittner-Eddy, P.D.; Crute, I.R.; Holub, E.B.; Beynon, J.L. RPP13 is a simple locus in Arabidopsis thaliana for alleles that specify downy mildew resistance to different avirulence determinants in Peronospora parasitica. Plant J. 2000, 21, 177–188. [Google Scholar] [CrossRef]
- Van Der Vossen, E.; Sikkema, A.; Te Lintel Hekkert, B.; Gros, J.; Stevens, P.; Muskens, M.; Wouters, D.; Pereira, A.; Stiekema, W.; Allefs, S. An ancient R gene from the wild potato species Solanum bulbocastanum confers broad-spectrum resistance to Phytophthora infestans in cultivated potato and tomato. Plant J. 2003, 36, 867–882. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Goritschnig, S.; Dong, X.; Li, X. A gain-of-function mutation in a plant disease resistance gene leads to constitutive activation of downstream signal transduction pathways in Suppressor of npr1-1, Constitutive 1. Plant Cell 2003, 15, 2636–2646. [Google Scholar] [CrossRef] [PubMed]
- Murray, S.L.; Ingle, R.A.; Petersen, L.N.; Denby, K.J. basal resistance against Pseudomonas syringae in Arabidopsis involves WRKY53 and a protein with homology to a nematode resistance protein. MPMI 2007, 20, 1431–1438. [Google Scholar] [CrossRef]
- Castel, B.; Ngou, P.M.; Cevik, V.; Redkar, A.; Kim, D.S.; Yang, Y.; Ding, P.; Jones, J.D.G. Diverse NLR immune receptors activate defence via the RPW8-NLR NRG1. New Phytol. 2019, 222, 966–980. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Yang, S.; Li, Y.; Hua, J. The Arabidopsis BAP1 and BAP2 genes are general inhibitors of programmed cell death. Plant Physiol. 2007, 145, 135–146. [Google Scholar] [CrossRef]
- Marathe, R.; Anandalakshmi, R.; Liu, Y.; Dinesh-Kumar, S.P. The tobacco mosaic virus resistance gene. N. Mol. Plant. Pathol. 2002, 3, 167–172. [Google Scholar] [CrossRef]
- Beltrán, M.F.; Correa, F.; Zamorano, A.; Fiore, N.; Rubilar, C.; Pizarro, L.S.; Sagredo, B. Comparative Genomics of Pseudomonas syringae pv. Syringae Reveals a New Convergent Group Associated with Specialization onto Cherry (Prunus avium); Centro Regional Rayentué, Instituto de Investigaciones Agropecuarias (INIA): Rengo, Chile, 2023. [Google Scholar]
- Vorwerke, S.; Schiff, C.; Santamaria, M.; Koh, S.; Nishimura, M.; Vogel, J.; Somerville, C.; Somerville, S. EDR2 negatively regulates salicylic acid-based defenses and cell death during powdery mildew infections of Arabidopsis thaliana. BMC Plant. Biol. 2007, 7, 35. [Google Scholar] [CrossRef]
- Riedlmeier, M.; Ghirardo, A.; Wenig, M.; Knappe, C.; Koch, K.; Georgii, E.; Dey, S.; Parker, J.E.; Schnitzler, J.P.; Vlot, A.C. Monoterpenes support systemic acquired resistance within and between plants. Plant Cell 2017, 29, 1440–1459. [Google Scholar] [CrossRef]
- Wenig, M.; Ghirardo, A.; Sales, J.H.; Pabst, E.S.; Breitenbach, H.H.; Antritter, F.; Weber, B.; Lange, B.; Lenk, M.; Cameron, R.K.; et al. Systemic acquired resistance networks amplify airborne defense cues. Nat. Commun. 2019, 10, 3813. [Google Scholar] [CrossRef]
- Sowden, R.G.; Watson, S.J.; Jarvis, P. The Role of Chloroplasts in Plant Pathology. Essays Biochem. 2018, 62, 21–39. [Google Scholar]
- Tran, B.Q.; Jung, S. Modulation of chloroplast components and defense responses during programmed cell death in tobacco infected with Pseudomonas syringae. Biochem. Biophys. Res. Commun. 2020, 528, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S. Cell wall signaling in plant development and defense. Annu. Rev. Plant Biol. 2022, 73, 323–353. [Google Scholar] [CrossRef] [PubMed]
- van Butselaar, T.; Van den Ackerveken, G. Salicylic acid steers the growth–immunity tradeoff. Trends Plant Sci. 2020, 25, 566–576. [Google Scholar] [CrossRef] [PubMed]
- Huot, B.; Yao, J.; Montgomery, B.L.; He, S.Y. Growth-defense tradeoffs in plants: A balancing act to optimize fitness. Mol. Plant 2014, 7, 1267–1287. [Google Scholar] [CrossRef]
- Berger, S.; Sinha, A.K.; Roitsch, T. Plant physiology meets phytopathology: Plant primary metabolism and plant–pathogen interactions. J. Exp. Bot. 2007, 58, 4019–4026. [Google Scholar] [CrossRef]
- Zeng, Q.; Man, X.; Dai, Y.; Liu, H. Pseudomonas spp. enriched in endophytic community of healthy cotton plants inhibit cotton Verticillium wilt. Front. Microbiol. 2022, 13, 906732. [Google Scholar] [CrossRef]
- Arnold, D.L.; Preston, G.M. Pseudomonas syringae: Enterprising epiphyte and stealthy parasite. Microbiology 2019, 165, 251–253. [Google Scholar] [CrossRef]
- Beltrán, M.F.; Osorio, V.; Lemus, G.; Millas, P.; France, A.; Correa, F.; Sagredo, B. Bacterial community associated with canker disease from sweet cherry orchards of central valley of Chile presents high resistance to copper. Chil. J. Agric. Res. 2021, 81, 378–389. [Google Scholar] [CrossRef]
- Lelliott, R.A.; Billing, E.; Hayward, A.C. A determinative scheme for the fluorescent plant pathogenic pseudomonads. J. Appl. Bacteriol. 1966, 29, 470–489. [Google Scholar] [CrossRef]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef]
- Broom, B.M.; Ryan, M.C.; Brown, R.E.; Ikeda, F.; Stucky, M.; Kane, D.W.; Melott, J.; Wakefield, C.; Casasent, T.D.; Akbani, R.; et al. A galaxy implementation of next-generation clustered heatmaps for interactive exploration of molecular profiling data. Cancer Res. 2017, 77, e23–e26. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.C.; Stucky, M.; Wakefield, C.; Melott, J.; Akbani, R.; Weinstein, J.N. Interactive Clustered Heat Map Builder: An easy web-based tool for creating sophisticated clustered heat maps. F1000Research 2019, 8, 1750. [Google Scholar] [CrossRef]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar] [CrossRef]
- Lienqueo, I.; Villar, L.; Beltrán, F.; Correa, F.; Sagredo, B.; Guajardo, V.; Moreno, M.A.; Almada, R. Molecular, Phenotypic and Histological Analysis Reveals a Multi-Tiered Immune Response and Callose Deposition in Stone Fruit Rootstocks (Prunus spp.) against Pseudomonas syringae pv. syringae (Pss) Infection; Centro de Estudios Avanzados en Fruticultura (CEAF), Laboratorio de Fisiología del Estrés: Rengo, Chile, 2023. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).