
The Research Process of PSK Biosynthesis, Signaling Transduction, and Potential Applications in Brassica napus


Abstract
:1. Introduction
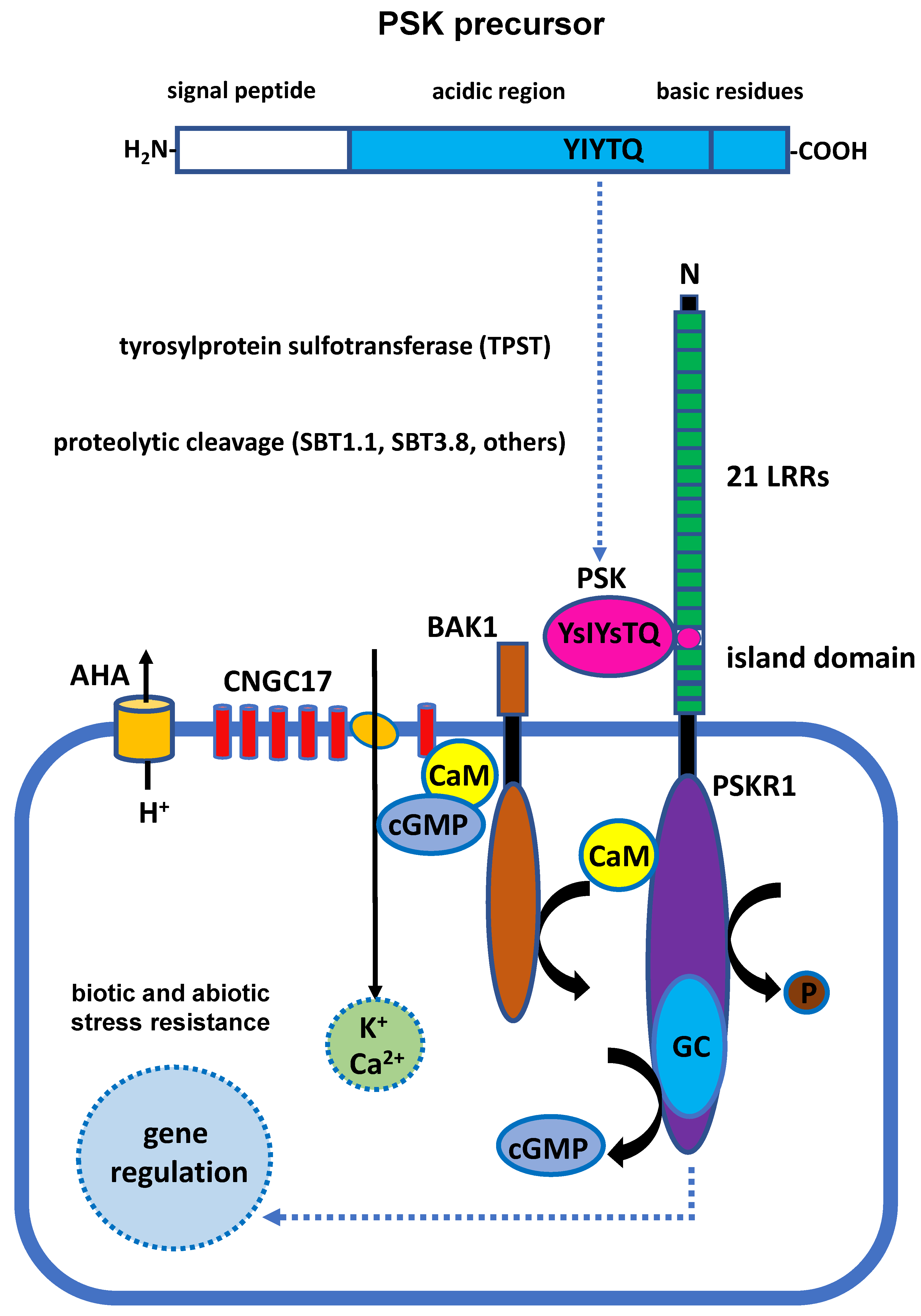
2. Synthesis and Signal Transduction of PSK Peptide
2.1. Biosynthesis of the PSK Peptide
2.2. Characteristics of PSK Receptors
2.3. PSKR-Dependent Signal Transduction
3. Physiological Functions of PSK
3.1. PSK Controls the Growth and Development of Plant Cells

{kind=link}
{kind=link}
{kind=link}
| Major Function | Function in Detail | Species | Reference |
|---|---|---|---|
| PSK controls the growth and development of plant cells | cell proliferation | Asparagus officinalis, Oryza sativa, Daucus carota | [1]; [15]; [29] |
| induced cell differentiation and somatic cell embryogenesis | Cucumis sativus, Daucus carota, Cryptomeria japonica, Cunninghamia lanceolata | [30]; [5]; [31]; [32] | |
| regulation of root elongation and fiber development | Arabidopsis, Gossypium species, Triticum aestivum | [36]; [40]; [41] | |
| increased crop yield | Triticum aestivum, Glycine max | [42]; [41] | |
| regulatory factor in pollen population effects | Nicotiana tabacum | [43] | |
| guide pollen into the embryo sac and enable them to elongate normally | Arabidopsis Pyrus bretschneideri | [45]; [46] | |
| PSK promotes callus formation and efficiency of tissue culture in vitro | induced cell regeneration | Brassica oleracea | [47] |
| enhanced the transformation efficiency of candidate genes | Daucus carota | [48] | |
| PSK regulates plant adaptation in response to environmental stresses | enhanced heat resistance | Arabidopsis | [49] |
| enhanced drought resistance | Oryza sativa, Arabidopsis Solanum lycopersicum | [50]; [7]; [51] | |
| delayed senescence | Fragaria ×ananassa, Rosa hybrida, Brassica oleracea var. Italica | [52]; [53]; [54] | |
| preserved the quality and delayed cold damage | Musa paradisiaca, Prunus persica | [55]; [56] | |
| pathogen resistance | Solanum lycopersicum, Arabidopsis, Oryza sativa | [28,57]; [58,59]; [60] | |
| balance plant immune signaling | Arabidopsis | [61] |
3.2. PSK Promotes Callus Formation and Efficiency of Tissue Culture In Vitro
3.3. PSK Regulates Plant Adaptation in Response to Environmental Stresses
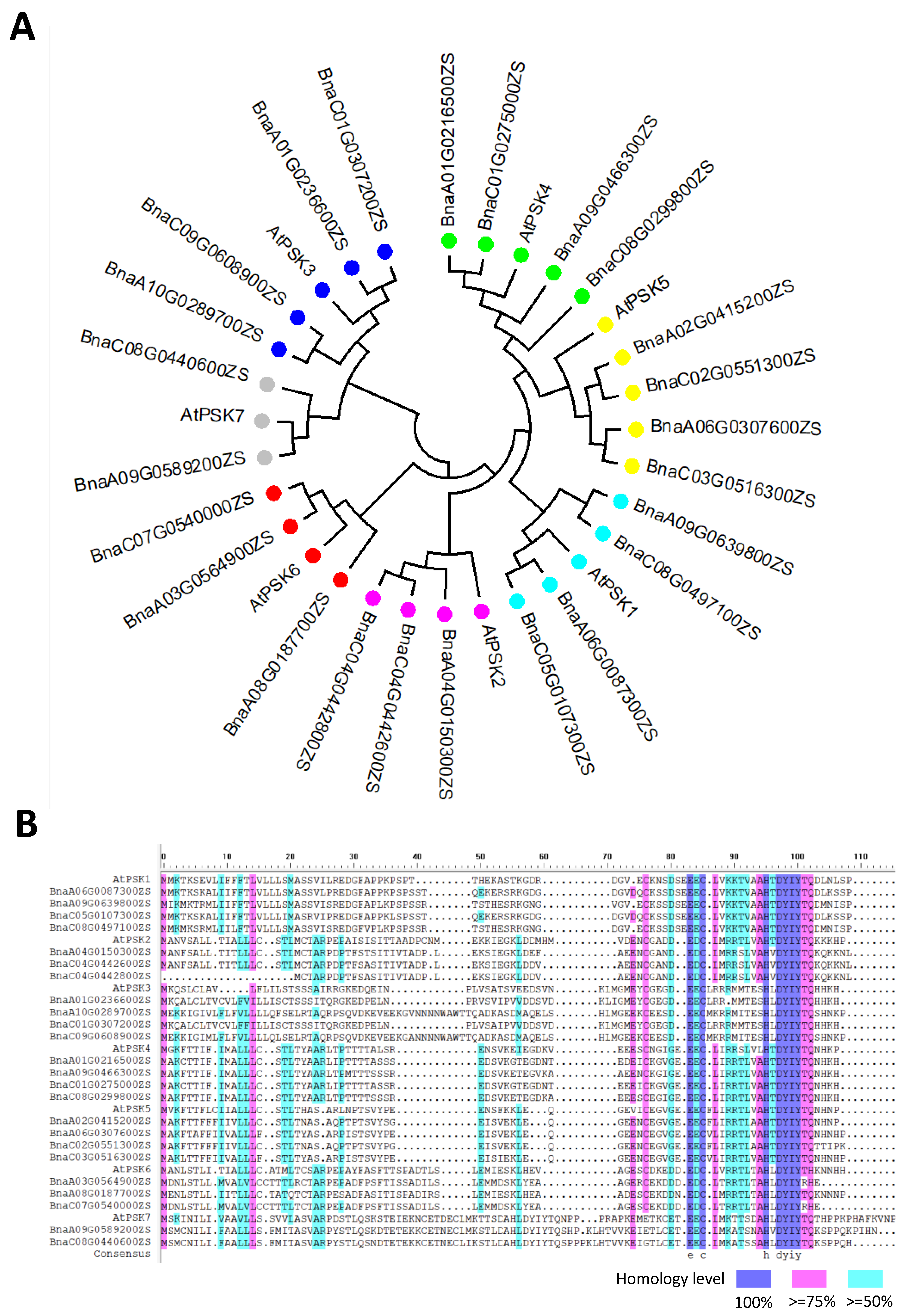
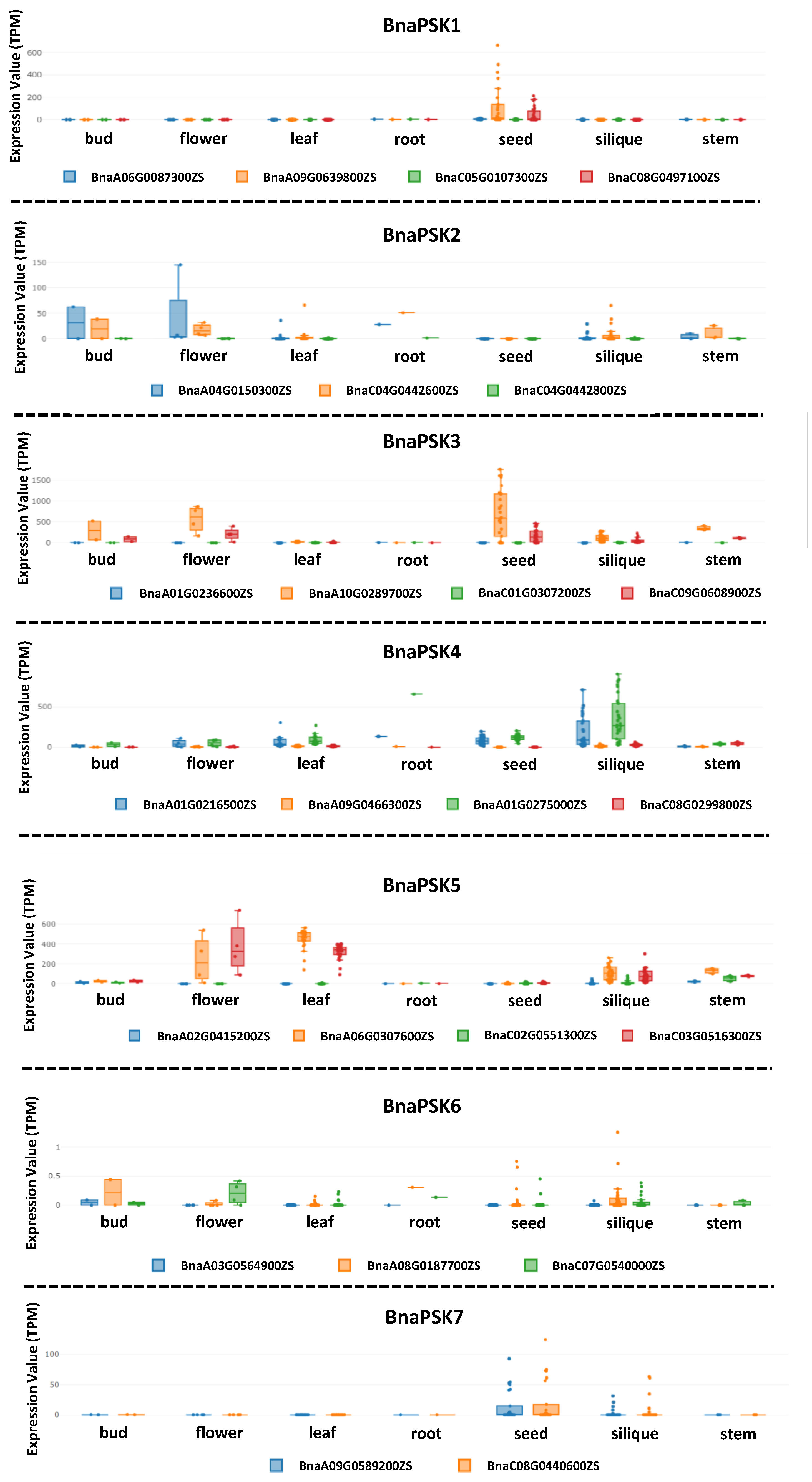
4. PSK-Related Genetic Correlation between Rapeseed and Arabidopsis
5. Conclusions and Perspectives
5.1. Bottleneck and Gaps of PSK-Related Research
5.2. Putative Function of PSK on Callus Formation during Genetic Transformation in B. napus
5.3. Potential Use of PSK-Related Genes in Agricultural Breeding
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Matsubayashi, Y.; Sakagami, Y. Phytosulfokine, sulfated peptides that induce the proliferation of single mesophyll cells of Asparagus officinalis L. Proc. Natl. Acad. Sci. USA 1996, 93, 7623–7627. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Matsubayashi, Y.; Nakamura, K.; Sakagami, Y. Oryza sativa PSK gene encodes a precursor of phytosulfokine-α, a sulfated peptide growth factor found in plants. Proc. Natl. Acad. Sci. USA 1999, 96, 13560–13565. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, D.; Li, M.; Yan, J.; Luo, L.; Yu, L. Overexpression of PSK-γ in Arabidopsis promotes growth without influencing pattern-triggered immunity. Plant Signal. Behav. 2019, 14, 1684423. [Google Scholar] [CrossRef]
- Di, Q.; Li, Y.; Zhang, D.; Wu, W.; Zhang, L.; Zhao, X.; Luo, L.; Yu, L. A novel type of phytosulfokine, PSK-ε, positively regulates root elongation and formation of lateral roots and root nodules in Medicago truncatula. Plant Signal.Behav. 2022, 17, 2134672. [Google Scholar] [CrossRef] [PubMed]
- Hanai, H.; Nakayama, D.; Yang, H.; Matsubayashi, Y.; Hirota, Y.; Sakagami, Y. Existence of a plant tyrosylprotein sulfotransferase: Novel plant enzyme catalyzing tyrosine O-sulfation of preprophytosulfokine variants in vitro. FEBS Lett. 2000, 470, 97–101. [Google Scholar] [CrossRef]
- Srivastava, R.; Liu, J.X.; Howell, S.H. Proteolytic processing of a precursor protein for a growth-promoting peptide by a subtilisin serine protease in Arabidopsis. Plant J. 2008, 56, 219–227. [Google Scholar] [CrossRef]
- Stuhrwohldt, N.; Buhler, E.; Sauter, M.; Schaller, A. Phytosulfokine (PSK) precursor processing by subtilase SBT3.8 and PSK signaling improve drought stress tolerance in Arabidopsis. J. Exp. Bot. 2021, 72, 3427–3440. [Google Scholar] [CrossRef]
- Ladwig, F.; Dahlke, R.I.; Stuhrwohldt, N.; Hartmann, J.; Harter, K.; Sauter, M. Phytosulfokine regulates growth in Arabidopsis through a response module at the plasma membrane that includes CYCLIC NUCLEOTIDE-GATED CHANNEL17, H+-ATPase, and BAK1. Plant Cell 2015, 27, 1718–1729. [Google Scholar] [CrossRef]
- Matsubayashi, Y.; Sakagami, Y. Peptide hormones in plants. Annu. Rev. Plant Biol. 2006, 57, 649–674. [Google Scholar] [CrossRef]
- Stührwohldt, N.; Schaller, A. Regulation of plant peptide hormones and growth factors by post-translational modification. Plant Biol. 2019, 21 (Suppl. S1), 49–63. [Google Scholar] [CrossRef]
- Yang, H.; Matsubayashi, Y.; Nakamura, K.; Sakagami, Y. Diversity of Arabidopsis genes encoding precursors for phytosulfokine, a peptide growth factor. Plant Physiol. 2001, 127, 842–851. [Google Scholar] [CrossRef]
- Lorbiecke, R.; Sauter, M. Comparative analysis of PSK peptide growth factor precursor homologs. Plant Sci. 2002, 163, 321–332. [Google Scholar] [CrossRef]
- Stuhrwohldt, N.; Scholl, S.; Lang, L.; Katzenberger, J.; Schumacher, K.; Schaller, A. The biogenesis of CLEL peptides involves several processing events in consecutive compartments of the secretory pathway. eLife 2020, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Sauter, M. Phytosulfokine peptide signalling. J. Exp. Bot. 2015, 66, 5161–5169. [Google Scholar] [CrossRef]
- Matsubayashi, Y.; Takagi, L.; Sakagami, Y. Phytosulfokine-α, a sulfated pentapeptide, stimulates the proliferation of rice cells by means of specific high- and low-affinity binding sites. Proc. Natl. Acad. Sci. USA 1997, 94, 13357–13362. [Google Scholar] [CrossRef] [PubMed]
- Matsubayashi, Y.; Sakagami, Y. Characterization of specific binding sites for a mitogenic sulfated peptide, phytosulfokine-α, in the plasma-membrane fraction derived from Oryza sativa L. Eur. J. Biochem. 1999, 262, 666–671. [Google Scholar] [CrossRef]
- Matsubayashi, Y.; Sakagami, Y. 120- and 160-kDa receptors for endogenous mitogenic peptide, phytosulfokine-α, in rice plasma membranes. J. Biol. Chem. 2000, 275, 15520–15525. [Google Scholar] [CrossRef]
- Matsubayashi, Y.; Ogawa, M.; Morita, A.; Sakagami, Y. An LRR receptor kinase involved in perception of a peptide plant hormone, phytosulfokine. Science 2002, 296, 1470–1472. [Google Scholar] [CrossRef]
- Wang, J.; Li, H.; Han, Z.; Zhang, H.; Wang, T.; Lin, G.; Chang, J.; Yang, W.; Chai, J. Allosteric receptor activation by the plant peptide hormone phytosulfokine. Nature 2015, 525, 265–268. [Google Scholar] [CrossRef]
- Morillo, S.A.; Tax, F.E. Functional analysis of receptor-like kinases in monocots and dicots. Curr. Opin. Plant Biol. 2006, 9, 460–469. [Google Scholar] [CrossRef]
- Kaufmann, C.; Sauter, M. Pull-down assay to characterize Ca2+/calmodulin binding to plant receptor kinases. Methods Mol. Biol. 2017, 1621, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Matsubayashi, Y.; Ogawa, M.; Kihara, H.; Niwa, M.; Sakagami, Y. Disruption and overexpression of Arabidopsis phytosulfokine receptor gene affects cellular longevity and potential for growth. Plant Physiol. 2006, 142, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Amano, Y.; Tsubouchi, H.; Shinohara, H.; Ogawa, M.; Matsubayashi, Y. Tyrosine-sulfated glycopeptide involved in cellular proliferation and expansion in Arabidopsis. Proc. Natl. Acad. Sci. USA 2007, 104, 18333–18338. [Google Scholar] [CrossRef] [PubMed]
- Kwezi, L.; Ruzvidzo, O.; Wheeler, J.I.; Govender, K.; Iacuone, S.; Thompson, P.E.; Gehring, C.; Irving, H.R. The phytosulfokine (PSK) receptor is capable of guanylate cyclase activity and enabling cyclic GMP-dependent signaling in plants. J. Biol. Chem. 2011, 286, 22580–22588. [Google Scholar] [CrossRef]
- Hartmann, J.; Fischer, C.; Dietrich, P.; Sauter, M. Kinase activity and calmodulin binding are essential for growth signaling by the phytosulfokine receptor PSKR1. Plant J. 2014, 78, 192–202. [Google Scholar] [CrossRef]
- Stuhrwohldt, N.; Dahlke, R.I.; Steffens, B.; Johnson, A.; Sauter, M. Phytosulfokine-α controls hypocotyl length and cell expansion in Arabidopsis thaliana through phytosulfokine receptor 1. PLoS ONE 2011, 6, e21054. [Google Scholar] [CrossRef]
- Talke, I.N.; Blaudez, D.; Maathuis, F.J.; Sanders, D. CNGCs: Prime targets of plant cyclic nucleotide signalling? Trends Plant Sci. 2003, 8, 286–293. [Google Scholar] [CrossRef]
- Hu, Z.; Fang, H.; Zhu, C.; Gu, S.; Ding, S.; Yu, J.; Shi, K. Ubiquitylation of PHYTOSULFOKINE RECEPTOR 1 modulates the defense response in tomato. Plant Physiol. 2023, 192, 2507–2522. [Google Scholar] [CrossRef]
- Eun, C.-H.; Ko, S.-M.; Matsubayashi, Y.; Sakagami, Y.; Kamada, H. Phytosulfokine-α requires auxin to stimulate carrot non-embryogenic cell proliferation. Plant Physiol. Biochem. 2003, 41, 447–452. [Google Scholar] [CrossRef]
- Yamakawa, S.; Sakuta, C.; Matsubayashi, Y.; Sakagami, Y.; Kamada, H.; Satoh, S. The promotive effects of a peptidyl plant growth factor, phytosulfokine-α, on the formation of adventitious roots and expression of a gene for a root-specific cystatin in cucumber hypocotyls. J. Plant Res. 1998, 111, 453–458. [Google Scholar] [CrossRef]
- Igasaki, T.; Akashi, N.; Ujino-Ihara, T.; Matsubayashi, Y.; Sakagami, Y.; Shinohara, K. Phytosulfokine stimulates somatic embryogenesis in Cryptomeria japonica. Plant Cell Physiol. 2003, 44, 1412–1416. [Google Scholar] [CrossRef]
- Hao, Z.; Wu, H.; Zheng, R.; Li, R.; Zhu, Z.; Chen, Y.; Lu, Y.; Cheng, T.; Shi, J.; Chen, J. The plant peptide hormone phytosulfokine promotes somatic embryogenesis by maintaining redox homeostasis in Cunninghamia lanceolata. Plant J. 2023, 113, 716–733. [Google Scholar] [CrossRef] [PubMed]
- Stahl, Y.; Simon, R. Plant stem cell niches. Int. J. Dev. Biol. 2005, 49, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Kumpf, R.; Thorstensen, T.; Rahman, M.A.; Heyman, J.; Nenseth, H.Z.; Lammens, T.; Herrmann, U.; Swarup, R.; Veiseth, S.V.; Emberland, G.; et al. The ASH1-RELATED3 SET-domain protein controls cell division competence of the meristem and the quiescent center of the Arabidopsis primary root. Plant Physiol. 2014, 166, 632–643. [Google Scholar] [CrossRef]
- Rovere, F.D.; Fattorini, L.; Ronzan, M.; Falasca, G.; Altamura, M.M. The quiescent center and the stem cell niche in the adventitious roots of Arabidopsis thaliana. Plant Signal. Behav. 2016, 11, e1176660. [Google Scholar] [CrossRef] [PubMed]
- Mussig, C.; Shin, G.H.; Altmann, T. Brassinosteroids promote root growth in Arabidopsis. Plant Physiol. 2003, 133, 1261–1271. [Google Scholar] [CrossRef]
- Wildwater, M.; Campilho, A.; Perez-Perez, J.M.; Heidstra, R.; Blilou, I.; Korthout, H.; Chatterjee, J.; Mariconti, L.; Gruissem, W.; Scheres, B. The RETINOBLASTOMA-RELATED gene regulates stem cell maintenance in Arabidopsis roots. Cell 2005, 123, 1337–1349. [Google Scholar] [CrossRef]
- Hartmann, J.; Stuhrwohldt, N.; Dahlke, R.I.; Sauter, M. Phytosulfokine control of growth occurs in the epidermis, is likely to be non-cell autonomous and is dependent on brassinosteroids. Plant J. 2013, 73, 579–590. [Google Scholar] [CrossRef]
- Komori, R.; Amano, Y.; Ogawa-Ohnishi, M.; Matsubayashi, Y. Identification of tyrosylprotein sulfotransferase in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 15067–15072. [Google Scholar] [CrossRef]
- Han, J.; Tan, J.; Tu, L.; Zhang, X. A peptide hormone gene, GhPSK promotes fibre elongation and contributes to longer and finer cotton fibre. Plant Biotechnol. J. 2014, 12, 861–871. [Google Scholar] [CrossRef]
- Geng, Y.; Jian, C.; Xu, W.; Liu, H.; Hao, C.; Hou, J.; Liu, H.; Zhang, X.; Li, T. miR164-targeted TaPSK5 encodes a phytosulfokine precursor that regulates root growth and yield traits in common wheat (Triticum aestivum L.). Plant Mol. Biol. 2020, 104, 615–628. [Google Scholar] [CrossRef]
- Yu, L.; Liu, Y.; Zeng, S.; Yan, J.; Wang, E.; Luo, L. Expression of a novel PSK-encoding gene from soybean improves seed growth and yield in transgenic plants. Planta 2019, 249, 1239–1250. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.F.; Matsubayashi, Y.; Sakagami, Y. Peptide growth factor phytosulfokine-α contributes to the pollen population effect. Planta 2000, 211, 752–755. [Google Scholar] [CrossRef]
- Tsuwamoto, R.; Fukuoka, H.; Takahata, Y. Identification and characterization of genes expressed in early embryogenesis from microspores of Brassica napus. Planta 2007, 225, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Stuhrwohldt, N.; Dahlke, R.I.; Kutschmar, A.; Peng, X.; Sun, M.X.; Sauter, M. Phytosulfokine peptide signaling controls pollen tube growth and funicular pollen tube guidance in Arabidopsis thaliana. Physiol. Plantarum 2015, 153, 643–653. [Google Scholar] [CrossRef]
- Kou, X.; Liu, Q.; Sun, Y.; Wang, P.; Zhang, S.; Wu, J. The peptide PbrPSK2 from phytosulfokine family induces reactive oxygen species (ROS) production to regulate pear pollen tube growth. Front. Plant Sci. 2020, 11, 601993. [Google Scholar] [CrossRef]
- Kiełkowska, A.; Adamus, A. Peptide growth factor phytosulfokine-α stimulates cell divisions and enhances regeneration from B. oleracea var. capitata L. protoplast culture. J. Plant Growth Regul. 2018, 38, 931–944. [Google Scholar] [CrossRef]
- Matsubayashi, Y.; Goto, T.; Sakagami, Y. Chemical nursing: Phytosulfokine improves genetic transformation efficiency by promoting the proliferation of surviving cells on selective media. Plant Cell Rep. 2004, 23, 155–158. [Google Scholar] [CrossRef]
- Yamakawa, S.; Matsubayashi, Y.; Sakagami, Y.; Kamada, H.; Satoh, S. Promotive effects of the peptidyl plant growth factor, phytosulfokine-α, on the growth and chlorophyll content of Arabidopsis seedlings under high night-time temperature conditions. Biosci. Biotechnol. Biochem. 1999, 63, 2240–2243. [Google Scholar] [CrossRef]
- Nagar, P.; Kumar, A.; Jain, M.; Kumari, S.; Mustafiz, A. Genome-wide analysis and transcript profiling of PSKR gene family members in Oryza sativa. PLoS ONE 2020, 15, e0236349. [Google Scholar] [CrossRef] [PubMed]
- Reichardt, S.; Piepho, H.P.; Stintzi, A.; Schaller, A. Peptide signaling for drought-induced tomato flower drop. Science 2020, 367, 1482–1485. [Google Scholar] [CrossRef] [PubMed]
- Soleimani Aghdam, M.; Sayyari, M.; Luo, Z. Exogenous phytosulfokine α application delays senescence and promotes antioxidant nutrient accumulation in strawberry fruit during cold storage by triggering endogenous phytosulfokine α signaling. Postharvest Biol. Technol. 2021, 175, 111473. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Ebrahimi, A.; Sheikh-Assadi, M. Phytosulfokine α (PSKα) delays senescence and reinforces SUMO1/SUMO E3 ligase SIZ1 signaling pathway in cut rose flowers (Rosa hybrida cv. Angelina). Sci. Rep. 2021, 11, 23227. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Flores, F.B. Employing phytosulfokine α (PSKα) for delaying broccoli florets yellowing during cold storage. Food Chem. 2021, 355, 129626. [Google Scholar] [CrossRef]
- Wang, D.; Huang, H.; Jiang, Y.; Duan, X.; Lin, X.; Soleimani Aghdam, M.; Luo, Z. Exogenous phytosulfokine α (PSKα) alleviates chilling injury of banana by modulating metabolisms of nitric oxide, polyamine, proline, and gamma-aminobutyric acid. Food Chem. 2022, 380, 132179. [Google Scholar] [CrossRef] [PubMed]
- Jiao, C. PpCBF6 is involved in phytosulfokine α-retarded chilling injury by suppressing the expression of PpLOX5 in peach fruit. Front. Plant Sci. 2022, 13, 874338. [Google Scholar] [CrossRef] [PubMed]
- Mosher, S.; Seybold, H.; Rodriguez, P.; Stahl, M.; Davies, K.A.; Dayaratne, S.; Morillo, S.A.; Wierzba, M.; Favery, B.; Keller, H.; et al. The tyrosine-sulfated peptide receptors PSKR1 and PSY1R modify the immunity of Arabidopsis to biotrophic and necrotrophic pathogens in an antagonistic manner. Plant J. 2013, 73, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Hu, Z.; Lei, C.; Zheng, C.; Wang, J.; Shao, S.; Li, X.; Xia, X.; Cai, X.; Zhou, J.; et al. A plant phytosulfokine peptide initiates auxin-dependent immunity through cytosolic Ca2+ signaling in tomato. Plant Cell 2018, 30, 652–667. [Google Scholar] [CrossRef]
- Loivamaki, M.; Stuhrwohldt, N.; Deeken, R.; Steffens, B.; Roitsch, T.; Hedrich, R.; Sauter, M. A role for PSK signaling in wounding and microbial interactions in Arabidopsis. Physiol. Plant 2010, 139, 348–357. [Google Scholar] [CrossRef]
- Yang, W.; Zhang, B.; Qi, G.; Shang, L.; Liu, H.; Ding, X.; Chu, Z. Identification of the phytosulfokine receptor 1 (OsPSKR1) confers resistance to bacterial leaf streak in rice. Planta 2019, 250, 1603–1612. [Google Scholar] [CrossRef]
- Igarashi, D.; Tsuda, K.; Katagiri, F. The peptide growth factor, phytosulfokine, attenuates pattern-triggered immunity. Plant J. 2012, 71, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Grzebelus, E.; Szklarczyk, M.; Greń, J.; Śniegowska, K.; Jopek, M.; Kacińska, I.; Mrożek, K. Phytosulfokine stimulates cell divisions in sugar beet (Beta vulgaris L.) mesophyll protoplast cultures. Plant Growth Regul. 2012, 67, 93–100. [Google Scholar] [CrossRef]
- Yasuda, M.; Ishikawa, A.; Jikumaru, Y.; Seki, M.; Umezawa, T.; Asami, T.; Maruyama-Nakashita, A.; Kudo, T.; Shinozaki, K.; Yoshida, S.; et al. Antagonistic interaction between systemic acquired resistance and the abscisic acid-mediated abiotic stress response in Arabidopsis. Plant Cell 2008, 20, 1678–1692. [Google Scholar] [CrossRef] [PubMed]
- Boller, T.; Felix, G. A renaissance of elicitors: Perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu. Rev. Plant Biol. 2009, 60, 379–406. [Google Scholar] [CrossRef]
- Murata, M.; Orton, T.J. Callus initiation and regeneration capacities in Brassica species. Plant Cell Tissue Organ Cult. 1987, 11, 111–123. [Google Scholar] [CrossRef]
- Farooq, N.; Nawaz, M.A.; Mukhtar, Z.; Ali, I.; Hundleby, P.; Ahmad, N. Investigating the in vitro regeneration potential of commercial cultivars of Brassica. Plants 2019, 8, 558. [Google Scholar] [CrossRef]
- Dai, C.; Li, Y.; Li, L.; Du, Z.; Lin, S.; Tian, X.; Li, S.; Yang, B.; Yao, W.; Wang, J.; et al. An efficient Agrobacterium-mediated transformation method using hypocotyl as explants for Brassica napus. Mol. Breed. 2020, 40, 96. [Google Scholar] [CrossRef]
- Maheshwari, P.; Selvaraj, G.; Kovalchuk, I. Optimization of Brassica napus (canola) explant regeneration for genetic transformation. New Biotechnol. 2011, 29, 144–155. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, X.; Stührwohldt, N.; Lin, C. The Research Process of PSK Biosynthesis, Signaling Transduction, and Potential Applications in Brassica napus. Plants 2023, 12, 3075. https://doi.org/10.3390/plants12173075
Shen X, Stührwohldt N, Lin C. The Research Process of PSK Biosynthesis, Signaling Transduction, and Potential Applications in Brassica napus. Plants. 2023; 12(17):3075. https://doi.org/10.3390/plants12173075
Chicago/Turabian StyleShen, Xuwen, Nils Stührwohldt, and Chen Lin. 2023. "The Research Process of PSK Biosynthesis, Signaling Transduction, and Potential Applications in Brassica napus" Plants 12, no. 17: 3075. https://doi.org/10.3390/plants12173075
APA StyleShen, X., Stührwohldt, N., & Lin, C. (2023). The Research Process of PSK Biosynthesis, Signaling Transduction, and Potential Applications in Brassica napus. Plants, 12(17), 3075. https://doi.org/10.3390/plants12173075