Abstract
Auxins are a class of plant hormones playing crucial roles in a plant’s growth, development, and stress responses. Phenylacetic acid (PAA) is a phenylalanine-derived natural auxin found widely in plants. Although the auxin activity of PAA in plants was identified several decades ago, PAA homeostasis and its function remain poorly understood, whereas indole-3-acetic acid (IAA), the most potent auxin, has been used for most auxin studies. Recent studies have revealed unique features of PAA distinctive from IAA, and the enzymes and intermediates of the PAA biosynthesis pathway have been identified. Here, we summarize the occurrence and function of PAA in plants and highlight the recent progress made in PAA homeostasis, emphasizing PAA biosynthesis and crosstalk between IAA and PAA homeostasis.
1. Introduction
Auxins are best known as plant hormones crucial for plant growth, development, and survival. Several endogenous auxins are found in plants [1,2,3]. Among them, indole-3-acetic acid (IAA) has been commonly used for studying auxin’s function and its biosynthesis. Phenylacetic acid (PAA) is a phenylalanine-derived auxin. Although the auxin activity of PAA was demonstrated nearly a century ago and has been detected widely in plants, PAA homeostasis and its function remain poorly understood. Here, we review the occurrence and function of PAA and the recent findings on PAA homeostasis, focusing on PAA biosynthesis.
2. Occurrence of PAA
Auxin activity of PAA was reported in the 1930s, however, PAA was first isolated in plants in the 1960s. Acidic fractions of aqueous extract from etiolated seedlings of Phaseolus were shown to promote the growth of oat coleoptile sections, and further analysis identified that these acidic fractions contained PAA [4]. Subsequently, PAA was detected in various plants. Interestingly, most organs and species accumulate PAA to significantly higher levels than IAA [5,6,7]. In Arabidopsis, the PAA contents range from 200 to 3500 pmol/gFW depending on the organs, which is higher than IAA in most organs, except silique [7,8,9,10,11] (Table 1). Other dicots, such as tomato, pea, sunflower, and tobacco accumulate around 600 to 1600 pmol/gFW of PAA in their shoots, which is also several folds greater than their IAA contents [5,12]. However, the PAA content in Tropaeolum majus (<16 pmol/gFW) is lower than IAA (>70 pmol/gFW) [13]. Monocots and non-vascular plants accumulate 300 to 5000 pmol/gFW of PAA, similar to other dicots [5,7,11,12,14,15] (Table 1). Taken together, PAA is widely distributed in the plant kingdom, and overall, PAA accumulates more than IAA in most plants.

Table 1.
Occurrence of PAA in plants, showing plant tissue, PAA, and IAA contents (if available).
3. Biological Function of PAA
The auxin activity of PAA was identified through three classical auxin activity tests: the pea test, cylinder test, and oat bending test [17]. All three tests revealed PAA has less than 10% of IAA activity [17]. One of the most representative functions of PAA is the promotion of root growth and development [6,8,13,18,19]. PAA induced the root formation of tomato, sunflower, marigold, artichoke, buckwheat, dahlia, and tobacco when applied to the stem [20]. PAA application promoted the formation of adventitious roots of cress hypocotyls, sugar-beet seedlings, and pea epicotyls [21]. Furthermore, leaf explants of Ajuga bracteosa on growth media supplemented with PAA increased the frequency of root induction and biomass [19]. Arabidopsis seedlings treated with PAA increased the formation of lateral roots, although PAA showed a 10- to 20-fold lower activity than IAA [7]. However, some studies showed stronger activity of PAA than IAA. In pea seedlings, PAA induced more lateral root primordia and emerged lateral roots and longer lengths of lateral roots compared to those of IAA [6,18].
PAA impacts the aerial parts of plants as well [7,11,22,23]. Daily exogenous application of PAA on tomato plants for 2 weeks significantly increased tomato height [22]. The PAA application led to epinastic leaves of artichokes [20], stimulated the elongation of wheat coleoptile and moss gametophore [7,23], and promoted the elongation of the Phaseolus internode [24]. Arabidopsis plants with increased PAA production showed elongated hypocotyls and epinastic leaves, similar to those observed in high-IAA Arabidopsis plants [11]. The supplementation of PAA also induced callus formation in tobacco, sunflower, chickpea, and lentil, but the optimal concentration of PAA for tobacco callus induction was 3–4 times higher than IAA [25,26]. Similar to IAA, PAA application to leafless cotyledon retarded the petiole abscission in cotton and inhibited ethylene evolution [16].
PAA, itself, has anti-microbial and anti-fungal activities, and the exogenous application of PAA enhances tolerance to pathogen infections, whereas increased IAA in plants enhances susceptibility to pathogens [27,28,29]. For example, the exogenous application of PAA on citrus inhibits the incidence of fungus molds caused by Penicillium digitatum and P. italicum [30]. Herbivore infestation in maize, poplar, and plumeria increases PAA production [31,32,33]. The application of PAA in oilseed rape enhances the prevention of Sclerotinia sclerotiorum, and the in vitro treatment with PAA demonstrated adverse effects through the disruption of the cell wall and cytoplasm in mycelia [34]. However, any biological role of PAA in a plant’s defense remains unknown.
The first step of auxin action starts from auxin sensing by the auxin receptors (TIR1 and AFBs), which results in the degradation of transcription repressors Aux/IAAs [35]. Shimizu-Mitao and Kakimoto showed PAA-dependent degradation of Aux/IAA [35]. Interaction of Aux/IAAs with auxin receptors, TIR1 or AFB2, leads to the degradation of Aux/IAA [35]. They showed that PAA induced Aux/IAA degradation with lower activity than IAA [35]. Sugawara et al. showed that PAA application rescued the growth defects of IAA-deficient plant, yuc quadruple mutant (yucQ) [7]. In the same study, a yeast two-hybrid assay and a pull-down assay revealed that PAA promoted the interaction of auxin receptors and Aux/IAA in vitro, suggesting that the sensing and signaling modes of PAA are similar to IAA [7].
It is noteworthy, however, that PAA does not engage in polar auxin transport [7,16,36,37,38]. The labeled PAA transport assay using pea epicotyls revealed that the transport of PAA barely occurs in both the basipetal and acropetal directions [36]. PAA applied to the apical bud of intact pea plants did not move in the long-distance basipetal transport [38]. Auxin polar transport inhibitor, naphthylphthalamic acid (NPA), inhibited IAA transport, but NPA did not affect the PAA gradient patterns in cotton, pea, and maize [7,16,37]. Unlike IAA, PAA did not form concentration gradients in response to the gravitropic stimulation via active and directional transport in maize [7]. However, PAA inhibited the IAA polar transport in the internode segments and long-distance movement of the pea apical bud [38].
4. PAA Homeostasis
As auxins regulate a vast array of processes, changes in the content or distribution of auxins can have profound effects on plant growth and development and, in extreme cases, can lead to severe dwarfism or sterility [39,40,41,42]. Auxin homeostasis refers to the spatio-temporal distribution of auxin throughout plant tissues and organs, which governs plant growth and development. Although several processes, including auxin transport, conjugation, and degradation, influence auxin homeostasis, de novo biosynthesis directly affects the local concentration of auxins.
4.1. PAA Biosynthesis
The main route of IAA biosynthesis from tryptophan in plants is the YUCCA pathway [43,44,45,46,47,48,49,50,51,52] (Figure 1a). The first step of this pathway is the conversion of tryptophan to indole-3-pyruvate (IPA) by the enzymes belonging to the Tryptophan Aminotransferase of Arabidopsis (TAA) family [53,54,55,56]. Then, flavin-containing mono-oxygenases belonging to the YUCCA (YUC) family convert the IPA to IAA [42,55,57,58,59,60]. This pathway is also believed to contribute towards PAA biosynthesis (Figure 1b), as several studies have shown that the TAA and YUC enzymes can convert phenylalanine to phenylpyruvate (PPA) and PPA to PAA in vitro, respectively [7,54,59,61,62]. YUCCA overexpression increases endogenous PAA or PAA conjugate content [7]. However, the TAA and YUC mutants sometimes show little or no change in the PAA content, despite the significant alterations to the IAA content. For example, the maize YUC1-deficient mutant de18 has an over 90% reduction in free and total IAA contents, while the PAA content is not significantly affected, and the tar2-1 pea mutant displays a near complete loss of free IAA but no change in the PAA levels [61]. Similarly, in Arabidopsis, the yuc1yuc2yuc6 triple and yuc3yuc5yuc7yuc8yuc9 quintuple mutants have wild-type levels of PAA but 40–50% reductions in the IAA content [7]. However, the TAA1 deficient mutant wei8-1 displays a 20% and 80% reduction in PAA and IAA, respectively [7]. These findings suggest that these enzymes may function in the PAA biosynthesis in plants but may not be major players in the PAA biosynthesis in some species [35].
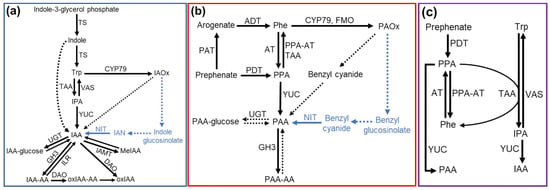
Figure 1.
Schematic diagrams of IAA biosynthesis: (a) PAA biosynthesis, (b) and a link between IAA and PAA homeostasis in plants (c). The pathways only include steps and enzymes that are demonstrated via genetic evidence. Solid arrows represent single reactions catalyzed by known enzymes, and dotted arrows represent predicted single or multiple steps. Blue arrows and metabolites represent pathways and metabolites present only in Brassicales species. IAA, indole-3-acetic acid; IAA-AA, amino acid-conjugated IAA; IAN, indole-3-acetonitrile; IAOx, indole-3-acetaldoxime; IPA, indole-3-pyruvate; MeIAA, methyl-conjugated IAA; oxIAA, 2-oxindole-3-acetic acid; oxIAA-AA, amino acid-conjugated oxIAA; PAA, phenylacetic acid; PAA-AA, amino acid-conjugated PAA; PAOx, phenylacetaldoxime; Phe, phenylalanine; PPA, phenylpyruvate; Trp, tryptophan; AT, amino transferase; CYP79, cytochrome P450 mono-oxygenase of the 79 family; DAO, dioxygenase for auxin oxidation; FMO, flavin-containing mono-oxygenase; GH3, Gretchen Hagen 3 auxin-amido synthetase; ILR, IAA-Leu Resistant IAA-Amino hydrolase; IAMT, IAA carboxymethyltransferase; NIT, nitrilase; PAT, prephenate aminotransferase; PDT, prephenate dehydratase; PPA-AT, phenylpyruvate aminotransferase; TAA, tryptophan aminotransferase of Arabidopsis; TS, tryptophan synthase; UGT, UDP-glucuronosyltransferase; YUC, YUCCA family of flavin-containing mono-oxygenase; VAS; methionine aminotransferase.
In plants and prokaryotes, phenylalanine biosynthesis from prephenate occurs through two routes: transamination of prephenate to arogenate, which is subsequently decarboxylated and dehydrated into phenylalanine, or decarboxylation and dehydration of prephenate to form phenylpyruvate (PPA), which is converted to phenylalanine through transamination [63,64,65] (Figure 1c). Generally, the PPA pathway is more commonly found in prokaryotes, while the arogenate pathway is the major route of phenylalanine biosynthesis in plants; however, there is evidence for both pathways existing and making significant contributions in several plant and bacterial species [63,64,65]. Thus, the modification of the PPA metabolism may impact PAA biosynthesis not only directly by being converted to PAA but also indirectly by affecting phenylalanine pools in plants (Figure 1c).
Aoi et al. demonstrated that arogenate dehydratase (ADT), the enzyme that catalyzes the conversion of arogenate to phenylalanine, affects PAA biosynthesis, as the overexpression or knockout of ADTs result in increased or decreased PAA contents, respectively [10]. As arogenate serves as a precursor of phenylalanine, increased ADT activity results in increases in phenylalanine-derived metabolites, such as PAA. They also detected changes in the PPA levels upon the modulation of ADT activity that followed the trends measured for the PAA content (i.e., ADT overexpression resulted in increased levels of PAA and PPA) [10], supporting the role of PPA as a precursor of PAA.
Another PAA biosynthesis pathway is the aldoxime pathway, using phenylalanine-derived aldoxime, phenylacetaldoxime (PAOx), as an intermediate [8,10,11,15,66]. Aldoximes, such as indole-3-acetaldoxime (IAOx) and PAOx, as well as the aldoximes derived from other amino acids, are well characterized as precursors of various specialized metabolites, such as glucosinolates, cyanogenic glycosides, and camalexin as well as nitrogenous volatiles [31,33,67,68,69,70,71,72]. Several studies have shown that IAA can be made from IAOx in Brassicales and monocots [11,73,74,75] (Figure 1a). Similarly, it was shown that PAOx is a precursor of PAA in Arabidopsis and monocots [8,11,15] (Figure 1b). Arabidopsis plants overproducing PAOx increase PAA and display altered morphology, such as epinasty leaves and elongated hypocotyls, similar to those shown in plants with increased IAA [8,11,42]. Maize and sorghum plants fed with labeled PAOx produce labeled PAA [11,15]. These findings indicate a wider distribution of the aldoxime-derived auxin biosynthesis pathway throughout the plant kingdom rather than being limited to Brassicales.
The first step of the aldoxime pathway is the production of IAOx or PAOx by the cytochrome P450 mono-oxygenases of the 79 family (CYP79s) [8,11,15,32,66]. In addition to the CYP79 enzymes, flavin-containing mono-oxygenases (FMOs) in two fern species have been shown to catalyze the conversion of phenylalanine to PAOx [76]. Aldoximes contribute to auxin pools through two routes (Figure 1a,b). In Brassicales plants, both IAOx and PAOx are precursors of family-specific glucosinolates. Glucosinolates and their degradation enzyme beta-thiol-glucosidases, known as myrosinases, are stored in separate cellular and subcellular compartments [77,78,79,80]. However, upon herbivore or pathogen attack, these compartments are compromised, resulting in glucosinolate hydrolysis by myrosinases and the rapid release of toxic metabolites, such as isothiocyanates, nitriles and epithionitriles [81,82,83]. Glucosinolate turnover has also been shown to occur in vivo without tissue damage or disruption [84,85,86,87]. Nitriles, such as indole-3-acetonitrile (IAN) and benzyl cyanide, are byproducts of glucosinolate degradation. These nitriles can then be acted upon by nitrilase enzymes to generate IAA and PAA [13,88,89,90,91,92]. On the other hand, aldoximes can be converted to auxins through the aldoxime-derived auxin pathway, which is glucosinolate-independent and is present in both the Brassicales and non-Brassicales species as maize and sorghum [11,15]. A recent study demonstrated that benzyl cyanide serves as an intermediate of PAOx-derived PAA biosynthesis in maize and sorghum [15]. Application of benzyl cyanide increases PAA in maize and sorghum, and both species convert labeled benzyl cyanide to labeled PAA [15], suggesting that nitriles may be key intermediates in both routes of the aldoxime pathway (Figure 1a,b). Unlike the YUCCA pathway, the aldoxime-derived auxin pathways do not appear to be the main route of auxin biosynthesis under normal growth conditions. For example, the Arabidopsis IAOx deficient mutant, cyp79b2 cyp79b3 (b2b3) double mutant, grows normally under optimal temperatures [73], and the CYP79A2 gene encoding the PAOx production enzyme is barely expressed in the vegetative tissue of Arabidopsis ecotype Col-0 [11]. However, at high temperatures and under salt stress, the b2b3 mutant displays a low auxin growth phenotype [73,93], suggesting that the aldoxime pathway contributes significantly towards stress-induced auxin production. Indeed, many CYP79 enzymes have their expression induced by stressors such as herbivory or by treatment with stress hormones such as jasmonic acid [31,32,68,72,94]. Given that aldoximes often serve as precursors of defense metabolites, the aldoxime pathway may play a role in modulating plant growth during the defense response.
Aside from PPA, PAOx, and benzyl cyanide, several other metabolites have been implicated in PAA biosynthesis, although where they fit within the known biosynthetic pathways is unclear. Several labeled feeding experiments have demonstrated that phenylacetaldehyde (PAAld) is derived from the phenylalanine metabolism and produced along with labeled PAA [5,95]. PAAld biosynthesis from PPA has been shown to occur in the rose through the actions of phenylpyruvate decarboxylases [96]. Additionally, in roses as well as other species, PAAld has been shown to be directly synthesized from phenylalanine by the action of aromatic aldehyde synthases or aromatic amino acid decarobylases [96,97,98,99,100]. Once PAAld is synthesized, it can potentially be acted upon by the aldehyde oxidases to generate PAA, which has been demonstrated to have activity for PAAld in maize [101]. Another metabolite, 4-phenylbutyric acid (4PBA), was recently shown to display an auxin-like effect during plant regeneration via conversion to PAA using a mechanism independent of IBR3-catalyzed oxidation [102]. Further study showing altered PAA contents upon the removal of intermediate biosynthesis enzymes may reveal the role(s) that these metabolites play in PAA biosynthesis.
4.2. PAA Inactivation
Another major facet of auxin homeostasis is the conversion of active auxins to inactive forms. The inactivation of auxin not only supports the formation of auxin gradients and the maintenance of auxin levels but is also necessary to prevent cytotoxic levels of auxins from accumulating in cells. IAA inactivation proceeds through two pathways: reversible IAA conjugation (to glucose, methyl, or amino acids) and irreversible IAA conjugation (to amino acids) and oxidation, with recent findings demonstrating that amino acid-conjugated IAA is oxidized and then subsequently hydrolyzed to form oxidized IAA [103,104]. As with biosynthesis, knowledge of PAA inactivation is limited compared to our understanding of IAA inactivation. Multiple studies have demonstrated that some Gretchen Hagen 3 IAA-amido synthetase (GH3) and UDP-glucuronosyltransferase (UGT) enzymes have activity towards PAA to generate PAA conjugates, such as PAA-asp, PAA-glu, and PAA-glucose, respectively [7,105,106,107,108] (Figure 1b).
4.3. Metabolic Interaction between IAA and PAA
More recent studies have shown a link between IAA and PAA homeostasis. The homeostasis of IAA and PAA was shown to be maintained through the modulation of auxin conjugation, with the accumulation of PAA resulting in the induction of GH3 or UGT genes that preferentially act upon IAA and vice versa [8,9]. Lynch et al. showed that PPA could, in addition to its previously defined and proposed impacts of PAA biosynthesis, impact IAA biosynthesis by serving as an amino acceptor in the TAA-catalyzed conversion of tryptophan to IPA [109]. This interaction not only promotes the production of IAA but may also impact PAA biosynthesis, as an increased flux through the PPA route of phenylalanine biosynthesis was shown to decrease steady-state levels of phenylalanine [109] (Figure 1c). Perez et al. demonstrated that the accumulation of PAA results in the transcriptional downregulation of genes related to tryptophan and IAA biosynthesis in Arabidopsis [11], demonstrating a complex regulatory network for maintaining auxin homeostasis.
5. Conclusions
In the past decades, several biochemical and genetic studies have identified key metabolites, enzymes, and pathways that contribute towards IAA metabolism. While many questions remain regarding PAA homeostasis, recent studies have greatly expanded our understanding of how PAA is synthesized and inactivated. The role of PPA as a metabolite linking together phenylalanine, IAA, and PAA biosynthesis has been supported by genetic studies and suggests that the PPA-derived PAA biosynthesis is more complex than the corresponding IAA biosynthetic pathway. Meanwhile, the occurrence of PAOx as a PAA precursor in Brassicales and monocots suggests that this hidden pathway may be distributed widely in the plant kingdom and contribute towards PAA homeostasis wherever PAOx is produced. Additionally, the identification of PAA-amino acid and PAA-glucose conjugates within Arabidopsis has provided mechanisms for PAA inactivation, which may be shared among other species and may employ similar pathways used in IAA inactivation. Future investigation is needed to reveal other potential PAA biosynthesis and inactivation pathways, as well as the physiological roles of these pathways.
Author Contributions
V.C.P., H.Z. and M.L. made tables and figures. V.C.P., H.Z., M.L. and J.K. designed and wrote the review. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Science Foundation CAREER Award (IOS-2142898).
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Simon, S.; Petrášek, J. Why Plants Need More than One Type of Auxin. Plant Sci. 2011, 180, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Damodaran, S.; Strader, L.C. Indole 3-Butyric Acid Metabolism and Transport in Arabidopsis thaliana. Front. Plant Sci. 2019, 10, 851. [Google Scholar] [CrossRef] [PubMed]
- Enders, T.A.; Strader, L.C. Auxin Activity: Past, Present, and Future. Am. J. Bot. 2015, 102, 180–196. [Google Scholar] [CrossRef]
- Okamoto, T.; Koizumi, T.; Isogai, Y. Studies on Plant Growth Regulators. II. Isolation of Indole-3-Acetic Acid, Phenylacetic Acid, and Several Plant Growth Inhibitors from Etiolated Seedlings of Phaseolus. Chem. Pharm. Bull. 1967, 15, 159–163. [Google Scholar] [CrossRef]
- Wightman, F.; Lighty, D.L. Identification of Phenylacetic Acid as a Natural Auxin in the Shoots of Higher Plants. Physiol. Plant. 1982, 55, 17–24. [Google Scholar] [CrossRef]
- Schneider, E.A.; Kazakoff, C.W.; Wightman, F. Gas Chromatography-Mass Spectrometry Evidence for Several Endogenous Auxins in Pea Seedling Organs. Planta 1985, 165, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, S.; Mashiguchi, K.; Tanaka, K.; Hishiyama, S.; Sakai, T.; Hanada, K.; Kinoshita-Tsujimura, K.; Yu, H.; Dai, X.; Takebayashi, Y.; et al. Distinct Characteristics of Indole-3-Acetic Acid and Phenylacetic Acid, Two Common Auxins in Plants. Plant Cell Physiol. 2015, 56, 1641–1654. [Google Scholar] [CrossRef]
- Aoi, Y.; Tanaka, K.; Cook, S.D.; Hayashi, K.I.; Kasahara, H. GH3 Auxin-Amido Synthetases Alter the Ratio of Indole-3-Acetic Acid and Phenylacetic Acid in Arabidopsis. Plant Cell Physiol. 2020, 61, 596–605. [Google Scholar] [CrossRef]
- Aoi, Y.; Hira, H.; Hayakawa, Y.; Liu, H.; Fukui, K.; Dai, X.; Tanaka, K.; Hayashi, K.I.; Zhao, Y.; Kasahara, H. UDP-Glucosyltransferase UGT84B1 Regulates the Levels of Indole-3-Acetic Acid and Phenylacetic Acid in Arabidopsis. Biochem. Biophys. Res. Commun. 2020, 532, 244–250. [Google Scholar] [CrossRef]
- Aoi, Y.; Oikawa, A.; Sasaki, R.; Huang, J.; Hayashi, K.; Kasahara, H. Arogenate Dehydratases Can Modulate the Levels of Phenylacetic Acid in Arabidopsis. Biochem. Biophys. Res. Commun. 2020, 524, 83–88. [Google Scholar] [CrossRef]
- Perez, V.C.; Dai, R.; Bai, B.; Tomiczek, B.; Askey, B.C.; Zhang, Y.; Rubin, G.M.; Ding, Y.; Grenning, A.; Block, A.K.; et al. Aldoximes Are Precursors of Auxins in Arabidopsis and Maize. New Phytol. 2021, 231, 1449–1461. [Google Scholar] [CrossRef]
- Wightman, F.; Rauthan, B.S. Evidence for the Biosynthesis and Natural Occurrence of the Auxin, Phenylacetic Acid, in Shoots of Higher Plants. In Plant Growth Substances, Proceedings of the International Conference; Hirokawa: Tokyo, Japan, 1974; pp. 15–27. [Google Scholar]
- Ludwig-Müller, J.; Cohen, J.D. Identification and Quantification of Three Active Auxins in Different Tissues of Tropaeolum Majus. Physiol. Plant. 2002, 115, 320–329. [Google Scholar] [CrossRef]
- Schneider, E.A.; Wightman, F. Auxins of Non-Flowering Plants. I. Occurrence of 3-Indoleacetic Acid and Phenylacetic Acid in Vegetative and Fertile Fronds of the Ostrich Fern (Matteucia struthiopteris). Physiol. Plant. 1986, 68, 396–402. [Google Scholar] [CrossRef]
- Perez, V.C.; Dai, R.; Tomiczek, B.; Mendoza, J.; Wolf, E.S.A.; Grenning, A.; Vermerris, W.; Block, A.K.; Kim, J. Metabolic Link between Auxin Production and Specialized Metabolites in Sorghum Bicolor. J. Exp. Bot. 2022, 74, 364–376. [Google Scholar] [CrossRef] [PubMed]
- Suttle, J.C.; Mansager, E.R. The Physiological Significance of Phenylacetic Acid in Abscising Cotton Cotyledons. Plant Physiol. 1986, 81, 434–438. [Google Scholar] [CrossRef]
- Haagen-Smit, S.A.J.; Went, F.W. A Physiological Analysis of Growth Substances. Proc. K. Ned. Akad. Van Wet. Amst. 1935, 38, 852–857. [Google Scholar]
- Wightman, F.; Schneider, E.A.; Thimann, K.V. Hormonal Factors Controlling the Initiation and Development of Lateral Roots. Physiol. Plant. 1980, 49, 304–314. [Google Scholar] [CrossRef]
- Saeed, S.; Ali, H.; Khan, T.; Kayani, W.; Khan, M.A. Impacts of Methyl Jasmonate and Phenyl Acetic Acid on Biomass Accumulation and Antioxidant Potential in Adventitious Roots of Ajuga Bracteosa Wall Ex Benth., a High Valued Endangered Medicinal Plant. Physiol. Mol. Biol. Plants 2017, 23, 229–237. [Google Scholar] [CrossRef]
- Zimmerman, P.W.; Wilcoxon, F. Several Chemical Growth Substances Which Cause Initiation of Roots and Other Responses in Plants. Contrib. Boyce Thompson Inst. 1935, 7, 209–229. [Google Scholar]
- Wheeler, A. Auxin-like Growth Activity of Phenylacetonitrile. Ann. Bot. 1977, 41, 867–872. [Google Scholar] [CrossRef]
- Pearse, H.L. Effect of Phenylacetic Acid on the Growth of Tomato Plants. Nature 1936, 138, 363–364. [Google Scholar] [CrossRef]
- Abe, H.; Uchiyama, M.; Sato, R. Isolation of Phenylacetic Acid and Its p -Hydroxy Derivative as Auxin-like Substances from Undaria Pinnatifida. Agric. Biol. Chem. 1974, 38, 897–898. [Google Scholar] [CrossRef]
- Small, D.K.; Morris, D.A. Promotion of Elongation and Acid Invertase Activity in Phaseolus Vulgaris L. Internode Segments by Phenylacetic Acid. Plant Growth Regul. 1990, 9, 329–340. [Google Scholar] [CrossRef]
- Milborrow, B.V.; Purse, J.G.; Wightman, F. On the Auxin Activity of Phenylacetic Acid. Ann. Bot. 1975, 39, 1143–1146. [Google Scholar]
- Leuba, V.; LeTourneau, D. Auxin Activity of Phenylacetic Acid in Tissue Culture. J. Plant Growth Regul. 1990, 9, 71–76. [Google Scholar] [CrossRef]
- Chen, Z.; Agnew, J.L.; Cohen, J.D.; He, P.; Shan, L.; Sheen, J.; Kunkel, B.N. Pseudomonas Syringae Type III Effector AvrRpt2 Alters Arabidopsis Thaliana Auxin Physiology. Proc. Natl. Acad. Sci. USA 2007, 104, 20131–20136. [Google Scholar] [CrossRef] [PubMed]
- Mutka, A.M.; Fawley, S.; Tsao, T.; Kunkel, B.N. Auxin Promotes Susceptibility to Pseudomonas Syringae via a Mechanism Independent of Suppression of Salicylic Acid-Mediated Defenses. Plant J. 2013, 74, 746–754. [Google Scholar] [CrossRef]
- Kunkel, B.N.; Harper, C.P. The Roles of Auxin during Interactions between Bacterial Plant Pathogens and Their Hosts. J. Exp. Bot. 2018, 69, 245–254. [Google Scholar] [CrossRef]
- Liu, P.; Cheng, Y.; Yang, M.; Liu, Y.; Chen, K.; Long, C.; Deng, X. Mechanisms of Action for 2-Phenylethanol Isolated from Kloeckera Apiculata in Control of Penicillium Molds of Citrus Fruits. BMC Microbiol. 2014, 14, 242. [Google Scholar] [CrossRef]
- Irmisch, S.; McCormick, A.C.; Boeckler, G.A.; Schmidt, A.; Reichelt, M.; Schneider, B.; Block, K.; Schnitzler, J.P.; Gershenzon, J.; Unsicker, S.B.; et al. Two Herbivore-Induced Cytochrome P450 Enzymes CYP79D6 and CYP79D7 Catalyze the Formation of Volatile Aldoximes Involved in Poplar Defense. Plant Cell 2013, 25, 4737–4754. [Google Scholar] [CrossRef]
- Irmisch, S.; Zeltner, P.; Handrick, V.; Gershenzon, J.; Köllner, T.G. The Maize Cytochrome P450 CYP79A61 Produces Phenylacetaldoxime and Indole-3-Acetaldoxime in Heterologous Systems and Might Contribute to Plant Defense and Auxin Formation. BMC Plant Biol. 2015, 15, 128. [Google Scholar] [CrossRef] [PubMed]
- Dhandapani, S.; Jin, J.; Sridhar, V.; Chua, N.H.; Jang, I.C. CYP79D73 Participates in Biosynthesis of Floral Scent Compound 2-Phenylethanol in Plumeria rubra. Plant Physiol. 2019, 180, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Cheng, Q.; Wang, X.; Jia, W.; Xie, J.; Fan, G.; Han, C.; Zhao, X. Selenium Improved Phenylacetic Acid Content in Oilseed Rape and Thus Enhanced the Prevention of Sclerotinia Sclerotiorum by Dimethachlon. J. Fungi 2022, 8, 1193. [Google Scholar] [CrossRef] [PubMed]
- Shimizu-Mitao, Y.; Kakimoto, T. Auxin Sensitivities of All Arabidopsis Aux/IAAs for Degradation in the Presence of Every TIR1/AFB. Plant Cell Physiol. 2014, 55, 1450–1459. [Google Scholar] [CrossRef]
- ProchÁzka, S.; Borkovec, V. Transport and Regulative Properties of Phenylacetic Acid. Biol. Plant. 1984, 26, 358–363. [Google Scholar] [CrossRef]
- Johnson, C.F.; Morris, D.A. Regulation of Auxin Transport in Pea (Pisum sativum L.) by Phenylacetic Acid: Effects on the Components of Transmembrane Transport of Indol-3yl-Acetic Acid. Planta 1987, 172, 400–407. [Google Scholar] [CrossRef]
- Morris, D.A.; Johnson, C.F. Regulation of Auxin Transport in Pea (Pisum sativum L.) by Phenylacetic Acid: Inhibition of Polar Auxin Transport in Intact Plants and Stem Segments. Planta 1987, 172, 408–416. [Google Scholar] [CrossRef]
- Boerjan, W.; Cervera, M.T.; Delarue, M.; Beeckman, T.; Dewitte, W.; Bellini, C.; Caboche, M.; Onckelen, H.V.; Montagu, M.V.; Inzé, D. Superroot, a Recessive Mutation in Arabidopsis, Confers Auxin Overproduction. Plant Cell 1995, 7, 1405–1419. [Google Scholar] [CrossRef]
- Seo, M.; Akaba, S.; Oritani, T.; Delarue, M.; Bellini, C.; Caboche, M.; Koshiba, T. Higher Activity of an Aldehyde Oxidase in the Auxin-Overproducing Superroot1 Mutant of Arabidopsis thaliana. Plant Physiol. 1998, 116, 687–693. [Google Scholar] [CrossRef]
- van der Graaff, E.; Boot, K.; Granbom, R.; Sandberg, G.; Hooykaas, P.J.J. Increased Endogenous Auxin Production in Arabidopsis thaliana Causes Both Earlier Described and Novel Auxin-Related Phenotypes. J. Plant Growth Regul. 2003, 22, 240–252. [Google Scholar] [CrossRef]
- Kim, J.I.; Sharkhuu, A.; Jin, J.B.; Li, P.; Jeong, J.C.; Baek, D.; Lee, S.Y.; Blakeslee, J.J.; Murphy, A.S.; Bohnert, H.J.; et al. Yucca6, a Dominant Mutation in Arabidopsis, Affects Auxin Accumulation and Auxin-Related Phenotypes. Plant Physiol. 2007, 145, 722–735. [Google Scholar] [CrossRef] [PubMed]
- Woo, Y.M.; Park, H.J.; Su’udi, M.; Yang, J.I.; Park, J.J.; Back, K.; Park, Y.M.; An, G. Constitutively Wilted 1, a Member of the Rice YUCCA Gene Family, Is Required for Maintaining Water Homeostasis and an Appropriate Root to Shoot Ratio. Plant Mol. Biol. 2007, 65, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Kamiya, N.; Morinaka, Y.; Matsuoka, M.; Sazuka, T. Auxin Biosynthesis by the YUCCA Genes in Rice. Plant Physiol. 2007, 143, 1362–1371. [Google Scholar] [CrossRef] [PubMed]
- Gallavotti, A.; Barazesh, S.; Malcomber, S.; Hall, D.; Jackson, D.; Schmidt, R.J.; McSteen, P. Sparse Inflorescence1 Encodes a Monocot-Specific YUCCA-like Gene Required for Vegetative and Reproductive Development in Maize. Proc. Natl. Acad. Sci. USA 2008, 105, 15196–15201. [Google Scholar] [CrossRef] [PubMed]
- Abu-Zaitoon, Y.M.; Bennett, K.; Normanly, J.; Nonhebel, H.M. A Large Increase in IAA during Development of Rice Grains Correlates with the Expression of Tryptophan Aminotransferase OsTAR1 and a Grain-Specific YUCCA. Physiol. Plant. 2012, 146, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Bernardi, J.; Lanubile, A.; Li, Q.-B.; Kumar, D.; Kladnik, A.; Cook, S.D.; Ross, J.J.; Marocco, A.; Chourey, P.S. Impaired Auxin Biosynthesis in the Defective Endosperm18 Mutant Is Due to Mutational Loss of Expression in the ZmYuc1 Gene Encoding Endosperm-Specific YUCCA1 Protein in Maize. Plant Physiol. 2012, 160, 1318–1328. [Google Scholar] [CrossRef]
- Bernardi, J.; Li, Q.B.; Gao, Y.; Zhao, Y.; Battaglia, R.; Marocco, A.; Chourey, P.S. The Auxin-Deficient Defective Kernel18 (Dek18) Mutation Alters the Expression of Seed-Specific Biosynthetic Genes in Maize. J Plant Growth Regul. 2016, 35, 770–777. [Google Scholar] [CrossRef]
- Kakei, Y.; Nakamura, A.; Yamamoto, M.; Ishida, Y.; Yamazaki, C.; Sato, A.; Narukawa-Nara, M.; Soeno, K.; Shimada, Y. Biochemical and Chemical Biology Study of Rice OsTAR1 Revealed That Tryptophan Aminotransferase Is Involved in Auxin Biosynthesis: Identification of a Potent OsTAR1 Inhibitor, Pyruvamine2031. Plant Cell Physiol. 2017, 58, 598–606. [Google Scholar] [CrossRef]
- Poulet, A.; Kriechbaumer, V. Bioinformatics Analysis of Phylogeny and Transcription of TAA/YUC Auxin Biosynthetic Genes. Int. J. Mol. Sci. 2017, 18, 1791. [Google Scholar] [CrossRef]
- Shao, A.; Ma, W.; Zhao, X.; Hu, M.; He, X.; Teng, W.; Li, H.; Tong, Y. The Auxin Biosynthetic TRYPTOPHAN AMINOTRANSFERASE RELATED TaTAR2.1-3A Increases Grain Yield of Wheat. Plant Physiol. 2017, 174, 2274–2288. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Yang, H.; Shang, C.; Ma, S.; Liu, L.; Cheng, J. The Roles of Auxin Biosynthesis YUCCA Gene Family in Plants. Int. J. Mol. Sci. 2019, 20, 6343. [Google Scholar] [CrossRef] [PubMed]
- Stepanova, A.N.; Robertson-Hoyt, J.; Yun, J.; Benavente, L.M.; Xie, D.Y.; Doležal, K.; Schlereth, A.; Jürgens, G.; Alonso, J.M. TAA1-Mediated Auxin Biosynthesis Is Essential for Hormone Crosstalk and Plant Development. Cell 2008, 133, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Ferrer, J.L.; Ljung, K.; Pojer, F.; Hong, F.; Long, J.A.; Li, L.; Moreno, J.E.; Bowman, M.E.; Ivans, L.J.; et al. Rapid Synthesis of Auxin via a New Tryptophan-Dependent Pathway Is Required for Shade Avoidance in Plants. Cell 2008, 133, 164–176. [Google Scholar] [CrossRef]
- Won, C.; Shen, X.; Mashiguchi, K.; Zheng, Z.; Dai, X.; Cheng, Y.; Kasahara, H.; Kamiya, Y.; Chory, J.; Zhao, Y. Conversion of Tryptophan to Indole-3-Acetic Acid by TRYPTOPHAN AMINOTRANSFERASES OF ARABIDOPSIS and YUCCAs in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 18518–18523. [Google Scholar] [CrossRef]
- Wang, Q.; Qin, G.; Cao, M.; Chen, R.; He, Y.; Yang, L.; Zeng, Z.; Yu, Y.; Gu, Y.; Xing, W.; et al. A Phosphorylation-Based Switch Controls TAA1-Mediated Auxin Biosynthesis in Plants. Nat. Commun. 2020, 11, 679. [Google Scholar] [CrossRef]
- Zhao, Y.; Christensen, S.K.; Fankhauser, C.; Cashman, J.R.; Cohen, J.D.; Weigel, D.; Chory, J. A Role for Flavin Monooxygenase-Like Enzymes in Auxin Biosynthesis. Science 2001, 291, 306–309. [Google Scholar] [CrossRef]
- Stepanova, A.N.; Yun, J.; Robles, L.M.; Novak, O.; He, W.; Guo, H.; Ljung, K.; Alonso, J.M. The Arabidopsis YUCCA1 Flavin Monooxygenase Functions in the Indole-3-Pyruvic Acid Branch of Auxin Biosynthesis. Plant Cell 2011, 23, 3961–3973. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Mashiguchi, K.; Chen, Q.; Kasahara, H.; Kamiya, Y.; Ojha, S.; DuBois, J.; Ballou, D.; Zhao, Y. The Biochemical Mechanism of Auxin Biosynthesis by an Arabidopsis YUCCA Flavin-Containing Monooxygenase. J. Biol. Chem. 2013, 288, 1448–1457. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, W.; Zuo, Y.; Zhu, L.; Hastwell, A.H.; Chen, L.; Tian, Y.; Su, C.; Ferguson, B.J.; Li, X. GmYUC2a Mediates Auxin Biosynthesis during Root Development and Nodulation in Soybean. J. Exp. Bot. 2019, 70, 3165–3176. [Google Scholar] [CrossRef]
- Cook, S.D.; Nichols, D.S.; Smith, J.; Chourey, P.S.; McAdam, E.L.; Quittenden, L.; Ross, J.J. Auxin Biosynthesis: Are the Indole-3-Acetic Acid and Phenylacetic Acid Biosynthesis Pathways Mirror Images? Plant Physiol. 2016, 171, 1230–1241. [Google Scholar]
- Cook, S.D. An Historical Review of Phenylacetic Acid. Plant Cell Physiol. 2019, 60, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Graindorge, M.; Giustini, C.; Kraut, A.; Moyet, L.; Curien, G.; Matringe, M. Three Different Classes of Aminotransferases Evolved Prephenate Aminotransferase Functionality in Arogenate-Competent Microorganisms. J. Biol. Chem. 2014, 289, 3198–3208. [Google Scholar] [CrossRef] [PubMed]
- Pascual, M.B.; El-Azaz, J.; de la Torre, F.N.; Cañas, R.A.; Avila, C.; Cánovas, F.M. Biosynthesis and Metabolic Fate of Phenylalanine in Conifers. Front. Plant Sci. 2016, 7, 1030. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Lynch, J.H.; Guo, L.; Rhodes, D.; Morgan, J.A.; Dudareva, N. Completion of the Cytosolic Post-Chorismate Phenylalanine Biosynthetic Pathway in Plants. Nat. Commun. 2019, 10, 15. [Google Scholar] [CrossRef]
- Wittstock, U.; Halkier, B.A. Cytochrome P450 CYP79A2 from Arabidopsis thaliana L. Catalyzes the Conversion of L-Phenylalanine to Phenylacetaldoxime in the Biosynthesis of Benzylglucosinolate. J. Biol. Chem. 2000, 275, 14659–14666. [Google Scholar] [CrossRef] [PubMed]
- Glawischnig, E.; Hansen, B.G.; Olsen, C.E.; Halkier, B.A. Camalexin Is Synthesized from Indole-3-Acetaldoxime, a Key Branching Point between Primary and Secondary Metabolism in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 8245–8250. [Google Scholar] [CrossRef]
- Luck, K.; Jirschitzka, J.; Irmisch, S.; Huber, M.; Gershenzon, J.; Köllner, T.G. CYP79D Enzymes Contribute to Jasmonic Acid-Induced Formation of Aldoximes and Other Nitrogenous Volatiles in Two Erythroxylum Species. BMC Plant Biol. 2016, 16, 215. [Google Scholar] [CrossRef]
- Kuwahara, Y.; Asano, Y. Generation of (2-Nitroethyl) Benzene and Related Benzenoids from L-Phenylalanine; Flower Scents of the Japanese Loquat Eriobotrya Japonica [Rosales: Rosaceae]. Biosci. Biotechnol. Biochem. 2018, 82, 1855–1866. [Google Scholar] [CrossRef]
- Sørensen, M.; Neilson, E.H.J.; Møller, B.L. Oximes: Unrecognized Chameleons in General and Specialized Plant Metabolism. Mol. Plant 2018, 11, 95–117. [Google Scholar] [CrossRef]
- Lai, D.; Maimann, A.B.; Macea, E.; Ocampo, C.H.; Cardona, G.; Pičmanová, M.; Darbani, B.; Olsen, C.E.; Debouck, D.; Raatz, B.; et al. Biosynthesis of Cyanogenic Glucosides in Phaseolus Lunatus and the Evolution of Oxime-Based Defenses. Plant Direct 2020, 4, e00244. [Google Scholar] [CrossRef]
- Liao, Y.; Zeng, L.; Tan, H.; Cheng, S.; Dong, F.; Yang, Z. Biochemical Pathway of Benzyl Nitrile Derived from L-Phenylalanine in Tea (Camellia sinensis) and Its Formation in Response to Postharvest Stresses. J. Agric. Food Chem. 2020, 68, 1397–1404. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Hull, A.K.; Gupta, N.R.; Goss, K.A.; Alonso, J.; Ecker, J.R.; Normanly, J.; Chory, J.; Celenza, J.L. Trp-Dependent Auxin Biosynthesis in Arabidopsis: Involvement of Cytochrome P450s CYP79B2 and CYP79B3. Genes Dev. 2002, 16, 3100–3112. [Google Scholar] [CrossRef]
- Sugawara, S.; Hishiyama, S.; Jikumaru, Y.; Hanada, A.; Nishimura, T.; Koshiba, T.; Zhao, Y.; Kamiya, Y.; Kasahara, H. Biochemical Analyses of Indole-3-Acetaldoxime-Dependent Auxin Biosynthesis in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 5430–5435. [Google Scholar] [CrossRef]
- Zhang, D.; Song, Y.H.; Dai, R.; Lee, T.G.; Kim, J. Aldoxime Metabolism Is Linked to Phenylpropanoid Production in Camelina Sativa. Front. Plant Sci. 2020, 11, 10.3389/fpls.2020.00017. [Google Scholar] [CrossRef] [PubMed]
- Thodberg, S.; Sørensen, M.; Bellucci, M.; Crocoll, C.; Bendtsen, A.K.; Nelson, D.R.; Motawia, M.S.; Møller, B.L.; Neilson, E.H.J. A Flavin-Dependent Monooxygenase Catalyzes the Initial Step in Cyanogenic Glycoside Synthesis in Ferns. Commun. Biol. 2020, 3, 507. [Google Scholar] [CrossRef] [PubMed]
- Koroleva, O.A.; Davies, A.; Deeken, R.; Thorpe, M.R.; Tomos, A.D.; Hedrich, R. Identification of a New Glucosinolate-Rich Cell Type in Arabidopsis Flower Stalk. Plant Physiol. 2000, 124, 599–608. [Google Scholar] [CrossRef]
- Koroleva, O.A.; Cramer, R. Single-Cell Proteomic Analysis of Glucosinolate-Rich S-Cells in Arabidopsis thaliana. Methods 2011, 54, 413–423. [Google Scholar] [CrossRef]
- Shroff, R.; Schramm, K.; Jeschke, V.; Nemes, P.; Vertes, A.; Gershenzon, J.; Svatoš, A. Quantification of Plant Surface Metabolites by Matrix-Assisted Laser Desorption–Ionization Mass Spectrometry Imaging: Glucosinolates on Arabidopsis thaliana Leaves. Plant J. 2015, 81, 961–972. [Google Scholar] [CrossRef]
- Burow, M.; Halkier, B.A. How Does a Plant Orchestrate Defense in Time and Space? Using Glucosinolates in Arabidopsis as Case Study. Curr. Opin. Plant Biol. 2017, 38, 142–147. [Google Scholar] [CrossRef]
- Cole, R.A. Isothiocyanates, Nitriles and Thiocyanates as Products of Autolysis of Glucosinolates in Cruciferae. Phytochemistry 1976, 15, 759–762. [Google Scholar] [CrossRef]
- Halkier, B.A.; Gershenzon, J. Biology and Biochemistry of Glucosinolates. Annu. Rev. Plant Biol. 2006, 57, 303–333. [Google Scholar] [CrossRef] [PubMed]
- Wittstock, U.; Kurzbach, E.; Herfurth, A.M.; Stauber, E.J. Chapter Six—Glucosinolate Breakdown. In Advances in Botanical Research; Kopriva, S., Ed.; Academic Press: Cambridge, MA, USA, 2016; Volume 80, pp. 125–169. [Google Scholar]
- Wittstock, U.; Burow, M. Glucosinolate Breakdown in Arabidopsis: Mechanism, Regulation and Biological Significance. Arab. Book 2010. [Google Scholar] [CrossRef]
- Jeschke, V.; Weber, K.; Moore, S.S.; Burow, M. Coordination of Glucosinolate Biosynthesis and Turnover Under Different Nutrient Conditions. Front. Plant Sci. 2019, 10, 1560. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, R.; Hirai, M.Y. Atypical Myrosinase as a Mediator of Glucosinolate Functions in Plants. Front. Plant Sci. 2019, 10, 1008. [Google Scholar] [CrossRef]
- Perez, V.C.; Dai, R.; Block, A.K.; Kim, J. Metabolite Analysis of Arabidopsis CYP79A2 Overexpression Lines Reveals Turnover of Benzyl Glucosinolate and an Additive Effect of Different Aldoximes on Phenylpropanoid Repression. Plant Signal. Behav. 2021, 16, 1966586. [Google Scholar] [CrossRef]
- Lambrix, V.; Reichelt, M.; Mitchell-Olds, T.; Kliebenstein, D.J.; Gershenzon, J. The Arabidopsis Epithiospecifier Protein Promotes the Hydrolysis of Glucosinolates to Nitriles and Influences Trichoplusia Ni Herbivory. Plant Cell 2001, 13, 2793–2807. [Google Scholar] [CrossRef]
- Vorwerk, S.; Biernacki, S.; Hillebrand, H.; Janzik, I.; Müller, A.; Weiler, E.W.; Piotrowski, M. Enzymatic Characterization of the Recombinant Arabidopsis thaliana Nitrilase Subfamily Encoded by the NIT2/NIT1/NIT3-Gene Cluster. Planta 2001, 212, 508–516. [Google Scholar] [CrossRef]
- Janowitz, T.; Trompetter, I.; Piotrowski, M. Evolution of Nitrilases in Glucosinolate-Containing Plants. Phytochemistry 2009, 70, 1680–1686. [Google Scholar] [CrossRef]
- Urbancsok, J.; Bones, A.M.; Kissen, R. Benzyl Cyanide Leads to Auxin-Like Effects Through the Action of Nitrilases in Arabidopsis thaliana. Front. Plant Sci. 2018, 9, 1240. [Google Scholar] [CrossRef]
- Vik, D.; Mitarai, N.; Wulff, N.; Halkier, B.A.; Burow, M. Dynamic Modeling of Indole Glucosinolate Hydrolysis and Its Impact on Auxin Signaling. Front. Plant Sci. 2018, 9, 550. [Google Scholar] [CrossRef]
- Julkowska, M.M.; Koevoets, I.T.; Mol, S.; Hoefsloot, H.; Feron, R.; Tester, M.A.; Keurentjes, J.J.B.; Korte, A.; Haring, M.A.; de Boer, G.J.; et al. Genetic Components of Root Architecture Remodeling in Response to Salt Stress. Plant Cell 2017, 29, 3198–3213. [Google Scholar] [CrossRef] [PubMed]
- Clavijo Mccormick, A.; Irmisch, S.; Reinecke, A.; Boeckler, G.A.; Veit, D.; Reichelt, M.; Hansson, B.S.; Gershenzon, J.; Köllner, T.G.; Unsicker, S.B. Herbivore-Induced Volatile Emission in Black Poplar: Regulation and Role in Attracting Herbivore Enemies. Plant Cell Environ. 2014, 37, 1909–1923. [Google Scholar] [CrossRef]
- Quittenden, L.J.; Davies, N.W.; Smith, J.A.; Molesworth, P.P.; Tivendale, N.D.; Ross, J.J. Auxin Biosynthesis in Pea: Characterization of the Tryptamine Pathway. Plant Physiol. 2009, 151, 1130–1138. [Google Scholar] [CrossRef] [PubMed]
- Hirata, H.; Ohnishi, T.; Tomida, K.; Ishida, H.; Kanda, M.; Sakai, M.; Yoshimura, J.; Suzuki, H.; Ishikawa, T.; Dohra, H.; et al. Seasonal Induction of Alternative Principal Pathway for Rose Flower Scent. Sci. Rep. 2016, 6, 20234. [Google Scholar] [CrossRef]
- Kaminaga, Y.; Schnepp, J.; Peel, G.; Kish, C.M.; Ben-Nissan, G.; Weiss, D.; Orlova, I.; Lavie, O.; Rhodes, D.; Wood, K.; et al. Plant Phenylacetaldehyde Synthase Is a Bifunctional Homotetrameric Enzyme That Catalyzes Phenylalanine Decarboxylation and Oxidation. J. Biol. Chem. 2006, 281, 23357–23366. [Google Scholar] [CrossRef] [PubMed]
- Gutensohn, M.; Klempien, A.; Kaminaga, Y.; Nagegowda, D.A.; Negre-Zakharov, F.; Huh, J.H.; Luo, H.; Weizbauer, R.; Mengiste, T.; Tholl, D.; et al. Role of Aromatic Aldehyde Synthase in Wounding/Herbivory Response and Flower Scent Production in Different Arabidopsis Ecotypes. Plant J. 2011, 66, 591–602. [Google Scholar] [CrossRef]
- Hirata, H.; Ohnishi, T.; Ishida, H.; Tomida, K.; Sakai, M.; Hara, M.; Watanabe, N. Functional Characterization of Aromatic Amino Acid Aminotransferase Involved in 2-Phenylethanol Biosynthesis in Isolated Rose Petal Protoplasts. J. Plant Physiol. 2012, 169, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Koeduka, T.; Fujita, Y.; Furuta, T.; Suzuki, H.; Tsuge, T.; Matsui, K. Aromatic Amino Acid Decarboxylase Is Involved in Volatile Phenylacetaldehyde Production in Loquat (Eriobotrya japonica) Flowers. Plant Biotechnol. 2017, 34, 193–198. [Google Scholar] [CrossRef]
- Koshiba, T.; Saito, E.; Ono, N.; Yamamoto, N.; Sato, M. Purification and Properties of Flavin- and Molybdenum-Containing Aldehyde Oxidase from Coleoptiles of Maize. Plant Physiol. 1996, 110, 781–789. [Google Scholar] [CrossRef]
- Iwase, A.; Takebayashi, A.; Aoi, Y.; Favero, D.S.; Watanabe, S.; Seo, M.; Kasahara, H.; Sugimoto, K. 4-Phenylbutyric Acid Promotes Plant Regeneration as an Auxin by Being Converted to Phenylacetic Acid via an IBR3-Independent Pathway. Plant Biotechnol. 2022, 39, 51–58. [Google Scholar] [CrossRef]
- Hayashi, K.; Arai, K.; Aoi, Y.; Tanaka, Y.; Hira, H.; Guo, R.; Hu, Y.; Ge, C.; Zhao, Y.; Kasahara, H.; et al. The Main Oxidative Inactivation Pathway of the Plant Hormone Auxin. Nat. Commun. 2021, 12, 6752. [Google Scholar] [CrossRef]
- Müller, K.; Dobrev, P.I.; Pěnčík, A.; Hošek, P.; Vondráková, Z.; Filepová, R.; Malínská, K.; Brunoni, F.; Helusová, L.; Moravec, T.; et al. DIOXYGENASE FOR AUXIN OXIDATION 1 Catalyzes the Oxidation of IAA Amino Acid Conjugates. Plant Physiol. 2021, 187, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Grubb, C.D.; Zipp, B.J.; Ludwig-Müller, J.; Masuno, M.N.; Molinski, T.F.; Abel, S. Arabidopsis Glucosyltransferase UGT74B1 Functions in Glucosinolate Biosynthesis and Auxin Homeostasis. Plant J. 2004, 40, 893–908. [Google Scholar] [CrossRef]
- Staswick, P.E.; Serban, B.; Rowe, M.; Tiryaki, I.; Maldonado, M.T.; Maldonado, M.C.; Suza, W. Characterization of an Arabidopsis Enzyme Family That Conjugates Amino Acids to Indole-3-Acetic Acid. Plant Cell 2005, 17, 616–627. [Google Scholar] [CrossRef]
- Westfall, C.S.; Sherp, A.M.; Zubieta, C.; Alvarez, S.; Schraft, E.; Marcellin, R.; Ramirez, L.; Jez, J.M. Arabidopsis thaliana GH3.5 Acyl Acid Amido Synthetase Mediates Metabolic Crosstalk in Auxin and Salicylic Acid Homeostasis. Proc. Natl. Acad. Sci. USA 2016, 113, 13917–13922. [Google Scholar] [CrossRef] [PubMed]
- Mashiguchi, K.; Hisano, H.; Takeda-Kamiya, N.; Takebayashi, Y.; Ariizumi, T.; Gao, Y.; Ezura, H.; Sato, K.; Zhao, Y.; Hayashi, K.; et al. Agrobacterium Tumefaciens Enhances Biosynthesis of Two Distinct Auxins in the Formation of Crown Galls. Plant Cell Physiol. 2019, 60, 29–37. [Google Scholar] [CrossRef]
- Lynch, J.H.; Qian, Y.; Guo, L.; Maoz, I.; Huang, X.-Q.; Garcia, A.S.; Louie, G.; Bowman, M.E.; Noel, J.P.; Morgan, J.A.; et al. Modulation of Auxin Formation by the Cytosolic Phenylalanine Biosynthetic Pathway. Nat. Chem. Biol. 2020, 16, 850–856. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).