
Different Physiological Responses to Continuous Drought between Seedlings and Younger Individuals of Haloxylon ammodendron

Abstract
:1. Introduction
2. Results
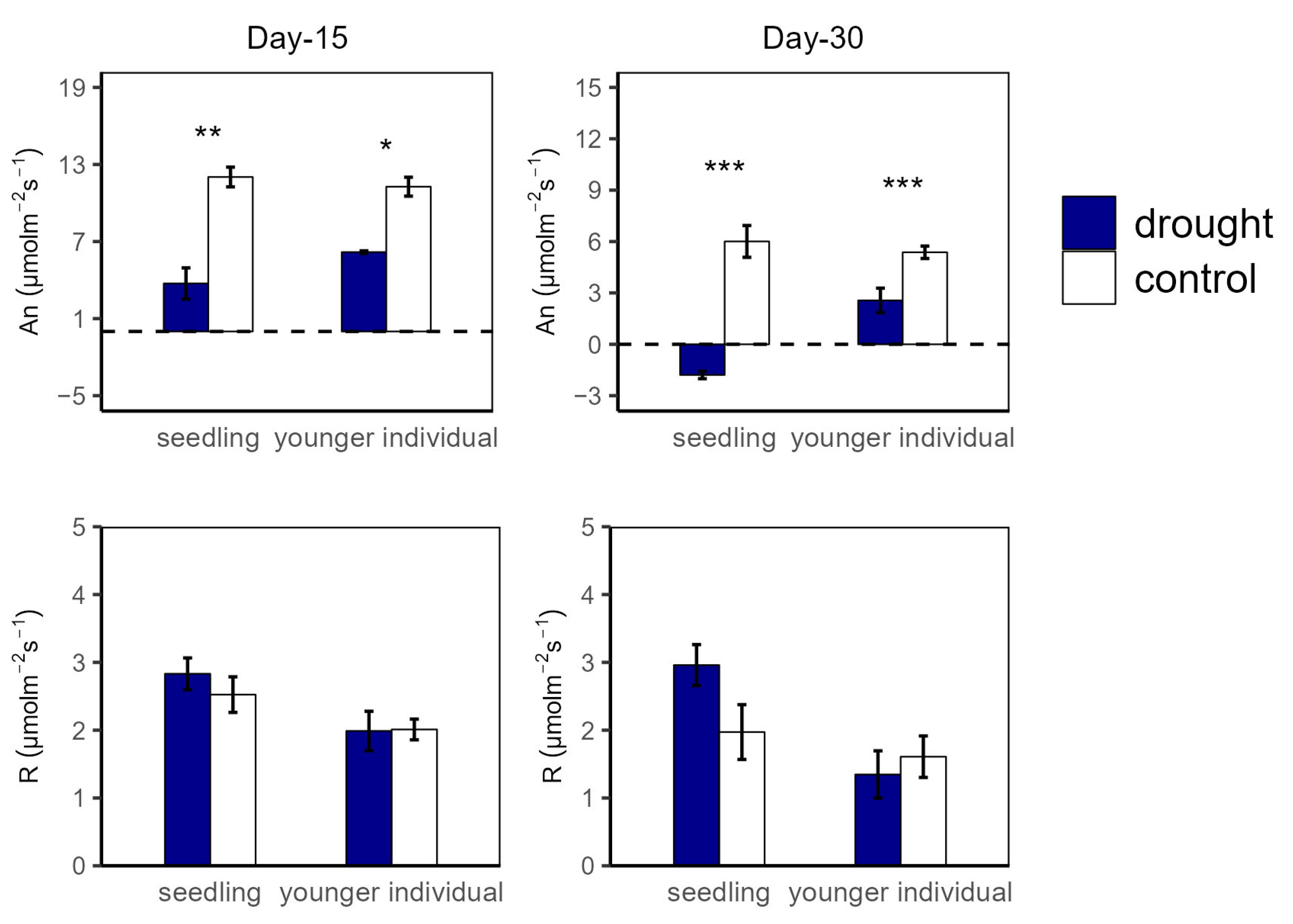
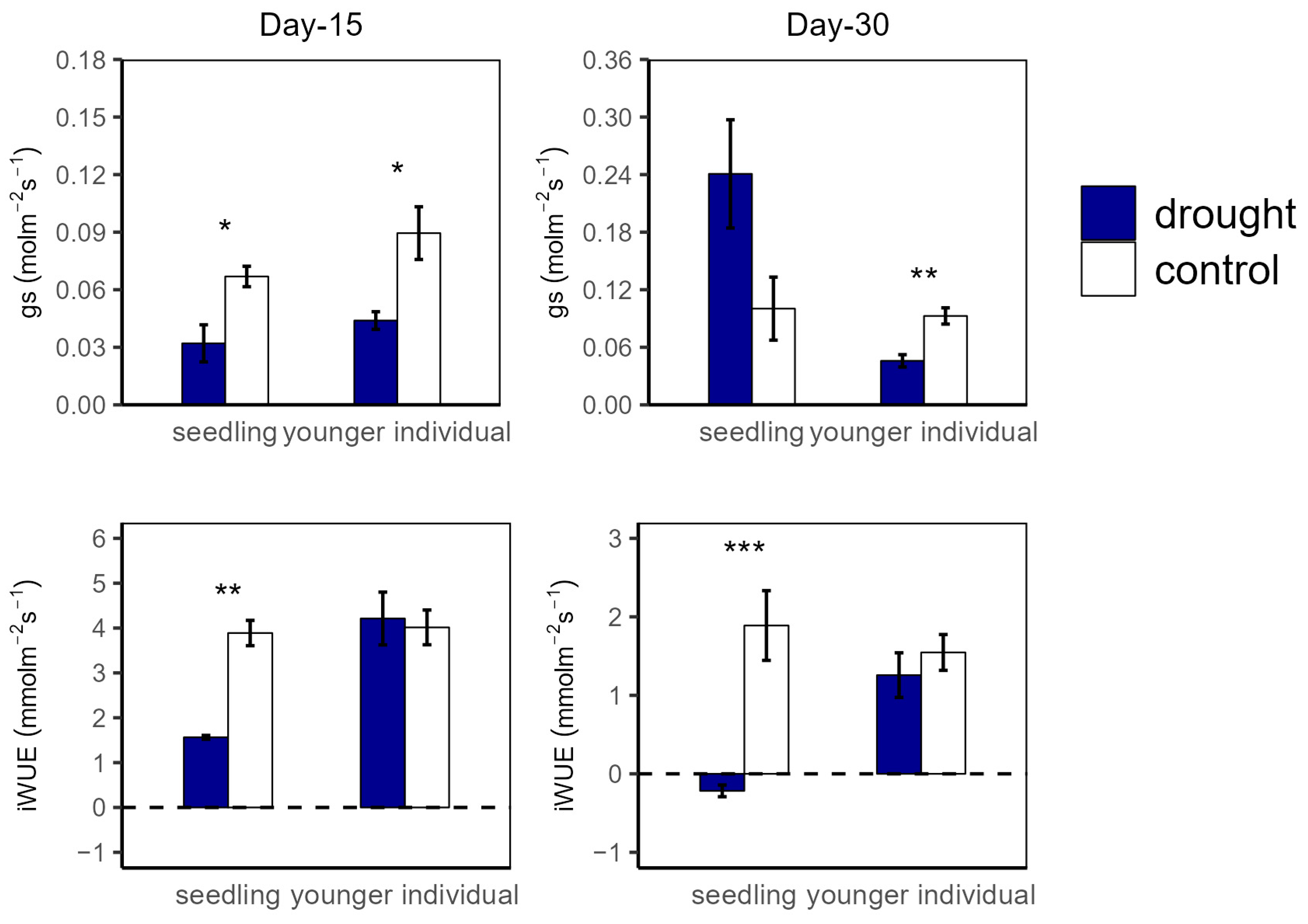
2.1. Leaf Photosynthetic Traits
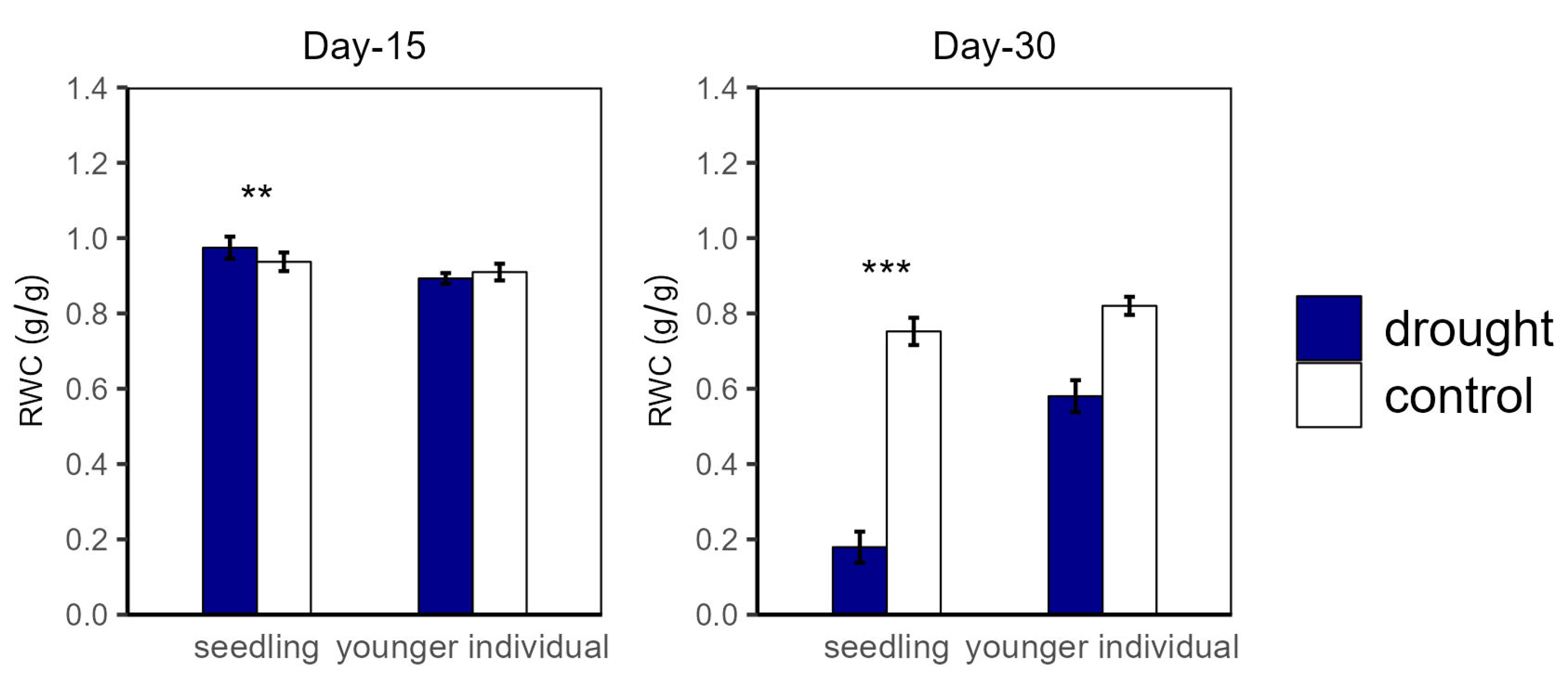
2.2. Plant Water Status
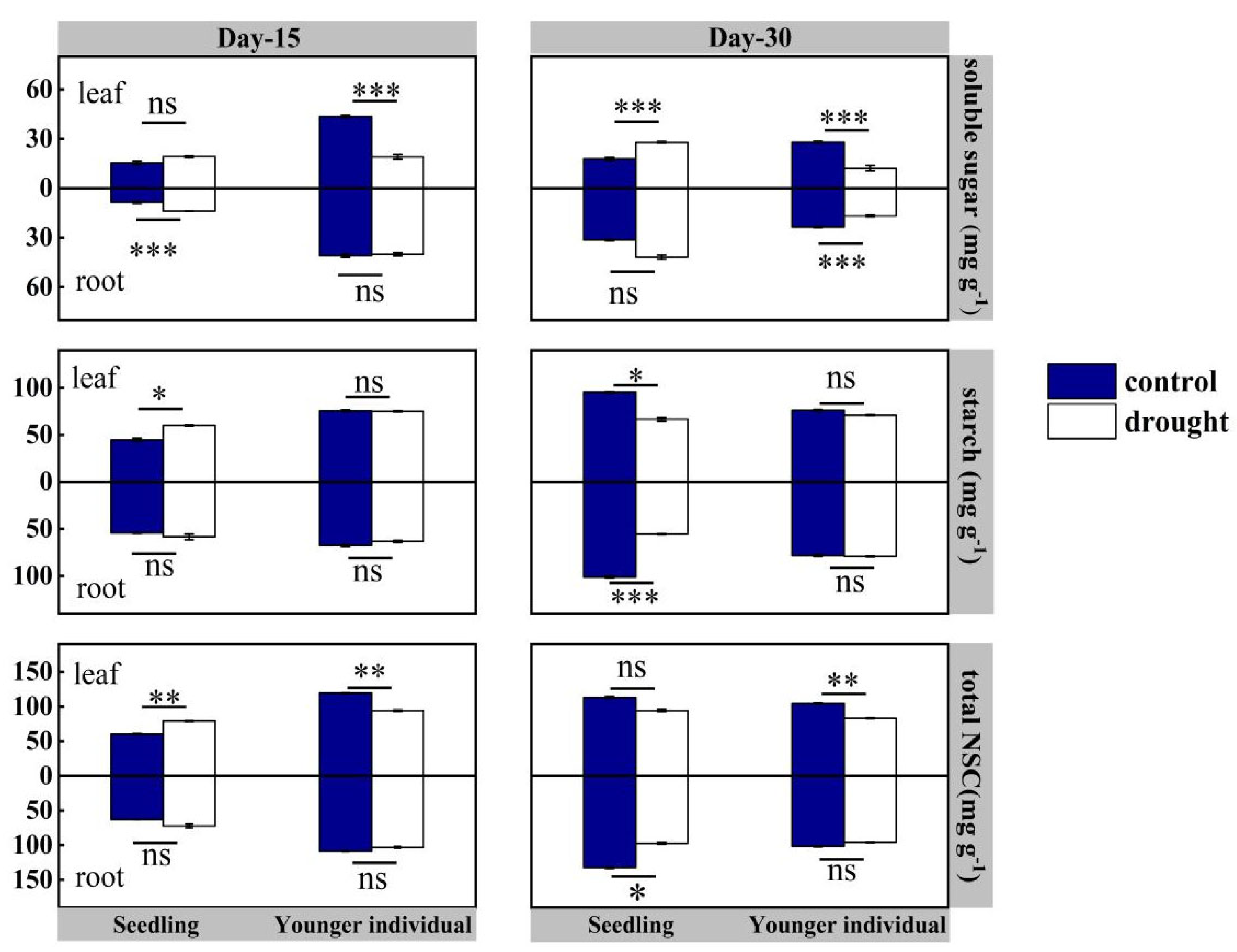
2.3. Non-Structural Carbohydrate
3. Materials and Methods
3.1. Plant Material and Experimental Design
3.2. Leaf Photosynthetic Traits
3.3. Water Status
3.4. Non-Structural Carbohydrates
3.5. Data Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barber, V.A.; Juday, G.P.; Finney, B.P. Reduced Growth of Alaskan White Spruce in the Twentieth Century from Temperature-Induced Drought Stress. Nature 2000, 405, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Simeone, C.; Maneta, M.P.; Holden, Z.A.; Sapes, G.; Sala, A.; Dobrowski, S.Z. Coupled Ecohydrology and Plant Hydraulics Modeling Predicts Ponderosa Pine Seedling Mortality and Lower Treeline in the US Northern Rocky Mountains. New Phytol. 2019, 221, 1814–1830. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, H.; Adams, D.H.; Anderegg, R.L.W.; Jansen, S.; Zeppel, J.B.M. Research Frontiers in Drought-Induced Tree Mortality Crossing Scales and Disciplines. New Phytol. 2015, 205, 965–969. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, H.; Moura, C.F.; Anderegg, R.L.W.; Ruehr, K.N.; Salmon, Y.; Allen, D.C.; Arndt, K.S.; Breshears, D.D.; Davi, H.; Galbraith, D.; et al. Research Frontiers for Improving Our Understanding of Drought-Induced Tree and Forest. New Phytol. 2018, 218, 15–28. [Google Scholar] [CrossRef]
- Babst, F.; Bouriaud, O.; Poulter, B.; Trouet, V.; Girardin, M.P.; Frank, D.C. Twentieth Century Redistribution in Climatic Drivers of Global Tree Growth. Sci. Adv. 2019, 5, eaat4313. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Nie, Y.; Chen, H.; Wang, K.; Querejeta, J.I. Water Uptake Depth Is Coordinated with Leaf Water Potential, Water-Use Efficiency and Drought Vulnerability in Karst Vegetation. New Phytol. 2021, 229, 1339–1353. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.M.R.; Lopez, R.; Nolf, M.; Hutley, L.B.; Wardlaw, T.; Cernusak, L.A.; Choat, B. Living on the Edge: A Continental-Scale Assessment of Forest Vulnerability to Drought. Glob. Chang. Biol. 2021, 27, 3620–3641. [Google Scholar] [CrossRef]
- Bennett, A.C.; Dargie, G.C.; Cuni-Sanchez, A.; Tshibamba, M.J.; Hubau, W.; Mukinzi, J.M.; Phillips, O.L.; Malhi, Y.; Sullivan, M.J.P.; Cooper, D.L.M.; et al. Resistance of African Tropical Forests to an Extreme Climate Anomaly. Proc. Natl. Acad. Sci. USA 2021, 118, e2003169118. [Google Scholar] [CrossRef] [PubMed]
- Breshears, D.D.; Cobb, N.S.; Rich, P.M.; Price, K.P.; Allen, C.D.; Balice, R.G.; Romme, W.H.; Kastens, J.H.; Floyd, M.L.; Belnap, J.; et al. Regional Vegetation Die-Off in Response to Global-Change-Type Drought. Proc. Natl. Acad. Sci. USA 2005, 102, 15144–15148. [Google Scholar] [CrossRef]
- De Kauwe, M.G.; Medlyn, B.E.; Ukkola, A.M.; Mu, M.; Sabot, M.E.B.; Pitman, A.J.; Meir, P.; Cernusak, L.A.; Rifai, S.W.; Choat, B.; et al. Identifying Areas at Risk of Drought-Induced Tree Mortality across South-Eastern Australia. Glob. Chang. Biol. 2020, 26, 5716–5733. [Google Scholar] [CrossRef]
- Blackman, C.J.; Creek, D.; Maier, C.; Aspinwall, M.J.; Drake, J.E.; Pfautsch, S.; O’Grady, A.; Deizon, S.; Medlyn, B.E.; Tissue, D.T.; et al. Drought response strategies and hydraulic traits contribute to mechanistic understanding of plant dry-down to hydraulic failure. Tree Physiol. 2019, 39, 910–924. [Google Scholar] [CrossRef] [PubMed]
- Nolan, R.H.; Gauthey, A.; Losso, A.; Medlyn, B.E.; Smith, R.; Chhajed, S.S.; Fuller, K.; Song, M.; Li, X.; Beaumont, L.J.; et al. Hydraulic Failure and Tree Size Linked with Canopy Die-Back in Eucalypt Forest During Extreme Drought. New Phytol. 2021, 230, 1354–1365. [Google Scholar] [CrossRef] [PubMed]
- Barbeta, A.; Mejia-Chang, M.; Ogaya, R.; Voltas, J.; Dawson, T.E.; Penuelas, J. The Combined Effects of a Long-Term Experimental Drought and an Extreme Drought on the Use of Plant-Water Sources in a Mediterranean Forest. Glob. Chang. Biol. 2015, 21, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Tan, N.D.; Tissue, D.T.; Huang, J.; Duan, H.L.; Su, W.; Song, Y.T.; Liu, X.J.; Liu, Y.; Li, X.; et al. Physiological Traits and Response Strategies of Four Subtropical Tree Species Exposed to Drought. Environ. Exp. Bot. 2022, 203, 105046. [Google Scholar] [CrossRef]
- Li, X.; Piao, S.; Wang, K.; Wang, X.; Wang, T.; Ciais, P.; Chen, A.; Lian, X.; Peng, S.; Penuelas, J. Temporal Trade-Off between Gymnosperm Resistance and Resilience Increases Forest Sensitivity to Extreme Drought. Nat. Ecol. Evol. 2020, 8, 1075–1083. [Google Scholar] [CrossRef]
- Isaac-Renton, M.; Montwe, D.; Hamann, A.; Spiecker, H.; Cherubini, P.; Treydte, K. Northern Forest Tree Populations Are Physiologically Maladapted to Drought. Nat. Commun. 2018, 8, 5254. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Forner, N.; Adams, H.D.; Sevanto, S.; Collins, A.D.; Dickman, L.T.; Hudson, P.J.; Zeppel, M.J.; Jenkins, M.W.; Powers, H.; Martinez-Vilalta, J.; et al. Responses of Two Semiarid Conifer Tree Species to Reduced Precipitation and Warming Reveal New Perspectives for Stomatal Regulation. Plant Cell Environ. 2016, 39, 38–49. [Google Scholar] [CrossRef]
- Peltier, D.M.P.; Ogle, K. Tree Growth Sensitivity to Climate Is Temporally Variable. Ecol. Lett. 2020, 23, 1561–1572. [Google Scholar] [CrossRef]
- McDowell, N.; Pockman, W.T.; Allen, C.D.; Breshears, D.D.; Cobb, N.; Kolb, T.; Plaut, J.; Sperry, J.; West, A.; Williams, D.G.; et al. Mechanisms of Plant Survival and Mortality During Drought: Why Do Some Plants Survive While Others Succumb to Drought? New Phytol. 2008, 178, 719–739. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Reum Han, A.; Han, A.; Kim, H.S. Responses to Drought Stress in Prunus Sargentii and Larix Kaempferi Seedlings Using Morphological and Physiological Parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- DeSoto, L.; Cailleret, M.; Sterck, F.; Jansen, S.; Kramer, K.; Robert, E.M.R.; Aakala, T.; Amoroso, M.M.; Bigler, C.; Camarero, J.J.; et al. Low Growth Resilience to Drought Is Related to Future Mortality Risk in Trees. Nat. Commun. 2020, 11, 545. [Google Scholar] [CrossRef]
- Arani, B.M.S.; Carpenter, S.R.; Lahti, L.; van Nes, E.H.; Scheffer, M. Exit Time as a Measure of Ecological Resilience. Science 2021, 372, eaay4895. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.L.; Kumar, M.; Katul, G.G.; Porporato, A. Reduced Resilience as an Early Warning Signal of Forest Mortality. Nat. Clim. Change 2019, 9, 880–885. [Google Scholar] [CrossRef]
- Taylor, S.H.; Orr, D.J.; Carmo-Silva, E.; Long, S.P. During Photosynthetic Induction, Biochemical and Stomatal Limitations Differ between Brassica Crops. Plant Cell Environ. 2020, 43, 2623–2636. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Rodriguez, J.; Perez, P.; Martinez-Carrasco, R. Photosynthesis, Carbohydrate Levels and Chlorophyll Fluorescence-Estimated Intercellular CO2 in Water-Stressed Casuarina Equisetifolia Forst. & Forst. Plant Cell Environ. 1999, 22, 867–873. [Google Scholar] [CrossRef]
- Woodruff, D.R.; Meinzer, F.C.; Marias, D.E.; Sevanto, S.; Jenkins, M.W.; McDowell, N.G. Linking Nonstructural Carbohydrate Dynamics to Gas Exchange and Leaf Hydraulic Behavior in Pinus edulis and Juniperus monosperma. New Phytol. 2015, 206, 411–421. [Google Scholar] [CrossRef]
- Damour, G.; Vandame, M.; Urban, L. Long-term drought results in a reversible decline in photosynthetic capacity in mango leaves, not just a decrease in stomatal conductance. Tree Physiol. 2009, 29, 675–684. [Google Scholar] [CrossRef]
- Cernusak, L.A.; Hutley, L.B.; Beringer, J.; Holtum, J.A.M.; Turner, B.L. Photosynthetic physiology of eucalypts along a sub-continental rainfall gradient in northern Australia. Agric. For. Meteorol. 2011, 151, 1462–1470. [Google Scholar] [CrossRef]
- Arp, W.J.; Van Mierlo, J.E.M.; Berendse, F.; Snijders, W. Interactions between Elevated CO2 Concentration, Nitrogen and Water: Effects on Growth and Water Use of Six Perennial Plant Species. Plant Cell Environ. 1998, 21, 1–11. [Google Scholar] [CrossRef]
- Buckley, T.N. How Do Stomata Respond to Water Status? New Phytol. 2019, 224, 21–36. [Google Scholar] [CrossRef]
- Yang, Y.J.; Bi, M.H.; Nie, Z.F.; Jiang, H.; Liu, X.D.; Fang, X.W.; Brodribb, T.J. Evolution of Stomatal Closure to Optimize Water-Use Efficiency in Response to Dehydration in Ferns and Seed Plants. New Phytol. 2021, 230, 2001–2010. [Google Scholar] [CrossRef] [PubMed]
- McDowell, N.G. Mechanisms Linking Drought, Hydraulics, Carbon Metabolism, and Vegetation Mortality. Plant Physiol. 2011, 155, 1051–1059. [Google Scholar] [CrossRef]
- Cowan, I.R.; Farquhar, G.D. Stomatal function in relation to leaf metabolism and environment. Symp. Soc. Exp. Biol. 1977, 31, 471–505. [Google Scholar]
- Guo, J.S.; Hultine, K.R.; Koch, G.W.; Kropp, H.; Ogle, K. Temporal Shifts in Iso/Anisohydry Revealed from Daily Observations of Plant Water Potential in a Dominant Desert Shrub. New Phytol. 2020, 225, 713–726. [Google Scholar] [CrossRef]
- Huang, G.; Li, Y.; Padilla, F.M. Ephemeral Plants Mediate Responses of Ecosystem Carbon Exchange to Increased Precipitation in a Temperate Desert. Agric. For. Meteorol. 2015, 201, 141–152. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M. Strategy Shifts in Leaf Physiology, Structure and Nutrient Content between Species of High- and Low-Rainfall and High- and Low-Nutrient Habitats. Funct. Ecol. 2001, 15, 423–434. [Google Scholar] [CrossRef]
- McDowell, N.G.; Beerling, D.J.; Breshears, D.D.; Fisher, R.A.; Raffa, K.F.; Stitt, M. The Interdependence of Mechanisms Underlying Climate-Driven Vegetation Mortality. Trends Ecol. Evol. 2011, 26, 523–532. [Google Scholar] [CrossRef]
- Arend, M.; Link, R.M.; Patthey, R.; Hoch, G.; Schuldt, B.; Kahmen, A. Rapid Hydraulic Collapse as Cause of Drought-Induced Mortality in Conifers. Proc. Natl. Acad. Sci. USA 2021, 118, e2025251118. [Google Scholar] [CrossRef] [PubMed]
- Peltier, D.M.P.; Carbone, M.S.; McIntire, C.D.; Robertson, N.; Thompson, R.A.; Malone, S.; LeMoine, J.; Richardson, A.D.; McDowell, N.G.; Adams, H.D.; et al. Carbon Starvation Following a Decade of Experimental Drought Consumes Old Reserves in Pinus Edulis. New Phytol. 2023, 240, 92–104. [Google Scholar] [CrossRef]
- Ambrose, A.R.; Baxter, W.L.; Wong, C.S.; Naesborg, R.R.; Williams, C.B.; Dawson, T.E. Contrasting Drought-Response Strategies in California Redwoods. Tree Physiol. 2015, 35, 453–469. [Google Scholar] [CrossRef]
- Wu, J.J.; Wang, J.Y.; Hui, W.K.; Zhao, F.Y.; Wang, P.Y.; Su, C.Y.; Gong, W. Physiology of Plant Responses to Water Stress and Related Genes: A Review. Forests 2022, 13, 324. [Google Scholar] [CrossRef]
- Urli, M.; Porte, A.J.; Cochard, H.; Guengant, Y.; Burlett, R.; Delzon, S. Xylem embolism threshold for catastrophic hydraulic failure in angiosperm trees. Tree Physiol. 2013, 33, 672–683. [Google Scholar] [CrossRef] [PubMed]
- Tomasella, M.; Petrussa, E.; Petruzzellis, F.; Nardini, A.; Casolo, V. The Possible Role of Non-Structural Carbohydrates in the Regulation of Tree Hydraulics. Int. J. Mol. Sci. 2019, 21, 144. [Google Scholar] [CrossRef] [PubMed]
- Piper, F.I.; Paula, S. The Role of Nonstructural Carbohydrates Storage in Forest Resilience under Climate Change. Curr. For. Rep. 2020, 6, 1–13. [Google Scholar] [CrossRef]
- Korner, C. Carbon Limitation in Trees. J. Ecol. 2003, 91, 4–17. [Google Scholar] [CrossRef]
- Chapin, F.S.; Schulze, E.D.; Mooney, H.A. The ecology and economics of storage in plants. Annu. Rev. Ecol. Syst. 1990, 21, 423–447. [Google Scholar] [CrossRef]
- Martinez-Vilalta, J.; Sala, A.; Asensio, D.; Galiano, L.; Hoch, G.; Palacio, S.; Piper, F.I.; Lloret, F. Dynamics of non-structural carbohydrates in terrestrial plants: A global synthesis. Ecol. Monogr. 2014, 86, 495–516. [Google Scholar] [CrossRef]
- Kozlowski, T.T. Carbohydrate Sources and Sinks in Woody-plants. Bot. Rev. 1992, 58, 107–222. [Google Scholar] [CrossRef]
- Woodruff, D.R.; Meinzer, F.C. Water Stress, Shoot Growth and Storage of Non-Structural Carbohydrates Along a Tree Height Gradient in a Tall Conifer. Plant Cell Environ. 2011, 34, 1920–1930. [Google Scholar] [CrossRef]
- Ryan, M.; Epron, D.; Dannoura, M.; Hölttä, T.; Sevanto, S.; Dietrich, L.; Salmon, Y. Drought Impacts on Tree Phloem: From Cell-Level Responses to Ecological Significance. Tree Physiol. 2019, 39, 173–191. [Google Scholar] [CrossRef]
- Salleo, S.; Trifilò, P.; Esposito, S.; Nardini, A.; LoGullo, M.A. Starch-to-Sugar Conversion in Wood Parenchyma of Field-Growing Laurus nobilis Plants: A Component of the Signal Pathway for Embolism Repair? Funct. Plant Biol. 2009, 36, 815–825. [Google Scholar] [CrossRef]
- Adams, H.D.; Germino, M.J.; Breshears, D.D.; Barron-Gafford, G.A.; Guardiola-Claramonte, M.; Zou, C.B.; Huxman, T.E. Nonstructural Leaf Carbohydrate Dynamics of Pinus Edulis During Drought-Induced Tree Mortality Reveal Role for Carbon Metabolism in Mortality Mechanism. New Phytol. 2013, 197, 1142–1151. [Google Scholar] [CrossRef]
- He, W.Q.; Liu, H.Y.; Qi, Y.; Liu, F.; Zhu, X.R. Patterns in Nonstructural Carbohydrate Contents at the Tree Organ Level in Response to Drought Duration. Glob. Chang. Biol. 2020, 26, 3627–3638. [Google Scholar] [CrossRef]
- Xu, G.Q.; Yu, D.D.; Xie, J.B.; Tang, L.S.; Li, Y. What Makes Haloxylon persicum Grow on Sand Dunes While H. Ammodendron Grows on Interdune Lowlands: A Proof from Reciprocal Transplant Experiments. J. Arid Land 2014, 6, 581–591. [Google Scholar] [CrossRef]
- Li, C.J.; Han, H.; Ablimiti, M.; Liu, R.; Zhang, H.; Fan, J.L. Morphological and Physiological Responses of Desert Plants to Drought Stress in a Man-Made Landscape of the Taklimakan Desert Shelter Belt. Ecol. Indic. 2022, 10, 109037. [Google Scholar] [CrossRef]
- Xu, H.; Li, Y. Water-Use Strategy of Three Central Asian Desert Shrubs and Their Responses to Rain Pulse Events. Plant Soil 2006, 285, 5–17. [Google Scholar] [CrossRef]
- Dai, Y.; Zheng, X.J.; Tang, L.S.; Li, Y. Stable Oxygen Isotopes Reveal Distinct Water Use Patterns of Two Haloxylon Species in the Gurbantonggut Desert. Plant Soil 2014, 389, 73–87. [Google Scholar] [CrossRef]
- Li, C.J.; Abulimiti, M.; Fan, J.L.; Wang, H.F. Ecologic Service, Economic Benefits, and Sustainability of the Man-Made Ecosystem in the Taklamakan Desert. Front. Environ. Sci. 2022, 10, 813932. [Google Scholar] [CrossRef]
- Hoch, G.; Richter, A.; Körner, C. Non-Structural Carbon Compounds in Temperate Forest Trees. Plant Cell Environ. 2003, 26, 1067–1081. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environmentfor Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 21 August 2022).
- Beer, C.; Ciais, P.; Reichstein, M.; Baldocchi, D.; Law, B.E.; Papale, D.; Soussana, J.F.; Ammann, C.; Buchmann, N.; Frank, D.; et al. Temporal and among-Site Variability of Inherent Water Use Efficiency at the Ecosystem Level. Global Biogeochem. Cycles 2009, 23, GB2018. [Google Scholar] [CrossRef]
- Metcalfe, D.B.; Meir, P.; Aragao, L.E.; Lobo-do-Vale, R.; Galbraith, D.; Fisher, R.A.; Chaves, M.M.; Maroco, J.P.; da Costa, A.C.; de Almeida, S.S.; et al. Shifts in Plant Respiration and Carbon Use Efficiency at a Large-Scale Drought Experiment in the Eastern Amazon. New Phytol. 2010, 187, 608–621. [Google Scholar] [CrossRef] [PubMed]
- Trueba, S.; Pan, R.; Scoffoni, C.; John, G.P.; Davis, S.D.; Sack, L. Thresholds for Leaf Damage Due to Dehydration: Declines of Hydraulic Function, Stomatal Conductance and Cellular Integrity Precede Those for Photochemistry. New Phytol. 2019, 223, 134–149. [Google Scholar] [CrossRef] [PubMed]
- Bhusal, N.; Park, I.H.; Jeong, S.; Choi, B.H.; Han, S.G.; Yoon, T.M. Photosynthetic Traits and Plant Hydraulic Dynamics in Gamhong Apple Cultivar under Drought, Waterlogging, and Stress Recovery Periods. Sci. Hortic. 2023, 321, 112276. [Google Scholar] [CrossRef]
- Peltier, D.M.P.; Guo, J.; Nguyen, P.; Bangs, M.; Gear, L.; Wilson, M.; Jefferys, S.; Samuels-Crow, K.; Yocom, L.L.; Liu, Y.; et al. Temporal Controls on Crown Nonstructural Carbohydrates in Southwestern US Tree Species. Tree Physiol. 2021, 41, 388–402. [Google Scholar] [CrossRef] [PubMed]
- Aubrey, D.P.; Teskey, R.O. Stored Root Carbohydrates Can Maintain Root Respiration for Extended Periods. New Phytol. 2018, 218, 142–152. [Google Scholar] [CrossRef]
- Pagliarani, C.; Boccacci, P.; Chitarra, W.; Cosentino, E.; Sandri, M.; Perrone, I.; Mori, A.; Cuozzo, D.; Nerva, L.; Rossato, M.; et al. Distinct Metabolic Signals Underlie Clone by Environment Interplay in “Nebbiolo” Grapes Over Ripening. Front. Plant Sci. 2019, 10, 1575. [Google Scholar] [CrossRef]
- Jiménez-Castillo, M.; Fajardo, A.; Lobos-Catalán, P.; Torres-Morales, P.; Piper, F.I. No Carbon Shortage in Declining Trees of the Isohydric Species Araucaria araucana (Molina) K. Koch under Drought. Ann. For. Sci. 2022, 79, 10. [Google Scholar] [CrossRef]
- Dash, P.K.; Cao, Y.G.; Jailani, A.K.; Gupta, P.; Venglat, P.; Xiang, D.Q.; Rai, R.; Sharma, R.; Thirunavukkarasu, N.; Abdin, M.Z.; et al. Genome-Wide Analysis of Drought Induced Gene Expression Changes in Flax (Linum usitatissimum). GM Crops Food 2014, 5, 106–119. [Google Scholar] [CrossRef]
- Dash, P.K.; Rai, R.; Pradhan, S.K.; Shivaraj, S.M.; Deshmukh, R.; Sreevathsa, R.; Singh, N.K. Drought and Oxidative Stress in Flax (Linum usitatissimum L.) Entails Harnessing Non-Canonical Reference Gene for Precise Quantification of Qrt-Pcr Gene Expression. Antioxidants 2023, 12, 950. [Google Scholar] [CrossRef]
- Dash, P.K.; Rai, R.; Mahato, A.K.; Gaikwad, K.; Singh, N.K. Transcriptome Landscape at Different Developmental Stages of a Drought Tolerant Cultivar of Flax (Linum usitatissimum). Front. Chem. 2017, 5, 82. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | An μ mol m−2 s−1 | R μ mol m−2 s−1 | gs μ mol m−2 s−1 | iWUE μ mol m−2 s−1 | Fv/Fm | LDMC g g−1 | RWC g g−1 | L-Sugar mg g−1 | L-Starch mg g−1 | L-NSC mg g−1 | R-Sugar mg g−1 | R-Starch mg g−1 | R-NSC mg g−1 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Within subject | |||||||||||||
| Treatment duration | <0.001 | <0.001 | 0.017 | <0.001 | 0.006 | <0.001 | <0.001 | 0.021 | 0.004 | 0.010 | 0.629 | <0.001 | <0.001 |
| Treatment duration and development stage | 0.030 | <0.001 | 0.017 | 0.164 | 0.005 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.251 | 0.001 |
| Treatment duration and drought treatment | 0.218 | 0.089 | 0.016 | 0.624 | 0.002 | <0.001 | <0.001 | 0.028 | 0.010 | 0.043 | 0.501 | 0.0152 | 0.013 |
| Treatment duration and drought treatment and development stage | 0.392 | 0.769 | 0.008 | 0.522 | 0.019 | <0.001 | 0.003 | 0.777 | 0.009 | 0.008 | 0.009 | 0.006 | 0.023 |
| Between subject | |||||||||||||
| Drought treatment | <0.001 | 0.046 | 0.857 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | 0.078 | <0.001 | 0.704 | 0.005 | 0.015 |
| Developing stage | 0.024 | <0.001 | 0.034 | <0.001 | 0.002 | 0.035 | <0.001 | <0.001 | 0.007 | <0.001 | 0.017 | 0.139 | 0.021 |
| Drought treatment and development stage | 0.014 | 0.157 | 0.015 | <0.001 | 0.007 | 0.002 | 0.005 | <0.001 | 0.461 | <0.001 | 0.029 | 0.012 | 0.177 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Li, M.; Li, C.; Zheng, W.; Liu, R. Different Physiological Responses to Continuous Drought between Seedlings and Younger Individuals of Haloxylon ammodendron. Plants 2023, 12, 3683. https://doi.org/10.3390/plants12213683
Chen L, Li M, Li C, Zheng W, Liu R. Different Physiological Responses to Continuous Drought between Seedlings and Younger Individuals of Haloxylon ammodendron. Plants. 2023; 12(21):3683. https://doi.org/10.3390/plants12213683
Chicago/Turabian StyleChen, Lidan, Minqing Li, Congjuan Li, Weihua Zheng, and Ran Liu. 2023. "Different Physiological Responses to Continuous Drought between Seedlings and Younger Individuals of Haloxylon ammodendron" Plants 12, no. 21: 3683. https://doi.org/10.3390/plants12213683
APA StyleChen, L., Li, M., Li, C., Zheng, W., & Liu, R. (2023). Different Physiological Responses to Continuous Drought between Seedlings and Younger Individuals of Haloxylon ammodendron. Plants, 12(21), 3683. https://doi.org/10.3390/plants12213683