
Optimizing Medium Composition and Environmental Culture Condition Enhances Antioxidant Enzymes, Recovers Gypsophila paniculata L. Hyperhydric Shoots and Improves Rooting In Vitro

 ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
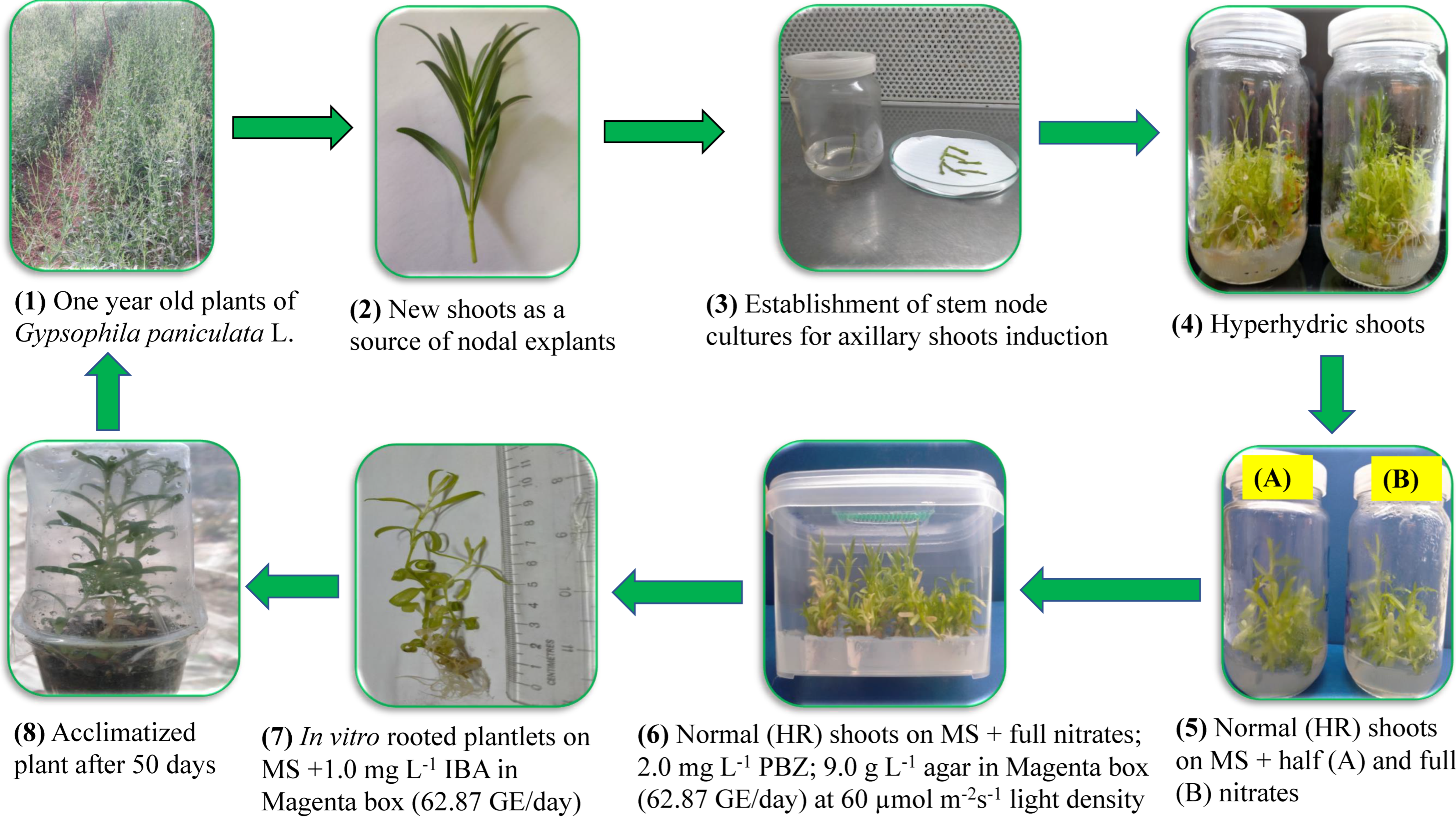
2.1. Plant Materials and Establishment of Aseptic Cultures
2.1.1. Effect of Nitrate Salt Concentrations on Recovery of Hyperhydric Shoots
2.1.2. Effect of Agar Concentrations on Recovery of Hyperhydric Shoots
2.1.3. Effect of Paclobutrazol on Recovery of Hyperhydric Shoots
2.1.4. Effects of Gas Exchange Rate and Light Density on Recovery of Hyperhydric Shoots
2.1.5. Culture Conditions
2.2. In Vitro Rooting of the Hyperhydricity Recovered Shoots
2.2.1. Antioxidant Enzymes Activity in In Vitro Rooted Plantlets
2.2.2. Electrolyte Leakage in In Vitro Rooted Plantlets
2.3. Acclimatization of In Vitro Rooted Plantlets
2.4. Statistical Analyses
3. Results
3.1. Influence of Nitrate Salts on Growth and Hyperhydricity Reversion of G. paniculata In Vitro Shoots
3.2. Influence of Agar and Nitrate Salts on Growth and Hyperhydricity Reversion of G. paniculata In Vitro Shoots
3.3. Influence of Paclobutrazol and Nitrate Salts on the Growth and Hyperhydricity Reversion of G. paniculata In Vitro Shoots
3.4. Influence of Gas Exchange Rate, Nitrate Salts and Light Density on Growth and Hyperhydricity Reversion of G. paniculata In Vitro Shoots
3.5. Influence of Gas Exchange Rate and Indole Butyric Acid on In Vitro Rooting of G. paniculata
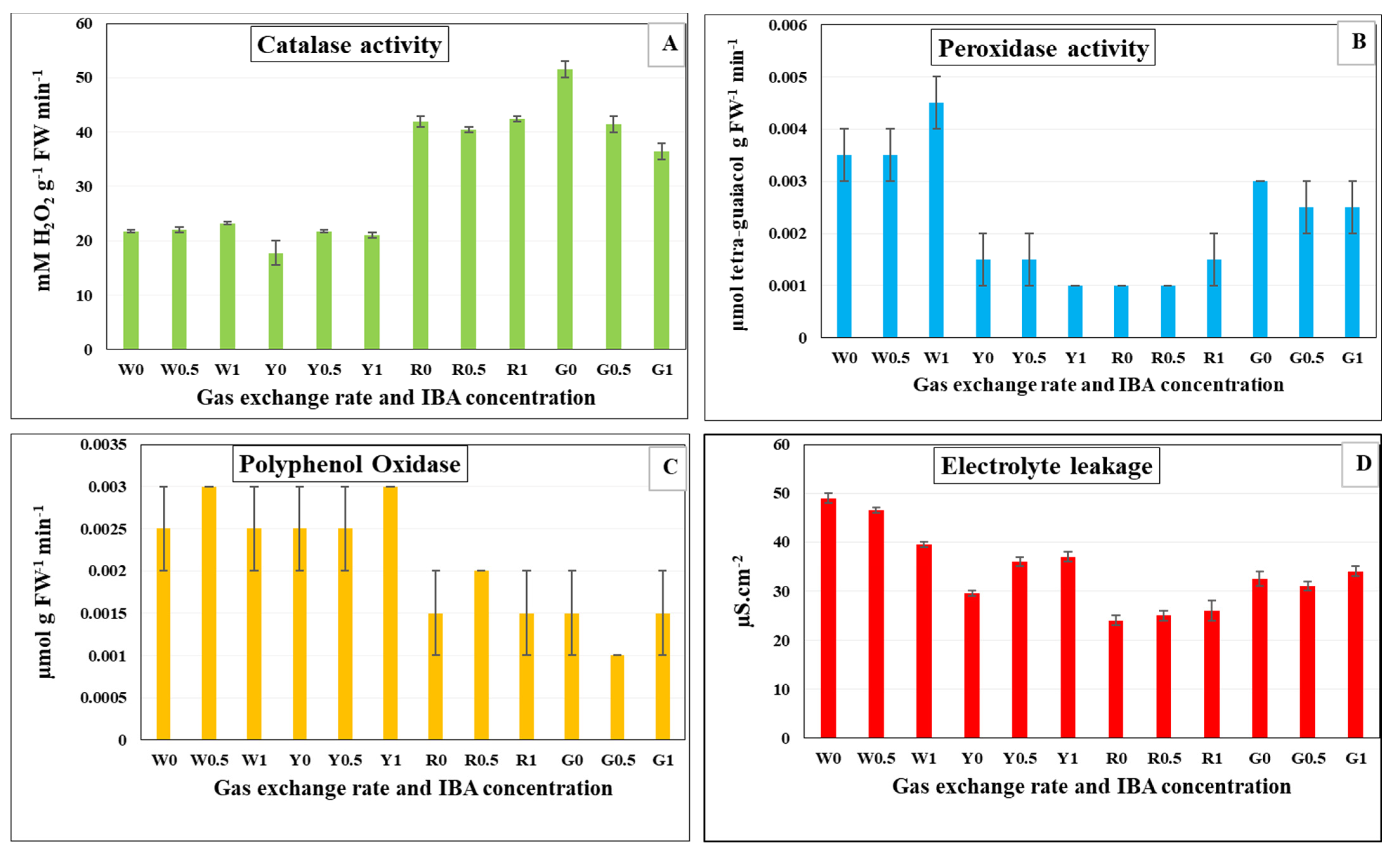
3.6. Antioxidant Enzymes Activity and Electrolyte Leakage in In Vitro Rooted Plantlets of G. paniculata
3.7. Acclimatization of In Vitro Rooted Plantlets
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Toaima, N.; Bosila, H.; El-Ateeq, A.A. In vitro Growth Regulators, Gelling Agents and Sucrose Levels Affect Micropropagation of Gypsophila paniculata L. Middle East J. Agric. Res. 2016, 5, 313–323. [Google Scholar]
- Thakur, S.; Dhiman, S.R.; Sharma, Y.D.; Singh, A.K. In vitro regeneration using nodal sections and shoot tips of gypsophila cv. ‘Bristol Fairy’. J. Hill Agric. 2013, 4, 74–77. [Google Scholar]
- Wang, S.M.; Piao, X.C.; Park, S.Y.; Lian, M.L. Improved micropropagation of Gypsophila paniculata L. with bioreactor and factors affecting ex vitro rooting in microponic system. Vitr. Cell. Dev. Biol.-Plant 2013, 49, 70–78. [Google Scholar] [CrossRef]
- Pensec, F.; Marmonier, A.; Marchal, A.; Gersch, S.; Nassr, N.; Chong, J.; Henry, M.; Demangeat, G.; Bertsch, C. Gypsophila paniculata L. root saponins as an environmentally safe treatment against two nematode, natural vectors of grapevine fanleaf degeneration. Aust. J. Grape Wine Res. 2013, 19, 439–445. [Google Scholar]
- Rady, M.R. In vitro culture of Gypsophila paniculata L. and random amplified polymorphic DNA analysis of the propagated plants. Biol. Plant. 2006, 50, 507–513. [Google Scholar] [CrossRef]
- Tian, J.; Cheng, Y.; Kong, X.; Liu, M.; Jiang, F.; Wu, Z. Induction of reactive oxygen species and the potential role of NADPH oxidase in hyperhydricity of garlic plantlets in vitro. Protoplasma 2017, 254, 379–388. [Google Scholar] [CrossRef]
- Dewir, Y.H.; El-Mahrouk, M.E.S.; Hafez, Y.M.; Teixeira da Silva, J.A.; Naidoo, Y. Hyperhydricity in African violet (Saintpaulia ionantha H. Wendl)—Biochemical aspects of normal versus hyperhydric shoots regenerated via direct adventitious shoots formation. Propag Ornam Plants 2015, 15, 53–62. [Google Scholar]
- Dewir, Y.H.; Naidoo, Y.; Teixeira da Silva, J.A. Thidiazuron-induced abnormalities in plant tissue cultures. Plant Cell Rep. 2018, 37, 1451–1470. [Google Scholar] [CrossRef]
- Saez, P.L.; Bravo, L.A.; Latsague, M.; Sanchez, M.; Rios, D. Increased light intensity during in vitro culture improves water loss control and photosynthetic performance of Castanea sativa grown in ventilated vessels. Sci. Hortic. 2012, 138, 7–16. [Google Scholar] [CrossRef]
- Muneer, S.; Park, Y.G.; Jeong, B.R. Red and blue light emitting diodes (LEDs) participate in mitigation of hyperhydricity in in vitro-grown carnation genotypes (Dianthus Caryophyllus). J. Plant Growth Regul. 2017, 37, 370–379. [Google Scholar] [CrossRef]
- Gao, H.; Xu, D.; Zhang, H.; Cheng, X.; Yang, Q. Effects of culture medium composition and PEG on hyperhydricity in Dendrobium officinale. Vitr. Cell Dev. Biol.-Plant 2020, 56, 143–149. [Google Scholar] [CrossRef]
- Dobránszki, J.; Magyar-Tábori, K.; Tombácz, E. Comparison of the rheological and diffusion properties of some gelling agents and blends and their effects on shoot multiplication. Plant Biotechnol. Rep. 2011, 5, 345–352. [Google Scholar] [CrossRef]
- Liu, M.; Jiang, F.; Kong, X.; Tian, J.; Wu, Z.; Wu, Z. Effects of multiple factors on hyperhydricity of Allium sativum L. Sci. Hortic. 2017, 217, 285–296. [Google Scholar] [CrossRef]
- Sreelekshmi, R.; Siril, E.A. Effective reversal of hyperhydricity leading to efficient micropropagation of Dianthus chinensis L. 3 Biotech 2021, 11, 95. [Google Scholar] [CrossRef]
- Abdalla, N.; El-Ramady, H.; Seliem, M.K.; El-Mahrouk, M.E.; Taha, N.; Bayoumi, Y.; Shalaby, T.A.; Dobránszki, J. An Academic and Technical Overview on Plant Micropropagation Challenges. Horticulturae 2022, 8, 677. [Google Scholar] [CrossRef]
- Barrales-López, A.; Robledo-Paz, A.; Trejo, C.; Espitia-Rangel, E.; Rodríguez-De, J.L. Improved in vitro rooting and acclimatization of Capsicum chinense Jacq. Plantlets. Vitr. Cell. Dev. Biol.-Plant 2015, 51, 274–283. [Google Scholar] [CrossRef]
- Dewir, Y.H.; Indoliya, Y.; Chakrabarty, D.; Paek, K.Y. Biochemical and physiological aspects of hyperhydricity in liquid culture system. In Production of Biomass and Bioactive Compounds Using Bioreactor Technology; Springer: Dordrecht, The Netherlands, 2014; pp. 693–709. [Google Scholar]
- Gao, H.; Xia, X.; An, L.; Xin, X.; Liang, Y. Reversion of hyperhydricity in pink (Dianthus chinensis L.) plantlets by AgNO3 and its associated mechanism during in vitro culture. Plant Sci. 2017, 254, 1–11. [Google Scholar] [CrossRef]
- El-Mahrouk, M.E.; El-Shereif, A.R.; Dewir, Y.H.; Hafez, Y.M.; Abdelaal, K.A.; El-Hendawy, A.S. Micropropagation of Banana: Reversion, Rooting, and Acclimatization of Hyperhydric Shoots. HortScience 2019, 54, 1384–1390. [Google Scholar] [CrossRef]
- Nikam, T.D.; Mulye, K.V.; Chambhare, M.R.; Nikule, H.A.; Ahire, M.L. Reduction in hyperhydricity and improvement in in vitro propagation of commercial hard fibre and medicinal glycoside yielding Agave sisalana Perr. ex Engelm by NaCl and polyethylene glycol. Plant Cell Tiss. Org. Cult. 2019, 138, 67–78. [Google Scholar] [CrossRef]
- Sreelekshmi, R.; Siril, E.A.; Muthukrishnan, S. Role of Biogenic Silver Nanoparticles on Hyperhydricity Reversion in Dianthus chinensis L. an In Vitro Model Culture. J. Plant Growth Regul. 2022, 41, 23–39. [Google Scholar] [CrossRef]
- Ramage, C.M.; Williams, R.R. Inorganic nitrogen requirements during shoot organogenesis in tobacco leaf discs. J. Exp. 2002, 53, 1437–1443. [Google Scholar] [CrossRef]
- Ivanova, M.; Van Staden, J. Nitrogen source, concentration, and NH4+:NO3– ratio influence shoot regeneration and hyperhydricity in tissue cultured Aloe polyphylla. Plant Cell Tissue Organ Cult. 2009, 99, 167–174. [Google Scholar] [CrossRef]
- Ivanova, M.; Van Staden, J. Influence of gelling agent and cytokinins on the control of hyperhydricity in Aloe polyphylla. Plant Cell Tiss. Org. Cult. 2011, 104, 13–21. [Google Scholar] [CrossRef]
- Casanova, E.; Moysset, L.; Trillas, M.I. Effects of agar concentration and vessel closure on the organogenesis and hyperhydricity of adventitious carnation shoots. Biol. Plant. 2008, 52, 1–8. [Google Scholar] [CrossRef]
- Lucho, S.R.; do Amaral, M.N.; Milech, C.; Bianchi, V.J.; Almagro, L.; Ferrer, M.A.; Calderón, A.A.; Braga, E.J.B. Gibberellin reverses the negative effect of paclobutrazol but not of chlorocholine chloride on the expression of SGs/Gas biosynthesis-related genes and increases the levels of relevant metabolites in Stevia rebaudiana. Plant Cell Tiss. Org. Cult. 2021, 146, 171–184. [Google Scholar] [CrossRef]
- Hajihashemi, S.; Ehsanpour, A.A. Influence of exogenously applied paclobutrazol on some physiological traits and growth of Stevia rebaudiana under in vitro drought stress. Biologia 2013, 68, 414–420. [Google Scholar] [CrossRef]
- Tsay, H.S.; Lee, C.Y.; Agrawal, D.C.; Basker, S. Influence of ventilation closure, gelling agent and explant type on shoot bud proliferation and hyperhydricity in scrophularia yoshimurae—A medicinal plant. Vitr. Cell. Dev. Biol.-Plant 2006, 42, 445–449. [Google Scholar] [CrossRef]
- Tabart, J.; Franck, T.; Kevers, C.; Dommes, J. Effect of polyamines and polyamine precursors on hyperhydricity in micropropagated apple shoots. Plant Cell Tiss. Org. Cult. 2015, 120, 11–18. [Google Scholar] [CrossRef]
- Dewir, Y.; Chakrabarty, D.; Ali, M.; Hahn, E.; Paek, K. Lipid peroxidation and antioxidant enzyme activities of Euphorbia millii hyperhydric shoots. Environ. Exp. Bot. 2006, 58, 93–99. [Google Scholar] [CrossRef]
- Balen, B.; Tkalec, M.; Pavoković, D.; Pevalek-Kozlina, B.; Krsnik-Rasol, M. Growth conditions in in vitro culture can induce oxidative stress in Mammillaria gracilis tissues. J. Plant Growth Regul. 2009, 28, 36–45. [Google Scholar] [CrossRef]
- Tian, J.; Jiang, F.; Wu, Z. The apoplastic oxidative burst as a key factor of hyperhydricity in garlic plantlet in vitro. Plant Cell Tiss. Org. Cult. 2014, 120, 571–584. [Google Scholar] [CrossRef]
- Muneer, S.; Soundararajan, P.; Jeong, B.R. Proteomic and antioxidant analysis elucidates the underlying mechanism of tolerance to hyperhydricity stress in in vitro shoot cultures of Dianthus caryophyllus. J. Plant Growth Regul. 2016, 35, 667–679. [Google Scholar] [CrossRef]
- Gupta, S.D.; Prasad, V.S.S. Shoot multiplication kinetics and hyperhydric status of regenerated shoots of gladiolus in agar-solidified and matrix-supported liquid cultures. Plant Biotechnol. Rep. 2010, 4, 85–94. [Google Scholar] [CrossRef]
- Chakrabarty, D.; Park, S.; Ali, M.; Shin, K.; Paek, K. Hyperhydricity in apple: Ultrastuctural and physiological aspects. Tree Physiol. 2005, 26, 377–388. [Google Scholar] [CrossRef]
- Han, B.H.; Paek, K.Y.; Choi, J.K. Micropropagation of Gypsophila paniculata L. using shoot tip culture in vitro. J. Korean Soc. hort. Sci. 1991, 32, 394–400. [Google Scholar]
- Han, B.H.; Paek, K.Y.; Choi, J.K. Prevention of vitrification of Gypsophila paniculata L. regenerated in vitro. J. Korean Soc. hort. Sci. 1991, 32, 518–524. [Google Scholar]
- Zamorano-Mendoza, J.J.; Mejia-Munoz, J.M. In vitro propagation of gypsophila (Gypsophila paniculata L.) cv. Perfecta. Revista-Chapingo. Ser. Hort. 1994, 1, 67–71. [Google Scholar]
- Zuker, A.; Ahroni, A.; Shejtman, H.; Vainstein, A. Adventitious shoot regeneration from leaf explants of Gypsophila paniculata L. Plant Cell Rep. 1997, 16, 775–778. [Google Scholar] [CrossRef]
- Salman, M.N. Establishment of callus and cell suspension cultures from Gypsophila paniculata L. leaf segments and study of the attachment of host cells by Erwinia herbicola pv. gypsophilae. Plant Cell Tiss. Org. Cult. 2002, 69, 189–196. [Google Scholar] [CrossRef]
- Rady, M.; Bekheet, S. Effect of Temperature, Sugar and Vessel Closure on in vitro Growth of Gypsophila paniculata L. Floric. Ornam. Biotechnol. 2008, 2, 73–76. [Google Scholar]
- Barakat, M.N.; El-Sammak, H. In vitro culture and plant regeneration from shoot tip and lateral bud explants of Gypsophila paniculata L. J. Med. Plant. Res. 2011, 5, 3351–3358. [Google Scholar]
- Bioplants, K.F. Gypsophila Manual KF Bioplants. Kirtane Baug, Mundhwa Road, Magarpatta, Hadapsar, Pune-411036, India, pp. 1–19. 2008. Available online: https://www.kfbioplants.com/images/cultivation-manuals/Gypsophila-manual.pdf (accessed on 30 November 2022).
- Aebi, H. Catalase in vitro. Methods Enzymo. 1984, 105, 121–126. [Google Scholar]
- Hammerschmidt, R.; Nuckles, E.M.; Kuc, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]
- Malik, C.P.; Singh, M.B. Plant Emynology and Histoenzymology; Kalyani Publishers: New Delhi, India, 1980; pp. 54–56. [Google Scholar]
- Whitlow, T.H.; Bassuk, N.L.; Ranney, T.G.; Reichert, D.L. An improved method for using electrolyte leakage to assess membrane competence in plant tissues. Plant Physiol. 1992, 98, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Szalai, G.; Janda, T.; Paldi, E.; Szigeti, Z. Role of light in the development of post-chilling symptoms in maize. J. Plant Physiol. 1996, 148, 378–383. [Google Scholar] [CrossRef]
- Dewir, Y.H.; El-Mahrouk, M.E.; AL-Shmgani, H.S.; Rihan, H.Z.; Teixeira Da Silva, J.A.; Fuller, M.P. Photosynthetic and biochemical characterization of in vitro-derived African violet (Saintpaulia ionantha H.Wendl) plants to ex vitro conditions. J. Plant Interact. 2015, 10, 101–108. [Google Scholar] [CrossRef]
- Costat; version 6.3111; Copyright 1998–2005. CoHort Software: Monteray, CA, USA, 1998.
- Duncan, D.B. Multiple Range and Multiple F Tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Polivanova, O.B.; Bedarev, V.A. Hyperhydricity in Plant Tissue Culture. Plants 2022, 11, 3313. [Google Scholar] [CrossRef]
- Poothong, S.; Reed, B.M. Optimizing shoot culture media for Rubus germplasm: The effects of NH4+, NO3−, and total nitrogen. Vitr. Cell. Dev. Biol. Plant 2016, 52, 265–275. [Google Scholar] [CrossRef]
- Debergh, P.; Harbaoui, Y.; Lemeur, R. Mass propagation of globe artichoke (Cynara scolymus): Evaluation of different hypotheses to overcome vitrification with special reference to water potential. Physiol. Plant. 1981, 53, 181–187. [Google Scholar] [CrossRef]
- Vlachou, G.; Trigka, M.; Papafotiou, M. Effect of plant growth regulators and agar concentration on shoot multiplication and hyperhydricity of Anthyllis barba-jovis. Acta Hortic. 2020, 1298, 341–346. [Google Scholar] [CrossRef]
- Ziv, M.; Ariel, T. Bud proliferation and plant regeneration in liquid-cultured Philodendron treated with ancymidol and paclobutrazol. J. Plant Growth Regul. 1991, 10, 53–57. [Google Scholar] [CrossRef]
- Chen, J.; Hall, D.E.; de Luca, V. Effects of the growth retardant paclobutrazol on large-scale micropropagation of daylily (hemerocallis spp.). Vitr. Cell. Dev. Biol.-Plant 2005, 41, 58–62. [Google Scholar] [CrossRef]
- Fletcher, R.A.; Gilley, A. Triazoles as plant growth regulators and stress protectants. Hort. Rev. 2000, 24, 55–138. [Google Scholar]
- Baninasab, B.; Ghobadi, C. Influence of paclobutrazol and application methods on high-temperature stress injury in cucumber seedlings. J. Plant Growth Regul. 2011, 30, 213–219. [Google Scholar] [CrossRef]
- Soumya, P.R.; Kumar, P.; Pal, M. Paclobutrazol: A novel plant growth regulator and multi-stress ameliorant. Ind. J. Plant Physiol. 2017, 22, 267–278. [Google Scholar] [CrossRef]
- Bortolotti da Silva, A.; Lima, P.P.; Simao de Oliveira, L.E.; Moreira, A.L. In vitro growth and leaf anatomy of Cattleya walkeriana (Gardner, 1839) grown in natural ventilation system. Rev. Ceres Viçosa 2014, 61, 883–890. [Google Scholar] [CrossRef]
- Kubota, C.; Kozai, T. Growth and net photosynthetic rate of Solanum tuberosum in vitro under forced and natural ventilation. HortScience 1992, 27, 1312–1314. [Google Scholar] [CrossRef]
- Fuentes, G.; Talavera, C.; Oropeza, C.; Desjardins, Y.; Santamaria, J.M. Exogenous sucrose can decrease in vitro photosynthesis but improve field survival and growth of coconut (Cocos nucifera L.) in vitro plantlets. Vitr. Plant 2005, 4, 69–76. [Google Scholar] [CrossRef]
- Pérez-Tornero, O.; Egea, J.; Olmos, E.; Burgos, L. Control of hyperhydricity in micropropagated apricot cultivars. Vitr. Cell. Dev. Biol.-Plant 2001, 37, 250–254. [Google Scholar] [CrossRef]
- Dillen, W.; Buysens, S. A simple technique to overcome vitrification in Gypsophila paniculata L. Plant Cell Tiss. Org. Cult. 1989, 19, 181–188. [Google Scholar] [CrossRef]
- Nguyen, Q.H.; Thi, L.T.; Park, Y.G.; Jeong, B.R. Promoted Growth and Development of Carnation Plantlets In Vitro by Ventilation and Combined Red and Blue Light. Flower Res. J. 2018, 26, 166–178. [Google Scholar] [CrossRef]
- Islam, M.T.; Dembele, P.D.; Keller, E.R.J. Influence of explant, temperature and different culture vessels on in vitro culture for germplasm maintenance of four mint accessions. Plant Cell Tiss. Org. Cult. 2005, 81, 123–130. [Google Scholar] [CrossRef]
- Kiferle, C.; Lucchesini, M.; Maggini, R.; Pardossi, A.; Mensuali-Sodi, A. In vitro culture of sweet basil: Gas exchanges, growth, and rosmarinic acid production. Biol. Plant. 2014, 58, 601–610. [Google Scholar] [CrossRef]
- Mamedes-Rodrigues, T.C.; Batista, D.S.; Napoleão, T.A.; Fortini, E.A.; Cruz, A.C.F.; Costa, M.G.C.; Otoni, W.C. Regulation of cell wall development in Brachypodium distachyon in vitro as affected by cytokinin and gas exchange. Plant Cell Tiss. Org. Cult. 2019, 136, 207–219. [Google Scholar] [CrossRef]
- Mills, D.; Yanqing, Z.; Benzioni, A. Improvement of Jojoba shoot multiplication in vitro by ventilation. Vitr. Cell. Dev. Biol.-Plant 2004, 40, 396–402. [Google Scholar] [CrossRef]
- Wojtania, A.; Skrzypek, E.; Marasek-Ciolakowska, A. Soluble sugar, starch and phenolic status during rooting of easy and difficult-to-root magnolia cultivars. Plant Cell Tiss. Org. Cult. 2019, 136, 499–510. [Google Scholar] [CrossRef]
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2016, 15, 71. [Google Scholar] [CrossRef]
- El-Banna, M.F.; Abdelaal, K.A.A. Response of strawberry plants grown in the hydroponic system to pretreatment with H2O2 before exposure to salinity stress. J. Plant Prod. 2018, 9, 989–1001. [Google Scholar] [CrossRef]
- Seliem, M.K.; El-Mahrouk, M.E.; El-Banna, A.N.; Hafez, Y.M.; Dewir, Y.H. Micropropagation of Philodendron selloum: Influence of copper sulfate on endophytic bacterial contamination, antioxidant enzyme activity electrolyte leakage, and plant survival. S. Afr. J. Bot. 2021, 139, 230–240. [Google Scholar] [CrossRef]
- Viczian, O.; Kunstler, A.; Hafez, Y.M.; Kiraly, L. Catalases may play different roles in influencing resistance to virus-induced hypersensitive necrosis. Acta Phytopathol. Entomol. Hung. 2014, 49, 189–200. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Treatments | Supplemented Salts (mg L−1) | Final Concentrations of K+, NO3−, and NH4+ Have Been Calculated in mg L−1 | |||||
|---|---|---|---|---|---|---|---|
| KNO3 | NH4NO3 | K+ (KNO3) | NO3− (KNO3) | NH4+ (NH4NO3) | NO3− (NH4NO3) | Total NO3− | |
| # Full nitrates MS | 1900 | 1650 | 730 | 1165 | 371 | 1278 | 2443 |
| ½ nitrates MS | 950 | 825 | 367 | 582 | 185 | 639 | 1803 |
| ¼ nitrates MS | 470 | 413 | 181 | 288 | 92.9 | 320 | 608 |
| 1/8 nitrates MS | 235 | 206 | 90.9 | 144 | 46.4 | 159 | 303 |
| 1/16 nitrates MS | 120 | 103 | 46.4 | 73.6 | 23.18 | 79.8 | 153.4 |
| Treatment (mg L−1) KNO3 + NH4NO3 | Shoot Length (cm) | No. of Shoots/Explant | No. of Leaves/Shoot | Shoot Fresh Weight (g) | Normal Shoots (%) |
|---|---|---|---|---|---|
| 120 + 103 | 3.00 d | 1.00 b | 9.00 c | 0.09 d | 33.00 d |
| 235 + 206 | 5.00 bc | 1.00 b | 11.70 b | 0.28 cd | 33.00 d |
| 470 + 413 | 3.83 cd | 1.70 b | 14.00 b | 0.47 bc | 50.00 c |
| 950 + 825 | 5.70 ab | 2.70 a | 18.00 a | 0.76 ab | 75.00 b |
| #1900 + 1650 | 6.70 a | 1.33 b | 13.33 b | 0.99 a | 85.00 a |
| Significance | ** | ** | *** | *** | ** |
| Treatments KNO3 + NH4NO3 | Shoot Length (cm) | No. of Shoots/Explant | No. of Leaves/Shoot | Shoot Fresh Weight (g) | Normal Shoots (%) | |
|---|---|---|---|---|---|---|
| Agar (g L−1) | (mg L−1) | |||||
| 7 | 950 + 825 | 8.00 b | 3.0 de | 21.67 ab | 1.70 a | 29.33 cd |
| 1900 + 1650 | 6.67 c | 3.0 de | 15.00 c | 1.30 ab | 25.00 d | |
| 8 | 950 + 825 | 8.30 ab | 4.0 bcd | 23.30 ab | 1.73 a | 33.30 c |
| 1900 + 1650 | 5.00 e | 4.0 bcd | 20.00 b | 1.30 ab | 41.67 c | |
| 9 | 950 + 825 | 9.00 a | 5.0 ab | 20.00 b | 0.37 c | 95.00 a |
| 1900 + 1650 | 6.00 cd | 6.0 a | 25.00 a | 1.83 a | 90.00 a | |
| 10 | 950 + 825 | 6.00 cd | 4.3 bc | 23.30 ab | 0.80 bc | 50.00 b |
| 1900 + 1650 | 6.00 cd | 1.3 f | 20.00 b | 1.60 a | 58.00 b | |
| 11 | 950 + 825 | 5.67 de | 3.3 cde | 21.67 ab | 1.50 a | 33.30 c |
| 1900 + 1650 | 5.00 e | 2.3 e | 23.30 ab | 1.73 a | 41.67c | |
| Significance | N | *** | ** | Ns | * | Ns |
| A | *** | *** | * | Ns | *** | |
| N × A | *** | *** | *** | *** | * | |
| Treatments | Shoot Length (cm) | No. of Shoots/Explant | No. of Leaves/Shoot | Shoot Fresh Weight (g) | Normal Shoots (%) | |
|---|---|---|---|---|---|---|
| Paclobutrazol (mg L−1) | KNO3 + NH4NO3 (mg L−1) | |||||
| 0 | 950 + 825 | 9.00 a | 3.67 ab | 21.67 bc | 1.70 ab | 50.00 c |
| 1900 + 1650 | 6.00 c | 1.33 fg | 15.00 d | 1.31 bcd | 35.00 d | |
| 1.0 | 950 + 825 | 4.00 d | 1.00 g | 15.00 d | 0.37 e | 35.00 d |
| 1900 + 1650 | 4.33 d | 2.67 cde | 25.00 a | 1.30 bcd | 65.00 c | |
| 1.5 | 950 + 825 | 6.50 c | 1.00 g | 15.00 d | 0.80 de | 25.00 e |
| 1900 + 1650 | 3.33 ef | 3.67 ab | 20.00 c | 1.63 ab | 50.00 c | |
| 2.0 | 950 + 825 | 6.00 c | 4.00 a | 25.00 a | 1.50 abc | 25.00 e |
| 1900 + 1650 | 3.50 efd | 2.33 de | 20.00 c | 1.60 abc | 100.00 a | |
| 2.5 | 950 + 825 | 6.00 b | 3.33 abc | 23.33 ab | 1.03 cd | 35.00 d |
| 1900 + 1650 | 3.33 ef | 3.00 bcd | 23.33 ab | 2.00 a | 25.00 e | |
| 3.0 | 950 + 825 | 6.00 c | 3.00 bcd | 21.67 bc | 2.10 a | 25.00 e |
| 1900 + 1650 | 2.67 f | 2.00 ef | 20.00 c | 1.67 ab | 25.00 e | |
| Significance | N | *** | NS | NS | ** | * |
| p | * | *** | *** | *** | *** | |
| N × p | *** | *** | *** | *** | ** | |
| Treatments | Shoot Length (cm) | No. of Shoots/ Explant | No. of Leaves/ Shoot | Shoot Fresh Weight (g) | Normal Shoots (%) | ||
|---|---|---|---|---|---|---|---|
| GER (GE/day) | KNO3 + NH4NO3 (mg L−1) | PPFD (µmol m−2s−1) | |||||
| GER1 (7.44) | 950 + 825 | 30 | 2.00 h | 1.00 d | 10.00 e | 0.90 e | 35.00 h |
| 1900 + 1650 | 30 | 3.67 cdef | 1.67 cd | 13.33 de | 1.17 de | 25.00 i | |
| 950 + 825 | 60 | 2.33 gh | 2.00 bcd | 15.00 cde | 1.00 e | 25.00 i | |
| 1900 + 1650 | 60 | 3.00 efgh | 1.00 d | 10.00 e | 0.90 e | 35.00 h | |
| GER2 (9.14) | 950 + 825 | 30 | 2.00 h | 1.00 d | 10.00 e | 0.90 e | 35.00 h |
| 1900 + 1650 | 30 | 5.00 b | 3.00 b | 20.00 bc | 1.50 cd | 35.00 h | |
| 950 + 825 | 60 | 2.67 fgh | 1.67 cd | 13.33 de | 0.97 e | 50.00 fg | |
| 1900 + 1650 | 60 | 4.00 bcde | 2.00 bcd | 15.00 cde | 1.00 e | 55.00 f | |
| GER3 (10.83) | 950 + 825 | 30 | 2.67 fgh | 2.00 bcd | 23.33 b | 1.83 bc | 50.00 fg |
| 1900 + 1650 | 30 | 3.33 defg | 1.33 cd | 12.67 de | 1.17 de | 66.67 de | |
| 950 + 825 | 60 | 5.00 b | 2.00 bcd | 35.00 a | 2.13 ab | 70.00 de | |
| 1900 + 1650 | 60 | 4.00 bcde | 2.00 bcd | 15.00 cde | 1.13 de | 85.00 bc | |
| GER4 (62.87) | 950 + 825 | 30 | 4.67 bc | 2.00 bcd | 17.33 cd | 1.27 de | 67.00 de |
| 1900 + 1650 | 30 | 4.33 bcd | 2.33 bc | 16.67 cd | 1.67 de | 75.00 d | |
| 950 + 825 | 60 | 7.00 a | 2.00 bcd | 15.00 cde | 1.17 de | 90.00 ab | |
| 1900 + 1650 | 60 | 6.33 a | 4.33 a | 40.00 a | 2.50 a | 95.00 a | |
| Significance | N | ** | * | Ns | Ns | Ns | |
| GER | *** | *** | *** | *** | *** | ||
| L | *** | Ns | *** | Ns | *** | ||
| N × GER | *** | ** | *** | *** | *** | ||
| N × L | ** | Ns | Ns | Ns | Ns | ||
| GER × L | *** | Ns | *** | ** | *** | ||
| N × GER × L | NS | ** | *** | *** | *** | ||
| Treatments | Plantlet Length (cm) | No. of Leaves | Plantlet Fresh Weight (g) | Rooting (%) | |
|---|---|---|---|---|---|
| GER (GE/day) | IBA (mg L−1) | ||||
| GER1 (7.44) | 0.0 | 5.30 ef | 18.33 bc | 2.50 cd | 25.00 e |
| 0.5 | 8.00 ab | 21.67 b | 4.27 a | 25.00 e | |
| 1.0 | 5.00 ef | 13.33 c | 1.47 d | 65.00 c | |
| GER2 (9.14) | 0.0 | 6.00 cdef | 15.00 bc | 2.00 cd | 25.00 e |
| 0.5 | 6.70 bcde | 13.33 c | 2.67 c | 25.00 e | |
| 1.0 | 7.30 abcd | 16.67 bc | 2.03 cd | 50.00 d | |
| GER3 (10.83) | 0.0 | 5.70 cde | 13.33 c | 2.33 cd | 65.00 c |
| 0.5 | 7.70 a | 30.00 a | 4.10 ab | 65.00 c | |
| 1.0 | 4.70 f | 16.67 bc | 2.83 c | 85.00 b | |
| GER4 (62.87) | 0.0 | 6.70 bcde | 18.33 bc | 2.27 cd | 50.00 d |
| 0.5 | 8.70 a | 30.00 a | 3.27 abc | 65.00 c | |
| 1.0 | 6.70 bcde | 18.33 bc | 3.00 bc | 95.00 a | |
| Significance | I | * | ** | ns | *** |
| GER | *** | *** | *** | *** | |
| I × GER | * | ** | * | *** | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohamed, S.M.; El-Mahrouk, M.E.; El-Banna, A.N.; Hafez, Y.M.; El-Ramady, H.; Abdalla, N.; Dobránszki, J. Optimizing Medium Composition and Environmental Culture Condition Enhances Antioxidant Enzymes, Recovers Gypsophila paniculata L. Hyperhydric Shoots and Improves Rooting In Vitro. Plants 2023, 12, 306. https://doi.org/10.3390/plants12020306
Mohamed SM, El-Mahrouk ME, El-Banna AN, Hafez YM, El-Ramady H, Abdalla N, Dobránszki J. Optimizing Medium Composition and Environmental Culture Condition Enhances Antioxidant Enzymes, Recovers Gypsophila paniculata L. Hyperhydric Shoots and Improves Rooting In Vitro. Plants. 2023; 12(2):306. https://doi.org/10.3390/plants12020306
Chicago/Turabian StyleMohamed, Suzan M., Mohammed E. El-Mahrouk, Antar N. El-Banna, Yaser M. Hafez, Hassan El-Ramady, Neama Abdalla, and Judit Dobránszki. 2023. "Optimizing Medium Composition and Environmental Culture Condition Enhances Antioxidant Enzymes, Recovers Gypsophila paniculata L. Hyperhydric Shoots and Improves Rooting In Vitro" Plants 12, no. 2: 306. https://doi.org/10.3390/plants12020306
APA StyleMohamed, S. M., El-Mahrouk, M. E., El-Banna, A. N., Hafez, Y. M., El-Ramady, H., Abdalla, N., & Dobránszki, J. (2023). Optimizing Medium Composition and Environmental Culture Condition Enhances Antioxidant Enzymes, Recovers Gypsophila paniculata L. Hyperhydric Shoots and Improves Rooting In Vitro. Plants, 12(2), 306. https://doi.org/10.3390/plants12020306