Abstract
Autophagy is a highly conserved self-degradation process that involves the degradation and recycling of cellular components and organelles. Although the involvement of autophagy in metabolic changes during fruit ripening has been preliminarily demonstrated, the variations in autophagic flux and specific functional roles in tomato fruit ripening remain to be elucidated. In this study, we analyzed the variations in autophagic flux during tomato fruit ripening. The results revealed differential expression of the SlATG8 family members during tomato fruit ripening. Transmission electron microscopy observations and dansylcadaverine (MDC) staining confirmed the presence of autophagy at the cellular level in tomato fruits. Furthermore, the overexpression of SlATG8f induced the formation of autophagosomes, increased autophagic flux within tomato fruits, and effectively enhanced the expression of ATG8 proteins during the color-transition phase of fruit ripening, thus promoting tomato fruit maturation. SlATG8f overexpression also led to the accumulation of vitamin C (VC) and soluble solids while reducing acidity in the fruit. Collectively, our findings highlight the pivotal role of SlATG8f in enhancing tomato fruit ripening, providing insights into the mechanistic involvement of autophagy in this process. This research contributes to a better understanding of the key factors that regulate tomato fruit quality and offers a theoretical basis for tomato variety improvement.
1. Introduction
Autophagy is a highly conserved intracellular self-degradation system in eukaryotic organisms that is utilized to eliminate and recycle cellular components, thereby breaking down toxic substances or damaged organelles and reclaiming essential nutrients [1,2]. Currently, three types of autophagy have been mainly identified in plants: microautophagy, macroautophagy (referred to as “autophagy” hereafter), and mega-autophagy [2]. Among them, autophagy is the most crucial and prevalent form in plants, involving the encapsulation of cellular materials within double-membrane structures known as autophagosomes. Subsequently, the outer membrane of the autophagosome fuses with a vacuolar membrane, leading to the transfer of cellular contents to the vacuole where they are degraded by vacuolar proteases and hydrolases [3].
The formation of autophagosomes is governed by a plethora of autophagy-related genes (ATGs). Initially discovered in yeast [4], homologous ATGs have been subsequently identified in animals and plants. The proteins encoded by ATGs participate in various stages of the autophagy process [5]. Among the numerous ATG proteins, the ubiquitin-like protein ATG8 plays a pivotal role in autophagy. It is involved in autophagosome formation, membrane elongation and fusion, degradation of cellular contents, and energy release [6]. ATG8 protein binds to phosphatidylethanolamine in a ubiquitin-like conjugation reaction, serving as a marker for growing autophagosomes and complete autophagic bodies. On the one hand, the loss of ATG8 function prevents autophagosome formation in yeast and other fungi [7]; however, this phenotype has not been observed in plants, which may be due to ATG8 gene redundancy. On the other hand, the overexpression of ATG8 promotes autophagosome formation in Arabidopsis and rice [8]. In addition to ATG8 lipidation, ATG8 protein is deaffixed by ATG4 and released from the autophagosome membrane, and the resulting released ATG8 protein is recycled to promote autophagosome formation in plants [9]. Consequently, ATG8 protein is often utilized as a reliable marker to assess autophagy induction and progression [10,11].
Research has indicated the significant role of autophagy in fruit ripening [9]. Transcriptome analysis of grape skin revealed elevated expression of ATG18g, ATG9, ATG11, and ATG2 during the late stages of grape maturation [12], implying the involvement of autophagy-related genes in the regulation of grape fruit aging. Subsequently, López-Vidal et al. investigated autophagy (ATG) components, ATG4 activity, autophagy receptor NBR1, protease Lon1, and protease Lon2 gene expression and protein content changes in two pepper varieties. Their findings confirmed the presence of autophagic structures in pepper fruit, highlighting their crucial role in fruit maturation and quality formation [13]. Similarly, Sánchez-Sevilla et al. identified autophagy-related structures at the cellular level in different stages of strawberry fruit ripening and demonstrated that blocking autophagy significantly impairs strawberry fruit growth and maturation [14]. These studies underscore the importance of autophagy in plant fruit development, yet the variations in autophagic flux during fruit ripening and its associated functional roles remain to be elucidated.
Tomato (Solanum lycopersicum L.), one of the most esteemed vegetables globally, is renowned for its diverse flavors, forms, colors, and abundant nutritional value [15]. During tomato fruit ripening, the color transitions from green to red, the flesh softens, and the content compounds increase [16]. This process involves cell division, cell expansion, and a series of metabolic reactions [17]. With continuous technological advancements, the enhancement of tomato fruit quality has become a significant focus. Considering the synthesis and cycling metabolism of nutrients during fruit development, acquiring more information about the role of autophagy in this process is imperative. In this study, through differential expression analysis of autophagy-related gene members of the ATG8 family during tomato fruit ripening, we demonstrate the existence of autophagic bodies within tomato fruit at the cellular level. Additionally, we observe the changes in ATG8 protein content necessary for autophagosome formation. Overexpression of SlATG8f leads to increased autophagic flux in tomato fruit, accelerating fruit ripening, upregulating ethylene-related genes, and enhancing fruit quality. Collectively, these findings suggest that autophagy is involved in regulating tomato fruit ripening and senescence, thereby enriching our understanding of the autophagic mechanisms in tomato fruit ripening and providing insights into the key factors that regulate fruit quality, ultimately contributing to the improvement of tomato varieties.
2. Results
2.1. Expression Analysis of SlATG8 Family Members during Tomato Fruit Ripening
In order to ascertain the involvement of autophagy in tomato fruit ripening, this study conducted an analysis of the expression levels of the core autophagy gene family, the ATG8 family members. Quantitative real-time polymerase chain reaction (qRT-PCR) was employed to quantify the transcript levels of SlATG8 family members during different stages of Micro-Tom tomato fruit maturation, including the green ripe, breaker, orange ripe, and red ripe stages (Figure 1). The results revealed distinct expression patterns among the SlATG8 family members. During the ripening process of tomato fruits, SlATG8a and SlATG8c exhibited lower expression levels. In contrast, SlATG8b displayed elevated expression specifically during the color-breaking stage of tomato fruit ripening, while its expression was reduced during the green, orange, and red ripening stages. SlATG8d exhibited higher expression levels in the color-breaking, orange ripening, and red ripening stages compared to the green ripening stage. SlATG8e demonstrated significantly higher expression levels during both the color-breaking and red ripening stages, approximately twice as much as in the green ripening stage. Conversely, SlATG8f displayed its lowest expression level during the green ripening stage, peaked during the color-breaking stage, and decreased gradually from the color-breaking stage to the red ripening stage. Lastly, the expression of the SlATG8g gene was notably higher during the red ripening stage, lowest during the color-breaking stage, and relatively consistent during the green and orange ripening stages.
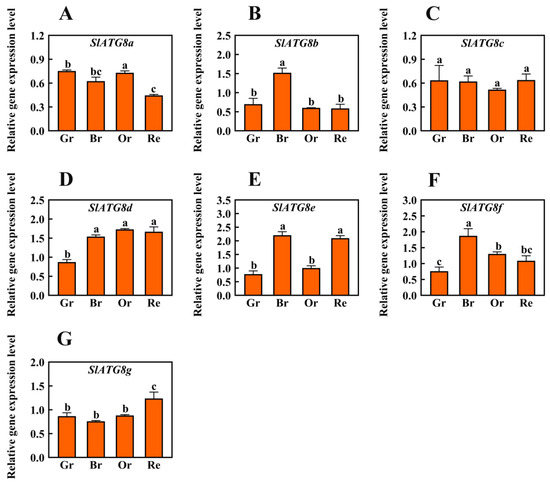
Figure 1.
Differential Expression Analysis of SlATG8s During Tomato Fruit Ripening. The expression profiles of SlATG8s during different stages of tomato fruit ripening are shown. The stages of fruit ripening are as follows: green ripe (Day 30), breaker (Day 35), orange ripe (Day 40), and red ripe (Day 45). Abbreviations: Gr—Green Ripe; Br—Breaker; Or—Orange Ripe; Re—Red Ripe. (A–G)—SlATG8a, SlATG8a, SlATG8b, SlATG8c, SlATG8d, SlATG8e, SlATG8f, SlATG8g genes relative expression. The presented data represent the mean values, and error bars represent the standard deviation. The same letter indicates no significant difference in mean values at p < 0.05 (Duncan’s test).
2.2. Overexpression of SlATG8f Promotes Pericarp Parenchyma Formation in Tomato Fruit
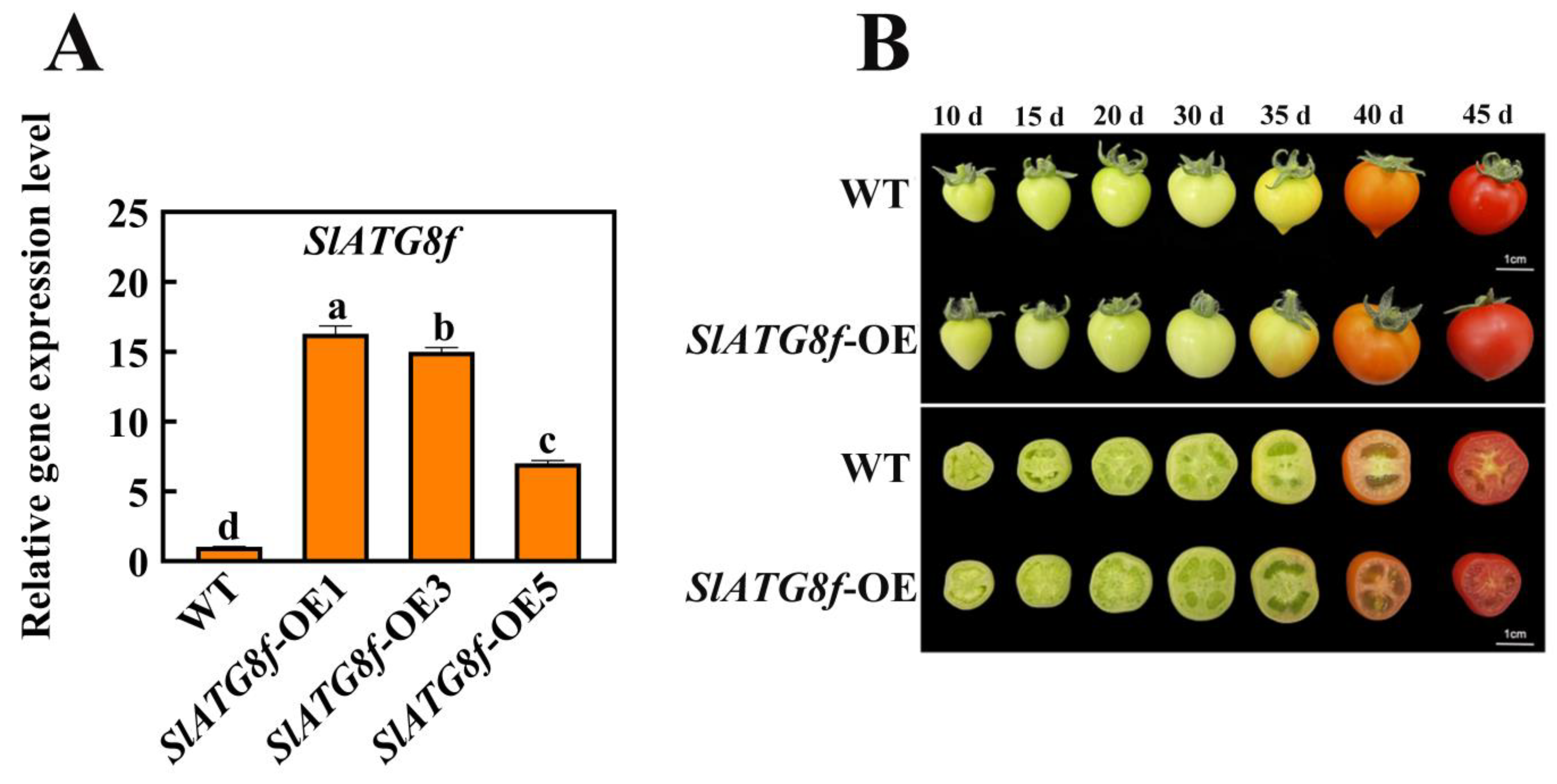
In this study, transgenic Micro-Tom tomato plants were generated with overexpression of the SlATG8f coding sequence (CDS) under the control of the 35 S promoter. Quantitative real-time polymerase chain reaction (qRT-PCR) results demonstrated a substantial increase in SlATG8f gene expression in the SlATG8f-overexpressing (OE) plants, reaching up to 16-fold higher levels compared to the wild-type (WT) plants (Figure 2A).
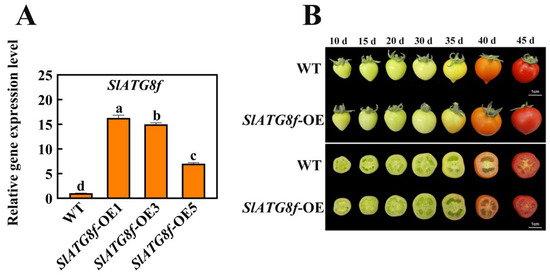
Figure 2.
Identification of SlATG8f OE Plants Line and Phenotypic Observation of Fruits. (A) Quantitative real-time PCR (qRT-PCR) analysis of SlATG8f gene expression in the SlATG8f OE plants line compared to the wild-type (WT) plants. (B) Phenotypic observations and cross-sections at 10 days, 15 days, 20 days, 30 days, 35 days, 40 days, and 45 days after flowering. Abbreviations: WT—Wild Type; SlATG8f-OE—SlATG8f overexpressing plants. The data shown are the mean values of three replicates, and the error bars represent the standard deviation. The same letter indicates no significant difference in mean values at p < 0.05 (Duncan’s test).
Phenotypic changes in fruit development were observed among the different tomato plants during fruit ripening (Figure 2B). While there were no significant differences in fruit phenotypes between the SlATG8f OE plants and WT fruits, a notable dissimilarity was observed upon slicing the fruits. The pericarp parenchyma of the SlATG8f OE plants’ fruits exhibited accelerated development compared to that of the WT fruits with the pericarp parenchyma being more plump and filled in the SlATG8f OE plants’ fruits than in the WT fruits. In the early stages of fruit development, the pericarp parenchyma in the WT fruits was relatively scant, whereas the pericarp tissue of the SlATG8f OE plants’ fruits was already completely filled. By the 35th day of fruit development, both the SlATG8f OE plants and WT fruits had entered the liquefaction phase. However, the SlATG8f OE plants’ fruits displayed a higher degree of liquefaction in their pericarp parenchyma. In the late stages of fruit development, both the SlATG8f OE plants’ and WT fruits were fully matured, and their pericarp parenchyma had undergone significant liquefaction with no conspicuous differences between them. Nonetheless, the pericarp parenchyma tissue of the SlATG8f OE plants’ fruits remained fuller compared to that of the WT fruits.
2.3. Overexpression of SlATG8f Enhances Autophagic Activity in Tomato Fruit
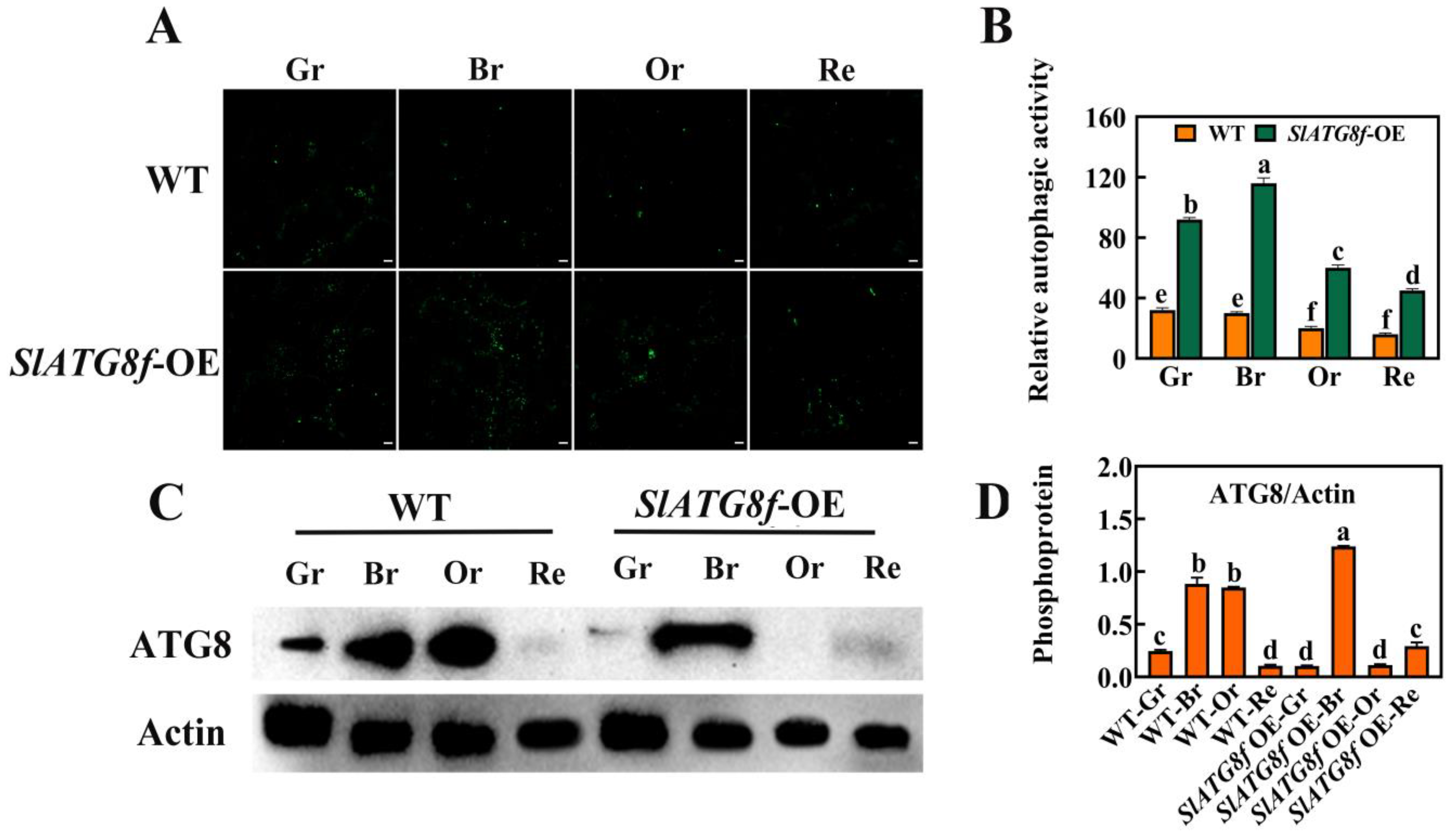
To investigate whether SlATG8f enhances autophagic activity in tomato fruit, this study employed MDC staining and protein immunoblotting (Western blotting) to assess the changes in autophagic activity between the SlATG8f OE plants’ and WT fruits at different stages of ripening (Figure 3). The MDC staining results revealed (Figure 3A,B) that, compared to the WT fruits, the SlATG8f OE plants’ fruits exhibited a higher intensity of green fluorescence corresponding to MDC-stained structures, indicative of autophagosomes. While the number of autophagosomes decreased from the green ripe stage to the red ripe stage in the WT fruits, the number of autophagosomes gradually increased from the green ripe stage to the breaker stage in the SlATG8f OE plants’ fruits, peaking at the breaker stage before declining.
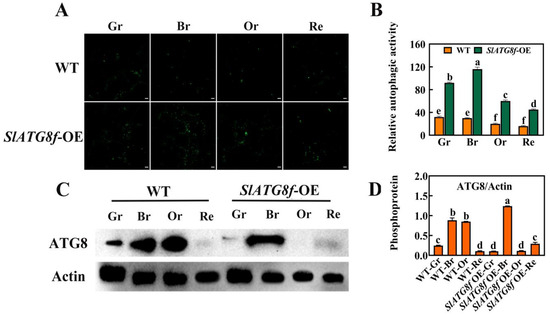
Figure 3.
Effects of SlATG8f OE Plants on Autophagic Activity in Tomato Fruits. (A) Autophagosomes stained with MDC (green) in SlATG8f OE plants’ and WT fruits during different stages: green ripe (Gr), breaker (Br), orange ripe (Or), and red ripe (Re). Scale bars = 20 μm. (B) Relative autophagic activity quantified by counting the number of autophagosomes stained with MDC in each image from Panel A. (C) Western blot analysis of ATG8 protein expression in SlATG8f OE plants’ and WT fruits at different stages using Actin as a loading control. (D) Quantification of the intensity of each band from Panel C normalized to the intensity of the Actin band. ATG8/Actin ratio represents the relative expression of ATG8 protein. Abbreviations: white scale—20 μm; Gr—Green Ripe; Br—Breaker; Or—Orange Ripe; Re—Red Ripe; WT—Wild Type; SlATG8f-OE—SlATG8f overexpressing plants. The data shown are the mean values of three replicates, and the error bars represent the standard deviation. The same letter indicates no significant difference in mean values at p < 0.05 (Duncan’s test).
Protein immunoblotting was employed to analyze the ATG8 protein levels during tomato fruit ripening, and the quantification of the corresponding bands was performed (Figure 3C,D). The results demonstrated that the ATG8 protein band intensity was strongest in the breaker stage of the SlATG8f OE plants’ fruits, whereas it was weakest in the green ripe, orange ripe, and red ripe stages, almost undetectable. In contrast, the ATG8 protein band signal was strong in the breaker and orange ripe stages of the WT fruits. The grayscale values of the extracted ATG8 protein bands indicated that, in the WT fruits, ATG8 protein expression gradually increased from the green ripe stage to the breaker stage followed by a decrease from the breaker stage to the orange ripe stage with the lowest expression in the red ripe stage. In the SlATG8f OE plants, ATG8 protein expression was significantly enhanced at the breaker stage, while it remained relatively low during the green ripe, orange ripe, and red ripe stages. These findings suggest that SlATG8f might be involved in the formation of autophagosomes during the breaker stage of tomato fruit.
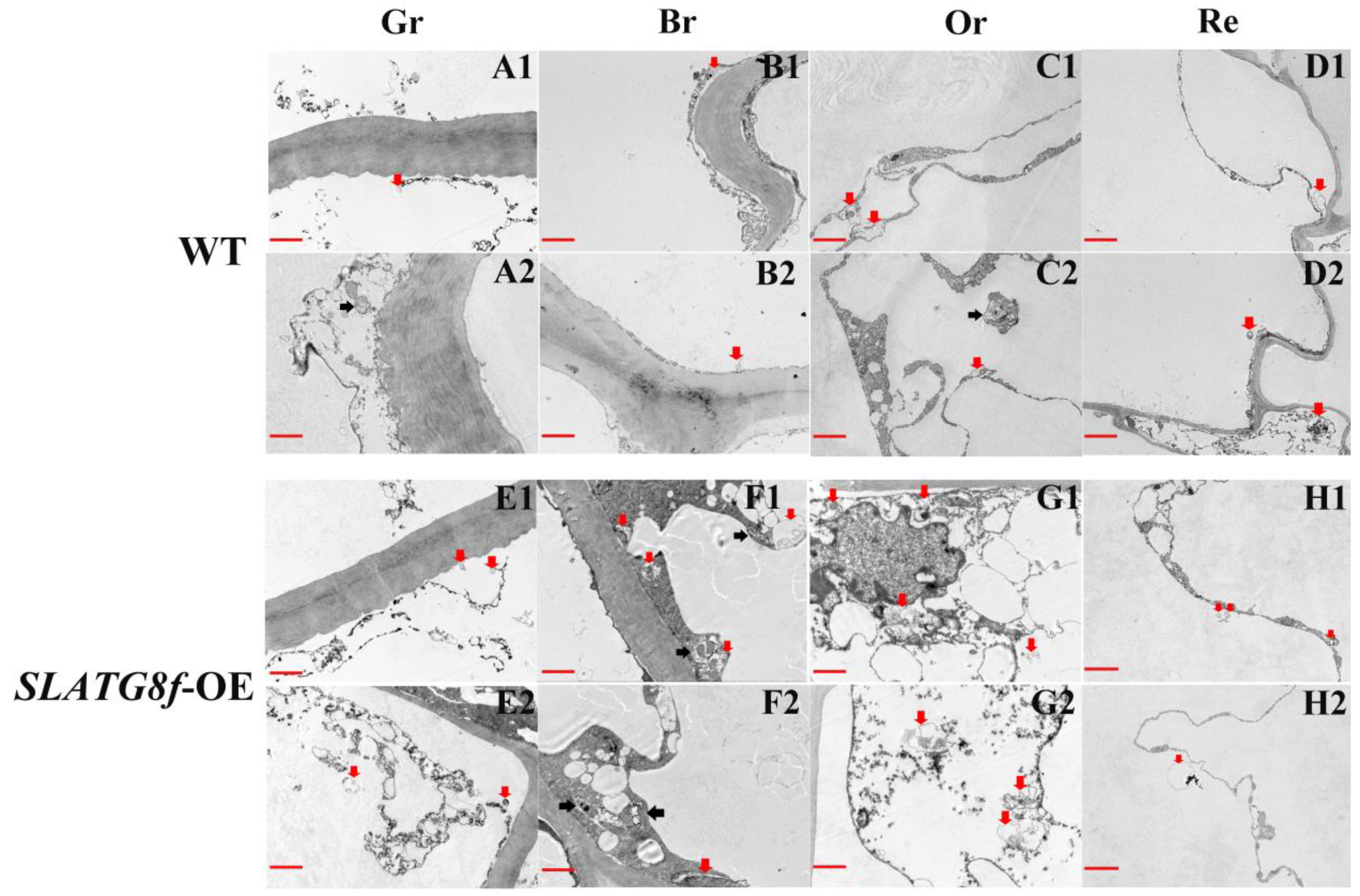
To gain deeper insights into whether the overexpression of ATG8f promotes an increase in autophagic flux during tomato fruit ripening, this study conducted an analysis of autophagic body structural changes using electron microscopy across four stages of fruit maturation (Figure 4). The results revealed distinct patterns of autophagic activity among the different fruit ripening stages. In the wild-type (WT) fruits, there was a relatively high number of autophagic bodies during the orange ripe stage, while autophagic activity was low in the other stages (Figure 4A–D). The SlATG8f OE plants’ fruits exhibited the highest autophagic activity during the breaker stage, displaying the highest number of visible autophagic bodies (Figure 4F). The following stage with notable autophagic activity was the orange ripe stage (Figure 4G), while autophagic activity was lower in the green ripe and red ripe stages (Figure 4E,H). Compared to the WT fruits, the SlATG8f OE plants’ fruits contained a greater abundance of autophagic structures within the cells, particularly evident during the breaker and orange ripe stages of the SlATG8f OE plants’ fruits. Notably, not only were numerous autolysosomal structures observed, but also, early autophagosome structures were evident (Figure 4F,G; indicated by double arrows and single arrows, respectively). These findings underscore that overexpression of SlATG8f significantly enhances autophagic flux during tomato fruit ripening.
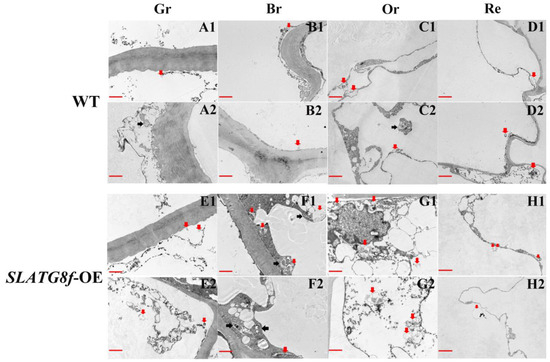
Figure 4.
Effects of SlATG8f OE Plants on Autophagic Activity in Tomato Fruits. (A1–D2) Autophagic structures in WT tomato fruits at four stages of ripening. (E1–H2) Autophagic structures in SlATG8f OE plants’ tomato fruits at four stages of ripening. Scale bars = 2 μm. WT—Wild Type; SlATG8f-OE—SlATG8f overexpressing plants. Abbreviations: red scale—2 μm; Red arrows indicate autolysosomes (single-membrane structures with cytoplasmic components degraded), and black arrows indicate autophagosomes (double or multi-membrane vesicular structures containing cytoplasmic components). Gr—Green Ripe; Br—Breaker; Or—Orange Ripe; Re—Red Ripe; WT—Wild Type.
2.4. Overexpression of SlATG8f Facilitates Tomato Fruit Ripening
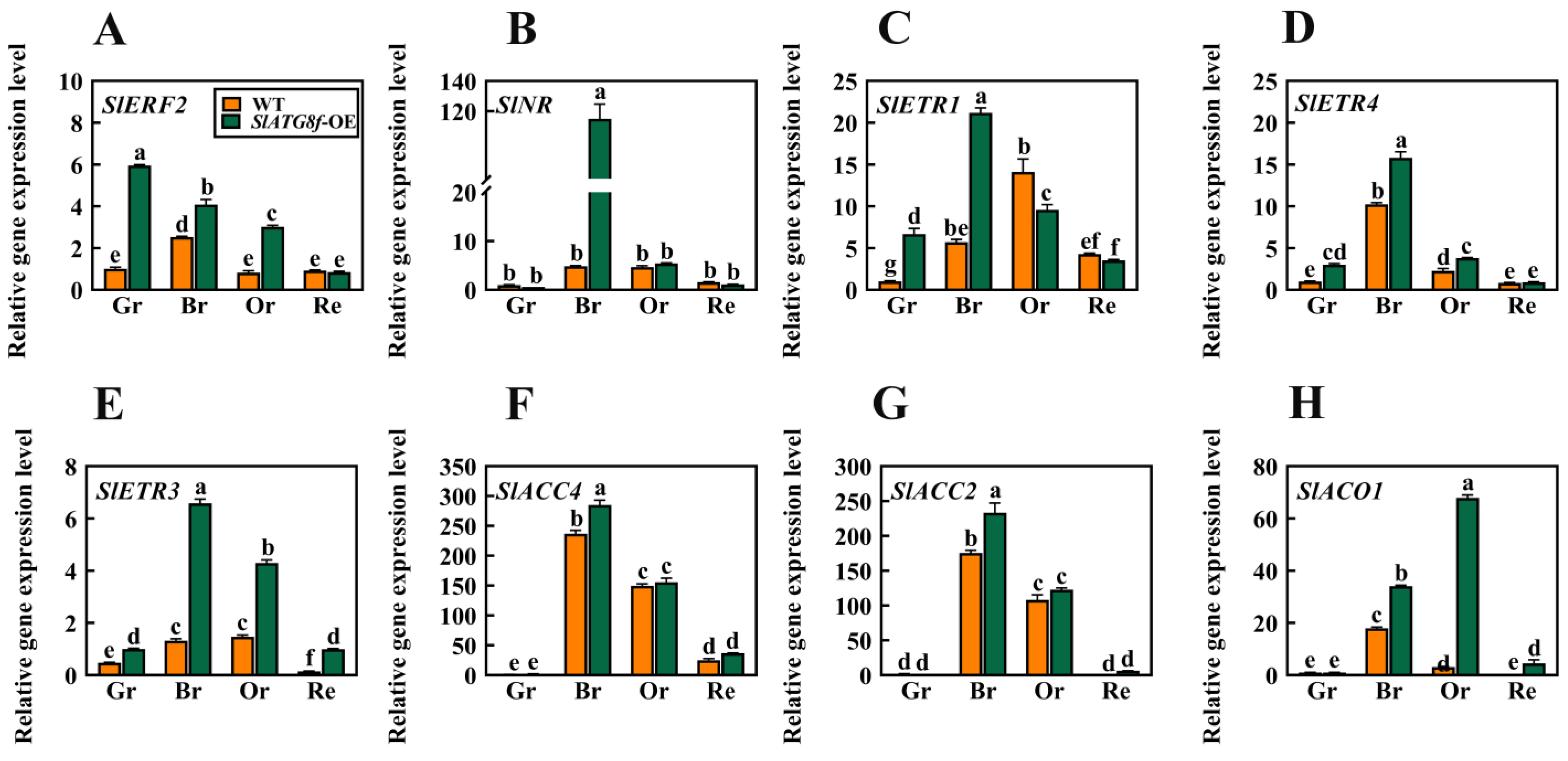
In earlier investigations, it was observed that the degree of liquefaction of the pericarp parenchyma in the SlATG8f OE plants was higher than that in the WT plants at the 35th day of fruit development. Based on this finding, this study hypothesized that SlATG8f might promote tomato fruit ripening. To validate this hypothesis, a selection of genes associated with ethylene signaling was screened, including ethylene signal transduction gene ERF2; ethylene receptor genes NR, ETR1, ETR3, and ETR4; and ethylene synthesis genes ACC2, ACC4, and ACO1. Their expression level changes during fruit ripening were analyzed (Figure 5).
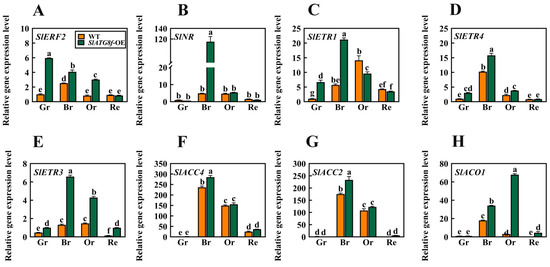
Figure 5.
Effects of SlATG8f OE Plants on the Expression of Ethylene-Related Genes in Tomato Fruits. (A) qRT-PCR analysis of the expression of the ethylene signal transduction gene ERF2. (B–E) qRT-PCR analysis of the expression of ethylene receptor genes NR, ETR1, ETR4, and ETR3. (F–H) qRT-PCR analysis of the expression of ethylene synthesis genes ACC4, ACO1, and ACC2. Annotations: Gr—Green Ripe; Br—Breaker; Or—Orange Ripe; Re—Red Ripe; WT—Wild Type; SlATG8f-OE—SlATG8f overexpressing plants.The data shown are the mean values of three replicates, and the error bars represent the standard deviation. The same letter indicates no significant difference in mean values at p < 0.05 (Duncan’s test).
The results indicated that, in the SlATG8f OE plants’ fruits, the expression of SlERF2 was upregulated during the green ripe, breaker, and orange ripe stages (Figure 5A). SlNR was significantly upregulated during the breaker stage of the SlATG8f OE plants’ fruits (Figure 5B). The expression levels of SlETR1 and SlETR4 were elevated during the green ripe and breaker stages of the SlATG8f OE plants’ fruits, but their expression levels were not notably high during the orange ripe and red ripe stages (Figure 5C,D). SlAETR3 exhibited increased expression during the breaker, orange ripe, and red ripe stages of the SlATG8f OE plants’ fruits (Figure 5E). SlACO1 showed significant upregulation during the breaker and orange ripe stages of the SlATG8f OE plants’ fruits (Figure 5G). However, the expression levels of SlACC4 and SlACC2 during the breaker and orange ripe stages of the SlATG8f OE plants’ fruits did not differ significantly from those of the wild type (Figure 5F,H). These findings suggest that SlATG8f might influence ethylene production by regulating the expression of SlACO1 and, thereby, promoting fruit ripening.
2.5. Overexpression of SlATG8f Improvements of Tomato Fruit Quality
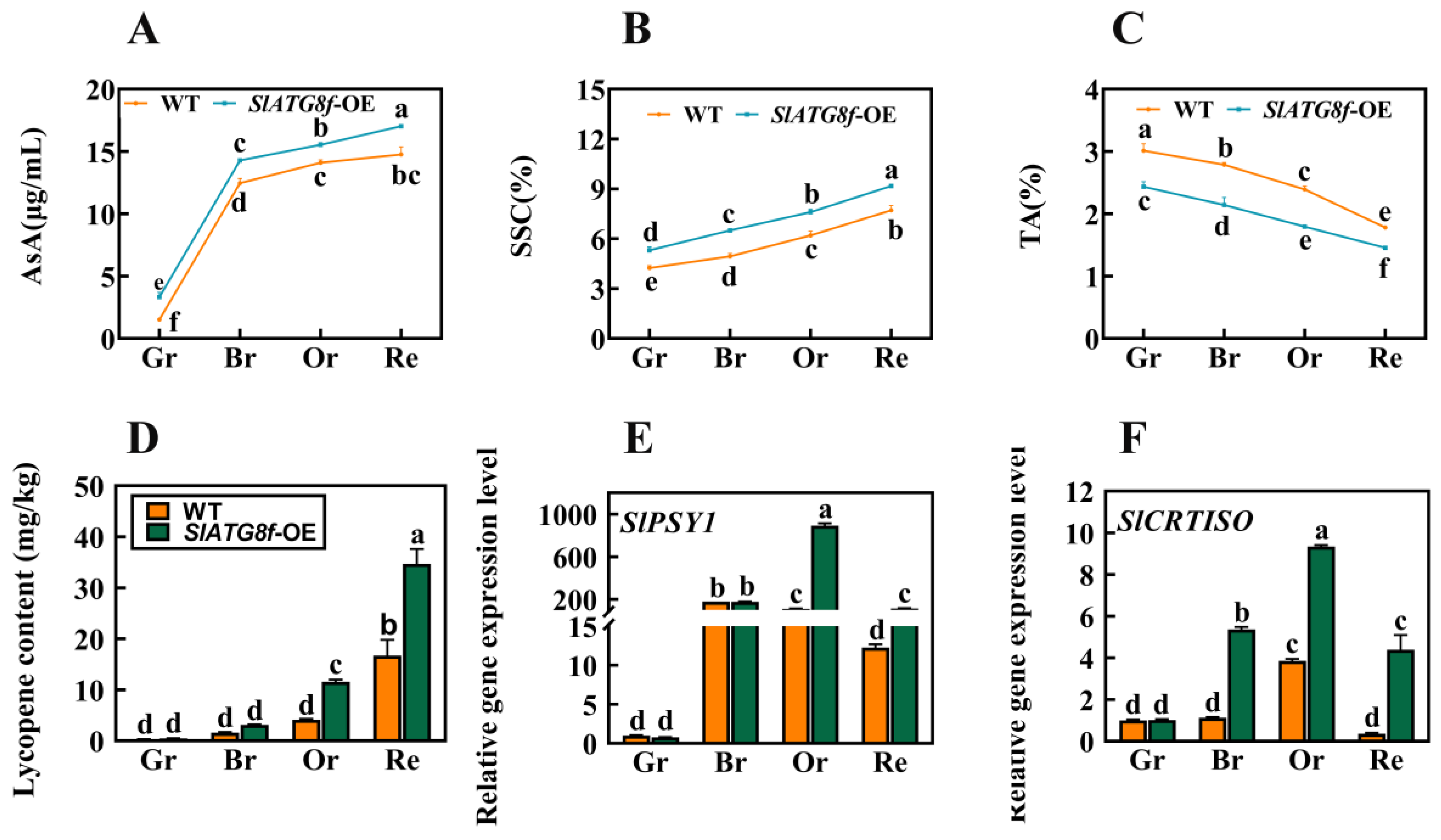
To further comprehend whether the promotion of fruit ripening by the SlATG8f OE plants influences fruit quality and flavor, this study evaluated the content of ascorbic acid (AsA), soluble solids content (SSC), titratable acidity (TA), and tomato lycopene levels. The results revealed notable variations in these components. At all ripening stages of the SlATG8f OE plants’ fruits, the content of both ascorbic acid and soluble solids was higher compared to that of the WT fruits (Figure 6A,B). Conversely, the titratable acidity content was consistently lower in the SlATG8f OE plants’ fruits compared to that in the WT (Figure 6C). During the orange ripe and red ripe stages of fruit development, the SlATG8f OE plants’ fruits exhibited significantly higher levels of tomato lycopene content compared to the WT fruits (Figure 6D). The key gene involved in tomato lycopene synthesis, SlPSY1, was notably upregulated during the orange ripe stage of the SlATG8f OE plants’ fruits (Figure 6E). Additionally, the expression of the tomato carotenoid isomerase gene, SlCRTISO, was elevated during the breaker, orange ripe, and red ripe stages of the SlATG8f OE plants’ fruits compared to that of the WT fruits (Figure 6F). These findings led to the hypothesis that overexpression of SlATG8f could enhance tomato fruit quality, potentially mediating the biosynthesis pathway of tomato lycopene and, thereby, influencing the color change in tomato fruits.
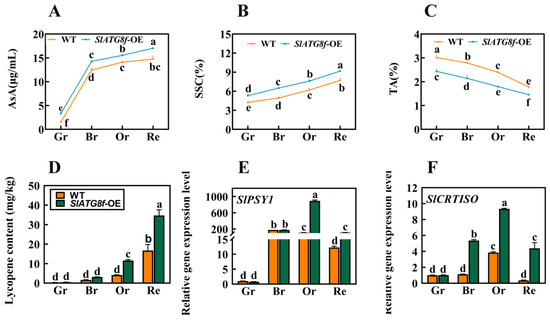
Figure 6.
Effects of SlATG8f OE Plants on Pigment Accumulation, Soluble Solids, and Acidity in Tomato Fruits. (A) Ascorbic acid (AsA) content in tomato fruits. (B) Soluble solids (SSC) content in tomato fruits (expressed as a percentage). (C) Titratable acidity (TA) content in tomato fruits (expressed as a percentage). (D) Lycopene content in tomato fruits. (E,F) qRT-PCR analysis of the expression levels of lycopene synthesis genes SlPSY1 and SlCRTISO. qRT-PCR primer sequences are provided in Supplementary Table S1. Annotations: Gr—Green Ripe; Br—Breaker; Or—Orange Ripe; Re—Red Ripe; WT—Wild Type; SlATG8f-OE—SlATG8f overexpressing plants. The data shown are the mean values of three replicates, and the error bars represent the standard deviation. The same letter indicates no significant difference in mean values at p < 0.05 (Duncan’s test).
3. Discussion
Autophagy is a crucial process in eukaryotic cells for the degradation and selective elimination of cytoplasmic components, playing a significant role in the bulk and selective degradation of targeted substances [18]. In plant cells, autophagy involves the transfer of damaged or redundant macromolecules and organelles to vacuoles for degradation, which holds key roles in maintaining cellular homeostasis, growth, and development as well as responding to environmental stress [19]. However, the role of autophagy in fruit development and ripening remains largely unexplored, and the analysis of ATG genes or ATG proteins in this context is seldom reported. Bernard et al. suggested that the transcriptional upregulation of ATG genes enhances autophagy [20]. In our study, the expression of tomato SlATG8s was detected during fruit ripening, indicating the presence of autophagy activity in these fruits. SlATG8b, SlATG8d, SlATG8e, and SlATG8f genes exhibited higher expression levels during the breaker stage; SlATG8d gene showed higher expression during the orange ripe stage; and SlATG8d, SlATG8e, and SlATG8g genes showed higher expression during the red ripe stage. This suggests that these genes might activate distinct regulatory mechanisms at different stages of tomato fruit ripening. In Arabidopsis, nine ATG8 homologous genes have been identified that encode proteins whose C-terminal cleavage is ATG4-dependent followed by the formation of ATG8-PE complexes [21]. ATG8-PE complexes promote the initiation of autophagosomes [22]. However, in our study, changes in the expression of ATG8 protein levels were detected using Western blotting, while the expression of ATG8-PE complexes was not assessed, suggesting the need for further investigation.
The presence of multiple autophagosome vesicles in tomato fruit vacuoles indicates the pivotal role of autophagy in fruit ripening. Through transmission electron microscopy, an increased number of vesicles was observed in the SlATG8f OE plants’ fruits compared to those in the WT with the peak vesicle count occurring during the turning stage, a result corroborated by MDC staining. As fruit undergoes various metabolic shifts during the transition from green to red, including the transformation of chloroplasts to chromoplasts [23], it suggests that different autophagy mechanisms might take place to sustain energy for fruit maturation. Prior research on peppers and strawberries indicated a higher autophagy activity in yellow peppers compared to that in red peppers as well as two strong autophagy fluxes during the white and ripe stages of strawberry maturation [12,14]. Our findings also suggest that the peak autophagy activity during the breaker and orange ripe stages of tomato fruit ripening, enhanced autophagy activity during the breaker stage in the SlATG8f OE plants’ genotype, is advantageous for energy balance during fruit maturation.
Autophagy primarily participates in plant aging and senescence [24]. The process of fruit ripening is also an aging process that is dependent on cellular vitality and gene regulation [25]. Among these, ethylene-related genes play pivotal roles in tomato fruit ripening [26]. Our research revealed increased liquefaction of the locule in the SlATG8f OE plants’ fruits during the breaker stage accompanied by upregulated expression of ethylene-related genes. This, in conjunction with the high expression of ATG8 protein during the breaker stage of the SlATG8f OE plants’ fruits, further confirms the promoting role of autophagy in tomato fruit ripening. Simultaneously, autophagy induced during fruit ripening enhances fruit coloration, increases levels of ascorbic acid and soluble solids, reduces acidity, and improves fruit texture and quality—possibly arising from the dual effect of autophagy. Previous studies reported that autophagy promotes strawberry fruit ripening while delaying senescence [27]. Our results suggest the presence of autophagy during tomato fruit ripening with the peak autophagy activity during the breaker and orange ripe stages; SlATG8f OE plants effectively enhance autophagy activity during the breaker stage, promoting fruit maturation and enhancing fruit quality. These findings have significant implications for understanding the role of autophagy in tomato fruit growth and development, exploring the regulatory mechanisms of autophagy in fruit, and advancing tomato breeding strategies.
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
Micro-Tom tomato seeds were disinfected with 0.3% sodium hypochlorite for 10 min followed by three rinses with sterile water. After soaking in water at 55 °C for 15 min and then in water at 28 °C for 6 h, the seeds were germinated in darkness at 28 °C. Once the seeds were visibly germinated, they were sown in trays containing soilless growth medium. The growth chamber conditions were set at 18 h light/6 h dark with a constant temperature of 25 °C. Fruit samples were collected at different ripening stages: green ripe (day 30), breaker (day 35), orange ripe (day 40), and red ripe (day 45). For each stage, 4–5 fruits were sampled, and the middle portion of the peel and flesh was harvested, frozen in liquid nitrogen, and stored at −80 °C. Gr: green ripe; Br: breaker; Or: orange ripe; Re: red ripe.
4.2. Generation of Transgenic Plants
The full-length SlATG8f coding sequence (CDS) was amplified from tomato cDNA using the specific primer (Supplemental Table S2) into pBWA(V)HS and then introduced into the plant overexpression vector. The resulting pro35S, SlATG8f, was transformed into Agrobacterium tumefaciens strain GV3101 and then transformed into Micro-Tom tomato with Agrobacterium-mediated transformation. During tomato infection, the tomato seeds were first surface-sterilized with 75% alcohol and sodium hypochlorite solution followed by immersion cleaning in sterilized distilled water to obtain sterile materials. After that, the sterilized seeds were spread on 1/2 MS solid agar medium (pH = 5.8) and left to germinate, and the cotyledons were assessed for Agrobacterium infection. Kanamycin was used for selecting stable transformants based on their resistance. Finally, the T2 generation was chosen as the experimental material.
4.3. Quantitative Real-Time PCR (qRT-PCR)
Quantitative real-time PCR was employed to study the expression of SlATG8s and ethylene-related genes in tomato fruits. Frozen tomato fruit samples were used for RNA extraction with a TIANGEN kit (DP432, TIANGEN), and cDNA synthesis was performed using a reverse transcription kit (KR118, TIANGEN). Specific primers were designed using Primer 5.0 software with the Actin gene selected as the internal reference (Supplementary Table S1). qRT-PCR was conducted using a 7500 Real-Time PCR System (4351107, Thermo Fisher Scientific, Guangzhou, China) and SYBR Green Supermix (RK02001, Tiangen, Shanghai, China). The experiments were carried out with three biological replicates and three technical replicates.
4.4. Transmission Electron Microscopy Observation
Tomato fruits were soaked in a 100 nmol/L rapamycin solution for 12 h to induce autophagy. The fruit tissues were rapidly cut into 1–2 mm3 pieces, fixed in electron microscopy fixative solution (G1102, Servicebio, Wuhan, China) at 4 °C for 2–4 h, and then dehydrated using a series of acetone gradients. The dehydrated tissues were infiltrated and embedded in mixtures of dehydrating agents and epoxy resin in ratios of 3:1, 1:1, and 1:3. Ultrathin sections (50 nm) were obtained using an ultrathin slicer, and these sections were placed on copper grids. The sections were stained with uranyl acetate for 10–15 min and then with lead citrate for 1–2 min at room temperature. Transmission electron microscopy was performed using a JEM-1400PLUS (JEM-1400 plus, Jeol, Tokyo, Japan) instrument.
4.5. Dansylcadaverine (MDC) Staining
Tomato fruits were soaked in a 100 nmol/L rapamycin solution for 12 h to induce autophagy. The tomato fruit sections were prepared according to a previous protocol [28]. The sections were immediately stained with 100 μM MDC (30432, Sigma-Aldrich, Shanghai, China) under vacuum and dark conditions for 30 min and then washed twice with phosphate-buffered saline (PBS, P1020, Solarbio, Beijing, China). Autophagosome structures were observed using an LSM-780 confocal microscope (LSM-780, Carl Zeiss, Shanghai, China) with excitation at 405 nm and emission ranging from 400 to 580 nm.
4.6. Protein Blot (Western Blotting)
Tomato fruits were soaked in a 100 nmol/L rapamycin solution for 12 h to induce autophagy. The fruit tissues were frozen in liquid nitrogen and used for total protein extraction using a kit (PTE011, Coolaber, Beijing, China). SDS-PAGE gel electrophoresis was performed (P1200, Solarbio, Beijing, China) to separate the proteins. The membrane was blocked with 5% skim milk for 2 h and then washed with TBST. The PVDF membrane was cut into strips and incubated with primary antibodies (1:1000) overnight at 4 °C. After washing with TBST three times, the membrane was incubated with secondary antibodies (1:1000) against rabbit ATG8a polyclonal antibody (Ab77003, Abcam, Shanghai, China) for 1 h at room temperature. Chemiluminescence was detected using a chemiluminescence detection kit (34080, Thermo Fisher Scientific, Guangzhou, China).
4.7. Detection of Lycopene Content, Ascorbic Acid, Soluble Solids (SSC), and Titratable Acidity (TA)
The lycopene content of the tomato fruits was determined according to Sun et al.’s method. Five grams of tomato flesh and peel tissue were frozen and ground into powder. The powder was mixed with 50 mL of hexane–acetone–ethanol (2:1:1, v/v) and shaken. After adding 15 mL of water and allowing for phase separation, the organic phase (hexane) absorbance was measured at 503 nm to calculate the lycopene concentration. The molar extinction coefficient was 17.2 L mol−1·m−1, and the units were expressed as mg·kg−1 based on the fresh weight [29]. Ascorbic acid was measured using a kit (A009-1-1, Nanjing Jiancheng Bioengineering Institute, Nanjing, China). The SSC and TA content measurements were performed using a PAL-BX/ACD1 sugar acidity meter (ATAGO, Tokyo, Japan) for SSC (%) and TA (%) in the fruit. The experiments included three biological replicates and three technical replicates.
5. Conclusions
In this study, quantitative analysis of the expression of the ATG8 gene family members during tomato fruit ripening revealed that the expression levels of most ATG8 family members were higher during the fruit color transition period or late stages of fruit ripening. Among them, the expression of SlATG8f gradually increased from the green ripe stage to the breaker stage and then gradually decreased from the breaker stage to the red ripe stage. In the SlATG8f OE plants line, the locular gel tissue of the tomato fruits appeared more plump compared to that in the wild type (WT), and the liquefaction rate of the locular gel tissue was faster, indicating that the SlATG8f gene positively regulates tomato fruit development. The MDC staining and electron microscopy observations revealed that the number of autophagosomes in the SlATG8f OE plants’ fruits was significantly higher than that in the WT, especially during the breaker stage, with a substantial increase in the number of autophagosomes and autolysosomes in the fruit flesh cells, indicating enhanced autophagic activity. Western blot results demonstrated a significant increase in ATG8 protein expression during the breaker stage in the SlATG8f OE plants’ fruits, and the expression level of SlATG8f correlated positively with autophagic activity. Overexpression of SlATG8f induced the formation of autophagosomes in the tomato fruit cells, enhancing autophagic activity. The increased autophagic flux due to SlATG8f overexpression accelerated fruit ripening and upregulated the expression of ethylene-related genes, regulating fruit ripening. Taken together, our findings suggest that autophagy is involved in regulating tomato fruit ripening and senescence. These discoveries contribute to a deeper understanding of the role of autophagy in the process of tomato fruit ripening, elucidate the key factors that regulate fruit quality, and provide a theoretical basis for improving tomato varieties.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants12183339/s1, Table S1: ATG8 family members, ethylene-related genes, SlPSY1, SlCRTISO primer design table, Table S2: SlATG8f clone-specific primers.
Author Contributions
C.W. and T.L., writing—original draft, formal analysis, data curation, software, methodology, and validation. Z.H., software and validation. W.X., Y.L. and J.Y., writing—review and editing. W.X., funding acquisition, project administration, resources, and supervision. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (32260754 and 31760594), the Special Project for Cultivating Innovative Talents of Thousand Levels in Guizhou Province ((2018)02), the Guizhou University of Research Start-up Fund for New Faculty ((2016)48), and Platform Construction Project of Facility Vegetable Engineering Research Center in Guizhou Higher Education Institutions (Guizhou Education and Technology [2022] No. 040).
Data Availability Statement
Data are available in the manuscript and in the Supplementary Materials.
Acknowledgments
The authors are grateful to the National and Local Vegetable Engineering Center (Guizhou), the Laboratory of the Department of Horticulture (Agricultural College of Guizhou University), and Zhejiang Provincial Key Laboratory of Horticultural Plant Integrative Biology for their support in this project.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Michaeli, S.; Galili, G.; Genschik, P.; Fernie, A.R.; Avin-Wittenberg, T. Autophagy in Plants—What’s New on the Menu? Trends Plant Sci. 2016, 21, 134–144. [Google Scholar] [CrossRef]
- Marshall, R.S.; Vierstra, R.D. Autophagy: The Master of Bulk and Selective Recycling. Annu. Rev. Plant Biol. 2018, 69, 173–208. [Google Scholar] [CrossRef]
- Wen, X.; Klionsky, D.J. An overview of macroautophagy in yeast. J. Mol. Biol. 2016, 428, 1681–1699. [Google Scholar] [CrossRef]
- Tsukada, M.; Ohsumi, Y. Isolation and characterization of autophagy-defective mutants of Saccharomyces cerevisiae. FEBS Lett. 1993, 333, 169–174. [Google Scholar] [CrossRef]
- Meijer, W.H.; van der Klei, I.J.; Veenhuis, M.; Kiel, J.A. ATG genes involved in non-selective autophagy are conserved from yeast to man, but the selective Cvt and pexophagy pathways also require organism-specific genes. Autophagy 2007, 3, 106–116. [Google Scholar] [CrossRef]
- Hafrén, A.; Macia, J.L.; Love, A.J.; Milner, J.J.; Drucker, M.; Hofius, D. Selective autophagy limits cauliflower mosaic virus infection by NBR1-mediated targeting of viral capsid protein and particles. Proc. Natl. Acad. Sci. USA 2017, 114, E2026–E2035. [Google Scholar] [CrossRef]
- Ren, W.; Liu, N.; Sang, C.; Shi, D.; Zhou, M.; Chen, C.; Qin, Q.; Chen, W. The Autophagy Gene BcATG8 Regulates the Vegetative Differentiation and Pathogenicity of Botrytis cinerea. Appl. Environ. Microbiol. 2018, 84, e02455-17. [Google Scholar] [CrossRef] [PubMed]
- Zhen, X.; Xu, F.; Zhang, W.; Li, N.; Li, X. Overexpression of rice gene OsATG8b confers tolerance to nitrogen starvation and increases yield and nitrogen use efficiency (NUE) in Arabidopsis. PLoS ONE 2019, 14, e0223011. [Google Scholar] [CrossRef]
- Yoshimoto, K.; Hanaoka, H.; Sato, S.; Kato, T.; Tabata, S.; Noda, T.; Ohsumi, Y. Processing of ATG8s, ubiquitin-like proteins, and their deconjugation by ATG4s are essential for plant autophagy. Plant Cell 2004, 16, 2967–2983. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.R.; Vierstra, R.D. Autophagic recycling: Lessons from yeast help define the process in plants. Curr. Opin. Plant Biol. 2005, 8, 165–173. [Google Scholar] [CrossRef]
- Zhai, Y.; Guo, M.; Wang, H.; Lu, J.; Liu, J.; Zhang, C.; Gong, Z.; Lu, M. Autophagy, a Conserved Mechanism for Protein Degradation, Responds to Heat, and Other Abiotic Stresses in Capsicum annuum L. Front. Plant Sci. 2016, 7, 131. [Google Scholar] [CrossRef] [PubMed]
- Ghan, R.; Petereit, J.; Tillett, R.L.; Schlauch, K.A.; Toubiana, D.; Fait, A.; Cramer, G.R. The common transcriptional subnetworks of the grape berry skin in the late stages of ripening. BMC Plant Biol. 2017, 17, 94. [Google Scholar] [CrossRef] [PubMed]
- López-Vidal, O.; Olmedilla, A.; Sandalio, L.M.; Sevilla, F.; Jiménez, A. Is Autophagy Involved in Pepper Fruit Ripening? Cells 2020, 9, 106. [Google Scholar] [CrossRef]
- Sánchez-Sevilla, J.F.; Botella, M.A.; Valpuesta, V.; Sanchez-Vera, V. Autophagy Is Required for Strawberry Fruit Ripening. Front. Plant Sci. 2021, 12, 688481. [Google Scholar] [CrossRef] [PubMed]
- Szabo, K.; Cătoi, A.F.; Vodnar, D.C. Bioactive Compounds Extracted from Tomato Processing by-Products as a Source of Valuable Nutrients. Plant Foods Hum. Nutr. 2018, 73, 268–277. [Google Scholar] [CrossRef]
- Ronen, G.; Cohen, M.; Zamir, D.; Hirschberg, J. Regulation of carotenoid biosynthesis during tomato fruit development: Expression of the gene for lycopene epsilon-cyclase is down-regulated during ripening and is elevated in the mutant Delta. Plant J. Cell Mol. Biol. 1999, 17, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Cheniclet, C.; Rong, W.Y.; Causse, M.; Frangne, N.; Bolling, L.; Carde, J.P.; Renaudin, J.P. Cell expansion and endoreduplication show a large genetic variability in pericarp and contribute strongly to tomato fruit growth. Plant Physiol. 2005, 139, 1984–1994. [Google Scholar] [CrossRef]
- Signorelli, S.; Tarkowski, Ł.P.; Van den Ende, W.; Bassham, D.C. Linking Autophagy to Abiotic and Biotic Stress Responses. Trends Plant Sci. 2019, 24, 413–430. [Google Scholar] [CrossRef]
- Qi, H.; Xia, F.N.; Xiao, S. Autophagy in plants: Physiological roles and post-translational regulation. J. Integr. Plant Biol. 2021, 63, 161–179. [Google Scholar] [CrossRef]
- Bernard, A.; Jin, M.; González-Rodríguez, P.; Füllgrabe, J.; Delorme-Axford, E.; Backues, S.K.; Joseph, B.; Klionsky, D.J. Rph1/KDM4 mediates nutrient-limitation signaling that leads to the transcriptional induction of autophagy. Curr. Biol. 2015, 25, 546–555. [Google Scholar] [CrossRef]
- Woo, J.; Park, E.; Dinesh-Kumar, S.P. Differential processing of Arabidopsis ubiquitin-like Atg8 autophagy proteins by Atg4 cysteine proteases. Proc. Natl. Acad. Sci. USA 2014, 111, 863–868. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.Q.; Ni, T.; Hong, B.; Wang, H.Y.; Jiang, F.J.; Zou, S.; Chen, Y.; Zheng, X.L.; Klionsky, D.J.; Liang, Y.; et al. Dual roles of Atg8-PE deconjugation by Atg4 in autophagy. Autophagy 2012, 8, 883–892. [Google Scholar] [CrossRef] [PubMed]
- McAtee, P.; Karim, S.; Schaffer, R.; David, K. A dynamic interplay between phytohormones is required for fruit development, maturation, and ripening. Front. Plant Sci. 2013, 4, 79. [Google Scholar] [CrossRef] [PubMed]
- Avila-Ospina, L.; Moison, M.; Yoshimoto, K.; Masclaux-Daubresse, C. Autophagy, plant senescence, and nutrient recycling. J. Exp. Bot. 2014, 65, 3799–3811. [Google Scholar] [CrossRef] [PubMed]
- Karlova, R.; Chapman, N.; David, K.; Angenent, G.C.; Seymour, G.B.; de Maagd, R.A. Transcriptional control of fleshy fruit development and ripening. J. Exp. Bot. 2014, 65, 4527–4541. [Google Scholar] [CrossRef]
- Wang, H.; Schippers, J.H.M. The Role and Regulation of Autophagy and the Proteasome During Aging and Senescence in Plants. Genes 2019, 10, 267. [Google Scholar] [CrossRef]
- Thomas, H. Senescence, ageing and death of the whole plant. New Phytol. 2013, 197, 696–711. [Google Scholar] [CrossRef]
- Shinozaki, Y.; Hao, S.; Kojima, M.; Sakakibara, H.; Ozeki-Iida, Y.; Zheng, Y.; Fei, Z.; Zhong, S.; Giovannoni, J.J.; Rose, J.K.; et al. Ethylene suppresses tomato (Solanum lycopersicum) fruit set through modification of gibberellin metabolism. Plant J. Cell Mol. Biol. 2015, 83, 237–251. [Google Scholar] [CrossRef]
- Sun, Q.; Zhang, N.; Wang, J.; Zhang, H.; Li, D.; Shi, J.; Li, R.; Weeda, S.; Zhao, B.; Ren, S.; et al. Melatonin promotes ripening and improves quality of tomato fruit during postharvest life. J. Exp. Bot. 2015, 66, 657–668. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).