

The Impact of 2-Ketones Released from Solid Lipid Nanoparticles on Growth Modulation and Antioxidant System of Lactuca sativa

 , ,
, ,  , , , ,
, , , ,  ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Results
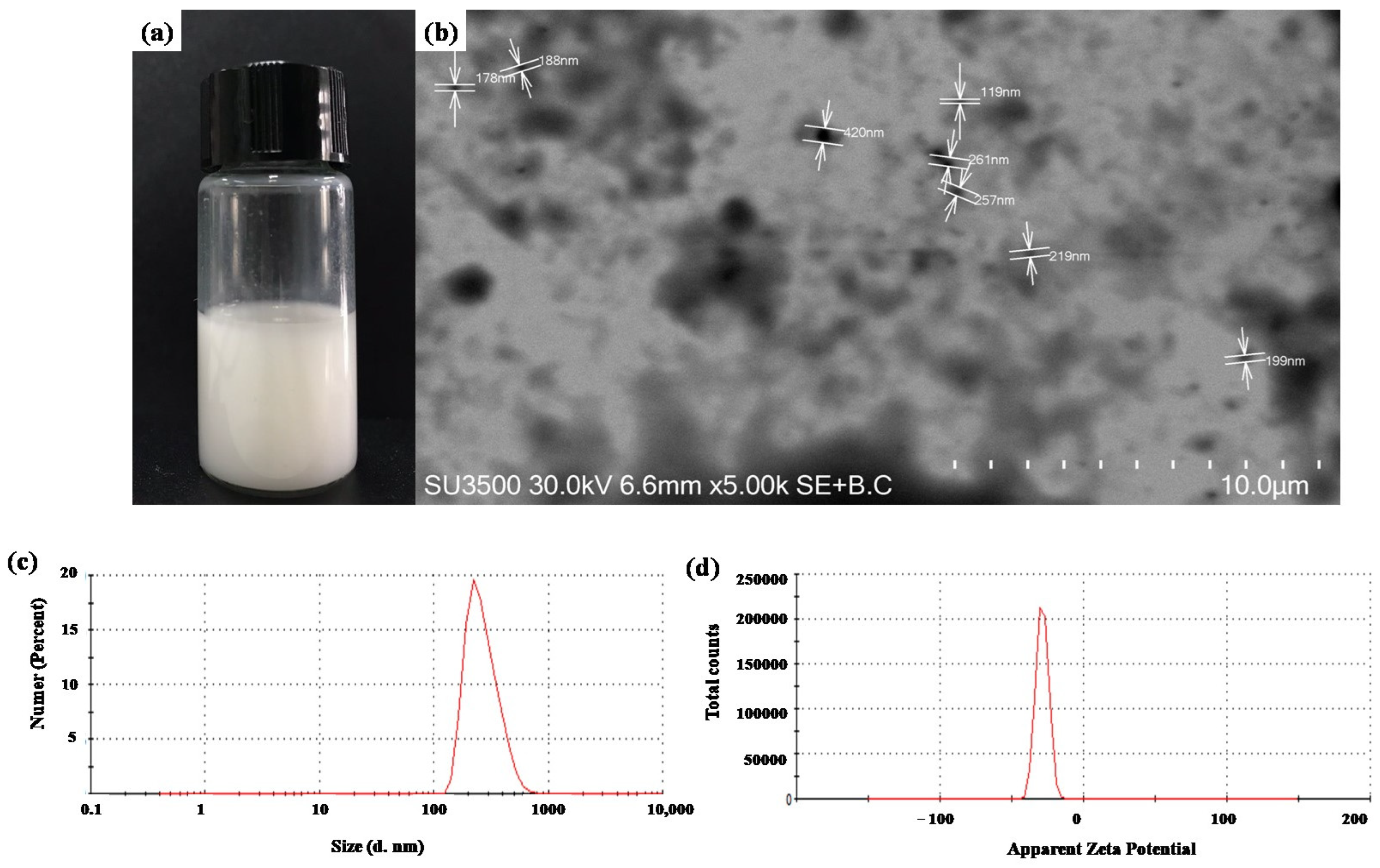
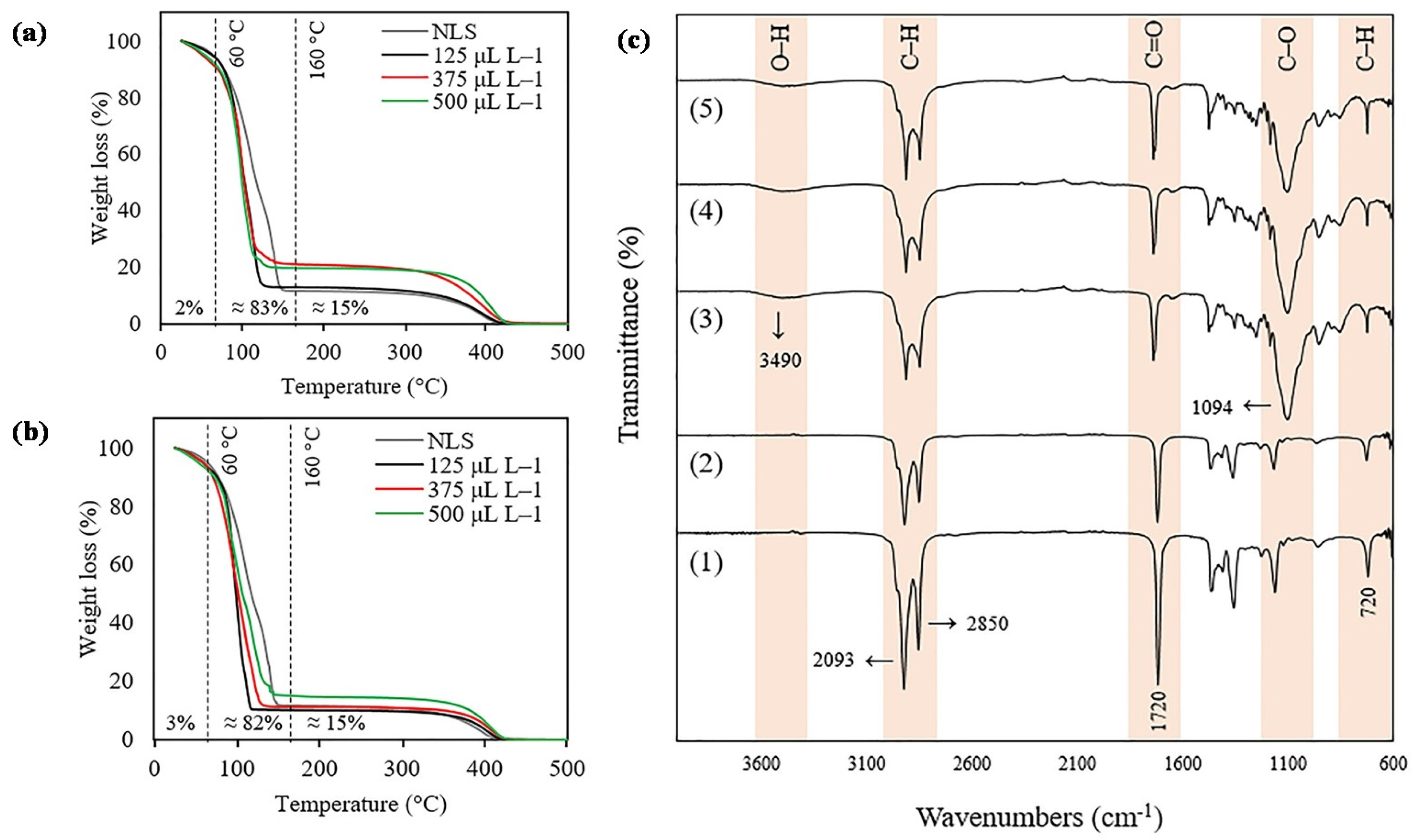
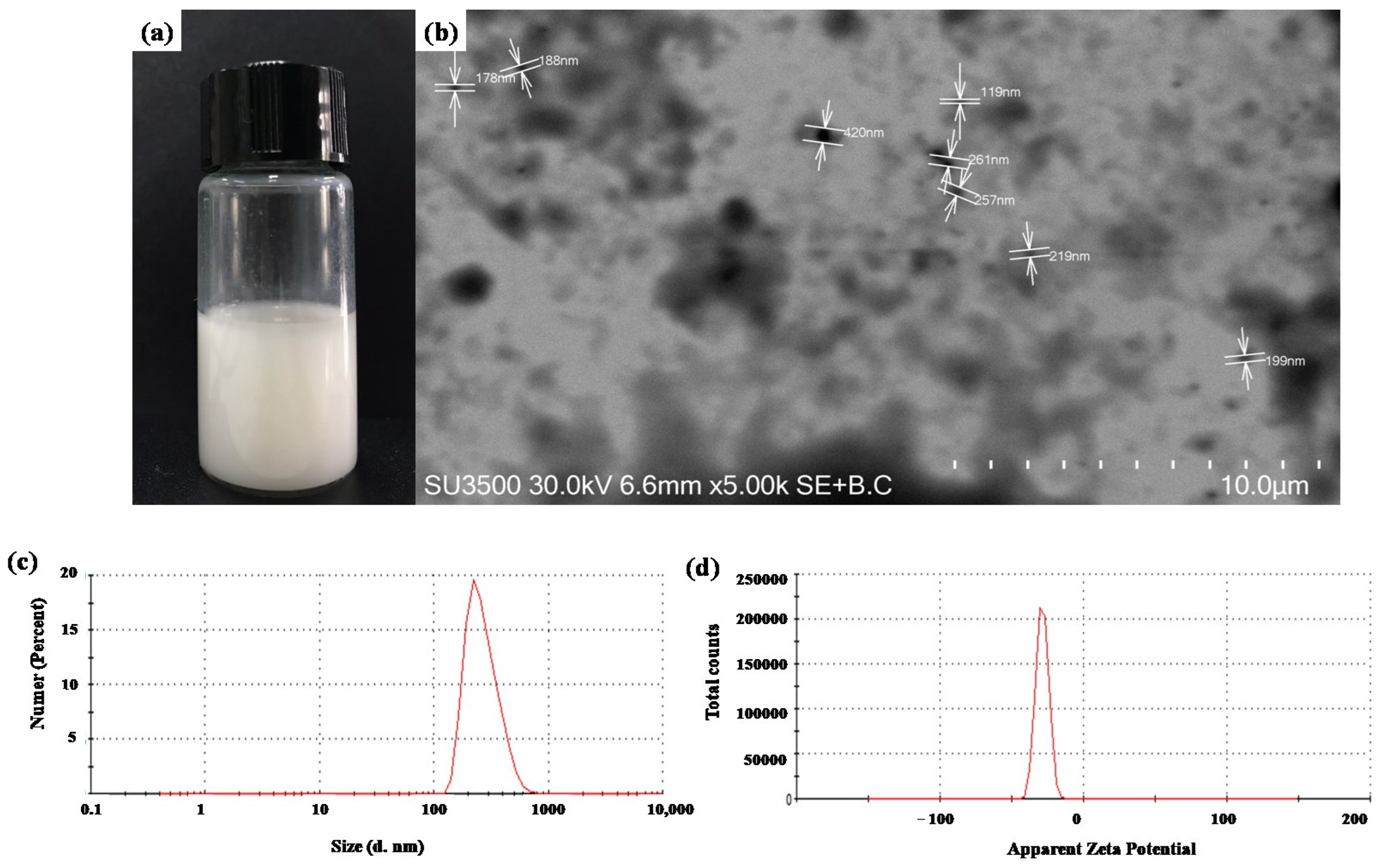
2.1. SLN Characterization
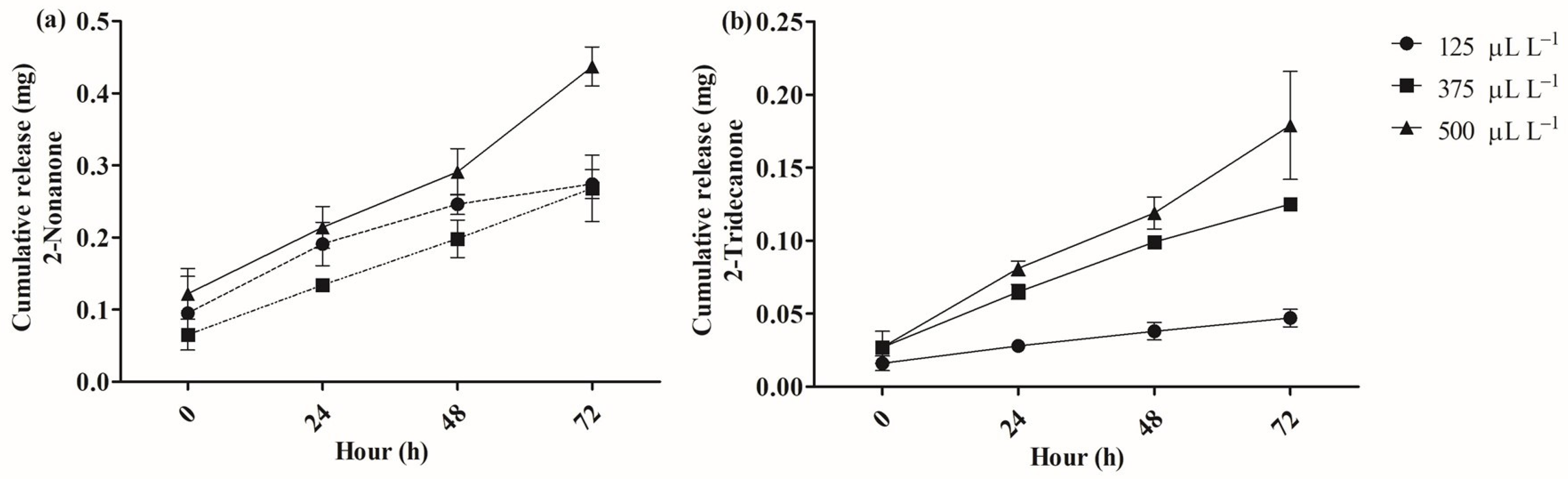
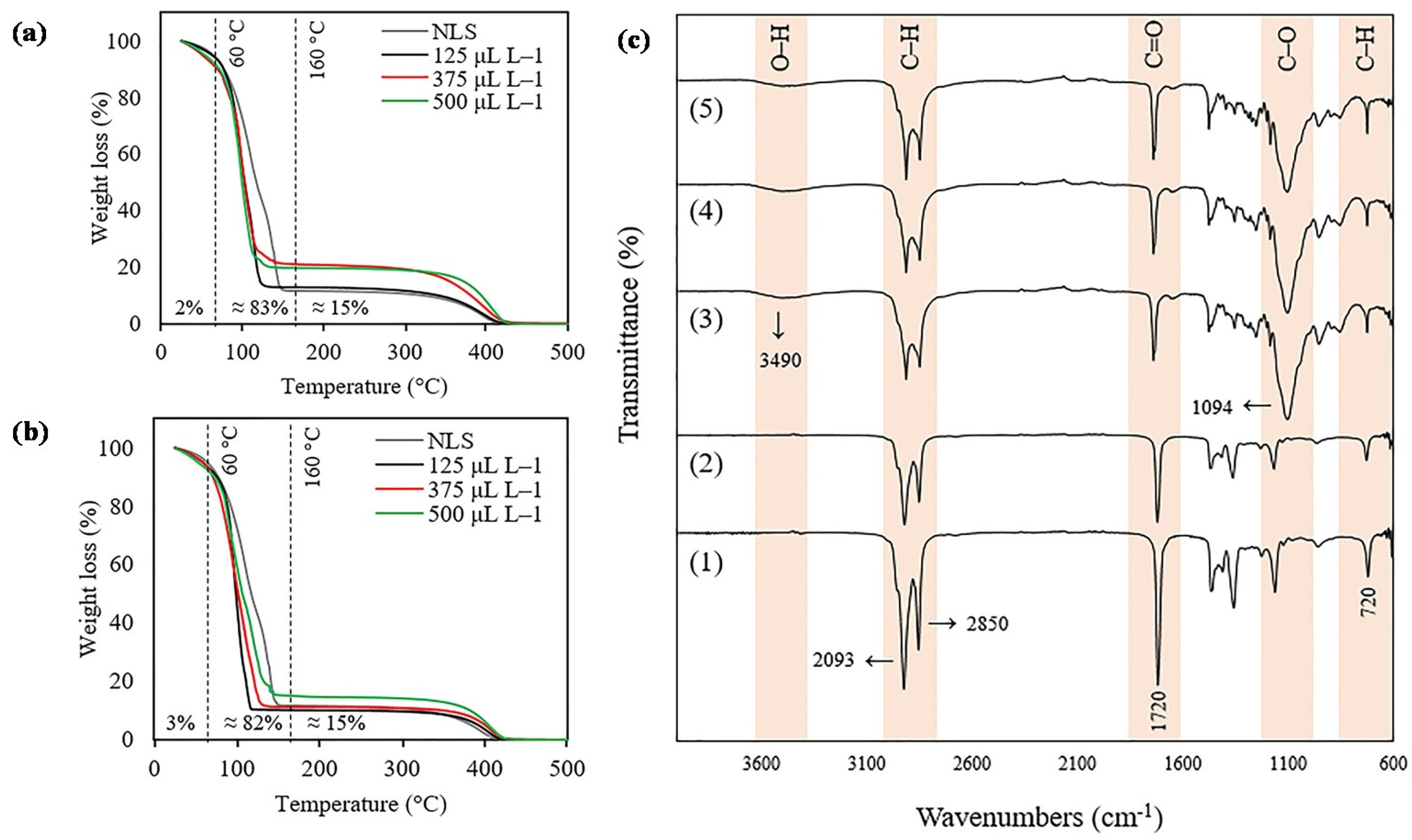
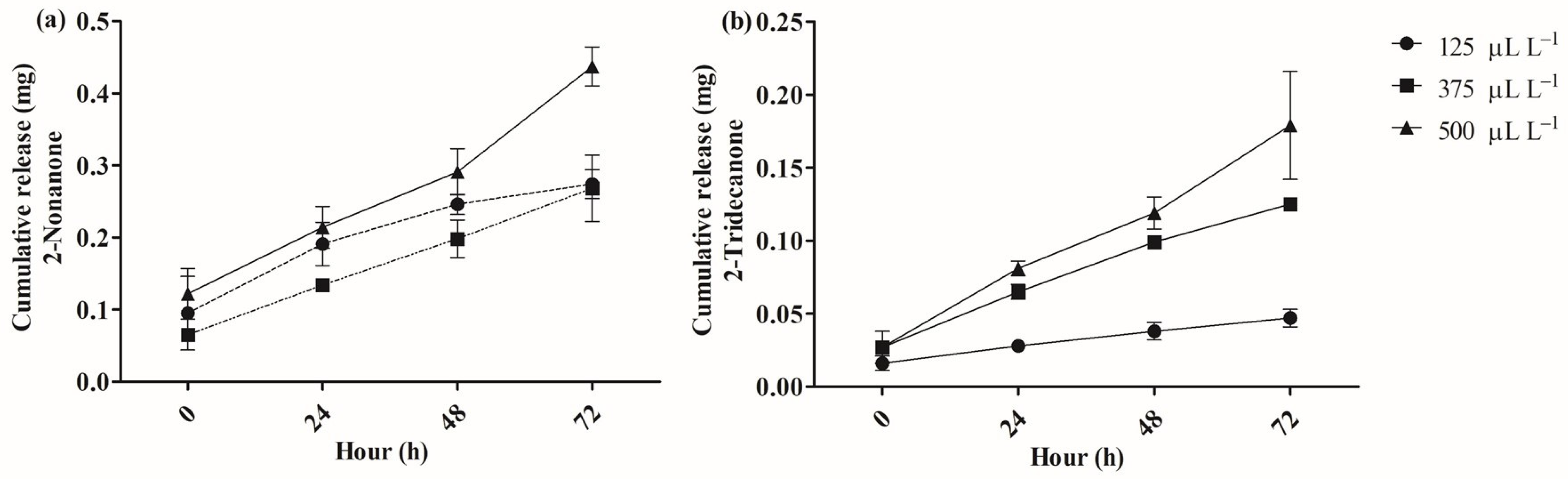
2.2. Release of 2-Nonanone and 2-Tridecanone from SLN
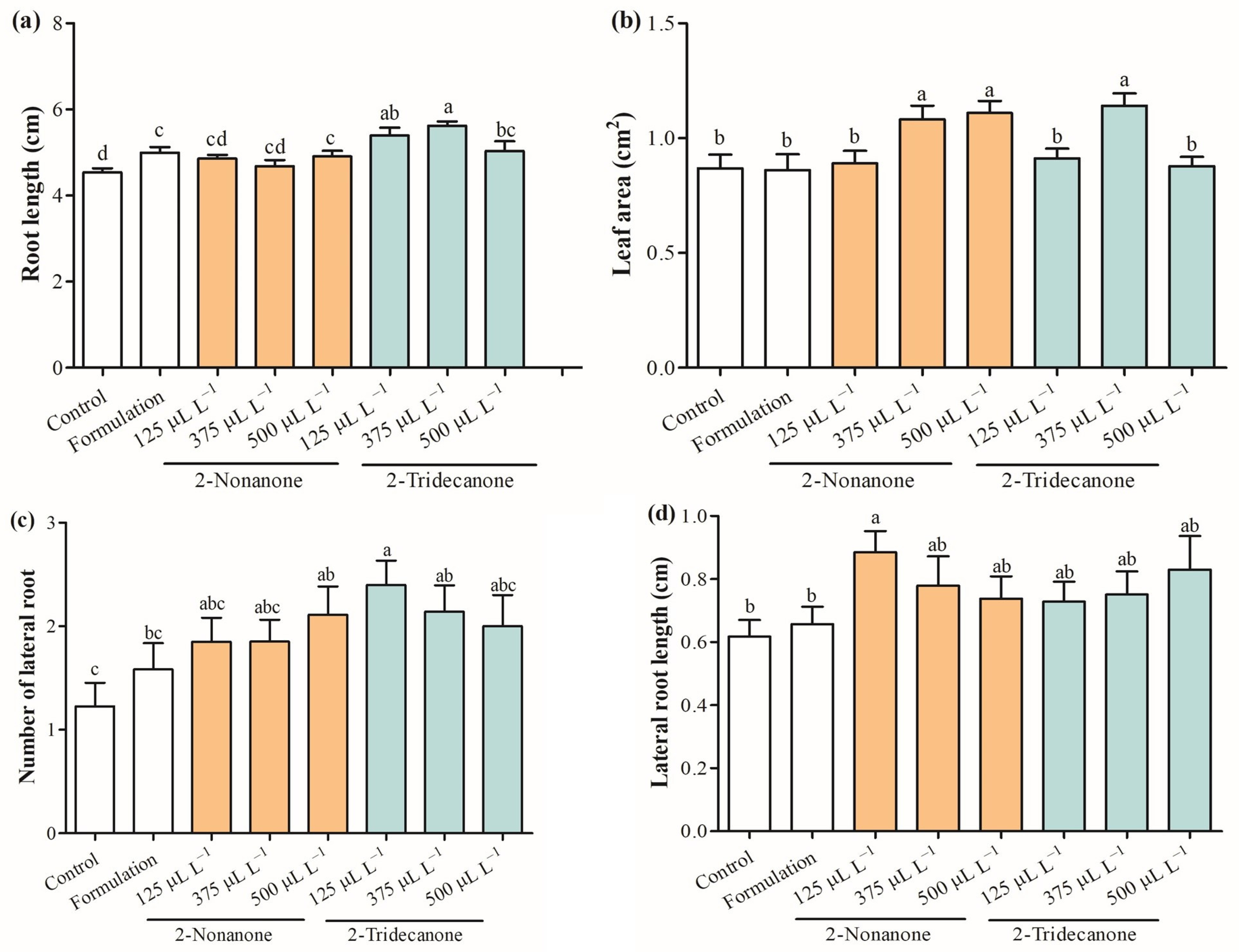
2.3. Plant Growth
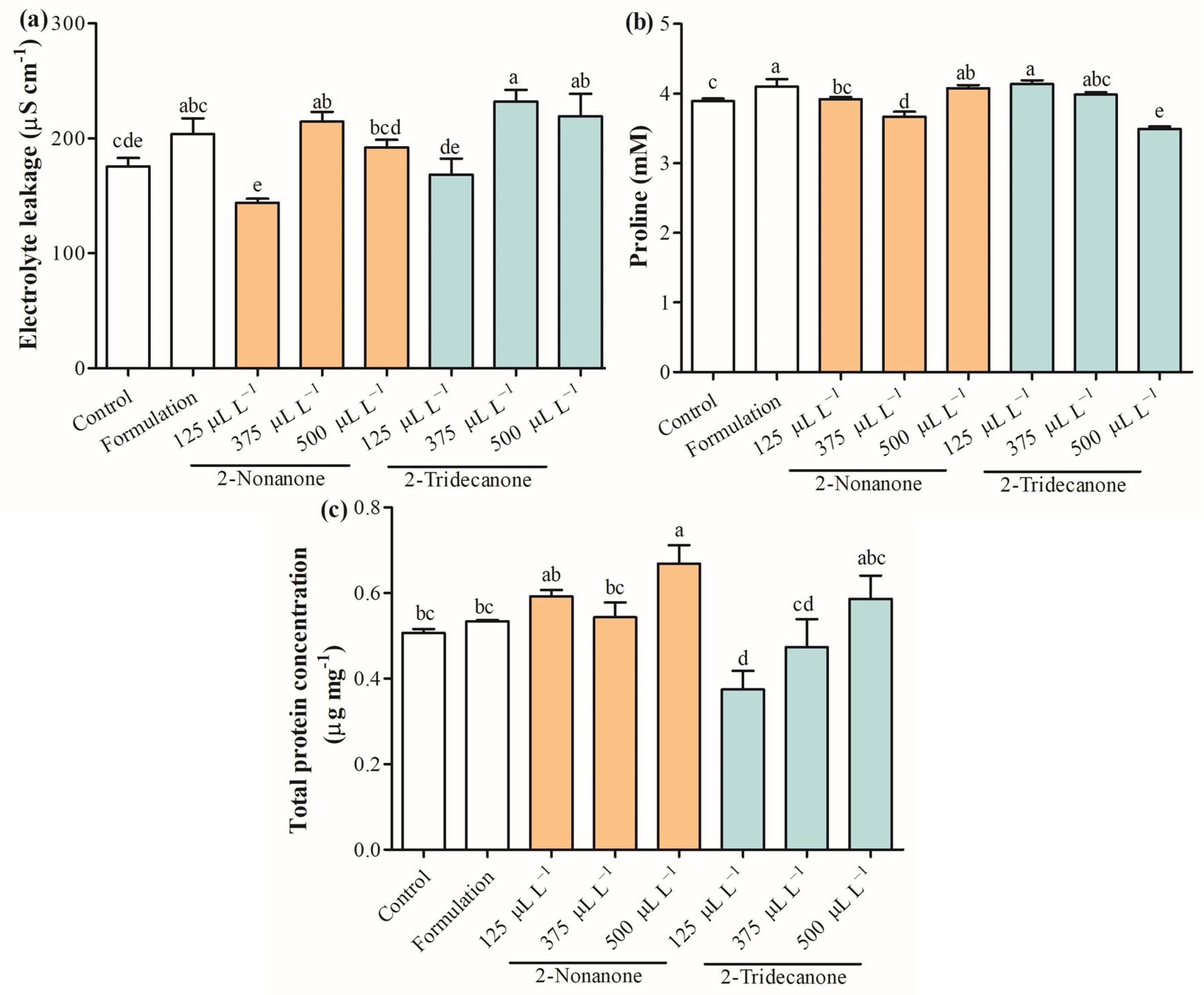
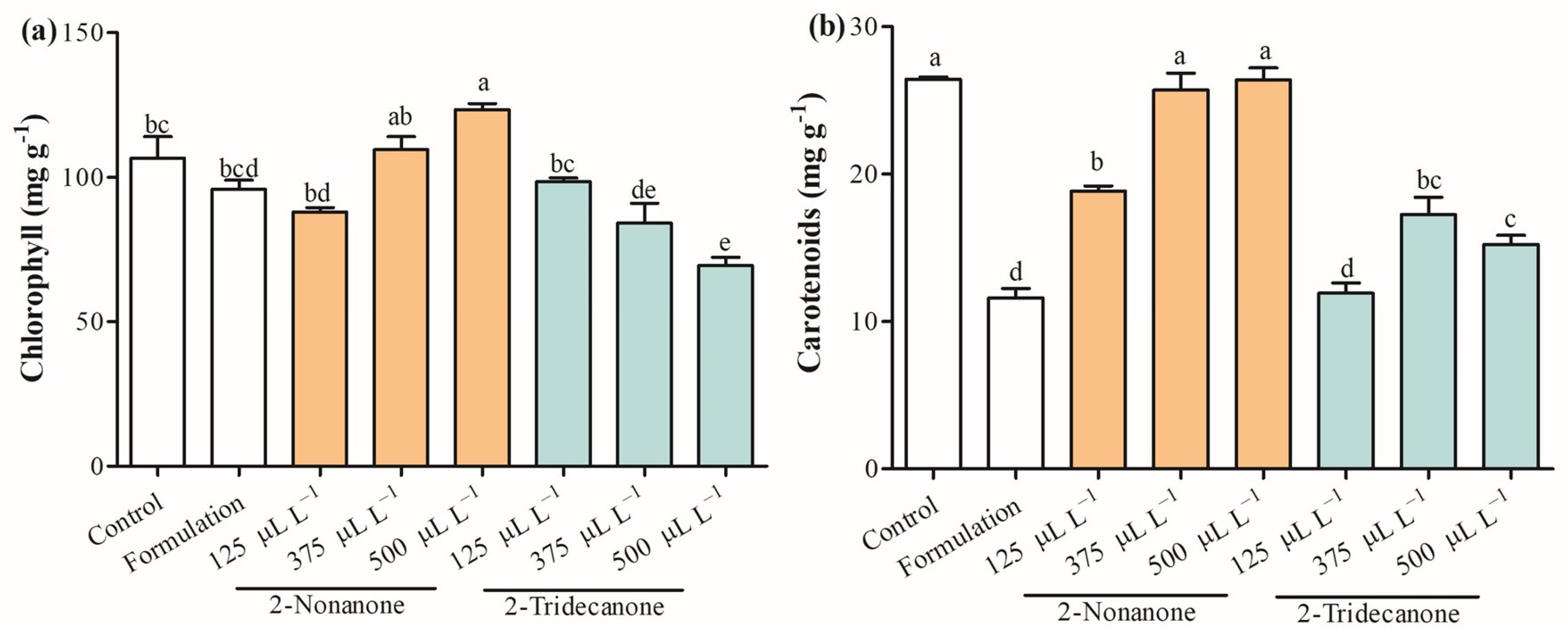
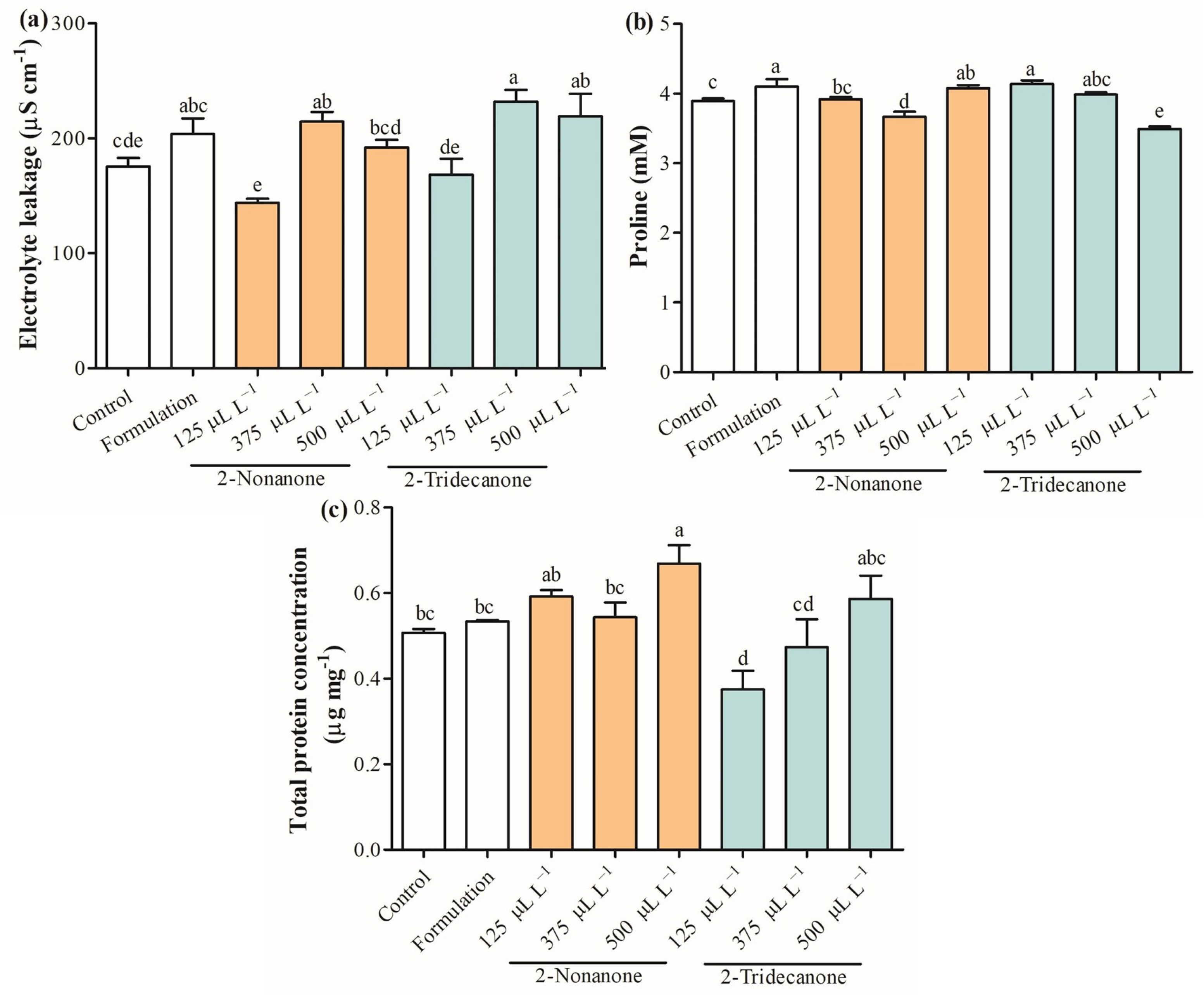
2.4. Photosynthetic Pigments and Physiological Parameters
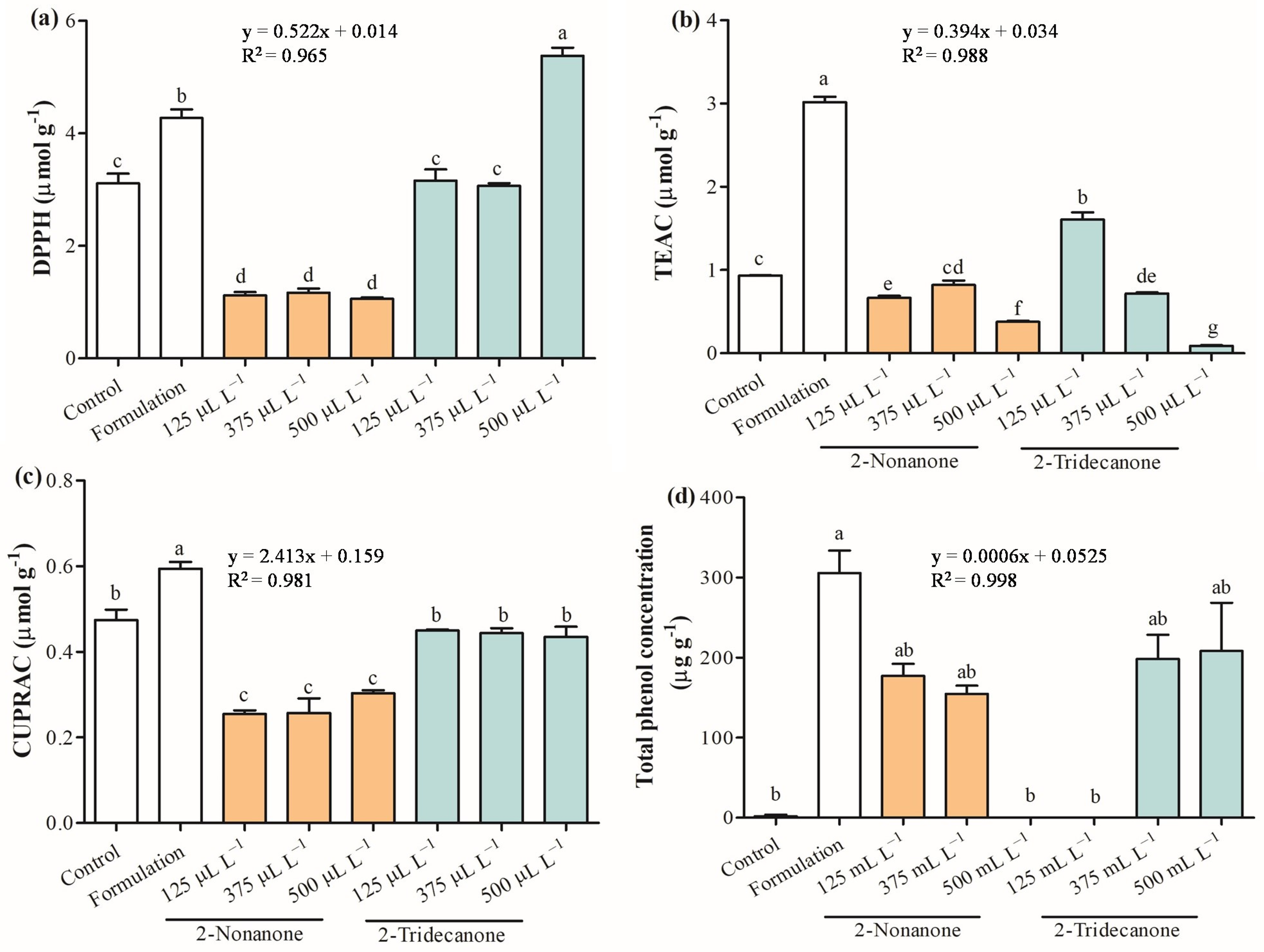
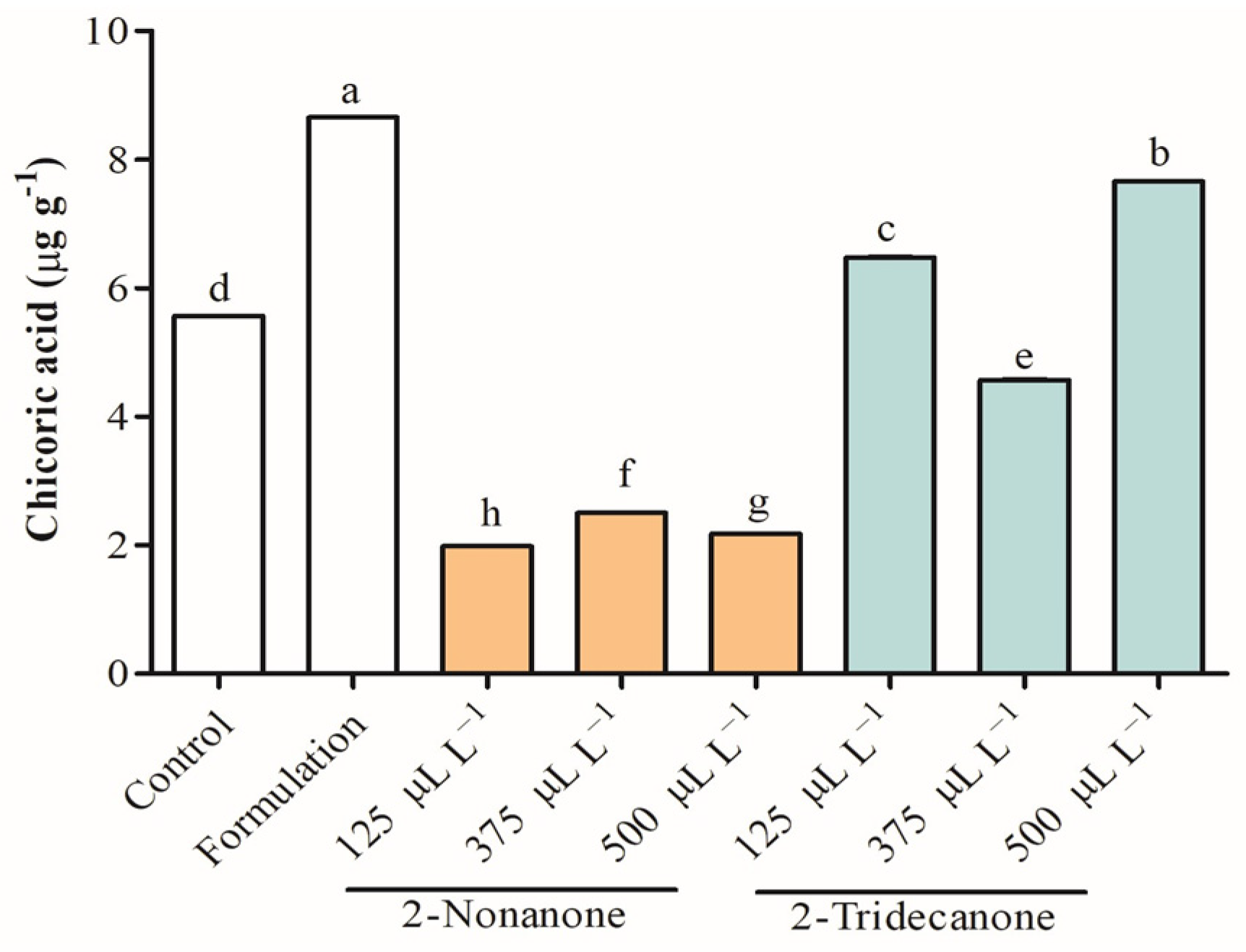
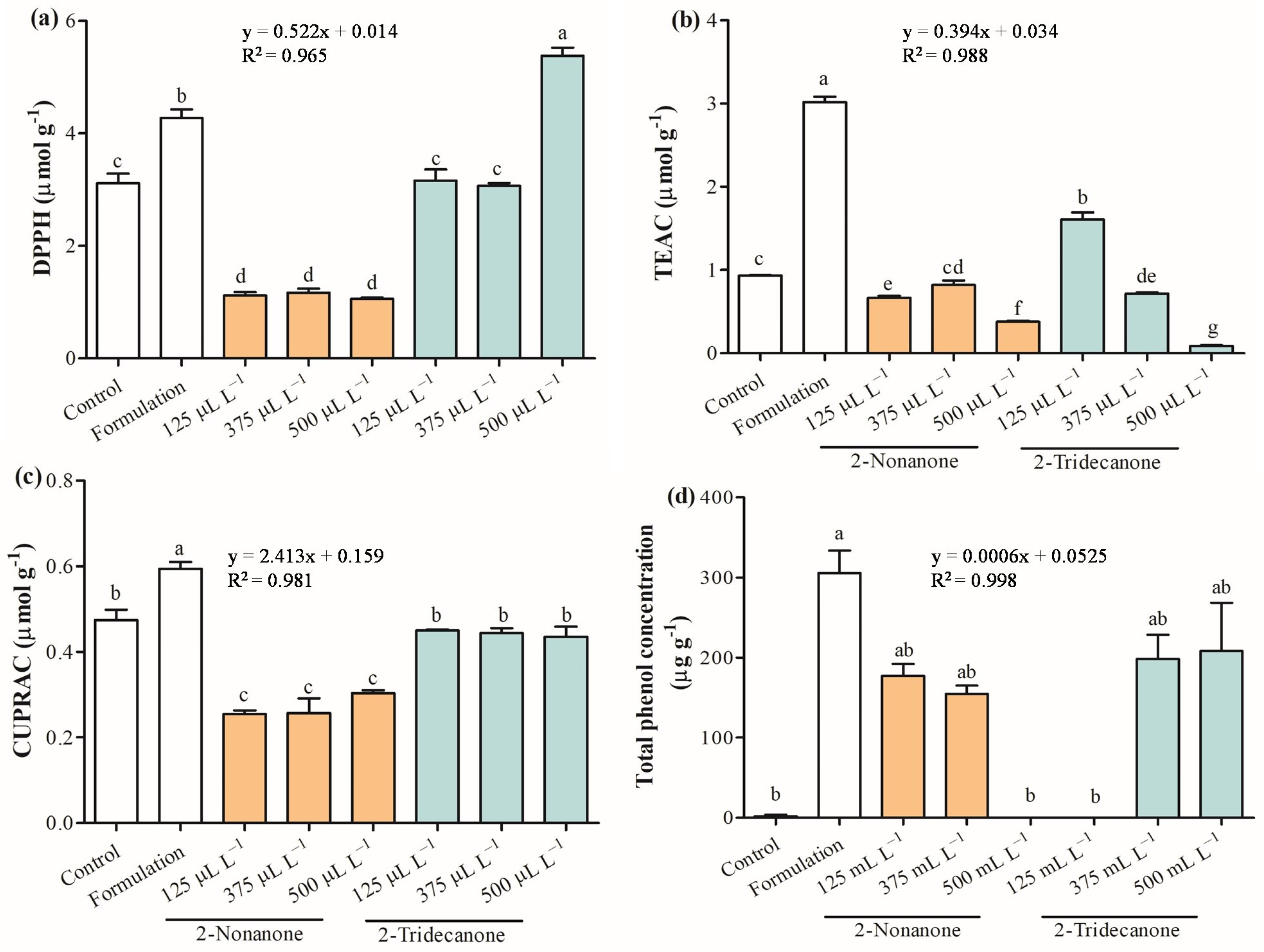
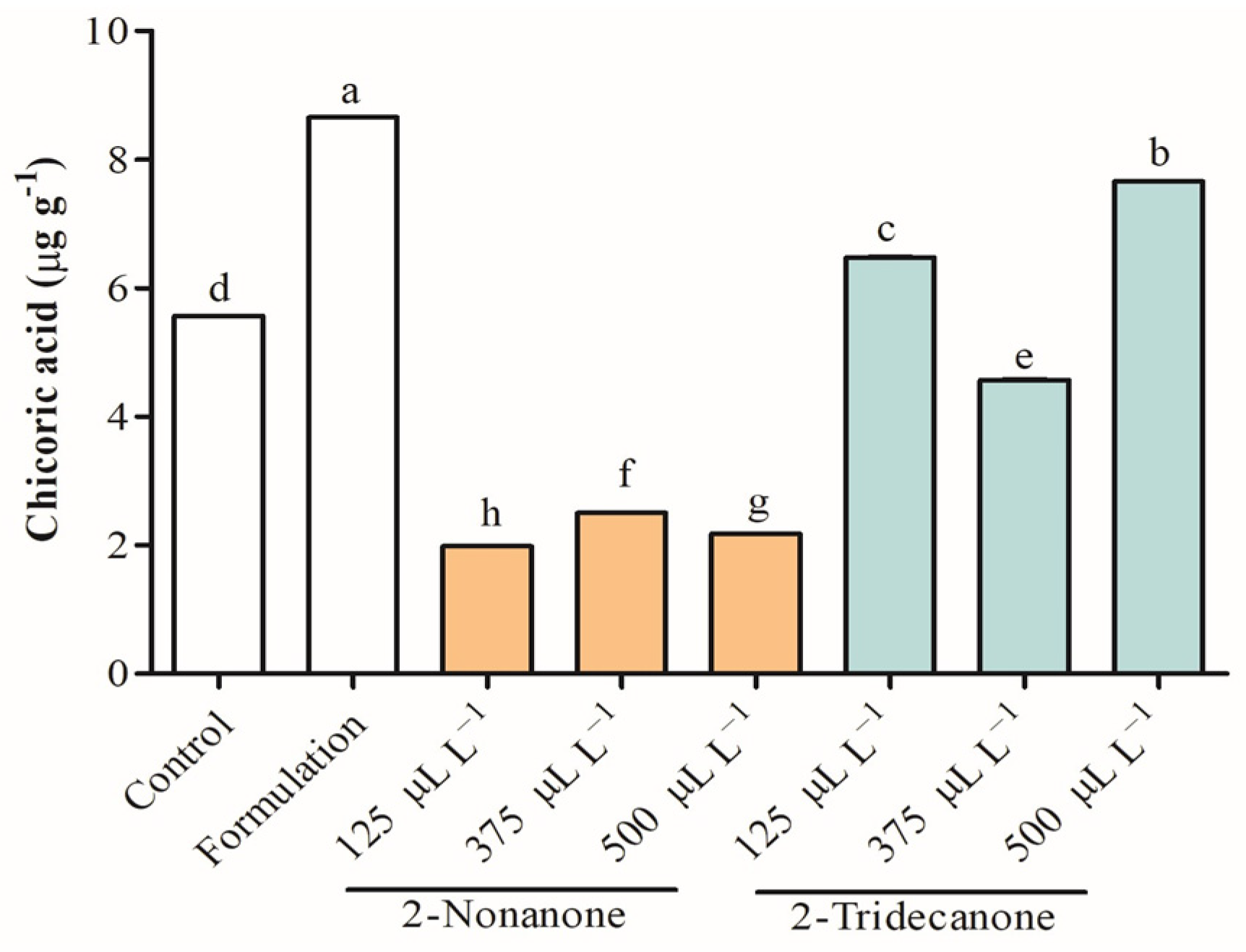
2.5. Antioxidant Capacity
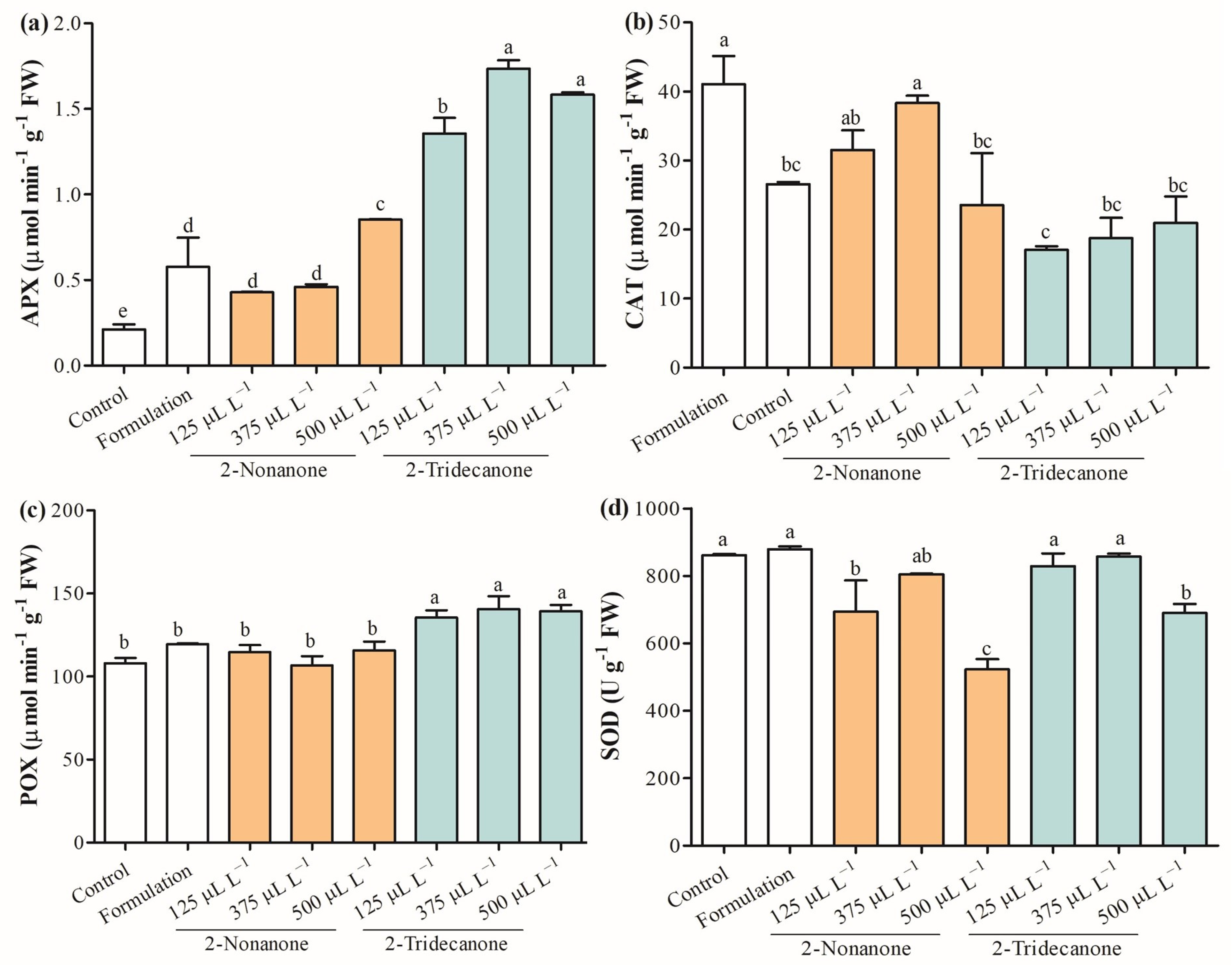
2.6. Antioxidant Enzymes
3. Discussion
4. Materials and Methods
4.1. Formulation of SLN Containing 2-Nonanone and 2-Tridecanone
4.2. Characterization of Solid Lipid Nanoparticles
4.3. Quantification of 2-Nonanone and 2-Tridecanone Released from SLN by HS-GC-FID
4.4. Plant Experiment and Growth Conditions
4.5. Photosynthetic Pigments
4.6. Physiological Parameters
4.7. Determination of Total Phenols
4.8. Identification of Chicoric Acid
4.9. Antioxidant Capacity
4.10. Antioxidant Enzyme Activities
4.11. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. World Fertilizer Trends and Outlook to 2022; FAO: Rome, Italy, 2019. [Google Scholar]
- Nisar Pahalvi, H.; Rafya, L.; Rashid, S.; Nisar, B.; Kamili, A.N. Chapter 1: Chemical fertilizers and their impact on soil health. In Microbiota and Biofertilizers; Dar, G.H., Bhat, R.A., Mehmood, M.A., Hakeem, K.R., Eds.; Springer: Cham, Switzerland, 2021; Volume 2. [Google Scholar]
- Penuelas, J.; Coello, F.; Sardans, J. A better use of fertilizers is needed for global food security and environmental sustainability. Agric. Food Secur. 2023, 12, 5. [Google Scholar]
- Bonanomi, G.; Lorito, M.; Vinale, F.; Woo, S.L. Organic amendments, beneficial microbes, and soil microbiota: Toward a unified framework for disease suppression. Annu. Rev. Phytopathol. 2018, 56, 1–20. [Google Scholar] [PubMed]
- Koskey, G.; Wambui Mburu, S.; Awino, R.; Mugendi Njeru, E.; Maingi, J.M. Potential use of beneficial microorganisms for soil amelioration, phytopathogen biocontrol, and sustainable crop production in smallholder agroecosystems. Front. Sustain. Food Syst. 2021, 5, 606308. [Google Scholar]
- Martín-Sánchez, L.; Ariotti, C.; Garbeva, P.; Vigani, G. Investigating the effect of belowground microbial volatiles on plant nutrient status: Perspective and limitations. J. Plant Interact. 2020, 15, 188–195. [Google Scholar]
- Srikamwang, S.; Eva Onsa, N.; Sunanta, P.; Sangta, J.; Chanway, C.P.; Thanakkasaranee, S.; Sommano, S.R. Role of microbial volatile organic compounds in promoting plant growth and disease resistance in horticultural production. Plant Signal.Behav. 2023, 27, 2227440. [Google Scholar]
- Kanchiswamy, C.N.; Malnoy, M.; Maffe, M.E. Bioprospecting bacterial and fungal volatiles for sustainable agriculture. Trends Plant Sci. 2015, 20, 206–211. [Google Scholar] [PubMed]
- Chandrasekaran, M.; Paramasivan, M.; Sahayarayan, J.J. Microbial volatile organic compounds: An alternative for chemical fertilizers in sustainable agriculture development. Microorganisms 2023, 11, 42. [Google Scholar]
- Russo, A.; Pollastri, S.; Ruocco, M.; Monti, M.M.; Loreto, F. Volatile organic compounds in the interaction between plants and beneficial microorganisms. J. Plant Interact. 2022, 17, 840–852. [Google Scholar]
- Fincheira, P.; Quiroz, A.; Tortella, G.; Diez, M.C.; Rubilar, O. Current advances in plant-microbe communication via volatile organic compounds as an innovative strategy to improve plant growth. Microbiol. Res. 2021, 247, 126726. [Google Scholar] [PubMed]
- Fincheira, P.; Parra, L.; Mutis, A.; Parada, M.; Quiroz, A. Volatiles emitted by Bacillus sp. BCT9 act as growth modulating agents on Lactuca sativa seedlings. Microbiol. Res. 2017, 203, 47–56. [Google Scholar] [CrossRef]
- Fincheira, P.; Quiroz, A. Physiological response of Lactuca sativa exposed to 2-nonanone emitted by Bacillus sp. BCT9. Microbiol. Res. 2019, 219, 49–55. [Google Scholar]
- Kumar, A.; Choudhary, A.; Kaur, H.; Mehta, S.; Husen, A. Smart nanomaterial and nanocomposite with advanced agrochemical activities. Nanoscale Res. Lett. 2021, 16, 156. [Google Scholar]
- Sampedro-Guerrero, J.; Vives-Peris, V.; Gomez-Cadenas, A.; Clausell-Terol, C. Efficient strategies for controlled release of nanoencapsulated phytohormones to improve plant stress tolerance. Plant Methods 2023, 19, 47. [Google Scholar]
- Kumar, S.; Nehra, M.; Dilbaghi, N.; Marrazza, G.; Aly Hassan, A.; Kim, K.H. Nano-based smart pesticide formulations: Emerging opportunities for agriculture. J. Control. Release 2019, 294, 131–153. [Google Scholar] [PubMed]
- An, C.; Sun, C.; Li, N.; Huang, B.; Jiang, J.; Shen, Y.; Wang, C.; Zhao, X.; Cui, B.; Wang, C.; et al. Nanomaterials and nanotechnology for the delivery of agrochemicals: Strategies towards sustainable agriculture. J. Nanobiotechnol. 2022, 20, 11. [Google Scholar]
- Kaliamurthi, S.; Selvaraj, G.; Hou, L.; Li, Z.; Wei, Y.; Gu, K.; Wei, D. Synergism of essential oils with lipid based nanocarriers: Emerging trends in preservation of grains and related food products. GOST 2019, 2, 21–26. [Google Scholar]
- Souto, E.B.; Severino, P.; Marques, C.; Andrade, L.N.; Durazzo, A.; Lucarini, M.; Atanasov, A.G.; El Maimouni, S.; Novellino, E.; Santini, A. Croton argyrophyllus Kunth essential oil-loaded solid lipid nanoparticles: Evaluation of release profile, antioxidant activity and cytotoxicity in a neuroblastoma cell line. Sustainability 2020, 12, 7697. [Google Scholar]
- Ganilho, C.; Bessa da Silva, M.; Paiva, C.; de Menezes, T.I.; Roncaglia dos Santos, M.; Pereira, C.M.; Pereira, R.; Andreani, T. Environmental safety assessments of lipid nanoparticles loaded with lambda-cyhalothrin. Nanomaterials 2022, 12, 2576. [Google Scholar]
- Daszkiewicz, T. Food production in the context of global developmental challenges. Agriculture 2022, 12, 832. [Google Scholar]
- Khan, M.N.; Mobin, M.; Abbas, Z.K.; Alamri, S.A. Fertilizers and Their Contaminants in Soils, Surface and Groundwater. In The Encyclopedia of the Anthropocene; DellaSala, D.A., Goldstein, M.I., Eds.; Elsevier: Oxford, UK, 2018; Volume 5, pp. 225–240. [Google Scholar]
- Cellini, A.; Spinelli, F.; Donati, I.; Ryu, C.M.; Kloepper, J.W. Bacterial volatile compound-based tools for crop management and quality. Trends Plant Sci. 2021, 29, 968–983. [Google Scholar]
- Abbas, F.; O’Neill Rothenberg, D.; Zhou, Y.; Ke, Y.; Wang, H.C. Volatile organic compounds as mediators of plant communication and adaptation to climate change. Physiol. Plant. 2022, 174, e13840. [Google Scholar] [PubMed]
- Fincheira, P.; Parada, M.; Quiroz, A. Volatile organic compounds stimulate plant growing and seed germination of Lactuca sativa. J. Soil Sci. Plant Nutr. 2017, 17, 853–867. [Google Scholar] [CrossRef]
- Rawat, M.K.; Jain, A.; Mishra, A.; Muthu, M.S.; Singh, S. Effect of lipid matrix on repaglinide-loaded solid lipid nanoparticles for oral delivery. Ther. Deliv. 2010, 1, 63–73. [Google Scholar] [CrossRef]
- Guo, S.J.; Ma, C.G.; Hu, Y.Y.; Bai, G.; Song, Z.J.; Cao, X.Q. Solid lipid nanoparticles for phytosterols delivery: The acyl chain number of the glyceride matrix affects the arrangement, stability, and release. Food Chem. 2022, 394, 133412. [Google Scholar]
- Ludtke, F.L.; Aparecida Stahl, M.; Grimaldi, R.; Soares Forte, M.B.; Gigante, M.L.; Badan Ribeiro, A.P. Optimization of high pressure homogenization conditions to produce nanostructured lipid carriers using natural and synthetic emulsifiers. Food Res. Int. 2022, 160, 111746. [Google Scholar]
- Oehlke, K.; Behsnilian, D.; Mayer-Miebach, E.; Weidler, P.G.; Greiner, R. Edible solid lipid nanoparticles (SLN) as carrier system for antioxidants of different lipophilicity. PLoS ONE 2017, 12, e0171662. [Google Scholar] [CrossRef]
- Fincheira, P.; Rubilar, O.; Tortella, G.; Medina, C.; Seabra, A.B.; Nascimento, M.H.M.; Diez, M.C.; Quiroz, A. Formulation of a controlled-release carrier for 2-ketones based on solid lipid nanoparticles to increase seedling growth in Lactuca sativa and Solanum lycopersicum. J. Soil Sci. Plant Nutr. 2021, 21, 3002–3015. [Google Scholar]
- Lowry, G.V.; Hill, R.J.; Harper, S.; Rawle, A.F.; Ogilvie Hendren, C.; Klaessig, F.; Nobbmann, U.; Sayre, P.; Rumble, J. Guidance to improve the scientific value of zeta-potential measurements in nanoEHS. Environ. Sci. Nano. 2016, 3, 953. [Google Scholar] [CrossRef]
- Lunardi, C.N.; Gomes, A.J.; Rocha, F.S.; De Tommaso, J.; Patience, G.S. Experimental methods in chemical engineering: Zeta potential. Can. J. Chem. Eng. 2020, 99, 627–639. [Google Scholar]
- Fincheira, P.; Quiroz, A.; Medina, C.; Tortella, G.; Hermosilla, E.; Diez, M.C.; Rubilar, O. Plant growth induction by volatile organic compound released from solid lipid nanoparticles and nanostructured lipid carriers. Colloids Surf. A Physicochem. Eng. Asp. 2020, 596, 124739. [Google Scholar]
- Shishkova, S.; Huang, L.; Rodríguez, R.E.; Ristova, D.; DelloIoio, R. Editorial: Root development: Towards understanding regulatory networks and complex interactions between cell populations. Front. Plant Sci. 2023, 13, 1108367. [Google Scholar] [CrossRef]
- Zhang, W.; Fang, D.; Dong, K.; Hu, F.; Ye, Z.; Cao, J. Insights into the environmental factors shaping lateral root development. Physiol. Plant. 2023, 175, e13878. [Google Scholar]
- Gao, Y.; Feng, J.; Wu, J.; Wang, K.; Wu, S.; Liu, H.; Jiang, M. Transcriptome analysis of the growth-promoting effect of volatile organic compounds produced by Microbacteriumaurantiacum GX14001on tobacco (Nicotiana benthamiana). BMC Plant Biol. 2022, 22, 208. [Google Scholar]
- Zhang, H.; Kim, M.S.; Krishnamachari, V.; Payton, P.; Sun, Y.; Grimson, M.; Farag, M.A.; Ryu, C.M.; Allen, R.; Melo, I.S.; et al. Rhizobacterial volatile emissions regulate auxin homeostasis and cell expansion in Arabidopsis. Planta 2007, 226, 839–851. [Google Scholar]
- Ryu, C.M.; Farag, M.A.; Hu, C.H.; Reddy, M.S.; Wei, H.X.; Pare, P.W. Bacterial volatiles promote growth in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 4927–4932. [Google Scholar] [PubMed]
- Zhang, H.; Ge, Y.; Xie, X.; Atefi, A.; Wijewardane, N.K.; Thapa, S. High throughput analysis of leaf chlorophyll content in sorghum using RGB, hyperspectral, and fluorescence imaging and sensor fusion. Plant Methods 2022, 18, 60. [Google Scholar]
- Stra, A.; Almarwaey, L.O.; Alagoz, Y.; Moreno, J.C.; Al-Babili, S. Carotenoid metabolism: New insights and synthetic approaches. Front. Plant Sci. 2023, 13, 1072061. [Google Scholar]
- Hatsugai, N.; Katagiri, F. Quantification of plant cell death by electrolyte leakage assay. Bio-protocol 2018, 8, e2758. [Google Scholar] [CrossRef]
- Zdunek-Zastocka, E.; Grabowska, A.; Michniewska, B.; Orzechowski, S. Proline concentration and its metabolism are regulated in a leaf age dependent manner but not by abscisic acid in pea plants exposed to cadmium stress. Cells 2021, 10, 946. [Google Scholar]
- Chun, S.C.; Paramasivan, M.; Chandrasekaran, M. Proline accumulation influenced by osmotic stress in arbuscular mycorrhizal symbiotic plants. Front. Microbiol. 2018, 9, 2525. [Google Scholar]
- Zaguri, M.; Kandel, S.; Rinehart, S.A.; Torsekar, V.R.; Hawlena, D. Protein quantification in ecological studies: A literature review and empirical comparisons of standard methodologies. Methods Ecol. Evol. 2020, 12, 1240–1251. [Google Scholar] [CrossRef]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive oxygen species signaling in plant stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar]
- Rajput, V.D.; Harish; Kumar Singh, R.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Singh Gour, V.; Minkina, T.; Sushkova, S.; et al. Recent developments in enzymatic antioxidant defence mechanism in plants with special reference to abiotic stress. Biology 2021, 10, 267. [Google Scholar] [PubMed]
- Rahman, M.M.; Islam, M.B.; Biswas, M.; Khurshid Alam, A.H.M. In vitro antioxidant and free radical scavenging activity of different parts of Tabebuia pallida growing in Bangladesh. BMC Res. Notes 2015, 8, 621. [Google Scholar] [CrossRef] [PubMed]
- Villegas-Aguilar, M.C.; Cádiz-Gurrea, M.L.; Arráez-Román, D.; Segura-Carretero, A. Comparative evaluation of the total antioxidant capacities of plant polyphenols in different natural sources. Med. Sci. Forum. 2021, 2, 1. [Google Scholar]
- Molole, G.J.; Gure, A.; Abdissa, N. Determination of total phenolic content and antioxidant activity of Commiphoramollis (Oliv.) Engl. resin. BMC Chem. Biol. 2022, 16, 48. [Google Scholar]
- Platzer, M.; Kiese, S.; Herfellner, T.; Schweiggert-Weisz, U.; Eisner, P. How does the phenol structure influence the results of the Folin-Ciocalteuassay? Antioxidants 2021, 10, 811. [Google Scholar] [CrossRef]
- Shi, L.; Zhao, W.; Yang, Z.; Subbiah, V.; Rasul Suleria, H.A. Extraction and characterization of phenolic compounds and their potential antioxidant activities. Environ. Sci. Pollut. Res. 2022, 29, 81112–81129. [Google Scholar]
- Yang, M.; Wu, C.; Zhang, T.; Shi, L.; Li, J.; Liang, H.; Lv, X.; Jing, F.; Qin, L.; Zhao, T.; et al. Chicoric acid: Natural occurrence, chemical synthesis, biosynthesis, and their bioactive effects. Front. Chem. 2022, 10, 888673. [Google Scholar]
- Dumanovic, J.; Nepovimova, E.; Natič, M.; Kuča, K.; Jačevič, V. The significance of reactive oxygen species and antioxidant defense system in plants: A concise overview. Front. Plant Sci. 2021, 11, 552969. [Google Scholar]
- Hasanuzzaman, M.; BorhannuddinBhuyan, M.H.M.; Zulfiqar, F.; Raza, A.; Mohammad Mohsin, S.; Al Mahmud, J.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Londhe, V.; Save, S. Zaltoprofen loaded solid lipid nanoparticles for topical delivery: Formulation design, In Vitro and Ex Vivo evaluation. MOJBioequiv. Bioavailab. 2017, 4, 248–254. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determination of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 603, 591–592. [Google Scholar] [CrossRef]
- Omezzine, F.; Ladhari, A.; Haouala, R. Physiological and biochemical mechanisms of allelochemicals in aqueous extracts of diploid and mixoploid Trigonellafoenum-graecum L. S. Afr. J. Bot. 2014, 93, 167–178. [Google Scholar] [CrossRef]
- Ercoli, S.; Cartes, J.; Cornejo, P.; Tereucán, G.; Winterhalter, P.; Contreras, B.; Ruiz, A. Stability of phenolic compounds, antioxidant activity and colour parameters of a coloured extract obtained from coloured-flesh potatoes. LWT—Food Sci. Technol. 2021, 136, 110370. [Google Scholar] [CrossRef]
- Valdebenito, A.; Nahuelcura, J.; Santander, C.; Cornejo, P.; Contreras, B.; Gómez-Alonso, S.; Ruiz, A. Physiological and metabolic effects of the inoculation of arbuscular mycorrhizal fungi in Solanum tuberosum Crops under water stress. Plants 2022, 11, 2539. [Google Scholar]
- Parada, J.; Valenzuela, T.; Gómez, F.; Tereucán, G.; García, S.; Cornejo, P.; Winterhalter, P.; Ruiz, A. Effect of fertilization and arbuscular mycorrhizal fungal inoculation on antioxidant profiles and activities in Fragaria ananassa fruit. J. Sci. Food Agric. 2019, 99, 1397–1404. [Google Scholar] [CrossRef]
- Nova-Baza, D.; Olivares-Caro, L.; Bustamante, L.; Pérez, A.J.; Vergara, C.; Fuentealba, J.; Mardones, C. Metabolic profile and antioxidant capacity of five Berberis leaves species: A comprehensive study to determine their potential as natural food or ingredient. Food Res. Int. 2022, 160, 111642. [Google Scholar] [PubMed]
- Fritz, V.; Tereucán, G.; Santander, C.; Contreras, B.; Cornejo, P.; Avelar Ferreira, P.A.; Ruiz, A. Effect of inoculation with arbuscular mycorrhizal fungi and fungicide application on the secondary metabolism of Solanum tuberosum Leaves. Plants 2022, 11, 278. [Google Scholar] [CrossRef] [PubMed]
- Kohatsu, M.Y.; Lange, C.N.; Pelegrino, M.T.; Pieretti, J.C.; Tortella, G.; Rubilar, O.; Batista, B.L.; Seabra, A.B.; Araujo de Jesus, T. Foliar spraying of biogenic CuO nanoparticles protects the defence system and photosynthetic pigments of lettuce (Lactuca sativa). J. Clean. Prod. 2021, 324, 129264. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2-Nonanone | 2-Tridecanone | |||||||
|---|---|---|---|---|---|---|---|---|
| F | 125 µL L−1 | 375 µL L−1 | 500 µL L−1 | F | 125 µL L−1 | 375 µL L−1 | 500 µL L−1 | |
| Month-1 | ||||||||
| Size (nm) | 352 ± 6.8 | 306 ±8.4 | 263 ± 10.7 | 306 ± 18.6 | 245 ± 4.6 | 253 ± 4.0 | 188± 10.0 | 261 ± 18.4 |
| PDI | 0.39 ± 0.07 | 0.38 ± 0.03 | 0.35 ± 0.03 | 0.35 ± 0.03 | 0.27 ± 0.01 | 0.25 ± 0.02 | 0.26 ± 0.01 | 0.25 ± 0.03 |
| ζ-potential (mV) | −22.4 ± 1.2 | −22.9 ± 0.7 | −22.1 ± 1.4 | −24.3 ± 0.9 | −19.2 ± 0.4 | −17.9 ± 0.9 | −16.0 ± 0.72 | −20.0 ± 1.21 |
| Month-2 | ||||||||
| Size (nm) | 281 ± 4.0 | 292 ± 30.7 | 246 ± 10.1 | 270 ± 11.3 | 234 ± 4.6 | 250 ± 4.4 | 187± 42.4 | 258 ± 17.6 |
| PDI | 0.24 ± 0.01 | 0.34 ± 0.12 | 0.28 ± 0.3 | 0.26 ± 0.2 | 0.26 ± 0.01 | 0.25 ± 0.01 | 0.26 ± 0.04 | 0.28 ± 0.07 |
| ζ-potential (mV) | −20.4 ±0.9 | −20.5 ±1.2 | −19.2 ±0.4 | −21.1 ±0.9 | −18.9 ± 0.4 | −17.8 ± 1.1 | −15.4 ± 2.0 | −16.8 ± 3.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fincheira, P.; Espinoza, J.; Vera, J.; Berrios, D.; Nahuelcura, J.; Ruiz, A.; Quiroz, A.; Bustamante, L.; Cornejo, P.; Tortella, G.; et al. The Impact of 2-Ketones Released from Solid Lipid Nanoparticles on Growth Modulation and Antioxidant System of Lactuca sativa. Plants 2023, 12, 3094. https://doi.org/10.3390/plants12173094
Fincheira P, Espinoza J, Vera J, Berrios D, Nahuelcura J, Ruiz A, Quiroz A, Bustamante L, Cornejo P, Tortella G, et al. The Impact of 2-Ketones Released from Solid Lipid Nanoparticles on Growth Modulation and Antioxidant System of Lactuca sativa. Plants. 2023; 12(17):3094. https://doi.org/10.3390/plants12173094
Chicago/Turabian StyleFincheira, Paola, Javier Espinoza, Joelis Vera, Daniela Berrios, Javiera Nahuelcura, Antonieta Ruiz, Andrés Quiroz, Luis Bustamante, Pablo Cornejo, Gonzalo Tortella, and et al. 2023. "The Impact of 2-Ketones Released from Solid Lipid Nanoparticles on Growth Modulation and Antioxidant System of Lactuca sativa" Plants 12, no. 17: 3094. https://doi.org/10.3390/plants12173094
APA StyleFincheira, P., Espinoza, J., Vera, J., Berrios, D., Nahuelcura, J., Ruiz, A., Quiroz, A., Bustamante, L., Cornejo, P., Tortella, G., Diez, M. C., Benavides-Mendoza, A., & Rubilar, O. (2023). The Impact of 2-Ketones Released from Solid Lipid Nanoparticles on Growth Modulation and Antioxidant System of Lactuca sativa. Plants, 12(17), 3094. https://doi.org/10.3390/plants12173094