
Comparative Physiological and Transcriptome Analyses Reveal Mechanisms of Salicylic-Acid-Reduced Postharvest Ripening in ‘Hosui’ Pears (Pyrus pyrifolia Nakai)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
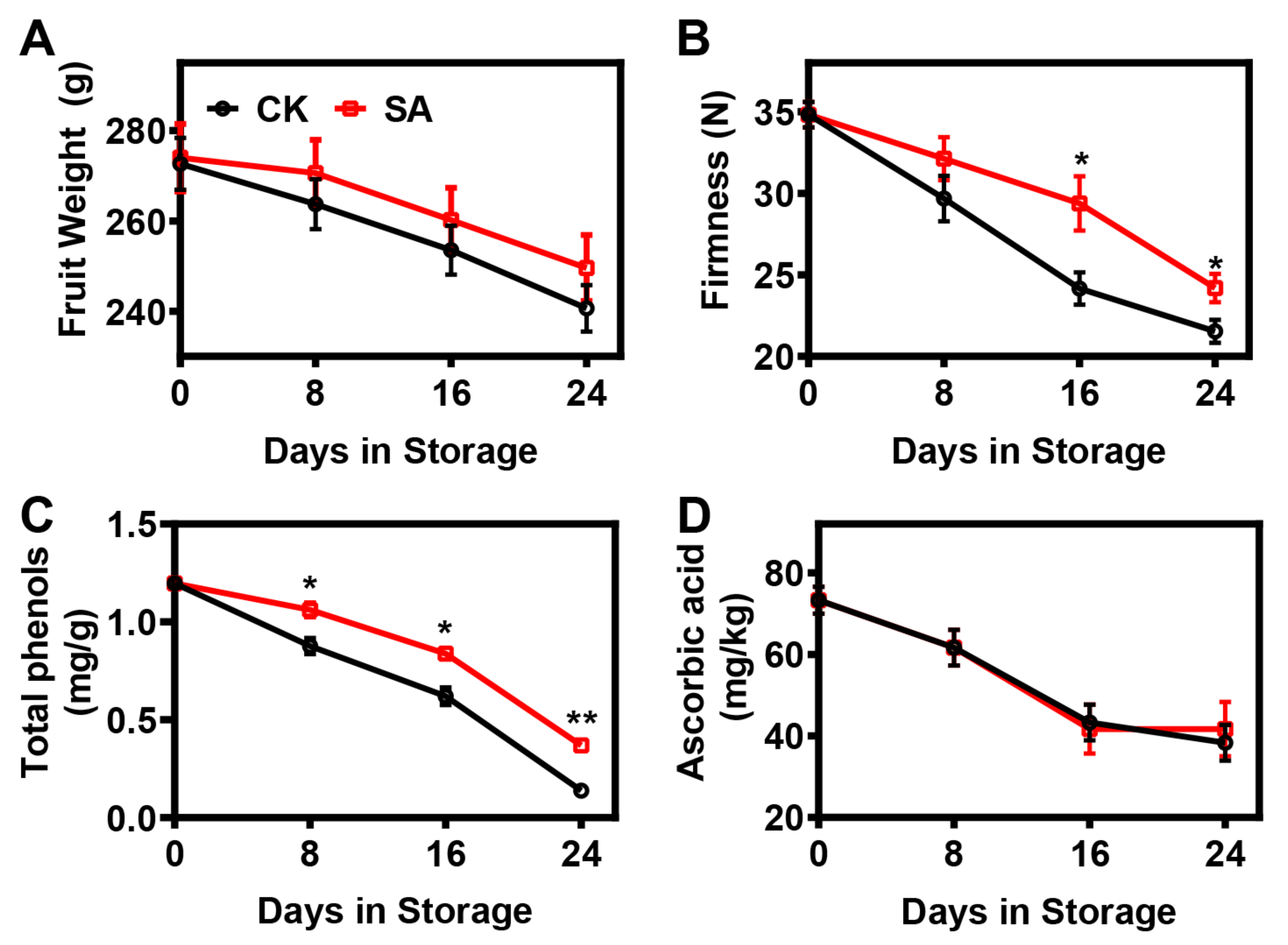
2.1. Effect of SA Treatment on the Postharvest Ripening Process of Pear Fruit
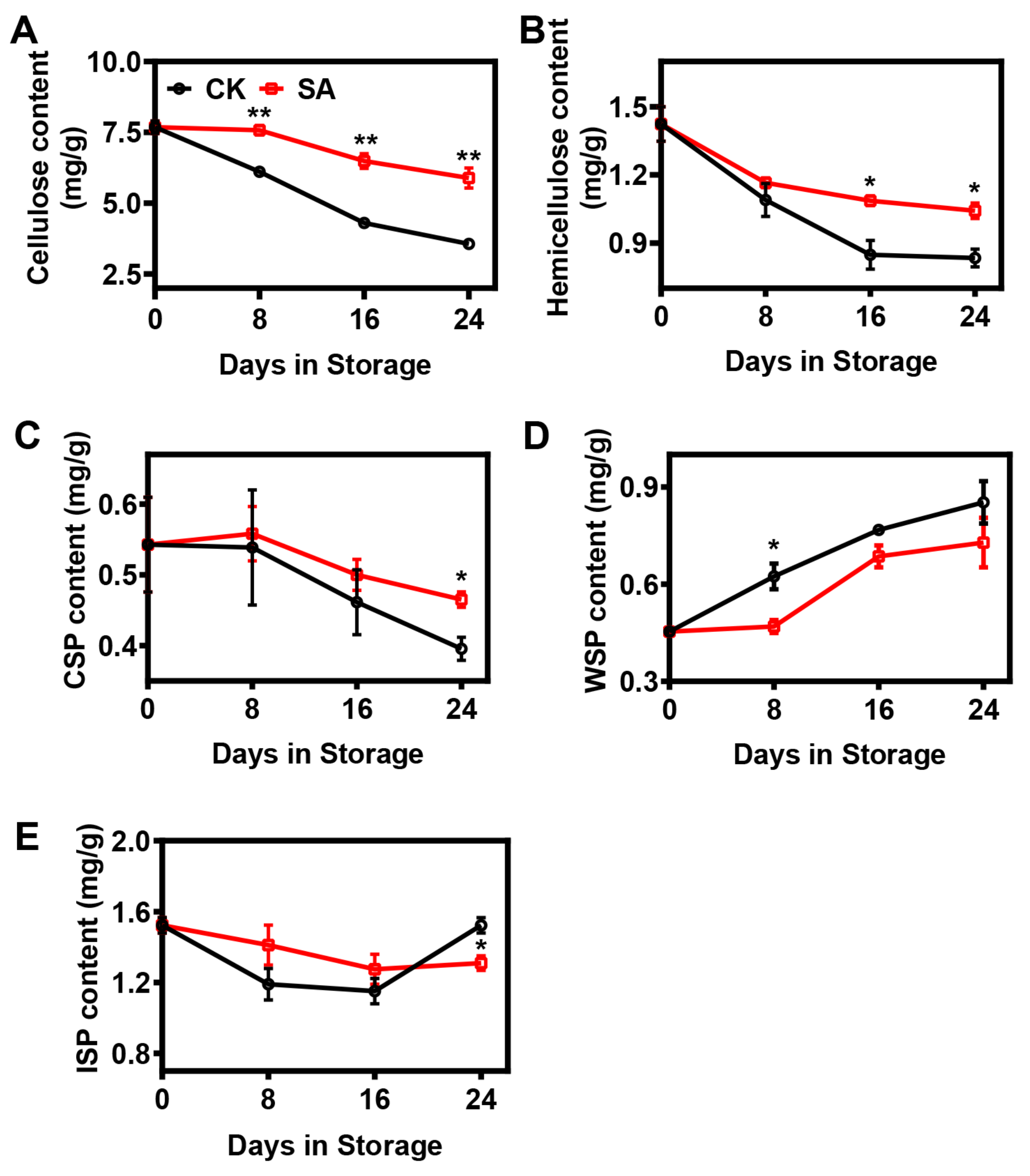
2.2. Effect of SA Treatment on the Cell Wall Composition of Pear Fruit during Room-Temperature Storage
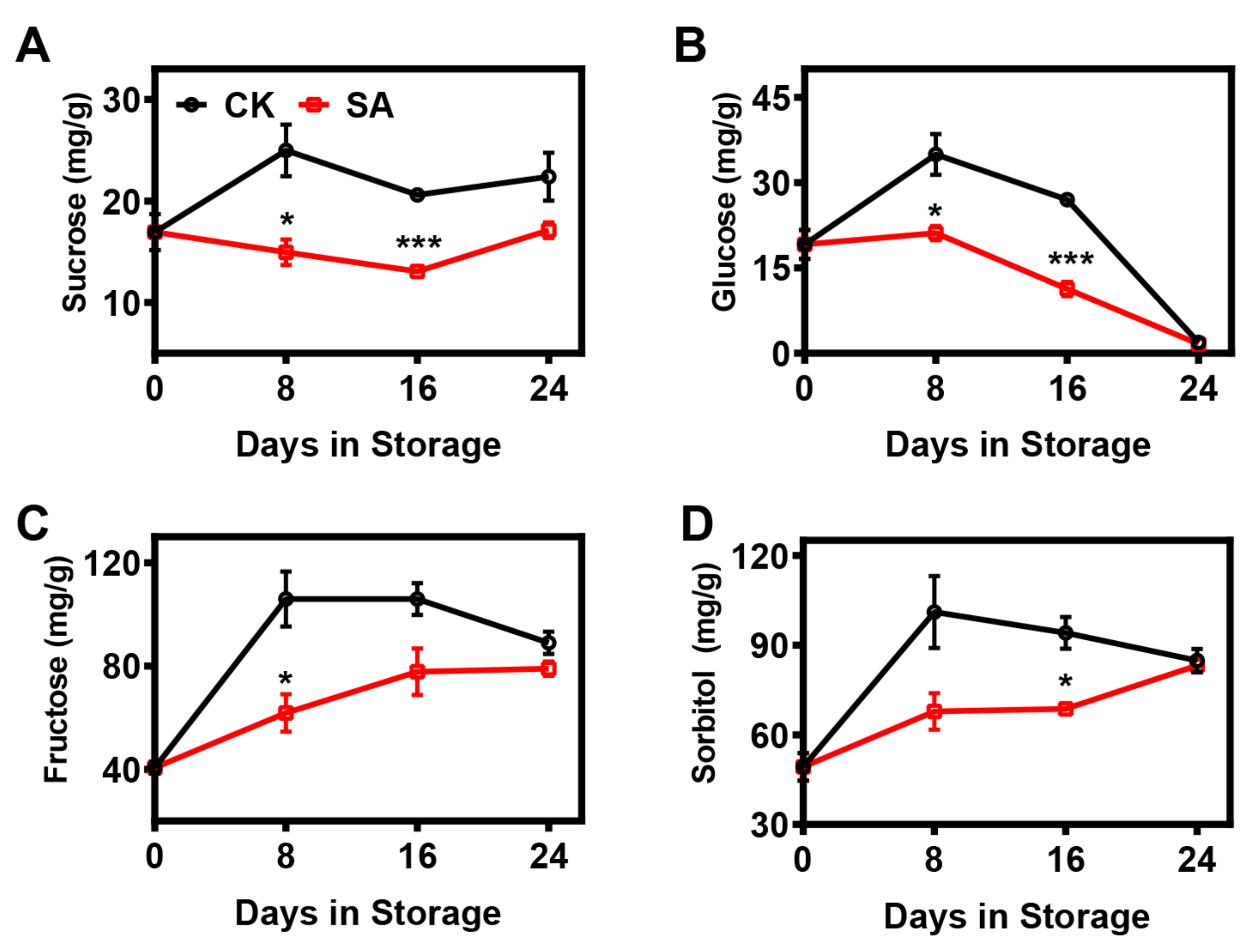
2.3. Effect of SA Treatment on the Soluble Sugar Composition of Pear Fruit during Room-Temperature Storage
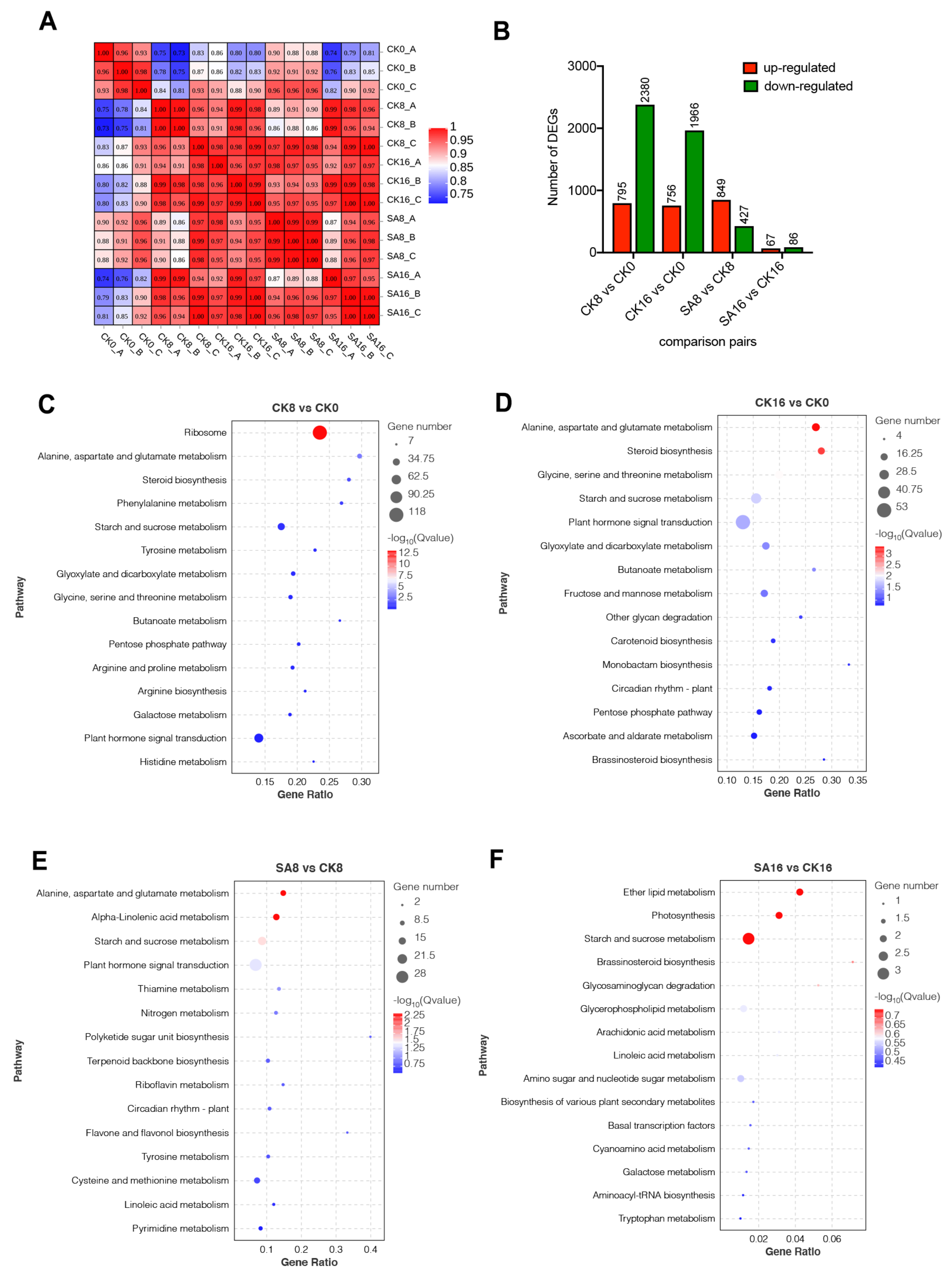
2.4. Characterization of Transcriptional Changes of Pear Fruit in Response to Salicylic Acid by RNA-Seq
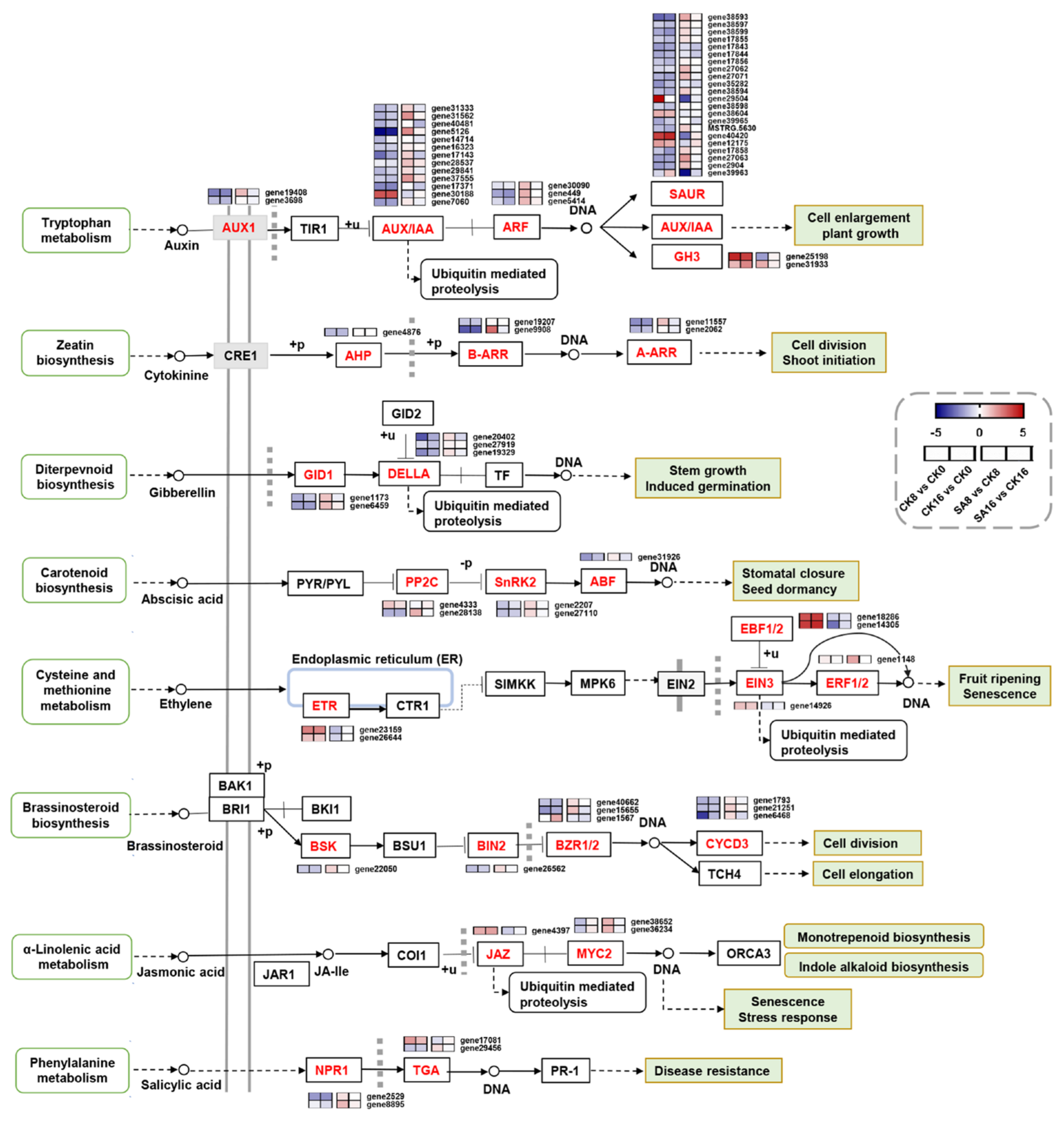
2.5. DEGs Related to Plant Hormone Signal Transduction
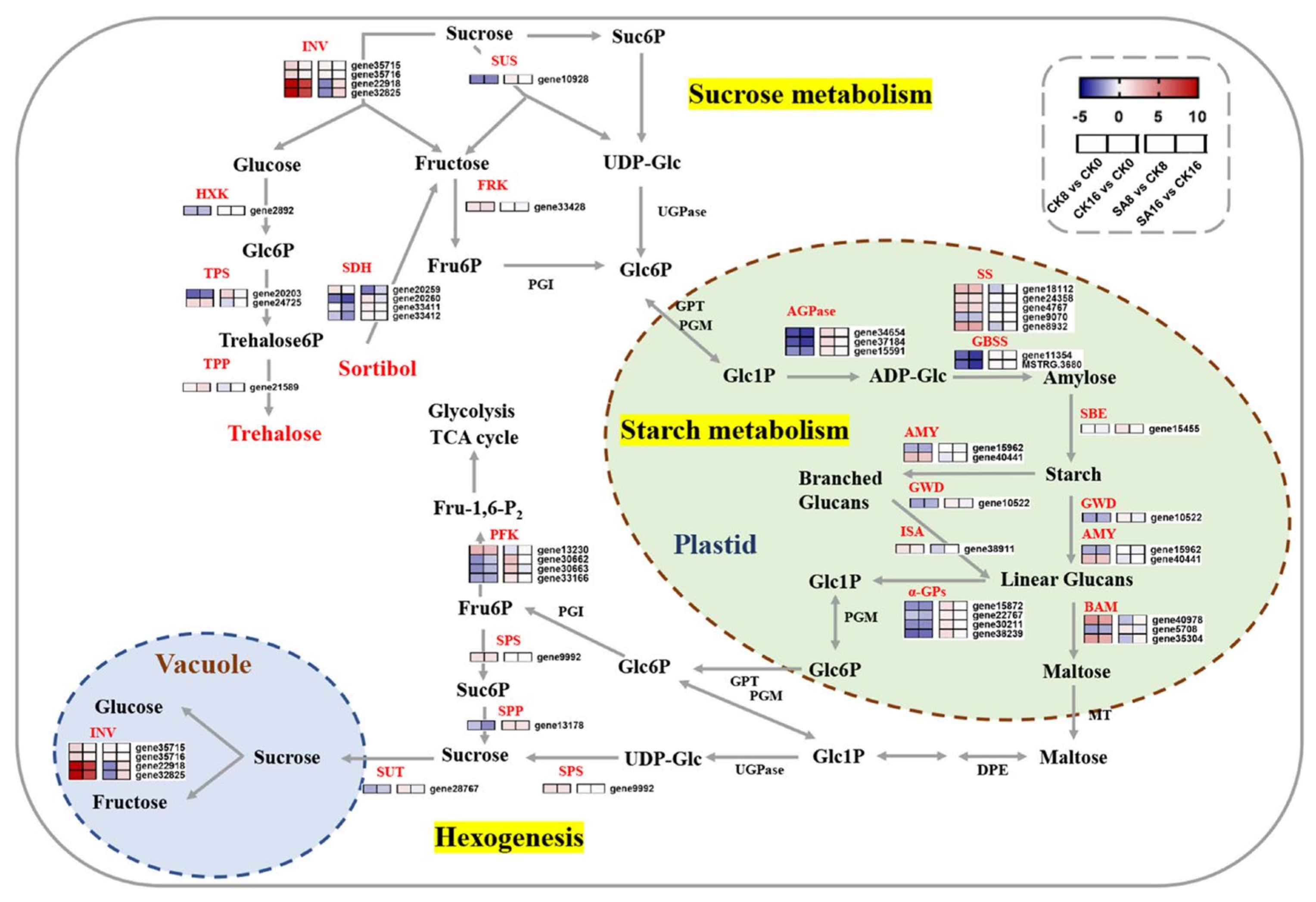
2.6. DEGs Related to Sugar Metabolism
2.7. DEGs Related to Fruit Softening
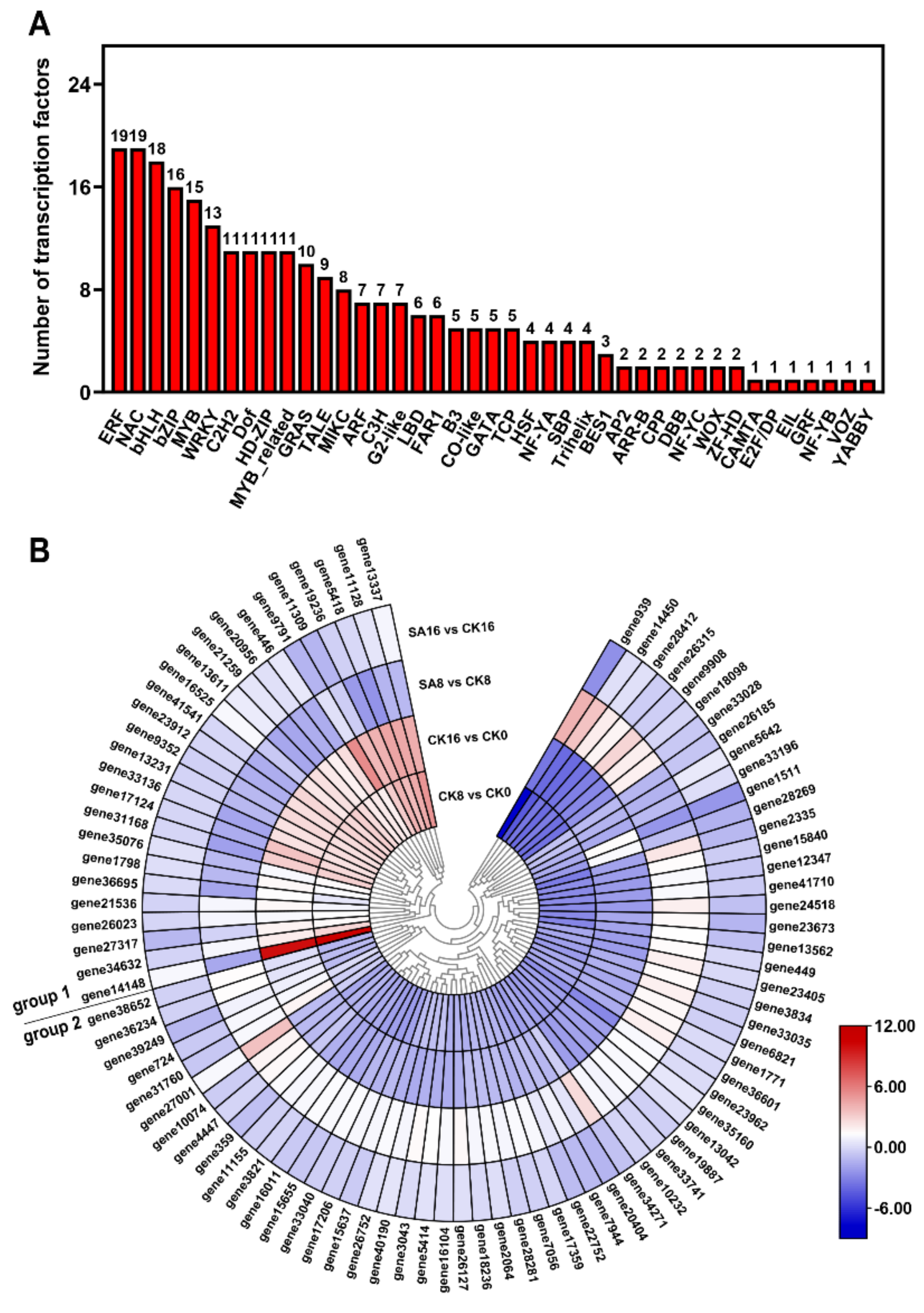
2.8. Differentially Expressed TFs
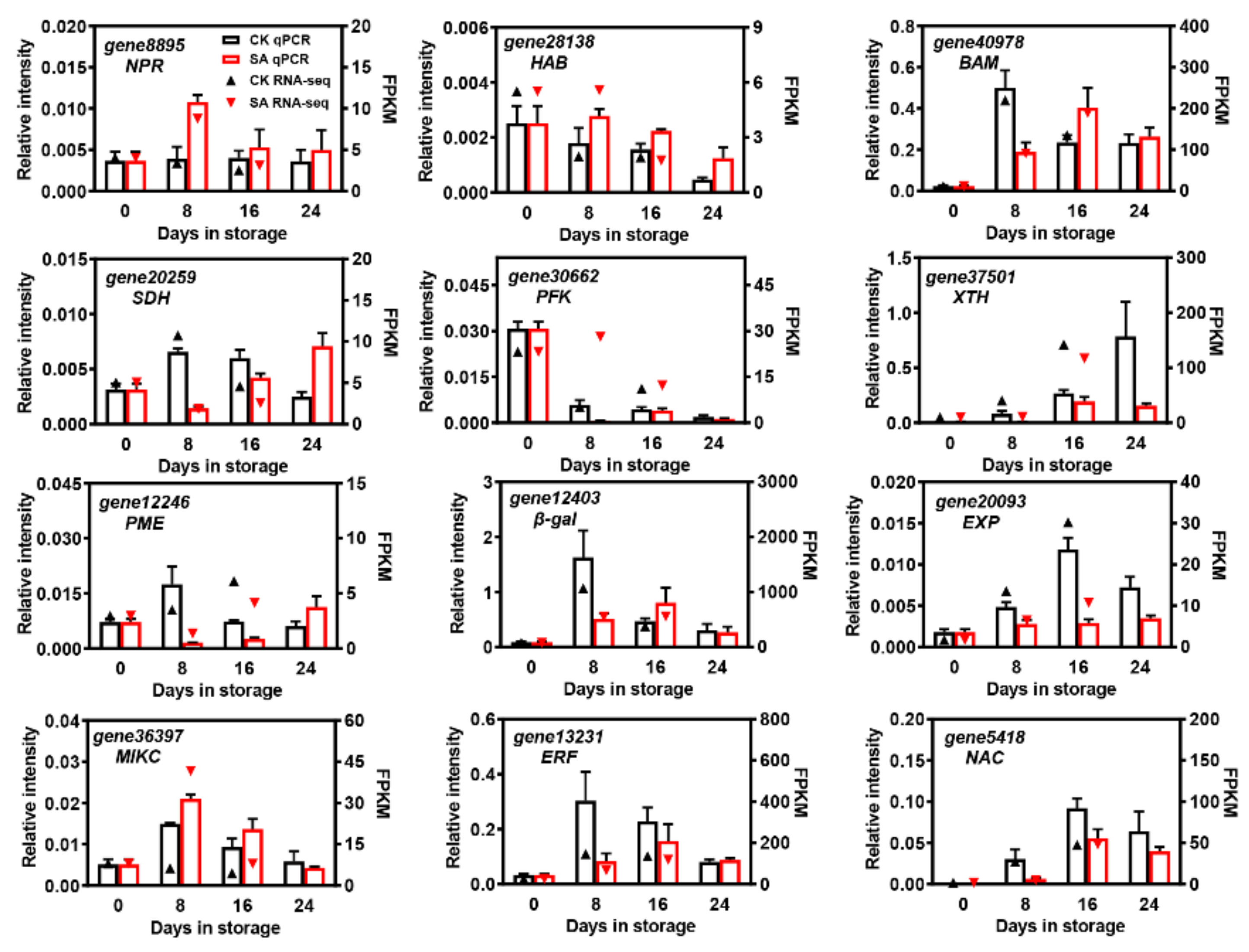
2.9. Validation of RNA-Seq by qRT-PCR Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material and Treatments
4.2. Weight and Firmness
4.3. Determination of Total Phenols and Ascorbic Acid
4.4. Determination of Cell Wall Components
4.5. Determination of Sugar Components
4.6. RNA Extraction and RNA-Seq
4.7. cDNA Synthesis and qRT-PCR Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, J.M.; Zhang, M.Y.; Li, X.L.; Khan, A.; Kumar, S.; Allan, A.C.; Lin-Wang, K.; Espley, R.V.; Wang, C.H.; Wang, R.Z.; et al. Pear genetics: Recent advances, new prospects, and a roadmap for the future. Hortic. Res. 2022, 9, uhab040. [Google Scholar] [CrossRef]
- Wu, J.; Wang, Y.T.; Xu, J.B.; Korbanet, S.S.; Feial, Z.J.; Tao, S.T.; Ming, R.; Tai, S.S.; Khan, A.M.; Postman, J.D.; et al. Diversification and independent domestication of Asian and European pears. Genome Biol. 2018, 19, 77. [Google Scholar] [CrossRef] [PubMed]
- Itai, A.; Kawata, T.; Tanabe, K.; Tamura, F.; Uchiyama, M.; Tomomitsu, M.; Shiraiwa, N. Identification of 1-aminocyclopropane-1-carboxylic acid synthase genes controlling the ethylene level of ripening fruit in Japanese pear (Pyrus pyrifolia Nakai). Mol. Genom. Genet. 1999, 262, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Dong, Y.; Yan, H.; Ge, W.; Shen, C.; Guan, J.; Liu, L.; Zhang, Y. Effects of 1-MCP on chlorophyll degradation pathway-associated genes expression and chloroplast ultrastructure during the peel yellowing of Chinese pear fruits in storage. Food Chem. 2012, 135, 415–422. [Google Scholar] [CrossRef]
- Xu, H.Y.; Chen, Y.Y.; Wang, L.B.; Xie, Z.H.; Gu, C.; Zhang, S.L. Transcriptome analysis reveals a regulation of ethylene-induced post-harvest senescence in pear fruit. Sci. Hortic. 2018, 240, 585–591. [Google Scholar] [CrossRef]
- Itai, A.; Kotaki, T.; Tanabe, K.; Tamura, F.; Kawaguchi, D.; Fukuda, M. Rapid identification of 1-aminocyclopropane-1-carboxylate (ACC) synthase genotypes in cultivars of Japanese pear (Pyrus pyrifolia Nakai) using CAPS markers. Theor. Appl. Genet. 2003, 106, 1266–1272. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Lai, T.; Xu, Y.; Tian, S. Changes in physiology and quality of Laiyang pear in long time storage. Sci. Hortic. 2013, 150, 31–36. [Google Scholar] [CrossRef]
- Zhang, S.L.; Ma, M.; Zhang, H.P.; Zhang, S.L.; Qian, M.; Zhang, Z.; Luo, W.Q.; Fan, J.B.; Liu, L.Q.; Wang, L.B. Genome-wide analysis of polygalacturonase gene family from pear genome and identification of the member involved in pear softening. BMC Plant Biol. 2019, 19, 587. [Google Scholar] [CrossRef] [PubMed]
- Nath, A.; Bidyut, C.D.; Singh, A.; Patel, R.K.; Paul, D.; Misra, L.K.; Ojha, H. Extension of shelf life of pear fruits using different packaging materials. J. Food Sci. Technol. 2012, 49, 556–563. [Google Scholar] [CrossRef]
- Singh, V.; Jawandha, S.K.; Gill, P.P.S.; Gill, M.S. Suppression of fruit softening and extension of shelf life of pear by putrescine application. Sci. Hortic. 2019, 256, 108623. [Google Scholar] [CrossRef]
- Golias, J.; Letal, J.; Balik, J.; Koziskova, J. Effect of controlled atmosphere storage on production of volatiles and ethylene from cv. Zaosuli pears. Hortic. Sci. 2016, 43, 117–125. [Google Scholar] [CrossRef]
- Wang, L.B.; Ma, M.; Zhang, Y.R.; Wu, Z.F.; Guo, L.; Luo, W.Q.; Wang, L.; Zhang, Z.; Zhang, S.L. Characterization of the genes involved in malic acid metabolism from pear fruit and their expression profile after postharvest 1-MCP/Ethrel treatment. J. Agric. Food Chem. 2018, 66, 8772–8782. [Google Scholar] [CrossRef] [PubMed]
- Adhikary, T.; Gill, P.P.S.; Jawandha, S.K.; Kaur, N. Postharvest quality response of pears with beeswax coatings during long term cold storage. J. Hortic. Sci. Biotechnol. 2022, 97, 785–798. [Google Scholar] [CrossRef]
- Li, J.Z.; Dai, X.N.; Li, Q.Y.; Jiang, F.D.; Xu, X.F.; Guo, T.T.; Zhang, H.X. Low temperatures inhibit the pectin degradation of ‘Docteur Jules Guyot’ pear (Pyrus communis L.). Int. J. Biol. Macromol. 2023, 242, 124719. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, N.; Khan, N.A.; Ferrante, A.; Trivellini, A.; Francini, A.; Khan, M.I.R. Ethylene role in plant growth, development and senescence: Interaction with other phytohormones. Front. Plant Sci. 2017, 8, 475. [Google Scholar] [CrossRef] [PubMed]
- Kou, X.H.; Feng, Y.; Yuan, S.; Zhao, X.Y.; Wu, C.E.; Wang, C.; Xue, Z.H. Different regulatory mechanisms of plant hormones in the ripening of climacteric and non-climacteric fruits: A review. Plant Mol. Biol. 2021, 107, 477–497. [Google Scholar] [CrossRef]
- Li, X.M.; Wang, X.M.; Zhang, Y.; Zhang, A.H.; You, C.X. Regulation of fleshy fruit ripening: From transcription factors to epigenetic modifications. Hortic. Res. 2022, 9, uhac013. [Google Scholar] [CrossRef]
- Song, B.B.; Li, X.L.; Cao, B.B.; Zhang, M.Y.; Korban, S.S.; Yu, L.A.; Yang, W.X.; Zhao, K.J.; Li, J.M.; Wu, J. An identical-by-descent segment harbors a 12-bp insertion determining fruit softening during domestication and speciation in Pyrus. BMC Biol. 2022, 20, 215. [Google Scholar] [CrossRef]
- Tucker, G.; Yin, X.R.; Zhang, A.D.; Wang, M.M.; Zhu, Q.G.; Liu, X.F.; Xie, X.L.; Chen, K.S.; Grierson, D. Ethylene and fruit softening. Food Qual. Saf. 2017, 1, 253–267. [Google Scholar] [CrossRef]
- Zhang, A.D.; Wang, W.Q.; Tong, Y.; Li, M.J.; Grierson, D.; Ferguson, I.; Chen, K.S.; Yin, X.R. Transcriptome analysis identifies a zinc finger protein regulating starch degradation in kiwifruit. Plant Physiol. 2018, 178, 850–863. [Google Scholar] [CrossRef]
- Hiwasa, K.; Nakano, R.; Hashimoto, A.; Matsuzaki, M.; Murayama, H.; Inaba, A.; Kubo, Y. European, Chinese and Japanese pear fruits exhibit differential softening characteristics during ripening. J. Exp. Bot. 2004, 55, 2281–2290. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Song, L.; Wang, Z.; Wang, Z.; Meng, G.; Zhai, R.; Cai, M.; Ma, F.; Xu, L. Screening of cell wall-related genes that are expressed differentially during ripening of pears with different softening characteristics. Postharvest Biol. Technol. 2016, 115, 1–8. [Google Scholar] [CrossRef]
- Itai, A.; Tanahashi, T. Inhibition of sucrose loss during cold storage in Japanese pear (Pyrus pyrifolia Nakai) by 1-MCP. Postharvest Biol. Technol. 2008, 48, 355–363. [Google Scholar] [CrossRef]
- Ma, M.; Wang, L.B.; Zhang, S.L.; Guo, L.; Zhang, Z.; Li, J.L.; Sun, Q.; Zhang, S.L. Acid vacuolar invertase 1 (PbrAc-Inv1) and invertase inhibitor 5 (PbrII5) were involved in sucrose hydrolysis during postharvest pear storage. Food Chem. 2020, 320, 126635. [Google Scholar] [CrossRef]
- Wang, J.W.; Dong, S.Z.; Jiang, Y.G.; He, H.S.; Liu, T.; Lv, M.; Ji, S.J. Influence of long-term cold storage on phenylpropanoid and soluble sugar metabolisms accompanied with peel browning of ‘Nanguo’ pears during subsequent shelf life. Sci. Hortic. 2020, 260, 108888. [Google Scholar] [CrossRef]
- Rivas-San Vicente, M.; Plasencia, J. Salicylic acid beyond defence: Its role in plant growth and development. J. Exp. Bot. 2011, 62, 3321–3338. [Google Scholar] [CrossRef]
- Asghari, M.; Aghdam, M.S. Impact of salicylic acid on postharvest physiology of horticultural crops. Trends Food Sci. Technol. 2010, 21, 502–509. [Google Scholar] [CrossRef]
- Zhou, Y.; Ma, J.; Xie, J.; Deng, L.; Yao, S.; Zeng, K. Transcriptomic and biochemical analysis of highlighted induction of phenylpropanoid pathway metabolism of citrus fruit in response to salicylic acid, Pichia membranaefaciens and oligochitosan. Postharvest Biol. Technol. 2018, 142, 81–92. [Google Scholar] [CrossRef]
- Chen, C.J.; Sun, C.C.; Wang, Y.H.; Gong, H.S.; Zhang, A.D.; Yang, Y.Q.; Guo, F.J.; Cui, K.B.; Fan, X.G.; Li, X.L. The preharvest and postharvest application of salicylic acid and its derivatives on storage of fruit and vegetables: A review. Sci. Hortic. 2023, 312, 111858. [Google Scholar] [CrossRef]
- Trejo, E.J.O.; Brizzolara, S.; Cardillo, V.; Ruperti, B.; Bonghi, C.; Tonutti, P. The impact of PGRs applied in the field on the postharvest behavior of fruit crops. Sci. Hortic. 2023, 318, 112103. [Google Scholar] [CrossRef]
- Kumar, N.; Tokas, J.; Raghavendra, M.; Singal, H.R. Impact of exogenous salicylic acid treatment on the cell wall metabolism and ripening process in postharvest tomato fruit stored at ambient temperature. Int. J. Food Sci. Technol. 2021, 56, 2961–2972. [Google Scholar] [CrossRef]
- Wang, J.; Liu, X.; Zhang, H.; Allan, A.C.; Wang, W.; Yin, X. Transcriptional and post-transcriptional regulation of ethylene biosynthesis by exogenous acetylsalicylic acid in kiwifruit. Hortic. Res. 2022, 9, uhac116. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Duan, W.; Xie, K.; Ren, C.; Zhu, C.; Chen, K.; Zhang, B. Effect of salicylic acid treatment on sensory quality, flavor-related chemicals and gene expression in peach fruit after cold storage. Postharvest Biol. Technol. 2020, 161, 111089. [Google Scholar] [CrossRef]
- Hassan, I.; Zhang, Y.; Du, G.; Wang, G.; Zhang, J. Effect of salicylic acid (SA) on delaying fruit senescence of Huang Kum pear. Front. Agric. China 2007, 1, 456–459. [Google Scholar]
- Adhikary, T.; Gill, P.S.; Jawandha, S.K.; Bhardwaj, R.D.; Anurag, R.K. Browning and quality management of pear fruit by salicylic acid treatment during low temperature storage. J. Sci. Food Agric. 2020, 101, 853–862. [Google Scholar] [CrossRef]
- Sinha, A.; Gill, P.P.S.; Jawandha, S.K.; Grewal, S.K. Composite coating of chitosan with salicylic acid retards pear fruit softening under cold and supermarket storage. Food Res. Int. 2022, 160, 11724. [Google Scholar] [CrossRef]
- Luo, M.L.; Ge, W.Y.; Sun, H.J.; Yang, Q.X.; Sun, Y.Y.; Zhou, X.; Zhou, Q.; Ji, S.J. Salicylic acid treatment alleviates diminished ester production in cold-stored Nanguo’ pear by promoting the transcription of PuAAT. Postharvest Biol. Technol. 2022, 187, 111849. [Google Scholar] [CrossRef]
- Liu, D. Effect of 1-MCP on postharvest physiology and quality of ‘Hosui’ pear produced from three gorges reservoir areas. Storage Process 2016, 16, 27–30. (In Chinese) [Google Scholar]
- Nie, Z.; Wan, C.; Chen, C.; Chen, J. Comprehensive evaluation of the postharvest antioxidant capacity of majiayou pomelo harvested at different maturities based on PCA. Antioxidants 2019, 8, 136. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, L.; Fan, C.; Wang, P.; Cui, M.; Liu, L.; Yang, H.; Wang, J. Impact of methyl salicylate on storage quality, ethylene action, and protein profiling of ‘Zaosu’ pear (Pyrus bretschneideri). Sci. Hortic. 2020, 264, 109196. [Google Scholar] [CrossRef]
- Mahajan, B.V.C.; Singh, K.; Dhillon, W.S. Effect of 1-methylcyclopropene (1-MCP) on storage life and quality of pear fruits. J. Food Sci. Technol. 2010, 47, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Lwin, H.P.; Rudell, R.D.; Lee, J. Metabolism and cold chain performance of ‘Chuhwangbae’ Asian pears as impacted by 1-MCP treatment. Sci. Hortic. 2021, 288, 110357. [Google Scholar] [CrossRef]
- Wang, Y.; Sugar, D. Ripening behavior and quality of modified atmosphere packed ‘Doyenne du Comice’ pears during cold storage and simulated transit. Postharvest Biol. Technol. 2013, 81, 51–59. [Google Scholar] [CrossRef]
- Hira, N.; Mitalo, O.W.; Okada, R.; Sangawa, M.; Masuda, K.; Fujita, N.; Ushijima, K.; Akagi, T.; Kubo, Y. The effect of layer-by-layer edible coating on the shelf life and transcriptome of ‘Kosui’ Japanese pear fruit. Postharvest Biol. Technol. 2022, 185, 111787. [Google Scholar] [CrossRef]
- Jiang, B.; Liu, R.L.; Fang, X.J.; Tong, C.; Chen, H.J.; Gao, H.Y. Effects of salicylic acid treatment on fruit quality and wax composition of blueberry (Vaccinium virgatum Ait). Food Chem. 2023, 368, 130757. [Google Scholar] [CrossRef]
- Chen, C.Y.; Huang, Q.; Peng, X.; Wan, C.P.; Zeng, J.K.; Zhang, Y.J.; Chen, J.Y. Alleviatory effects of salicylic acid on postharvest softening and cell wall degradation of ‘Jinshayou’ pummelo (Citrus maxima Merr.): A comparative physiological and transcriptomic analysis. Food Chem. 2023, 424, 136428. [Google Scholar] [CrossRef] [PubMed]
- Koyuncu, M.A.; Erbas, D.; Onursal, C.E.; Secmen, T.; Guneyli, A.; Uzumcu, S.S. Postharvest treatments of salicylic acid, oxalic acid and putrescine influences bioactive compounds and quality of pomegranate during controlled atmosphere storage. J. Food Sci. Technol. 2019, 56, 350–359. [Google Scholar] [CrossRef]
- Zhao, Y.Y.; Song, C.C.; Brimmel, D.A.; Qi, S.N.; Lin, Q.; Bi, J.F.; Duan, Y.Q. Salicylic acid treatment mitigates chilling injury in peach fruit by regulation of sucrose metabolism and soluble sugar content. Food Chem. 2021, 358, 129867. [Google Scholar] [CrossRef]
- Davarynejad, G.H.; Zarei, M.; Nasrabadi, M.E.; Ardakani, E. Effects of salicylic acid and putrescine on storability, quality attributes and antioxidant activity of plum cv. ‘Santa Rosa’. J. Food Sci. Technol. 2015, 52, 2053–2062. [Google Scholar] [CrossRef]
- Li, S.; Chen, K.; Grierson, D. Molecular and hormonal mechanisms regulating fleshy fruit ripening. Cells 2021, 10, 1136. [Google Scholar] [CrossRef]
- Hao, Y.; Hu, G.; Breitel, D.; Liu, M.; Mila, I.; Frasse, P.; Fu, Y.; Aharoni, A.; Bouzayen, M.; Zouine, M. Auxin Response Factor SlARF2 Is an Essential Component of the Regulatory Mechanism Controlling Fruit Ripening in Tomato. PLoS Genet. 2015, 11, e1005649. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.H.; Mila, I.; Liu, M.; Rodrigues, M.A.; Vernoux, T.; Pirrello, J.; Bouzayen, M. The RIN-regulated Small Auxin-Up RNA SAUR 69 is involved in the unripe-to-ripe phase transition of tomato fruit via enhancement of the sensitivity to ethylene. New Phytol. 2018, 222, 820–836. [Google Scholar] [CrossRef]
- Liu, K.; Yuan, C.; Feng, S.; Zhong, S.; Li, H.; Zhong, J.; Shen, C.; Liu, J. Genome-wide analysis and characterization of Aux/IAA family genes related to fruit ripening in papaya (Carica papaya L.). BMC Genom. 2017, 18, 351. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Tseng, Y.; Pham, K.; Liu, M.; Beckles, D.M. Starch and sugars as determinants of postharvest shelf life and quality: Some new and surprising roles. Curr. Opin. Biotechnol. 2022, 78, 102844. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, H.; Ichimura, K.; Imada, S.; Yamaki, S. Sucrose synthase and sucrose phosphate synthase, but not acid invertase, are regulated by cold acclimation and deacclimation in cabbage seedlings. J. Plant Physiol. 2001, 158, 847–852. [Google Scholar] [CrossRef]
- Kanayama, Y. Sugar metabolism and fruit development in the tomato. Hortic. J 2017, 86, 417–425. [Google Scholar] [CrossRef]
- Payasi, A.; Mishra, N.N.; Chaves, A.L.; Singh, R. Biochemistry of fruit softening: An overview. Physiol. Mol. Biol. Plants 2009, 15, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Li, B.J.; Su, G.; Zhang, M.X.; Grierson, D.; Chen, K.S. Transcriptional regulation of fleshy fruit texture. J. Integr. Plant Biol. 2022, 64, 1649–1672. [Google Scholar] [CrossRef]
- Miedes, E.; Herbers, K.; Sonnewald, U.; Lorences, E.P. Overexpression of a cell wall enzyme reduces xyloglucan depolymerization and softening of transgenic tomato fruits. J. Agric. Food Chem. 2010, 58, 5708–5713. [Google Scholar] [CrossRef]
- Wang, D.D.; Samsulrizal, N.H.; Yan, C.; Allcock, N.S.; Craigon, J.; Blanco Ulate, B.; Ortega-Salazar, I.; Marcus, S.E.; Bagheri, H.M.; Perez Fons, L.; et al. Characterization of CRISPR mutants targeting genes modulating pectin degradation in ripening tomato. Plant Physiol. 2019, 179, 544–557. [Google Scholar]
- Paniagua, C.; Blanco Portales, R.; Barcelo-Muñoz, M.; Garcia-Gago, J.A.; Waldron, K.W.; Quesada, M.A.; Muñoz-Blanco, J.; Mercado, J.A. Antisense down regulation of the strawberry β-galactosidase gene FaβGal4 increases cell wall galactose levels and reduces fruit softening. J. Exp. Bot. 2016, 67, 619–631. [Google Scholar] [CrossRef] [PubMed]
- Brummell, D.A.; Harpster, M.H.; Civello, P.M.; Palys, J.M.; Bennett, A.B.; Dunsmuir, P. Modification of expansin protein abundance in tomato fruit alters softening and cell wall polymer metabolism during ripening. Plant Cell 1999, 11, 2203–2216. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wei, W.; Fan, Z.Q.; Zhao, X.D.; Zhang, Y.P.; Jing, Y.; Zhu, B.Z.; Zhu, H.L.; Shan, W.; Chen, J.Y.; et al. Re-evaluation of the nor mutation and the role of the NAC-NOR transcription factor in tomato fruit ripening. J. Exp. Bot. 2020, 71, 3560–3574. [Google Scholar] [CrossRef] [PubMed]
- Kuang, J.F.; Chen, J.Y.; Liu, X.C.; Han, Y.C.; Xiao, Y.Y.; Shan, W.; Tang, Y.; Wu, K.Q.; He, J.X.; Lu, W.J. The transcriptional regulatory network mediated by banana (Musa acuminata) dehydration-responsive element binding (MaDREB) transcription factors in fruit ripening. New Phytol. 2017, 214, 762–781. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.Y.; Kuang, J.F.; Qi, X.N.; Ye, Y.J.; Wu, Z.X.; Chen, J.Y.; Lu, W.J. A comprehensive investigation of starch degradation process and identification of a transcriptional activator MabHLH6 during banana fruit ripening. Plant Biotechnol. J. 2018, 16, 151–164. [Google Scholar] [CrossRef]
- Mao, W.W.; Han, Y.; Chen, Y.T.; Sun, M.Z.; Feng, Q.Q.; Li, L.; Liu, L.P.; Zhang, K.K.; Wei, L.Z.; Han, Z.H.; et al. Low temperature inhibits anthocyanin accumulation in strawberry fruit by activating FvMAPK3-induced phosphorylation of FvMYB10 and degradation of chalcone synthase 1. Plant Cell 2022, 34, 1226–1249. [Google Scholar] [CrossRef]
- Sang, Y.Y.; Liu, Y.X.; Tang, Y.S.; Yang, W.T.; Guo, M.R.; Chen, G.G. Transcriptome sequencing reveals mechanism of improved antioxidant capacity and maintained postharvest quality of winter jujube during cold storage after salicylic acid treatment. Postharvest Biol. Technol. 2022, 189, 111929. [Google Scholar] [CrossRef]
- Sekhon-Loodu, S.; Warnakulasuriya, S.N.; Rupasinghe, H.V.; Shahidi, F. Antioxidant ability of fractionated apple peel phenolics to inhibit fish oil oxidation. Food Chem. 2013, 140, 189–196. [Google Scholar] [CrossRef]
- Hughes, D.E. Titrimetric determination of ascorbic acid with 2,6-dichlorophenol indophenol in commercial liquid diets. J. Pharm. Sci. 1983, 72, 126–129. [Google Scholar] [CrossRef]
- Lin, Y.F.; Lin, Y.Z.; Lin, Y.X.; Lin, M.S.; Chen, Y.H.; Wang, H.; Lin, H.T. A novel chitosan alleviates pulp breakdown of harvested longan fruit by suppressing disassembly of cell wall polysaccharides. Carbohydr. Polym. 2019, 217, 126–134. [Google Scholar] [CrossRef]
- Yang, H.Y.; Li, J.Z.; Li, X.H.; Wu, R.; Zhang, X.L.; Fan, X.G.; Li, G.T.; Gong, H.S.; Yin, X.R.; Zhang, A.D. The mechanism of gibberellins treatment suppressing kiwifruit postharvest ripening processes by transcriptome analysis. Postharvest Biol. Technol. 2023, 198, 112223. [Google Scholar] [CrossRef]
- Kintner, P.K., III; Van Buren, J.P. Carbohydrate interference and its correction in pectin analysis using the m-hydroxydiphenyl method. J. Food Sci. 1982, 47, 756–759. [Google Scholar] [CrossRef]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Miao, M.M.; Xu, X.F.; Chen, X.H.; Xue, L.B.; Cao, B.S. Cucumber carbohydrate metabolism and translocation under chilling night temperature. J. Plant Physiol. 2007, 164, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Rep. 1993, 11, 113–116. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.F.; Zhang, P.F.; Bian, Y.H.; Liu, Z.Y.; Zhang, C.; Liu, X.; Wang, C.L. An integrated metabolic and transcriptomic analysis reveals the mechanism through which fruit bagging alleviates exocarp semi-russeting in pear fruit. Tree Physiol. 2021, 41, 1306–1318. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Wen, M.; Dai, R.; Liu, X.; Wang, C. Comparative Physiological and Transcriptome Analyses Reveal Mechanisms of Salicylic-Acid-Reduced Postharvest Ripening in ‘Hosui’ Pears (Pyrus pyrifolia Nakai). Plants 2023, 12, 3429. https://doi.org/10.3390/plants12193429
Zhang J, Wen M, Dai R, Liu X, Wang C. Comparative Physiological and Transcriptome Analyses Reveal Mechanisms of Salicylic-Acid-Reduced Postharvest Ripening in ‘Hosui’ Pears (Pyrus pyrifolia Nakai). Plants. 2023; 12(19):3429. https://doi.org/10.3390/plants12193429
Chicago/Turabian StyleZhang, Jing, Mengmeng Wen, Rong Dai, Xiao Liu, and Chunlei Wang. 2023. "Comparative Physiological and Transcriptome Analyses Reveal Mechanisms of Salicylic-Acid-Reduced Postharvest Ripening in ‘Hosui’ Pears (Pyrus pyrifolia Nakai)" Plants 12, no. 19: 3429. https://doi.org/10.3390/plants12193429
APA StyleZhang, J., Wen, M., Dai, R., Liu, X., & Wang, C. (2023). Comparative Physiological and Transcriptome Analyses Reveal Mechanisms of Salicylic-Acid-Reduced Postharvest Ripening in ‘Hosui’ Pears (Pyrus pyrifolia Nakai). Plants, 12(19), 3429. https://doi.org/10.3390/plants12193429