


Effects of Light Intensity on Growth and Quality of Lettuce and Spinach Cultivars in a Plant Factory

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
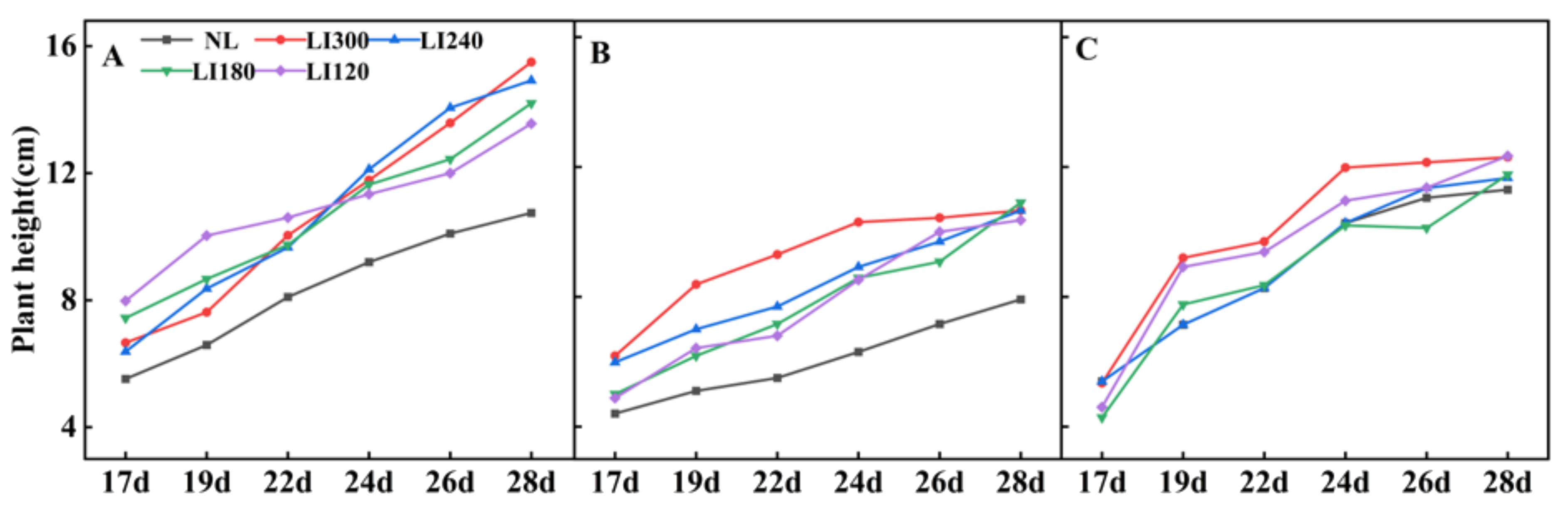
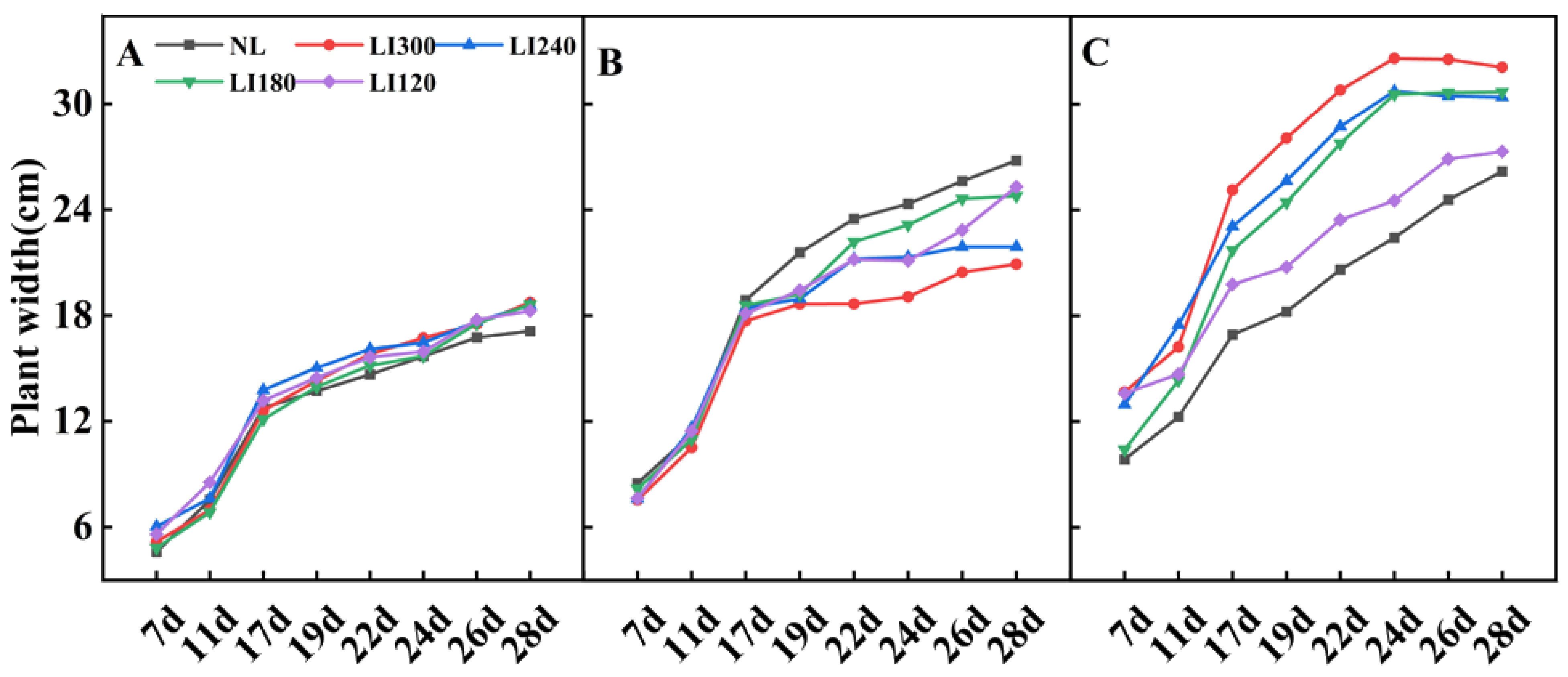
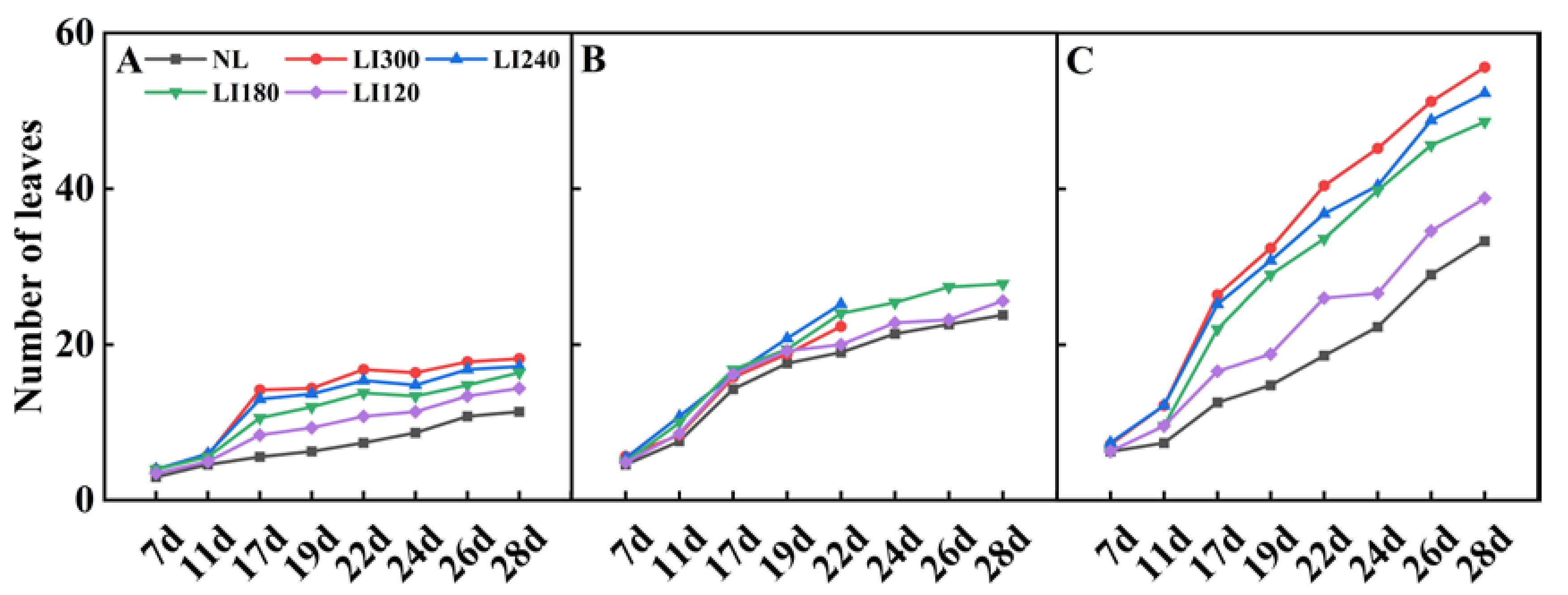
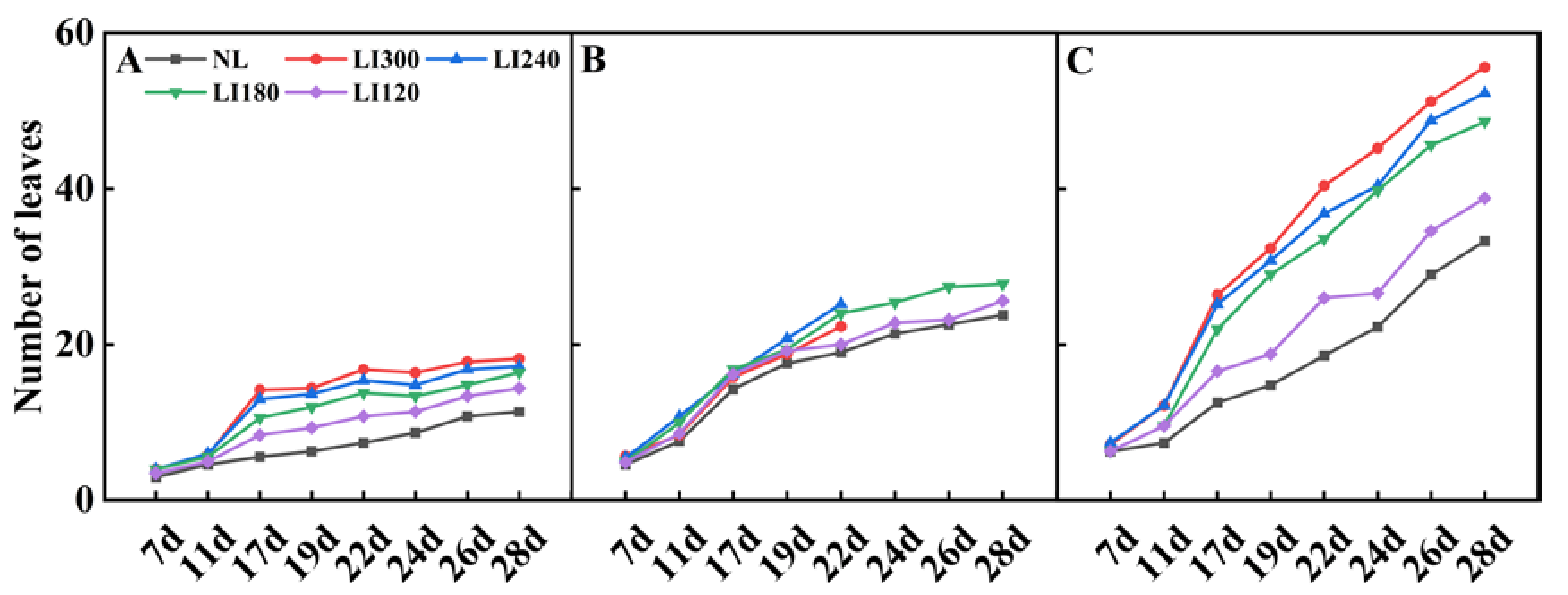
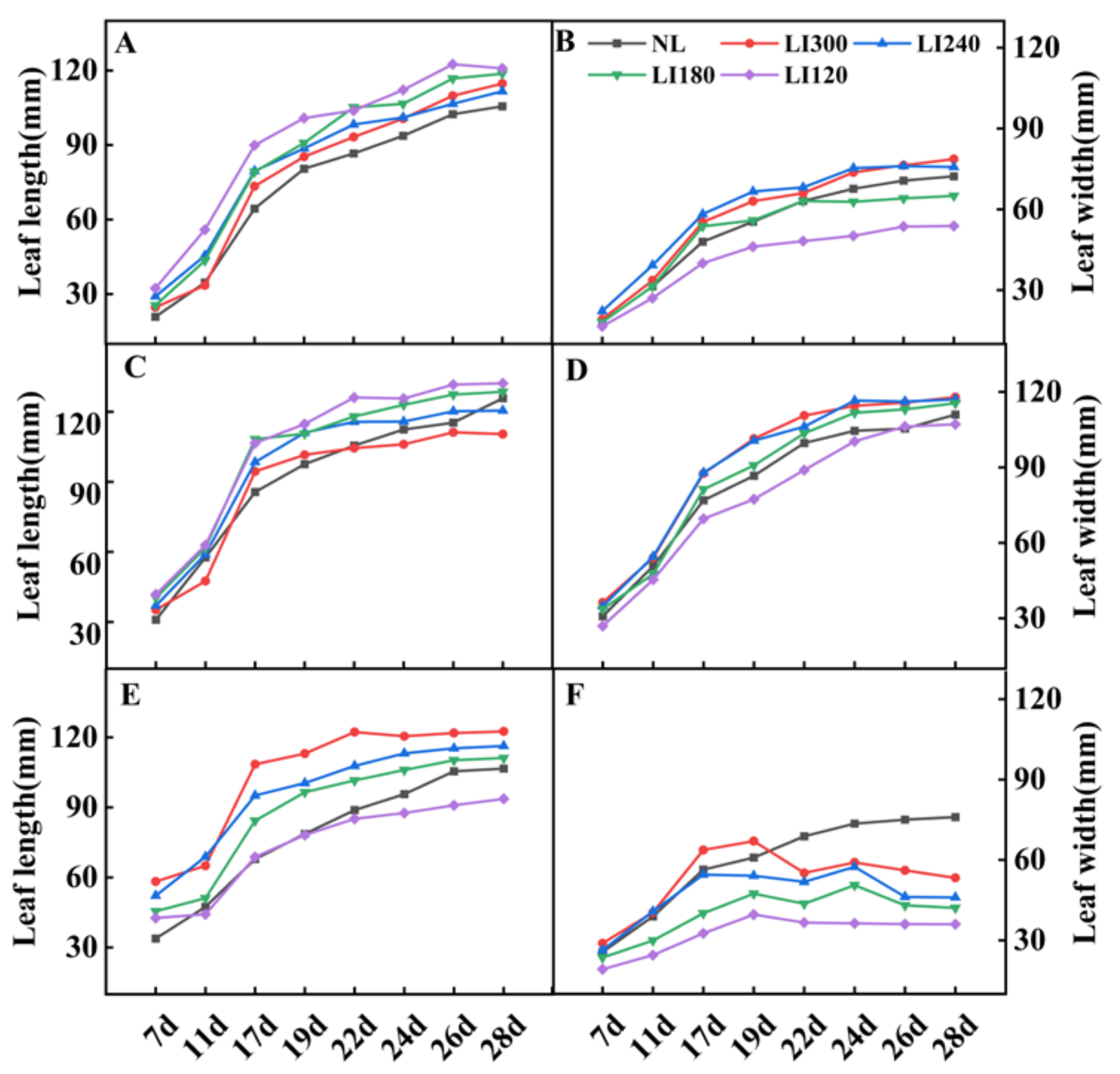
2.1. Plant Height, Plant Width, Number of Leaves, Leaf Length, Leaf Width, and Fresh Weight of Shoot and Root
2.2. Leaf Photosynthetic Net Rate and Chlorophyll Fluorescence
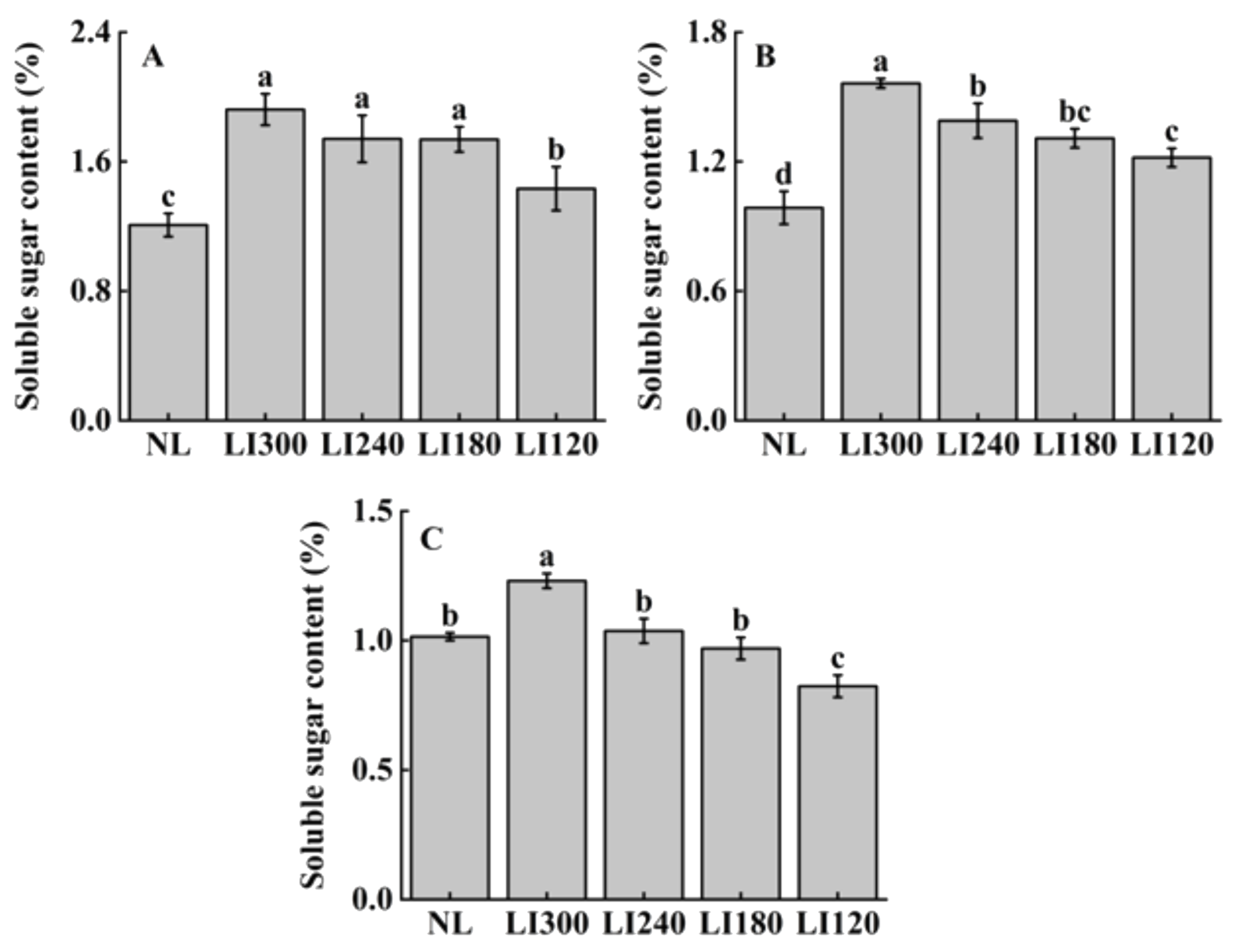
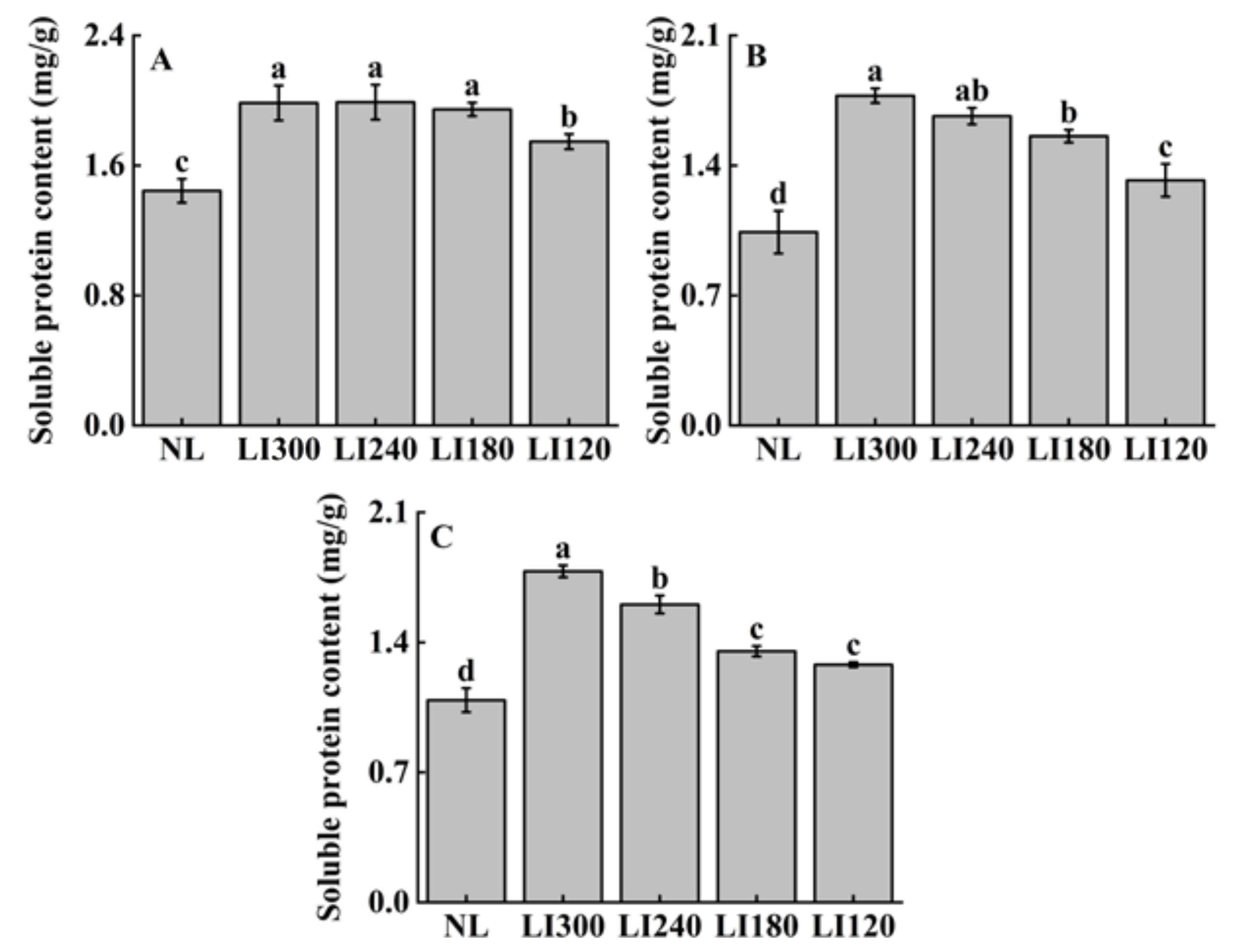
2.3. Soluble Sugar, Soluble Protein, and Ascorbic Acid Contents
3. Discussion
4. Materials and Methods
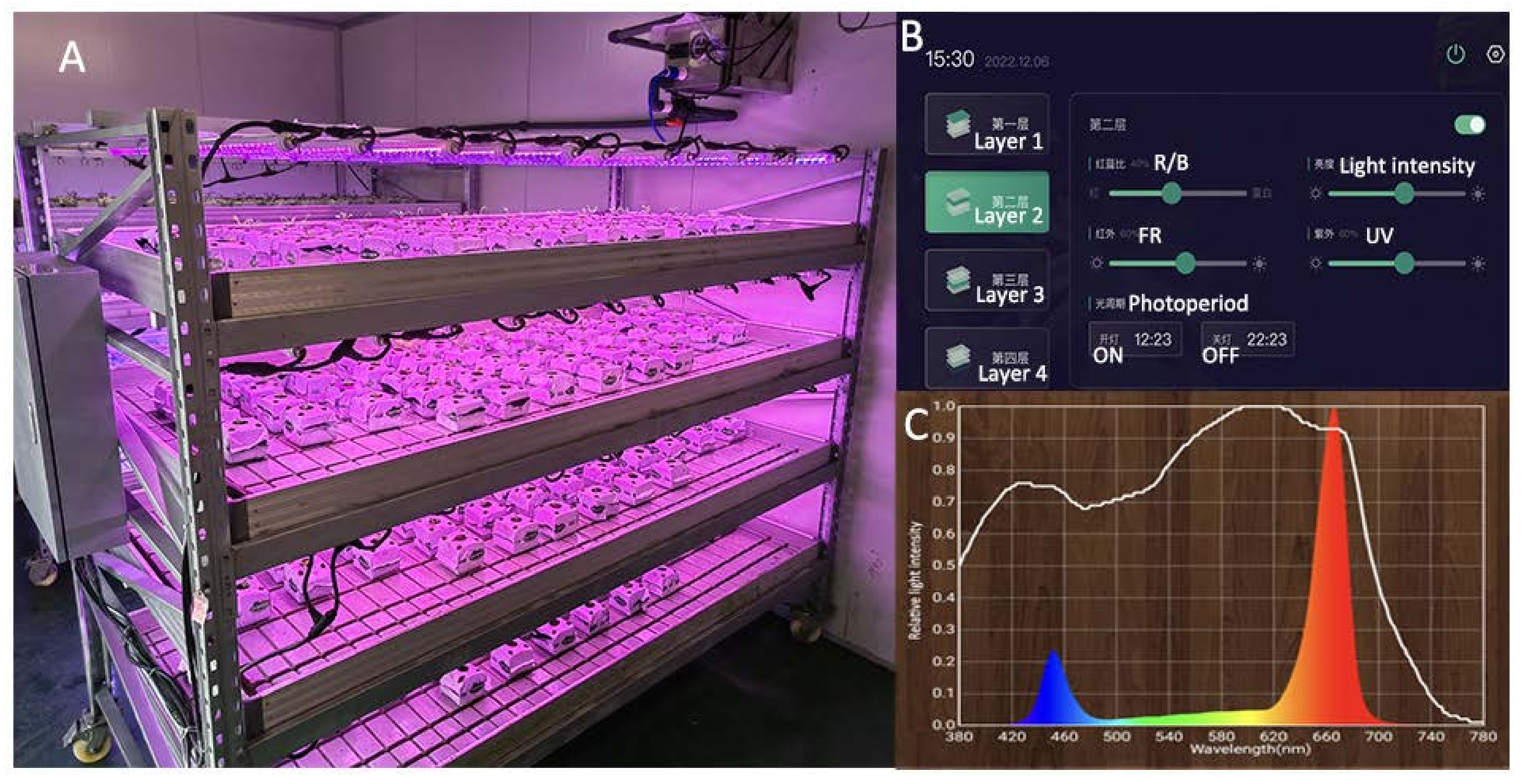
4.1. Materials and Treatment Conditions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Temperature (°C) (Day/Night) | Light Intensity (μmol m−2 s−1) | Photoperiod (Day/Night) |
|---|---|---|---|
| NL | 19.8/15.7 | 97 | 10 h/14 h |
| LI300 | 22 ± 2 | 300 | 16 h/8 h |
| LI240 | 240 | 16 h/8 h | |
| LI180 | 180 | 16 h/8 h | |
| LI120 | 120 | 16 h/8 h |
4.2. Sample Collection
4.3. Assessment of Various Growth Parameters
4.3.1. Plant Growth
4.3.2. Leaf Photosynthetic Rate and Chlorophyll Fluorescence
4.3.3. Soluble Sugar Content
4.3.4. Soluble Protein Content
4.3.5. Ascorbic Acid Content
4.4. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tewolde, F.T.; Lu, N.; Shiina, K.; Maruo, T.; Takagaki, M.; Kozai, T.; Yamori, W. Nighttime Supplemental LED Inter-Lighting Improves Growth and Yield of Single-Truss Tomatoes by Enhancing Photosynthesis in Both Winter and Summer. Front. Plant Sci. 2016, 7, 448. [Google Scholar] [CrossRef] [PubMed]
- Kozai, T. Sustainable Plant Factory: Closed Plant Production Systems with Artificial Light for High Resource Use Efficiencies and Quality Produce. Acta Hortic. 2013, 2012, 27–40. [Google Scholar] [CrossRef]
- Yamori, W.; Zhang, G. Feasibility Study of Rice Growth in Plant Factories. Rice Res. Open Access 2014, 2, 1–6. [Google Scholar] [CrossRef]
- Shimizu, H.; Saito, Y.; Nakashima, H.; Miyasaka, J.; Ohdoi, K. Light Environment Optimization for Lettuce Growth in Plant Factory. IFAC Proc. Vol. 2011, 44, 605–609. [Google Scholar] [CrossRef]
- Taulavuori, E.; Taulavuori, K.; Holopainen, J.K.; Julkunen-Tiitto, R.; Acar, C.; Dincer, I. Targeted Use of LEDs in Improvement of Production Efficiency through Phytochemical Enrichment. J. Sci. Food Agric. 2017, 97, 5059–5064. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Fu, Y.; Hu, D.; Yu, J.; Liu, H. Effect of Green, Yellow and Purple Radiation on Biomass, Photosynthesis, Morphology and Soluble Sugar Content of Leafy Lettuce via Spectral Wavebands “Knock Out”. Sci. Hortic. 2018, 236, 10–17. [Google Scholar] [CrossRef]
- Clouse, S.D. Integration of Light and Brassinosteroid Signals in Etiolated Seedling Growth. Trends Plant Sci. 2001, 6, 443–445. [Google Scholar] [CrossRef]
- Hamedalla, A.M.; Ali, M.M.; Ali, W.M.; Ahmed, M.A.A.; Kaseb, M.O.; Kalaji, H.M.; Gajc-Wolska, J.; Yousef, A.F. Increasing the Performance of Cucumber (Cucumis sativus L.) Seedlings by LED Illumination. Sci. Rep. 2022, 12, 852. [Google Scholar] [CrossRef]
- Zhang, T.; Shi, Y.; Piao, F.; Sun, Z. Effects of Different LED Sources on the Growth and Nitrogen Metabolism of Lettuce. Plant Cell Tissue Organ Cult. PCTOC 2018, 134, 231–240. [Google Scholar] [CrossRef]
- Chen, Y.; Li, T.; Yang, Q.; Zhang, Y.; Zou, J.; Bian, Z.; Wen, X. UVA Radiation Is Beneficial for Yield and Quality of Indoor Cultivated Lettuce. Front. Plant Sci. 2019, 10, 1563. [Google Scholar] [CrossRef]
- Zhang, X.; He, D.; Niu, G.; Yan, Z.; Song, J. Effects of Environment Lighting on the Growth, Photosynthesis, and Quality of Hydroponic Lettuce in a Plant Factory. Int. J. Agric. Biol. Eng. 2018, 11, 33–40. [Google Scholar] [CrossRef]
- Li, L.; Tong, Y.; Lu, J.; Li, Y.; Liu, X.; Cheng, R. Morphology, Photosynthetic Traits, and Nutritional Quality of Lettuce Plants as Affected by Green Light Substituting Proportion of Blue and Red Light. Front. Plant Sci. 2021, 12, 627311. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.J.; Bhandari, S.R.; Lee, G.; Lee, J.G. Optimization of Temperature and Light, and Cultivar Selection for the Production of High-Quality Head Lettuce in a Closed-Type Plant Factory. Hortic. Environ. Biotechnol. 2019, 60, 207–216. [Google Scholar] [CrossRef]
- Min, Q.; Marcelis, L.F.M.; Nicole, C.C.S.; Woltering, E.J. High Light Intensity Applied Shortly Before Harvest Improves Lettuce Nutritional Quality and Extends the Shelf Life. Front. Plant Sci. 2021, 12, 615355. [Google Scholar] [CrossRef]
- Hasperué, J.H.; Lemoine, L.; Vicente, A.R.; Chaves, A.R.; Martínez, G.A. Postharvest Senescence of Florets from Primary and Secondary Broccoli Inflorescences. Postharvest Biol. Technol. 2015, 104, 42–47. [Google Scholar] [CrossRef]
- Lin, K.H.; Huang, M.Y.; Huang, W.D.; Hsu, M.H.; Yang, Z.W.; Yang, C.M. The Effects of Red, Blue, and White Light-Emitting Diodes on the Growth, Development, and Edible Quality of Hydroponically Grown Lettuce (Lactuca sativa L. var. capitata). Sci. Hortic. 2013, 150, 86–91. [Google Scholar] [CrossRef]
- Ntagkas, N.; Woltering, E.J.; Marcelis, L.F.M. Light Regulates Ascorbate in Plants: An Integrated View on Physiology and Biochemistry. Environ. Exp. Bot. 2018, 147, 271–280. [Google Scholar] [CrossRef]
- Abreu, A.C.; Marín, P.; Aguilera-Sáez, L.M.; Tristán, A.I.; Peña, A.; Oliveira, I.; Simões, M.; Valera, D.; Fernández, I. Effect of a Shading Mesh on the Metabolic, Nutritional, and Defense Profiles of Harvested Greenhouse-Grown Organic Tomato Fruits and Leaves Revealed by NMR Metabolomics. J. Agric. Food Chem. 2019, 67, 12972–12985. [Google Scholar] [CrossRef]
- Yorio, N.C.; Goins, G.D.; Kagie, H.R.; Wheeler, R.M.; Sager, J.C. Improving Spinach, Radish, and Lettuce Growth under Red Light-Emitting Diodes (LEDs) with Blue Light Supplementation. HortScience 2001, 36, 380–383. [Google Scholar] [CrossRef]
- Weiguo, F.; Pingping, L.; Yanyou, W.; Jianjian, T. Effects of Different Light Intensities on Anti-Oxidative Enzyme Activity, Quality and Biomass in Lettuce. Hortic. Sci. 2012, 39, 129–134. [Google Scholar] [CrossRef]
- Zhan, L.; Hu, J.; Ai, Z.; Pang, L.; Li, Y.; Zhu, M. Light Exposure during Storage Preserving Soluble Sugar and L-Ascorbic Acid Content of Minimally Processed Romaine Lettuce (Lactuca sativa L. var. longifolia). Food Chem. 2013, 136, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, P.; Wang, J.; Fu, W. Growth, Photosynthesis, and Nutrient Uptake at Different Light Intensities and Temperatures in Lettuce. HortScience 2019, 54, 1925–1933. [Google Scholar] [CrossRef]
- Zhang, R.; Kong, Z.; Chen, S.; Ran, Z.; Ye, M.; Xu, J.; Zhou, C.; Liao, K.; Cao, J.; Yan, X. The Comparative Study for Physiological and Biochemical Mechanisms of Thalassiosira Pseudonana and Chaetoceros Calcitrans in Response to Different Light Intensities. Algal Res. 2017, 27, 89–98. [Google Scholar] [CrossRef]
- Ruban, A.V. Nonphotochemical Chlorophyll Fluorescence Quenching: Mechanism and Effectiveness in Protecting Plants from Photodamage. Plant Physiol. 2016, 170, 1903–1916. [Google Scholar] [CrossRef]
- Yudina, L.; Sukhova, E.; Gromova, E.; Mudrilov, M.; Zolin, Y.; Popova, A.; Nerush, V.; Pecherina, A.; Grishin, A.A.; Dorokhov, A.A.; et al. Effect of Duration of LED Lighting on Growth, Photosynthesis and Respiration in Lettuce. Plants 2023, 12, 442. [Google Scholar] [CrossRef]
- Eriksen, R.L.; Knepper, C.; Cahn, M.D.; Mou, B. Screening of Lettuce Germplasm for Agronomic Traits under Low Water Conditions. HortScience 2016, 51, 669–679. [Google Scholar] [CrossRef]
- Cammarisano, L.; Körner, O. Response of Cyanic and Acyanic Lettuce Cultivars to an Increased Proportion of Blue Light. Biology 2022, 11, 959. [Google Scholar] [CrossRef]
- Cammarisano, L.; Donnison, I.S.; Robson, P.R.H. The Effect of Red & Blue Rich LEDs vs Fluorescent Light on Lollo Rosso Lettuce Morphology and Physiology. Front. Plant Sci. 2021, 12, 603411. [Google Scholar] [CrossRef]
- Landi, M.; Zivcak, M.; Sytar, O.; Brestic, M.; Allakhverdiev, S.I. Plasticity of Photosynthetic Processes and the Accumulation of Secondary Metabolites in Plants in Response to Monochromatic Light Environments: A Review. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148131. [Google Scholar] [CrossRef]
- Sago, Y. Effects of Light Intensity and Growth Rate on Tipburn Development and Leaf Calcium Concentration in Butterhead Lettuce. HortScience 2016, 51, 1087–1091. [Google Scholar] [CrossRef]
- Kang, J.; Sugumaran, K.; Atulba, S.L.; Jeong, B.R.; Hwang, S. Light Intensity and Photoperiod Influence the Growth and Development of Hydroponically Grown Leaf Lettuce in a Closed-Type Plant Factory System. Horticulture 2013, 54, 501–509. [Google Scholar] [CrossRef]
- Miyake, C. Alternative Electron Flows (Water–Water Cycle and Cyclic Electron Flow Around PSI) in Photosynthesis: Molecular Mechanisms and Physiological Functions. Plant Cell Physiol. 2010, 51, 1951–1963. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.G.; Choi, C.S.; Jang, Y.A.; Jang, S.W.; Lee, S.G.; Um, Y.C. Effects of Air Temperature and Air Flow Rate Control on the Tipburn Occurrence of Leaf Lettuce in a Closed-Type Plant Factory System. Hortic. Environ. Biotechnol. 2013, 54, 303–310. [Google Scholar] [CrossRef]
- Frantz, J.M.; Ritchie, G.; Cometti, N.N.; Robinson, J.; Bugbee, B. Exploring the Limits of Crop Productivity: Beyond the Limits of Tipburn in Lettuce. J. Am. Soc. Hortic. Sci. 2004, 129, 331–338. [Google Scholar] [CrossRef]
- Hamidon, M.H.; Ahamed, T. Detection of Tip-Burn Stress on Lettuce Grown in an Indoor Environment Using Deep Learning Algorithms. Sensors 2022, 22, 7251. [Google Scholar] [CrossRef]
- Kozai, T. Resource Use Efficiency of Closed Plant Production System with Artificial Light: Concept, Estimation and Application to Plant Factory. Proc. Jpn. Acad. Ser. B 2013, 89, 447–461. [Google Scholar] [CrossRef] [PubMed]
- Ryder, E.J.; Waycott, W. Crisphead Lettuce Resistant to Tipburn: Cultivar Tiber and Eight Breeding Lines. HortScience 1998, 33, 903–904. [Google Scholar] [CrossRef]
- Jang, S.W.; Park, S.H.; Lee, J.N.; Seo, M.H.; Kim, D.G.; Lee, M.J.; Jeong, T.G.; In, M.S.; Lee, J.W.; Kim, U.J.; et al. Breeding of Lettuce ‘Sambokhacheong’ Tolerant to Tipburn and with Good Yield. Korean Soc. Breed. Sci. 2020, 52, 200–205. [Google Scholar] [CrossRef]
- Uno, Y.; Okubo, H.; Itoh, H.; Koyama, R. Reduction of Leaf Lettuce Tipburn Using an Indicator Cultivar. Sci. Hortic. 2016, 210, 14–18. [Google Scholar] [CrossRef]
- Hayashi, E.; Kozai, T. Phenotyping- and AI-Based Environmental Control and Breeding for PFAL. In Smart Plant Factory: The Next Generation Indoor Vertical Farms; Kozai, T., Ed.; Springer: Singapore, 2018; pp. 405–411. ISBN 9789811310652. [Google Scholar]
- He, D.; Yan, Z.; Sun, X.; Yang, P. Leaf Development and Energy Yield of Hydroponic Sweetpotato Seedlings Using Single-Node Cutting as Influenced by Light Intensity and LED Spectrum. J. Plant Physiol. 2020, 254, 153274. [Google Scholar] [CrossRef]
- Camejo, D.; Frutos, A.; Mestre, T.C.; del Carmen Piñero, M.; Rivero, R.M.; Martínez, V. Artificial Light Impacts the Physical and Nutritional Quality of Lettuce Plants. Hortic. Environ. Biotechnol. 2020, 61, 69–82. [Google Scholar] [CrossRef]
- Shafiq, I.; Hussain, S.; Raza, M.A.; Iqbal, N.; Asghar, M.A.; Raza, A.; Fan, Y.; Mumtaz, M.; Shoaib, M.; Ansar, M.; et al. Crop Photosynthetic Response to Light Quality and Light Intensity. J. Integr. Agric. 2021, 20, 4–23. [Google Scholar] [CrossRef]
- Burattini, C.; Mattoni, B.; Bisegna, F. The Impact of Spectral Composition of White LEDs on Spinach (Spinacia oleracea) Growth and Development. Energies 2017, 10, 1383. [Google Scholar] [CrossRef]
- Fan, X.; Xu, Z.; Liu, X.; Tang, C.; Wang, L.; Han, X. Effects of Light Intensity on the Growth and Leaf Development of Young Tomato Plants Grown under a Combination of Red and Blue Light. Sci. Hortic. 2013, 153, 50–55. [Google Scholar] [CrossRef]
- Fu, Y.; Li, H.; Yu, J.; Liu, H.; Cao, Z.; Manukovsky, N.S.; Liu, H. Interaction Effects of Light Intensity and Nitrogen Concentration on Growth, Photosynthetic Characteristics and Quality of Lettuce (Lactuca sativa L. var. youmaicai). Sci. Hortic. 2017, 214, 51–57. [Google Scholar] [CrossRef]
- Yu, A.; Xie, Y.; Pan, X.; Zhang, H.; Cao, P.; Su, X.; Chang, W.; Li, M. Photosynthetic Phosphoribulokinase Structures: Enzymatic Mechanisms and the Redox Regulation of the Calvin-Benson-Bassham Cycle [OPEN]. Plant Cell 2020, 32, 1556–1573. [Google Scholar] [CrossRef]
- Sekhar, S.; Panda, D.; Kumar, J.; Mohanty, N.; Biswal, M.; Baig, M.J.; Kumar, A.; Umakanta, N.; Samantaray, S.; Pradhan, S.K.; et al. Comparative Transcriptome Profiling of Low Light Tolerant and Sensitive Rice Varieties Induced by Low Light Stress at Active Tillering Stage. Sci. Rep. 2019, 9, 5753. [Google Scholar] [CrossRef]
- Sui, X.; Mao, S.; Wang, L.; Zhang, B.; Zhang, Z. Effect of Low Light on the Characteristics of Photosynthesis and Chlorophyll a Fluorescence During Leaf Development of Sweet Pepper. J. Integr. Agric. 2012, 11, 1633–1643. [Google Scholar] [CrossRef]
- Lu, C.; Lu, Q.; Zhang, J.; Zhang, Q.; Kuang, T. Xanthophyll Cycle, Light Energy Dissipation and Photosystem II down-Regulation in Senescent Leaves of Wheat Plants Grown in the Field. Funct. Plant Biol. 2001, 28, 1023–1030. [Google Scholar] [CrossRef]
- Qin, L.; Guo, S.; Ai, W.; Tang, Y. Selection of Candidate Salad Vegetables for Controlled Ecological Life Support System. Adv. Space Res. 2008, 41, 768–772. [Google Scholar] [CrossRef]
- Wang, L.; Deng, F.; Ren, W.-J.; Yang, W.-Y. Effects of Shading on Starch Pasting Characteristics of Indica Hybrid Rice (Oryza sativa L.). PLoS ONE 2013, 8, e68220. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Feng, L.; Liu, Q.; Wu, X.; Fan, Y.; Raza, M.A.; Cheng, Y.; Chen, J.; Wang, X.; Yong, T.; et al. Effect of Interactions between Light Intensity and Red-to- Far-Red Ratio on the Photosynthesis of Soybean Leaves under Shade Condition. Environ. Exp. Bot. 2018, 150, 79–87. [Google Scholar] [CrossRef]
- Wherley, B.G.; Gardner, D.S.; Metzger, J.D. Tall Fescue Photomorphogenesis as Influenced by Changes in the Spectral Composition and Light Intensity. Crop Sci. 2005, 45, 562–568. [Google Scholar] [CrossRef]
- Terashima, I.; Hanba, Y.T.; Tazoe, Y.; Vyas, P.; Yano, S. Irradiance and Phenotype: Comparative Eco-Development of Sun and Shade Leaves in Relation to Photosynthetic CO2 Diffusion. J. Exp. Bot. 2006, 57, 343–354. [Google Scholar] [CrossRef]
- Kalve, S.; Fotschki, J.; Beeckman, T.; Vissenberg, K.; Beemster, G.T.S. Three-Dimensional Patterns of Cell Division and Expansion throughout the Development of Arabidopsis Thaliana Leaves. J. Exp. Bot. 2014, 65, 6385–6397. [Google Scholar] [CrossRef]
- Terashima, I.; Miyazawa, S.-I.; Hanba, Y.T. Why Are Sun Leaves Thicker than Shade Leaves?—Consideration Based on Analyses of CO2 Diffusion in the Leaf. J. Plant Res. 2001, 114, 93–105. [Google Scholar] [CrossRef]
- Sage, R.F.; Kubien, D.S. The Temperature Response of C3 and C4 Photosynthesis. Plant Cell Environ. 2007, 30, 1086–1106. [Google Scholar] [CrossRef]
- Moore, C.E.; Meacham-Hensold, K.; Lemonnier, P.; Slattery, R.A.; Benjamin, C.; Bernacchi, C.J.; Lawson, T.; Cavanagh, A.P. The Effect of Increasing Temperature on Crop Photosynthesis: From Enzymes to Ecosystems. J. Exp. Bot. 2021, 72, 2822–2844. [Google Scholar] [CrossRef]
- Adams, S.R.; Langton, F.A. Photoperiod and Plant Growth: A Review. J. Hortic. Sci. Biotechnol. 2005, 80, 2–10. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; De Pascale, S.; Colla, G. Improving Vegetable Quality in Controlled Environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- Fondy, B.R.; Geiger, D.R.; Servaites, J.C. Photosynthesis, Carbohydrate Metabolism, and Export in Beta vulgaris L. and Phaseolus vulgaris L. during Square and Sinusoidal Light Regimes 1. Plant Physiol. 1989, 89, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.L.; Liu, W.K.; Yang, Q.C. Quality Changes in Hydroponic Lettuce Grown under Pre-Harvest Short-Duration Continuous Light of Different Intensities. J. Hortic. Sci. Biotechnol. 2012, 87, 429–434. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, Y.; Zhao, X.; Yu, H.-J.; Shi, K.; Yu, J.-Q. Impact of Light Variation on Development of Photoprotection, Antioxidants, and Nutritional Value in Lactuca sativa L. J. Agric. Food Chem. 2009, 57, 5494–5500. [Google Scholar] [CrossRef] [PubMed]
- Viršilė, A.; Brazaitytė, A.; Vaštakaitė-Kairienė, V.; Miliauskienė, J.; Jankauskienė, J.; Novičkovas, A.; Laužikė, K.; Samuolienė, G. The Distinct Impact of Multi-Color LED Light on Nitrate, Amino Acid, Soluble Sugar and Organic Acid Contents in Red and Green Leaf Lettuce Cultivated in Controlled Environment. Food Chem. 2020, 310, 125799. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Jiang, W.; Zhao, Y. Experiment Guidance of Postharvest Physiology and Biochemistry of Fruits and Vegetables (Chinese Edition); China Light Industry Press: Beijing, China, 2017; ISBN 978-7-5019-6003-3. [Google Scholar]





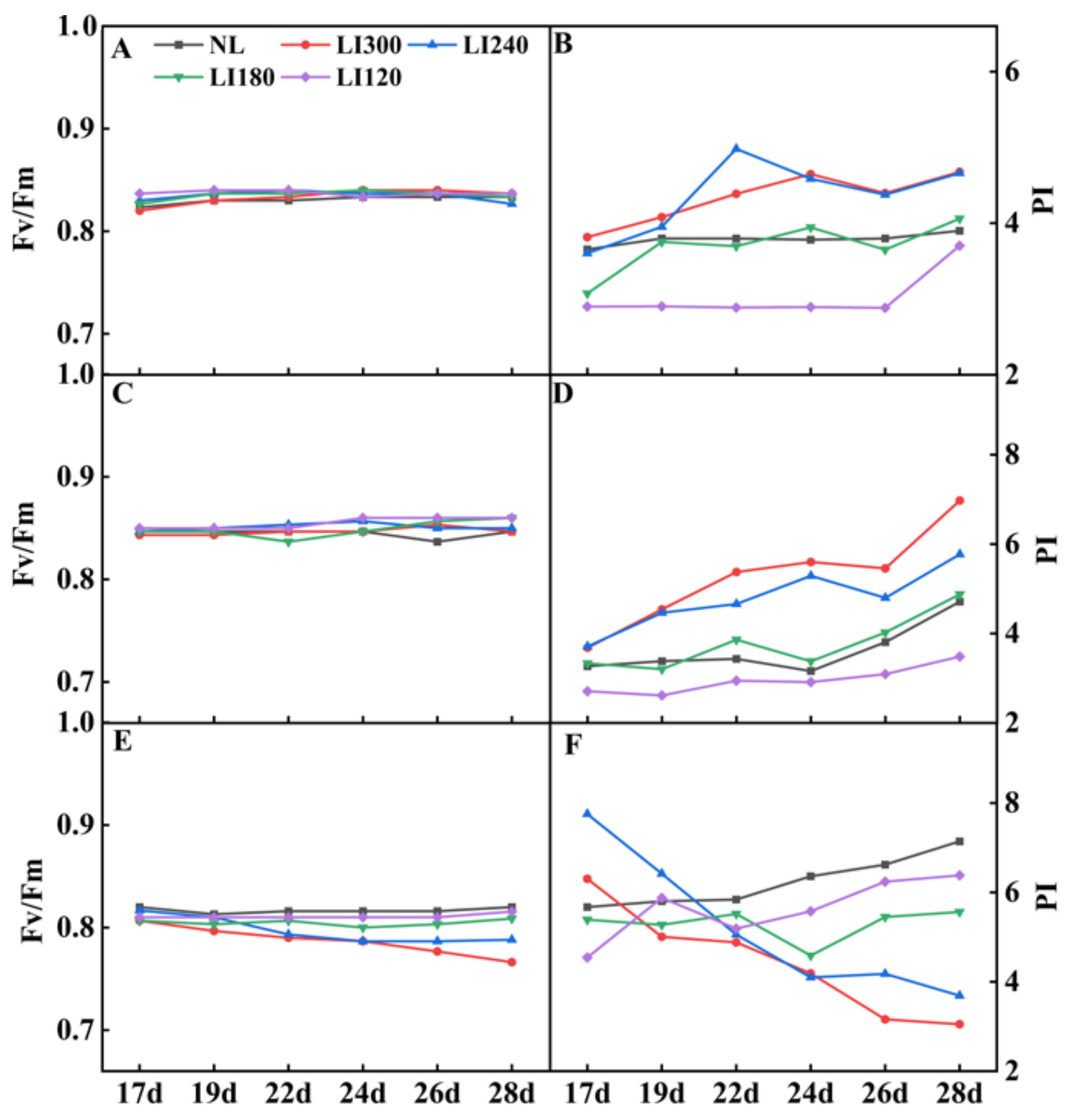
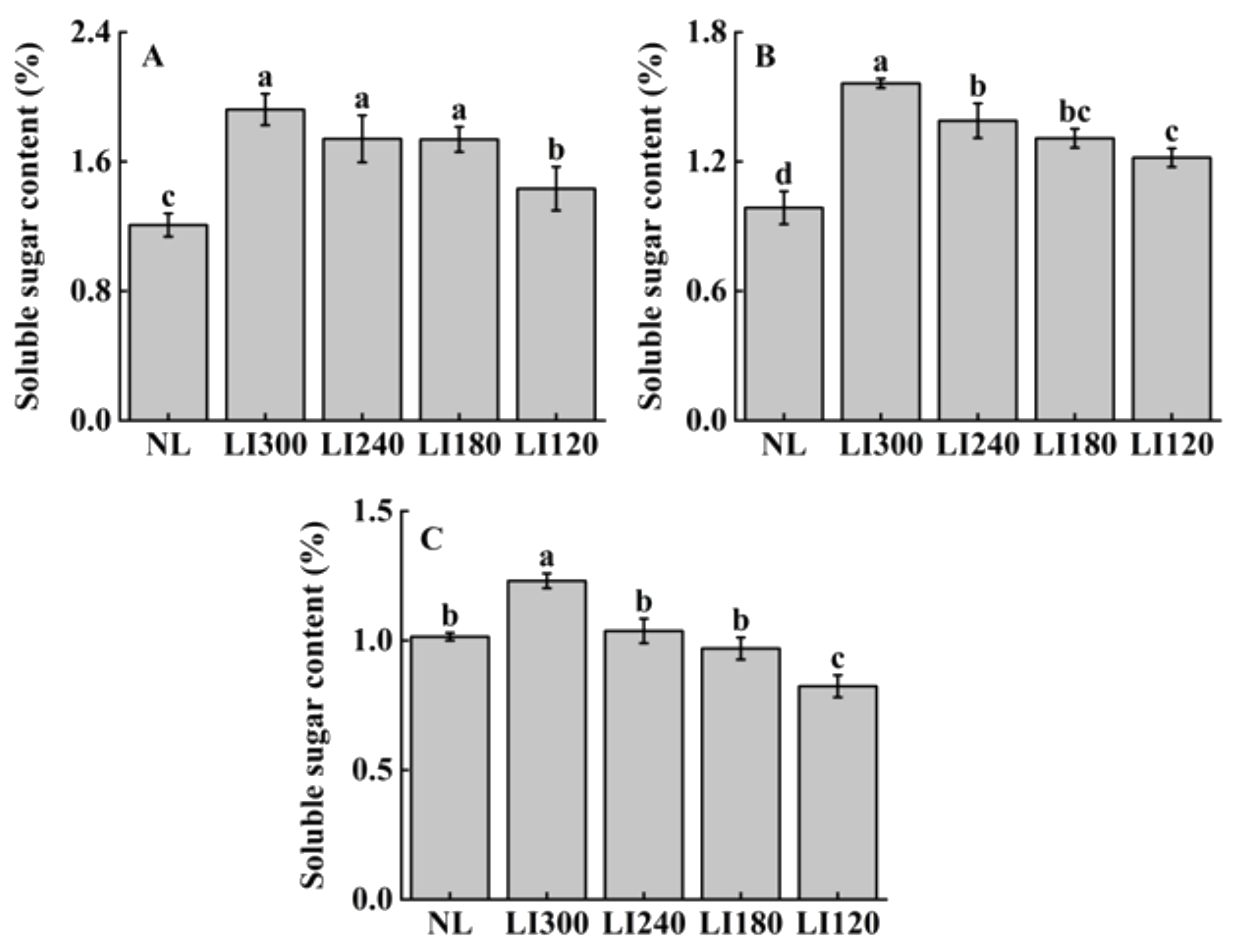
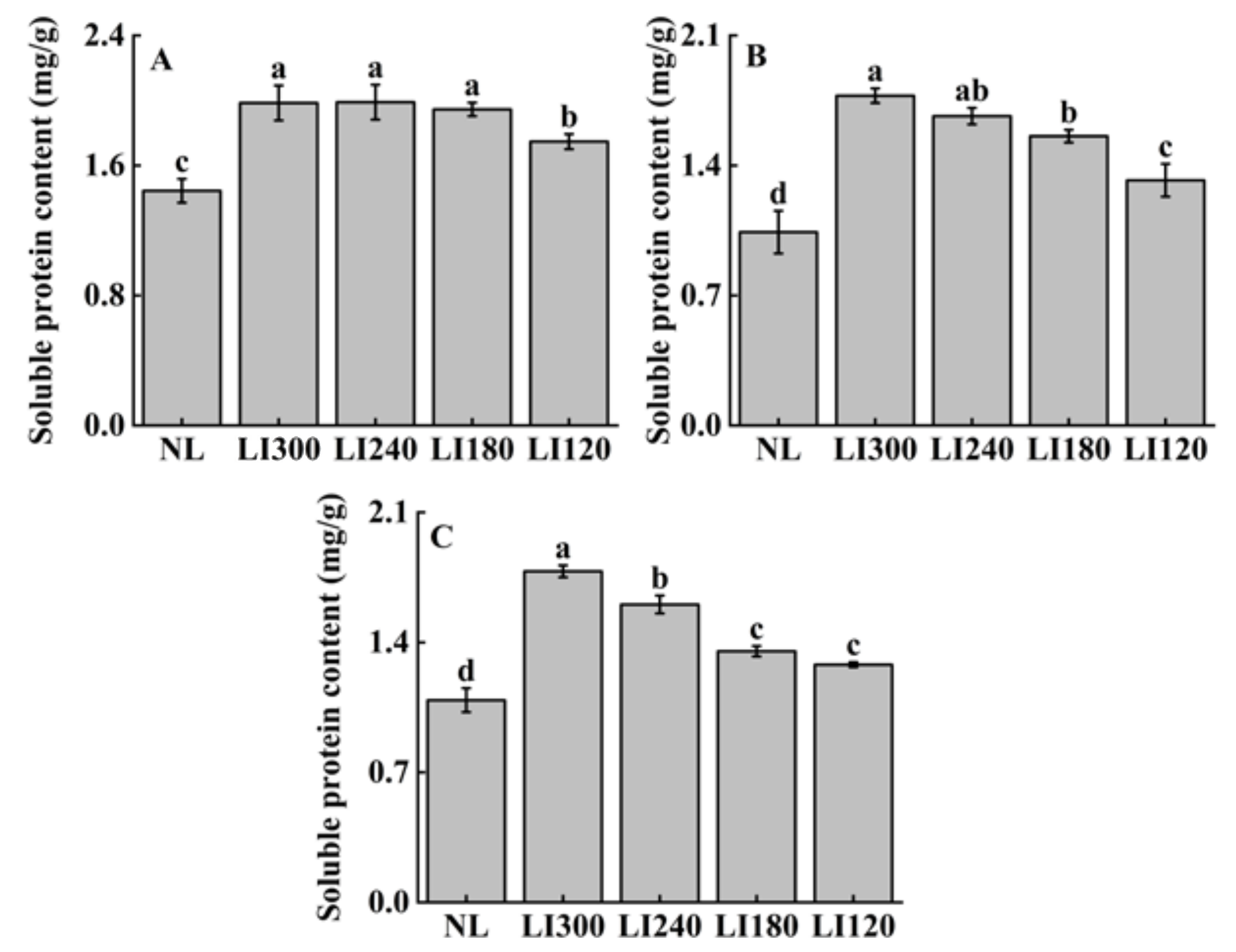


| Treatments | Pn (µmol CO2·m−2·s−1) | Ci (µmol CO2·mol−1) | Gs (mmol H2O·m−2·s−1) | Tr (mmol H2O·m−2·s−1) | WUE (mmol CO2·mol−1·H2O) | |
|---|---|---|---|---|---|---|
| Crunchy | NL | 7.8 ± 0.9 d | 144.7 ± 6.7 b | 47.0 ± 4.0 e | 0.9 ± 0.1 c | 8.7 ± 0.2 a |
| LI300 | 21.3 ± 0.4 a | 356.7 ± 2.5 a | 436.3 ± 20.1 a | 3.6 ± 0.1 a | 5.9 ± 0 b | |
| LI240 | 18.4 ± 0.4 b | 363.3 ± 4.2 a | 353.3 ± 24.5 b | 3.3 ± 0.1 a | 5.6 ± 0.1 b | |
| LI180 | 11.5 ± 0.1 c | 340 ± 14.1 a | 144.3 ± 18.2 c | 2.0 ± 0.2 b | 5.9 ± 0.5 b | |
| LI120 | 7.8 ± 1.0 d | 345 ± 33.8 a | 99.0 ± 37.3 d | 1.5 ± 0.6 b | 5.4 ± 1.3 b | |
| Deangelia | NL | 7.3 ± 1.5 d | 167.0 ± 3.0 c | 47.0 ± 8.9 d | 0.8 ± 0.1 c | 9.1 ± 0.2 a |
| LI300 | 23.0 ± 2.6 a | 342.7 ± 19.9 b | 395.3 ± 18.6 a | 3.8 ± 0.6 b | 6.1 ± 0.6 b | |
| LI240 | 20.5 ± 0.6 ab | 338.3 ± 47.4 b | 375.0 ± 9.8 ab | 4.0 ± 1.1 b | 5.3 ± 1.4 bc | |
| LI180 | 18.6 ± 0.6 b | 407.0 ± 1.0 a | 350.0 ± 29.8 b | 5.3 ± 0.2 a | 3.5 ± 0 d | |
| LI120 | 12.9 ± 1.2 c | 376.0 ± 2.6 ab | 301.3 ± 29.8 c | 3.0 ± 0.1 b | 4.3 ± 0.2 cd | |
| Shawen | NL | 10.0 ± 1.0 b | 124.7 ± 4.0 c | 50.0 ± 11.5 d | 0.79 ± 0.2 c | 11.5 ± 0.2 a |
| LI300 | 13.6 ± 1.1 a | 420.3 ± 9.5 ab | 387.3 ± 21.5 a | 5.3 ± 0.1 a | 2.6 ± 0.2 c | |
| LI240 | 14.7 ± 1.9 a | 416.3 ± 7.0 ab | 345.3 ± 25.5 ab | 5.4 ± 0.1 a | 2.7 ± 0.3 c | |
| LI180 | 16.0 ± 0.7 a | 441.7 ± 7.1 a | 331.7 ± 19.8 b | 5.5 ± 0.2 a | 2.9 ± 0.1 bc | |
| LI120 | 10.1 ± 1.3 b | 394.3 ± 34.1 b | 243.3 ± 34.6 c | 2.8 ± 0.7 b | 3.8 ± 1.2 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miao, C.; Yang, S.; Xu, J.; Wang, H.; Zhang, Y.; Cui, J.; Zhang, H.; Jin, H.; Lu, P.; He, L.; et al. Effects of Light Intensity on Growth and Quality of Lettuce and Spinach Cultivars in a Plant Factory. Plants 2023, 12, 3337. https://doi.org/10.3390/plants12183337
Miao C, Yang S, Xu J, Wang H, Zhang Y, Cui J, Zhang H, Jin H, Lu P, He L, et al. Effects of Light Intensity on Growth and Quality of Lettuce and Spinach Cultivars in a Plant Factory. Plants. 2023; 12(18):3337. https://doi.org/10.3390/plants12183337
Chicago/Turabian StyleMiao, Chen, Shaojun Yang, Jing Xu, Hong Wang, Yongxue Zhang, Jiawei Cui, Hongmei Zhang, Haijun Jin, Panling Lu, Lizhong He, and et al. 2023. "Effects of Light Intensity on Growth and Quality of Lettuce and Spinach Cultivars in a Plant Factory" Plants 12, no. 18: 3337. https://doi.org/10.3390/plants12183337
APA StyleMiao, C., Yang, S., Xu, J., Wang, H., Zhang, Y., Cui, J., Zhang, H., Jin, H., Lu, P., He, L., Yu, J., Zhou, Q., & Ding, X. (2023). Effects of Light Intensity on Growth and Quality of Lettuce and Spinach Cultivars in a Plant Factory. Plants, 12(18), 3337. https://doi.org/10.3390/plants12183337