
Efficacy of Imidacloprid Seed Treatments against Four Wheat Aphids under Laboratory and Field Conditions


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
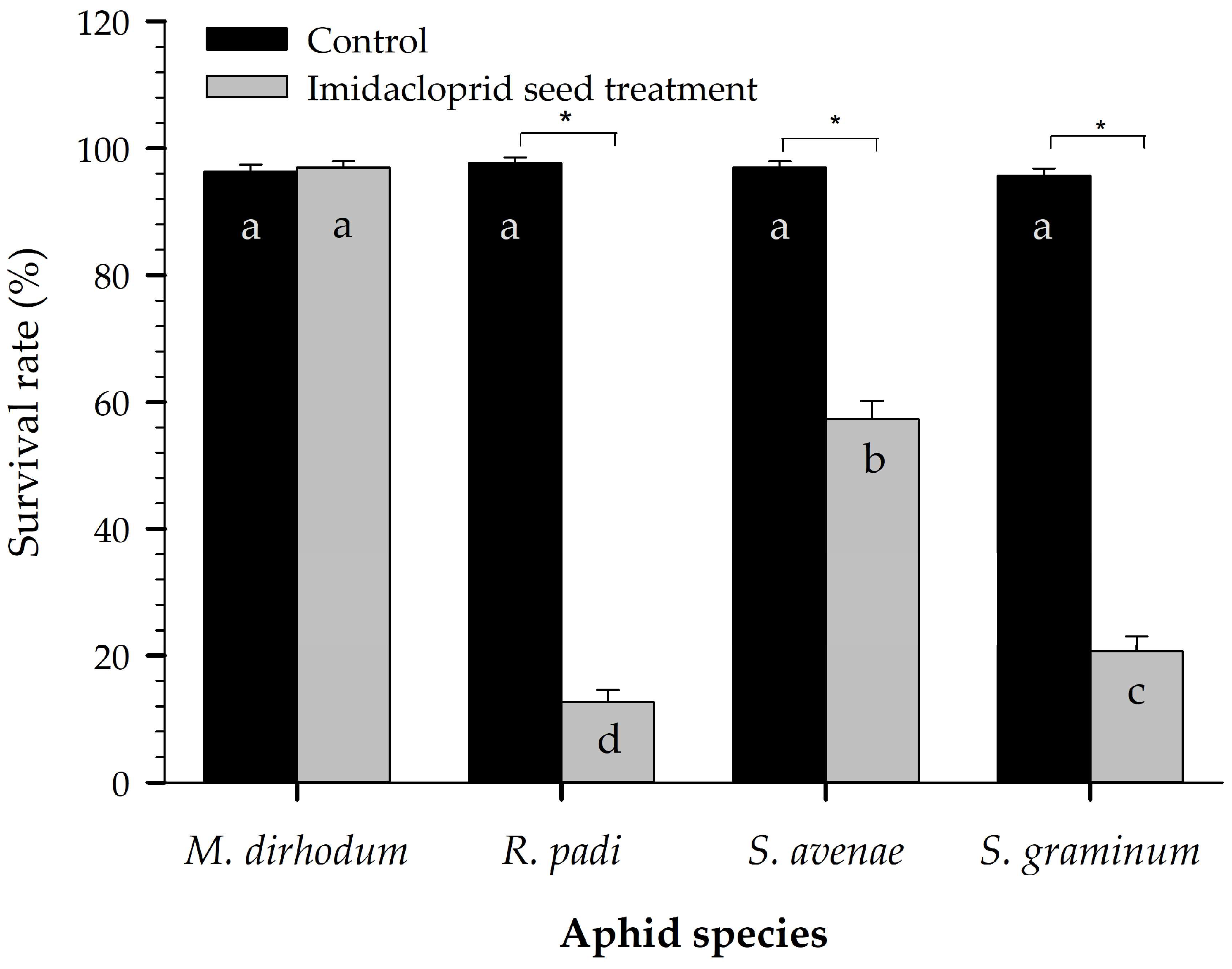
2.1. Laboratory Survival
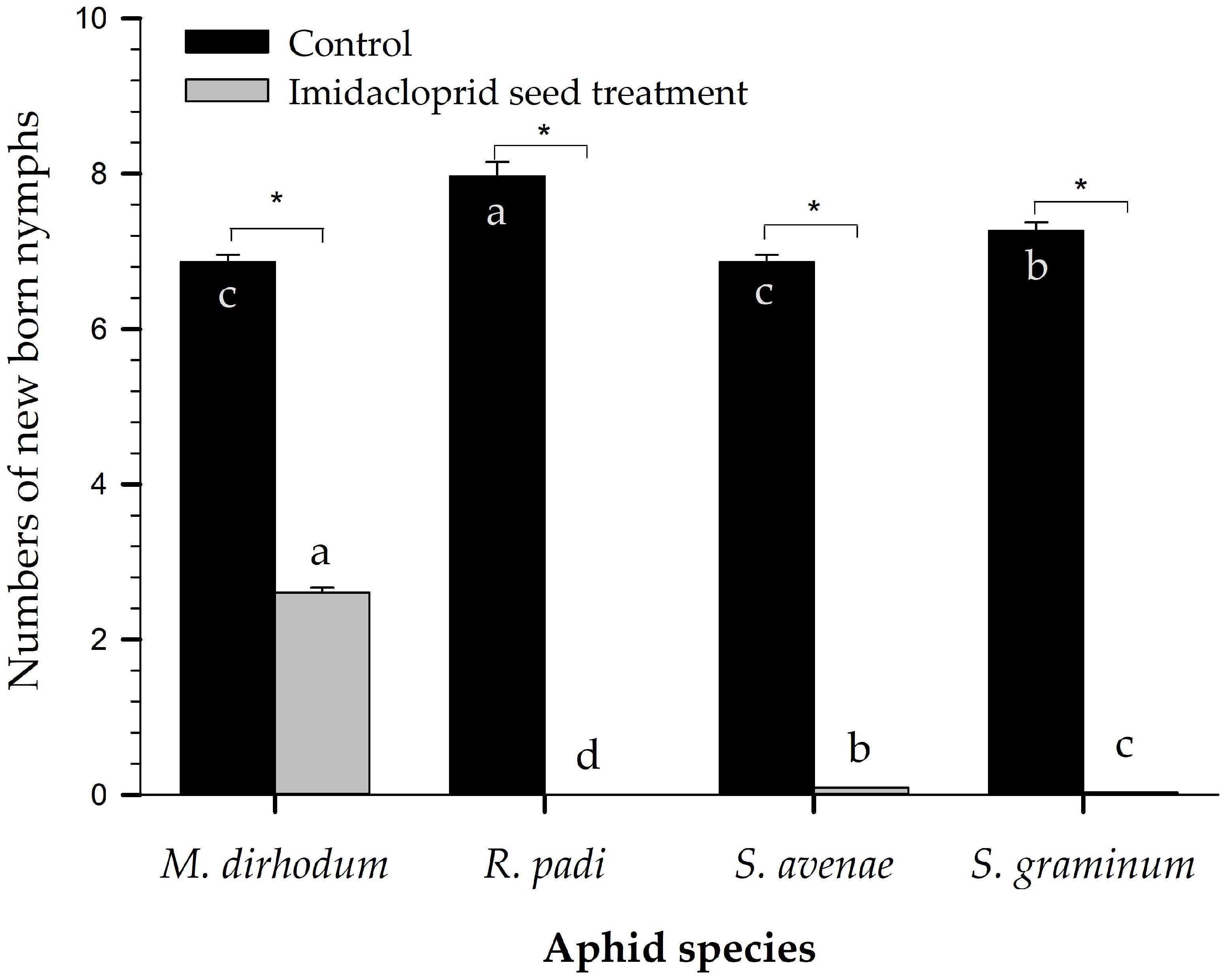
2.2. Laboratory Fecundity
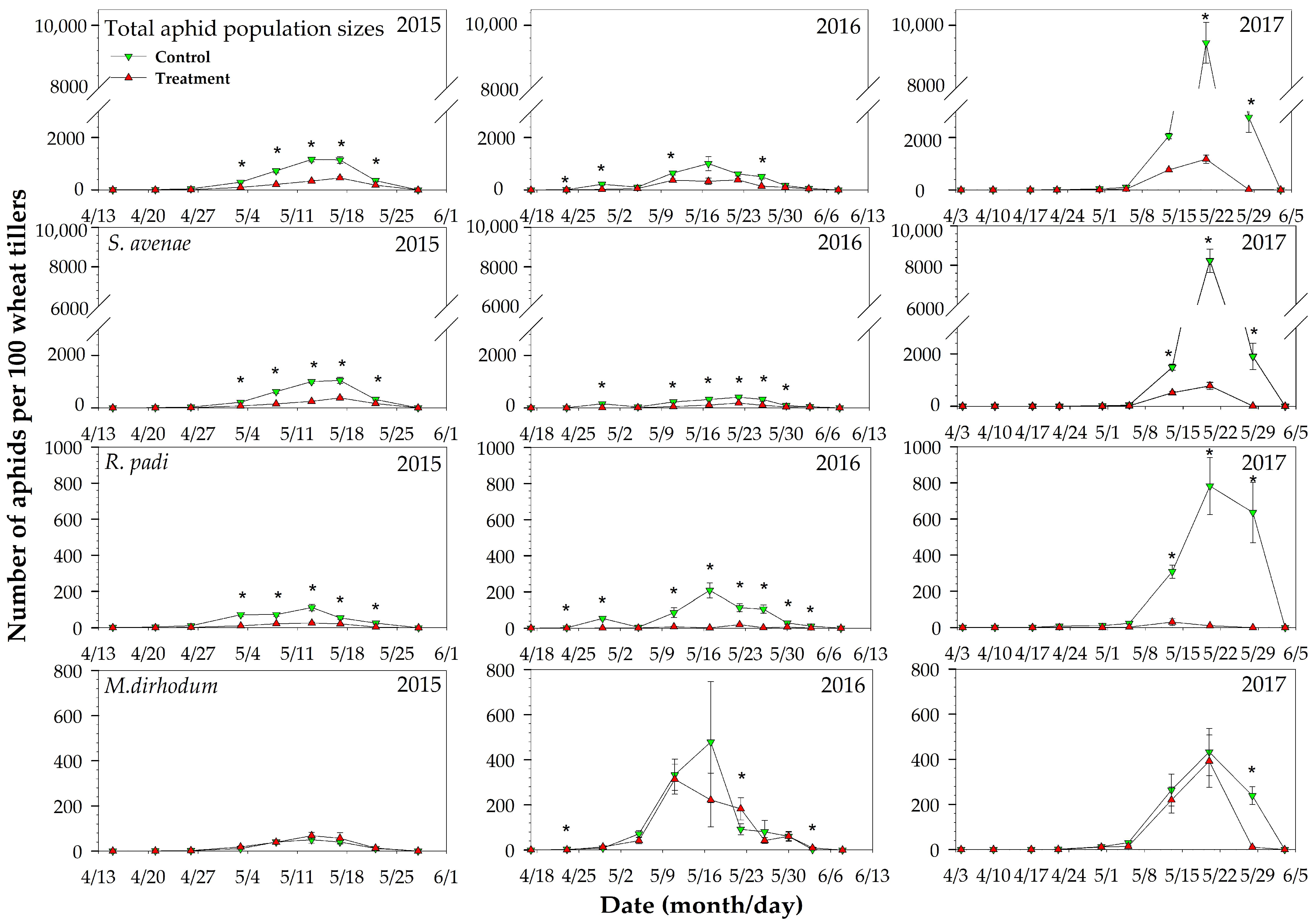
2.3. Field Population Dynamics
3. Discussion
4. Materials and Methods
4.1. Wheat Aphids
4.2. Plant Material
4.3. Seed Treatment
4.4. Laboratory Experiment
4.5. Field Experiment
4.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guo, Y.Y.; Cao, Y.Z.; Li, S.G.; Wu, Y.Q.; Liu, A.Z.; Hu, Y. Preliminary study on damage and dynamic control indexes of the mixedwheat aphids population in the ear stage. Plant Prot. 1988, 14, 2–5. [Google Scholar]
- Cao, Y.Z.; Yin, J.; Li, K.B.; Zhang, K.C.; Li, X.Q. Exploration of the factors causing the outbreak of wheat aphids and the control strategies. Plant Prot. 2006, 32, 72–75. [Google Scholar]
- Ma, C.S.; Ma, G.; Pincebourde, S. Survive a warming climate: Insect responses to extreme high temperatures. Ann. Rev. Entomol. 2020, 66, 163–184. [Google Scholar] [CrossRef]
- Wang, C.Z.; Huo, Z.G.; Guo, A.H.; Huang, H.; Zhang, L.; Lu, M.H.; He, Y.B.; Liu, W.C. Climatic risk assessment of wheat aphids in northern China. J. Appl. Meteorol. 2021, 32, 160–174. [Google Scholar]
- Sun, X.; Sun, Y.M.; Ma, L.; Liu, Z.; Wang, Q.Y.; Wang, D.L.; Zhang, C.J.; Yu, H.W.; Xu, M.; Ding, J.Q.; et al. Multidecadal, continent-level analysis indicates agricultural practices impact wheat aphid loads more than climate change. Commun. Biol. 2022, 5, 761. [Google Scholar] [CrossRef] [PubMed]
- Men, X.Y.; Dong, Z.K.; Li, L.L.; Yang, Q.F.; Zhang, Q.Q.; Ouyang, F.; Lu, Z.B.; Li, C.; Yu, Y.; Zhuang, Q.Y. Advances in the integrated management of wheat pests based on ecological regulation. Chin. J. Appl. Entomol. 2020, 57, 59–69. [Google Scholar]
- Huang, C.; Jiang, Y.Y.; Li, C.G. Occurrence, yield loss and dynamics of wheat diseases and insect pests in China from 1987 to 2018. Plant Prot. 2020, 46, 186–193. [Google Scholar]
- Zhang, Q.Q.; Men, X.Y.; Hui, C.; Ge, F.; Ouyang, F. Wheat yield losses from pests and pathogens in China. Agric. Ecosyst. Environ. 2022, 326, 107821. [Google Scholar] [CrossRef]
- Chen, J.L. Wheat aphid. In Institute of Plant Protection Chinese Academy of Agricultural Sciences and The China Society of Plant Protection (ed) Crop Disease and Insect Pests in China, 3rd ed.; Beijing Chinese Agricultural Press: Beijing, China, 2015; pp. 438–448. [Google Scholar]
- Albajes, R.; Lopez, C.; Pons, X. Predatory fauna in corn fields and response to imidacloprid seed treatment. J. Econ. Entomol. 2003, 96, 1805–01813. [Google Scholar] [CrossRef]
- Nault, B.A.; Taylor, A.G.; Urwiler, M.; Rabaey, T.; Hutchison, W.D. Neonicotinoid seed treatments for managing potato leafhopper infestations in snap bean. Crop Prot. 2004, 23, 147–154. [Google Scholar] [CrossRef]
- Kennedy, T.F.; Connery, J. Grain yield reductions in spring barley due to barley yellow dwarf virus and aphid feeding. Ir. J. Agric. Food Res. 2005, 44, 111–128. [Google Scholar]
- Magalhaes, L.C.; Hunt, T.E.; Siegfried, B.D. Efficacy of neonicotinoid seed treatments to reduce soybean aphid populations under field and controlled conditions in Nebraska. J. Econ. Entomol. 2009, 102, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Ni, H.X.; Li, G.B. Progress in research on integrated pest management of wheat in China. Chin. Agric. Sci. Bull. 1993, 9, 16–19. [Google Scholar]
- Dewar, A.M. Chemical control. In Aphids as Crop Pests; Emden, H.F.V., Harrington, R., Eds.; CABI: Wallingford, UK, 2007. [Google Scholar]
- Jansen, J.P. A three-year field study on the short-term effects of insecticides used to control cereal aphids on plant-dwelling aphid predators in winter wheat. Pest Manag. Sci. 2000, 56, 533–539. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, X.F.; Zhao, Y.H.; Wei, Y.; Mu, W.; Liu, F. Effects of imidacloprid and clothianidin seed treatments on wheat aphids and their natural enemies on winter wheat. Pest Manag. Sci. 2016, 72, 1141–1149. [Google Scholar] [CrossRef]
- Pike, K.S.; Reed, G.L.; Graf, G.T.; Allison, D. Compatibility of imidacloprid with fungicides as a seed-treatment control of Russian wheat aphid (Homoptera: Aphididae) and effect on germination, growth, and yield of wheat and barley. J. Econ. Entomol. 1993, 86, 586–593. [Google Scholar] [CrossRef]
- Burd, J.D.; Elliott, N.C.; Reed, D.K. Effects of the aphicides ‘gaucho’ and CGA-215944 on feeding behavior and tritrophic interactions of Russian wheat aphids. Southwest. Entomol. 1996, 21, 145–152. [Google Scholar]
- Elbert, A.; Haas, M.; Springer, B.; Thielert, W.; Nauen, R. Applied aspects of neonicotinoid uses in crop protection. Pest Manag. Sci. 2008, 64, 1099–1105. [Google Scholar] [CrossRef]
- Liu, A.Z.; Li, S.J.; Han, S. Preliminary study on controlling effect and yield increasing action of seed dressing with imidacloprid on wheat aphids. J. Henan Agric. Sci. 2005, 34, 63–64. [Google Scholar]
- Jeschke, P.; Nauen, R.; Schindler, M.; Elbert, A. Overview of the status and global strategy for neonicotinoids. J. Agric. Food Chem. 2011, 59, 2897–2908. [Google Scholar] [CrossRef]
- Shi, X.B.; Jiang, L.L.; Wang, H.Y.; Qiao, K.; Wang, D.; Wang, K.Y. Toxicities and sublethal effects of seven neonicotinoid insecticides on survival, growth and reproduction of imidacloprid-resistant cotton aphid, Aphis gossypii. Pest Manag. Sci. 2011, 67, 1528–1533. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Du, Z.B.; Wu, Y.Q.; Gong, Z.J.; Jiang, Y.L.; Duan, Y.; Li, T.; Lei, C.L. Sub-lethal effects of four neonicotinoid seed treatments on the demography and feeding behavior of the wheat aphid Sitobion avenae. Pest Manag. Sci. 2014, 70, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Krupke, C.H.; Long, E.Y. Intersections between neonicotinoid seed treatments and honey bees. Curr. Opin. Insect Sci. 2015, 10, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Pons, X.; Albajes, R. Control of maize pests with imidacloprid seed dressing treatment in Catalonia (NEIberian Peninsula) under traditional crop conditions. Crop Prot. 2002, 21, 943–950. [Google Scholar] [CrossRef]
- Bereś, P.K.; Górski, D.; Kucharczyk, H. Influence of seed treatments and foliar insecticides used against Oscinella frit L. in maize on the population of thrips. Acta Sci. Pol. Agric. 2017, 16, 3–16. [Google Scholar]
- Ahmed, N.E.; Kanan, H.O.; Inanaga, S.; Ma, Y.Q.; Sugimoto, Y. Impact of pesticide seed treatments on aphid control and yield of wheat in the Sudan. Crop Prot. 2001, 20, 929–934. [Google Scholar] [CrossRef]
- Drinkwater, T.W. Comparison of imidacloprid with carbamate insecticides, and the role of planting depth in the control of false wireworms, Somaticus species, in maize. Crop Prot. 1994, 13, 341–345. [Google Scholar] [CrossRef]
- Drinkwater, T.W.; Groenewald, L.H. Comparison of imidacloprid and furathiocarb seed dressing insecticides for the control of the black maize beetle, Heteronychus arator Fabricius (Coleoptera: Scarabaeidae), in maize. Crop Prot. 1994, 13, 421–424. [Google Scholar] [CrossRef]
- Gray, S.M.; Bergstrom, G.C.; Vaughan, R.; Smith, D.M.; Kalb, D.W. Insecticidal control of cereal aphids and its impact on the epidemiology of the barley yellow dwarf luteoviruses. Crop Prot. 1996, 15, 687–697. [Google Scholar] [CrossRef]
- Sloderbeck, P.E.; Witt, M.D.; Buschman, L.L. Effects of imidacloprid seed treatment on greenbug (Homopera: Aphididae) infestations on three sorghum hybrids. Southwest. Entomol. 1996, 21, 181–187. [Google Scholar]
- Kirkland, L.S.; Pirtle, E.L.; Umina, P.A. Responses of the Russian wheat aphid (Diuraphis noxia) and bird cherry oat aphid (Rhopalosiphum padi) to insecticide seed treatments in wheat. Crop Pasture Sci. 2018, 69, 966–973. [Google Scholar] [CrossRef]
- Li, Y.P.; Li, X.R.; Zhang, Y.H.; Zhu, X.; Cheng, D.F. Impacts of seed coating with imidacloprid on laboratory populations of Metopolophium dirhodum (Walker) (Hemiptera: Aphididae). Plant Prot. 2019, 45, 25–29. [Google Scholar]
- Gong, P.P.; Chen, D.F.; Wang, C.; Li, M.Y.; Li, X.A.; Zhang, Y.H.; Li, X.R.; Zhu, X. Susceptibility of four species of aphids in wheat to seven insecticides and its relationship to detoxifying enzymes. Front. Physiol. 2021, 11, 623612. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.R.; Fu, X.W.; Luo, M.H.; Yuan, G.H.; Zhang, Y. Studies on the niche of two species of wheat aphids and their natural enemies in the middle and late growing stage of wheat. J. Henan Agric. Univ. 2008, 42, 430–433. [Google Scholar]
- Blackman, R.L.; Eastop, V.F. Aphids on the World’s Crops: An Identification Guide; Wiley: Chichester, UK, 1984. [Google Scholar]
- Gong, P.P.; Li, X.A.; Wang, C.; Zhu, S.; Li, Q.C.; Zhang, Y.H.; Li, X.R.; Li, G.K.; Liu, E.L.; Gao, H.F.; et al. The Sensitivity of Field Populations of Metopolophium dirhodum (Walker) (Hemiptera: Aphididae) to Seven Insecticides in Northern China. Agronomy 2021, 11, 1556. [Google Scholar] [CrossRef]
- Weber, G. On the Ecological Genetics of Metopolophiurn dirhodurn (Walker) (Hemiptera, Aphididae). J. Appl. Entomol. 1985, 100, 451–458. [Google Scholar]
- Stamm, M.D.; Heng-Moss, T.M.; Baxendale, F.P.; Siegfried, B.D.; Blankenship, E.E.; Nauen, R. Uptake and translocation of imidacloprid, clothianidin and flupyradifurone in seed-treated soybeans. Pest Manag. Sci. 2016, 72, 1099–1109. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Ann. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Rundlöf, M.; Andersson, G.K.S.; Bommarco, R.; Fries, I.; Hederström, V.; Herbertsson, L.; Jonsson, O.; Klatt, B.K.; Pedersen, T.R.; Yourstone, J.; et al. Seed coating with a neonicotinoid insecticide negatively affects wild bees. Nature 2015, 521, 77–80. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Zhang, F.M.; Cheng, M.D.F.; Yin, M.X.T.; Li, M.X.R.; Fan, M.J. A Brief Bioassay Method of Wheat Aphids. Patent ZL201410117638.3 [P/OL], 16 July 2014. [Google Scholar]
- Zhou, H.B.; Chen, J.L.; Cheng, D.F.; Liu, Y.; Sun, J.R. Effects of wheat-pea intercropping on the population dynamics of Sitobion avenea (Homoptera: Aphididae) and its main natural enemies. Acta Entomol. Sin. 2009, 52, 775–782. [Google Scholar]
- Wei, M.F.; Chi, H.; Guo, Y.F.; Li, X.W.; Zhao, L.L.; Ma, R.Y. Demography of Cacopsylla chinensis (Hemiptera: Psyllidae) reared on four cultivars of Pyrus bretschneideri (Rosales: Rosaceae) and P. communis pears with estimations of confidence intervals of specific life table statistics. J. Econ. Entomol. 2020, 113, 2343–2353. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.Y.; Chen, Z.Z.; Zheng, F.Q.; Shi, A.J.; Guo, T.T.; Yeh, B.H.; Chi, H.; Xu, Y.Y. Demographic analysis, a comparison of the jackknife and bootstrap methods, and predation projection: A case study of Chrysopa pallens (Neuroptera: Chrysopidae). J. Econ. Entomol. 2013, 106, 1–9. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Li, Y.; Li, X.; Zhu, X.; Zhang, Y. Efficacy of Imidacloprid Seed Treatments against Four Wheat Aphids under Laboratory and Field Conditions. Plants 2023, 12, 238. https://doi.org/10.3390/plants12020238
Zhang Z, Li Y, Li X, Zhu X, Zhang Y. Efficacy of Imidacloprid Seed Treatments against Four Wheat Aphids under Laboratory and Field Conditions. Plants. 2023; 12(2):238. https://doi.org/10.3390/plants12020238
Chicago/Turabian StyleZhang, Zhi, Yaping Li, Xiangrui Li, Xun Zhu, and Yunhui Zhang. 2023. "Efficacy of Imidacloprid Seed Treatments against Four Wheat Aphids under Laboratory and Field Conditions" Plants 12, no. 2: 238. https://doi.org/10.3390/plants12020238
APA StyleZhang, Z., Li, Y., Li, X., Zhu, X., & Zhang, Y. (2023). Efficacy of Imidacloprid Seed Treatments against Four Wheat Aphids under Laboratory and Field Conditions. Plants, 12(2), 238. https://doi.org/10.3390/plants12020238