

Plant Extracts from the Yucatan Peninsula in the In Vitro Control of Curvularia lunata and Antifungal Effect of Mosannona depressa and Piper neesianum Extracts on Postharvest Fruits of Habanero Pepper

 , , and
, , and
Abstract
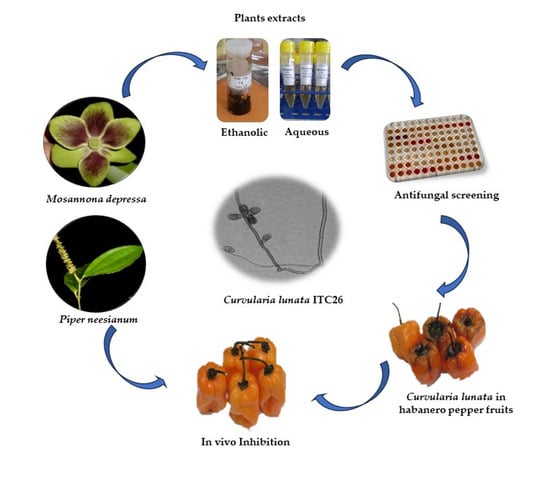
1. Introduction
2. Results
2.1. In Vitro Activity of Plant Extracts on Curvularia lunata
2.2. Determination of Minimum Inhibitory Concentration of Extracts, Fractions, and α-Asarone
2.3. Inhibitory Concentration (IC50 and IC95) of the Most Active Extracts, Fractions, and α-Asarone
2.4. Effect of Active Extracts on Curvularia lunata
2.5. In Vivo Effect of Extracts and α-Asarone against C. lunata on Habanero Pepper Fruits
3. Discussion
4. Materials and Methods
4.1. Plant Collection and Processing
4.2. Plant Extracts and Partition of Active Extracts
4.3. Fungal Cultures
4.4. Antifungal Microdilution Assay of Extracts
4.4.1. Preparation of Conidial Suspension
4.4.2. Bioassay with Aqueous Extracts
4.4.3. Bioassays with Ethanolic Extracts
4.4.4. Minimum Inhibitory Concentration
4.5. Evaluation of Ethanolic Extracts on Hyphal Morphology of Curvularia lunata ITC26
4.6. In Vivo Evaluation of Ethanolic Extracts and α-Asarona against C. lunata on Habanero Pepper Fruits
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Servicio de Información Agroalimentaria y Pesquera (SIAP). Panorama Agroalimentario. 2020, 62. Available online: https://nube.siap.gob.mx/gobmx_publicaciones_siap/pag/2020/Atlas-Agroalimentario-2020 (accessed on 15 April 2023).
- Anuario Estadístico de la Producción Agrícola SIAP. 2022. Available online: https://nube.siap.gob.mx/cierreagricola/ (accessed on 21 June 2023).
- Naves, E.R.; de Ávila Silva, L.; Sulpice, R.; kmmmAraújo, W.L.; Nunes-Nesi, A.; Pérez, L.E.; Zsögön, A. Capsaicinoids: Pungency beyond Capsicum. Trends. Plant. Sci. 2019, 24, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Mongkolporn, O.; Taylor, P.W.J. Chili anthracnose: Colletotrichum taxonomy and pathogenicity. Plant Pathol. 2018, 67, 1255–1263. [Google Scholar] [CrossRef]
- Ali, A.; Bordoh, P.K.; Singh, A.; Siddiqui, Y.; Droby, S. Post-harvest development of anthracnose in pepper (Capsicum spp.): Etiology and management strategies. Crop Prot. 2016, 90, 132–141. [Google Scholar] [CrossRef]
- Oo, M.M.; Lim, G.T.; Jang, H.A.; Oh, S.O. Characterization and pathogenicity of new record of anthracnose on various chili varieties caused by Colletotrichum scovillei in Korea. Mycobiology 2017, 45, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y.L.; Tao, S.; Sun, Y.F.; Feng, T.Z.; Long, H.B. First report of Capsicum frutescens leaf spot caused by Curvularia lunata in China. Plant Dis. 2018, 102, 241. [Google Scholar] [CrossRef]
- Menaria, D.; Rawal, P.; Mathur, K.; Saharan, V. Management of fruit rot (Curvularia lunata) of bell pepper. J. Mycol. Plant Pathol. 2012, 42, 317–320. [Google Scholar]
- Hoil-Cocom, P.A. Actividad Inhibitoria del Extracto Acuoso de Bonellia flammea Contra Hongos Postcosecha de Capsicum spp. Master’s Thesis, Instituto Tecnológico de Conkal, Mérida, Mexico, 2016; p. 49. [Google Scholar]
- Tun-Suárez, J.M.; Castillo Peraza, M.E.; Cristóbal Alejo, J.; Latournerie Moreno, L. Etiología de la mancha foliar del chile dulce (Capsicum annuum L.) y su control in vitro en Yucatán, México. Fitosanidad 2011, 15, 5–10. [Google Scholar]
- Cristóbal-Alejo, J.; Meléndez, E.Z.; Tun-Suárez, J.M.; Moreno, L.L.; Sánchez, E.R. Control químico y epidemiología de la mancha foliar del chile habanero (Capsicum chinense Jacq.) en Yucatán, México. Fitosanidad 2006, 10, 217–220. [Google Scholar]
- Cristóbal-Alejo, J.; Navarrete-Mapen, Z.; Herrera-Parra, E.; Mis-Mut, M.; Tun-Suárez, J.M.; Ruiz-Sánchez, E. Hifomicetos asociados a plantas tropicales del estado de Yucatán, México: Identificación genérica y evaluación de fungicidas para su control. Rev. Protección Veg. 2013, 28, 138–144. [Google Scholar]
- Khan, I.H.; Javaid, A. First report of Curvularia lunata causing postharvest fruit rot of banana in Pakistan. Int. J. Agric. Biol. 2020, 24, 1621–1624. [Google Scholar] [CrossRef]
- Zhou, H.K.; Liu, Y.L.; Tang, J.R.; Zhong, F.T.; Li, Y. First report of leaf spot caused by Curvularia lunata on wild rice in China. Plant Dis. 2021, 105, 3300. [Google Scholar] [CrossRef]
- Pandey, R.K.; Gupta, P.K.; Srivastava, M.; Singh, S.R.; Robin, G. First report of brown leaf spot disease caused by Curvularia lunata infecting Indian spinach or poi (Basella rubra). Indian Phytopathol. 2011, 64, 207. [Google Scholar]
- Abubakar, M.N.; Likita, M.S. Effect of combined fungicides on the mycelial growth of Curvularia lunata. Int. Innov. Agric. Biol. Res. 2021, 9, 1–9. [Google Scholar]
- Sharma, A.B.; SarbjitKaur; Lore, J.S. Prevalence of pathogens causing sheath rot of rice in North India and its management. Ind. Phytopathol. 2023, 1–15. [Google Scholar] [CrossRef]
- Beckerman, J.; Palmer, C.; Tedford, E.; Ypema, H. Fifty years of fungicide development, deployment, and future use. Phytopathology 2023, 113, 694–706. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, F.P. Pesticides, environment, and food safety. Food Energ. Sec. 2017, 6, 48–60. [Google Scholar] [CrossRef]
- Gaherwal, S.; Prakash, M.M.; Khasdeo, K.; Sharma, A. Impact of selected chemical and herbal pesticide on beneficial soil microorganism. Int. J. Microbiol. Res. 2015, 6, 236–239. Available online: https://idosi.org/ijmr/ijmr6(3)15/12.pdf (accessed on 15 June 2023).
- Shuping, D.S.S.; Eloff, J.N. The use of plants to protect plants and food against fungal pathogens: A review. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Sparks, T.C.; Sparks, J.M.; Duke, S.O. Natural product-based crop protection compounds─ origins and future prospects. J. Agric. Food Chem. 2023, 71, 2259–2269. [Google Scholar] [CrossRef]
- Jiménez-Reyes, M.F.; Carrasco, H.; Olea, A.F.; Silva-Moreno, E. Natural compounds: A sustainable alternative to the phytopathogens control. J. Chil. Chem. Soc. 2019, 64, 4459–4465. [Google Scholar] [CrossRef]
- Hussain, M.A.; Pandey, A.; Chandra, R. Lantana camara against Curvularia lunata, a common pathogen of Capsicum frutescens. Life Sci. Bull. 2009, 6, 289–290. [Google Scholar]
- Kumaran, R.S.; Gomathi, V.; Kannabiran, B. Fungitoxic effects of root extracts of certain plant species on Colletotrichum capsici causing anthracnose in Capsicum annuum. Indian Phytopathol. 2003, 56, 114–116. [Google Scholar]
- Moo-Koh, F.A.; Cristóbal-Alejo, J.; Tun-Suárez, J.M.; Medina-Baizabal, I.L.; Arjona-Cruz, A.A.; Gamboa-Angulo, M. Activity of aqueous extracts from native plants of the Yucatan Peninsula against fungal pathogens of tomato in Vitro and from Croton chichenensis against Corynespora cassiicola on Tomato. Plants 2022, 11, 2821. [Google Scholar] [CrossRef] [PubMed]
- Moo-Koh, F.A.; Cristóbal Alejo, J.; Reyes-Ramírez, A.; Tun-Suárez, J.M.; Sandoval-Luna, R.; Ramírez-Pool, J.A. Actividad in vitro del extracto acuoso del Bonellia flammea contra hongos fitopatógenos. Agrociencia 2014, 48, 833–845. [Google Scholar]
- Vargas-Díaz, A.A.; Gamboa Angulo, M.; Medina Baizabal, I.L.; Pérez Brito, D.; Cristóbal Alejo, J.; Ruiz Sánchez, E. Evaluation of native Yucatecan plant extracts against Alternaria chrysanthemi and antifungal spectrum of Acalypha gaumeri. Rev. Mex. Fitopatol. 2014, 32, 1–11. [Google Scholar]
- Cruz-Cerino, P.; Cristóbal-Alejo, J.; Ruiz-Carrera, V.; Carnevali, G.; Vera-Ku, M.; Martín, J.; Gamboa-Angulo, M. Extracts from six native plants of the Yucatán Peninsula hinder mycelial growth of Fusarium equiseti and F. oxysporum, pathogens of Capsicum chinense. Pathogens 2020, 9, 827. [Google Scholar] [CrossRef] [PubMed]
- Zambrano Orozco, L.M. Evaluación, Diagnóstico y Biocontrol de Fitopatógenos de Capsicum frutescens (Solanaceae) en Gauacarí Valle del Cauca. Undergraduate Thesis, Universidad del valle, Cali, Colombia, 2016. Available online: http://hdl.handle.net/10893/9974 (accessed on 14 June 2023).
- Pei, Y.; Sun, Y.; Feng, T.; Chen, Y.; Long, H. Pathogen identification for the new leaf spot disease of Capsicum annuum L. and its biological characteristics. Chin. J. Trop. Crops CJTC 2021, 42, 2659–2665. [Google Scholar] [CrossRef]
- Pearson, M.N.; Bull, P.B.; Speke, H. Anthracnose of capsicum in Papua New Guinea; varietal reaction and associated fungi. Int. J. Pest Manag. 1984, 30, 230–233. [Google Scholar] [CrossRef]
- Pérez-Chablé, C.L. Aislamiento e identificación de hongos en semillas de Capsicum spp. y su sensibilidad in vitro a extractos vegetales. Master’s Thesis, Instituto Tecnológico de Conkal, Conkal, Mexico, 2019; p. 49. [Google Scholar]
- Aviles-Gomez, J.; Cristóbal-Alejo, J.; Andrés, M.F.; González-Coloma, A.; Carnevali, G.; Pérez-Brito, D.; Gamboa-Angulo, M. Nematicidal screening of aqueous extracts from plants of the Yucatan Peninsula and ecotoxicity. Plants 2022, 11, 2138. [Google Scholar] [CrossRef]
- Esquivel-Chi, M.; Ruiz-Sánchez, E.; Ballina-Gómez, H.S.; Martín, J.; Reyes, F.; Carnevali, G.; Tapia-Muñoz, J.L.; Gamboa-Angulo, M. Repellent screening of ethanol extracts from plants of the Yucatan Peninsula against Bemisia tabaci and chemical profile of Malpighia glabra leaves. Ind. Crops Prod. 2023; submitted. [Google Scholar]
- Barupal, T.; Sharma, K. Effect of leaf extracts of Lawsonia Inermis Linn. On Curvularia lunata, caused leaf spot disease of maize. Int. J. Innov. Res. Adv. Stud. 2017, 4, 64–67. [Google Scholar]
- Gupta, S.P.; Rana, K.S.; Sharma, K.; Chhabra, B.S. Antifungal activity of aqueous leaf extract of Ocimum sanctum on dominant fungal species of monuments. Eur. Chem. Bull. 2014, 2, 609–611. [Google Scholar] [CrossRef]
- Sailaja, I. Antifungal activity of some wild plant extracts against fungal pathogens. Int. J. Intg. Med. Sci. 2014, 1, 41–44. [Google Scholar]
- Saha, A.; Rahman, M.S. Antimicrobial activity of crude extract from Calycopteris floribunsa. Bangladesh J. Microbiol. 2008, 25, 137–139. [Google Scholar] [CrossRef]
- Duraipandiyan, V.; Abdullah Al-Harbi, N.; Ignacimuthu, S.; Muthukumar, C. Antimicrobial activity of sesquiterpene lactones isolated from traditional medicinal plant, Costus speciosus (Koen ex. Retz.) Sm. BMC Complement. Altern. Med. 2012, 12, 13. [Google Scholar] [CrossRef] [PubMed]
- Singha, I.M.; Unni, B.G.; Kakoty, Y.; Das, J.; Wann, S.B.; Singh, L.; Kalita, M.C. Evaluation of in vitro antifungal activity of medicinal plants against phytopathogenic fungi. Arch. Phytopathol. 2011, 44, 1033–1040. [Google Scholar] [CrossRef]
- Begum, J.; Yusuf, M.; Chowdhury, J.U.; Khan, S.; Anwar, M.N. Antifungal activity of forty higher plants against phytopathogenic fungi. Bangladesh J. Microbiol. 2007, 24, 76–78. [Google Scholar] [CrossRef]
- Mathew, S.; Raju, R.; Zhou, X.; Bodkin, F.; Govindaraghavan, S.; Münch, G. A Method and formula for the quantitative analysis of the total bioactivity of natural products. Int. J. Mol. Sci. 2023, 24, 6850. [Google Scholar] [CrossRef]
- Ju, J.; Xie, Y.; Yu, H.; Guo, Y.; Cheng, Y.; Qian, H.; Yao, W. Analysis of the synergistic antifungal mechanism of eugenol and citral. LWT Food. Sci. Technol. 2020, 123, 109128. [Google Scholar] [CrossRef]
- Rajput, S.B.; Karuppayil, S.M. Small molecules inhibit growth, viability and ergosterol biosynthesis in Candida albicans. SpringerPlus 2013, 2, 26. [Google Scholar] [CrossRef]
- FMC Launches Fracture Fungicide. (n.d.). Fruit Growers News. Available online: https://fruitgrowersnews.com/news/fmc-launches-fracture-fungicide/ (accessed on 28 July 2023).
- Burg, J.S.; Espenshade, P.J. Regulation of HMG-CoA reductase in mammals andyeast. Prog. Lipid Res. 2011, 50, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Pavón, D.; Ortiz-Álvarez, J.; Sánchez-Sandoval, E.; Tamariz, J.; Hernández-Rodríguez, C.; Ibarra, J.A.; Villa-Tanaca, L. Inhibition of recombinant enzyme 3-hydroxy-3-methylglutaryl-CoA reductase from Candida glabrata by α-asarone-based synthetic compounds as antifungal agents. J. Biotechnol. 2019, 292, 64–67. [Google Scholar] [CrossRef]
- Mejía, R. Guatteria gaumeri, Malmea depressa o Yumel, una revisión sobre su historia, sus propiedades y su uso en la homeopatía. Homeopatia Méx. 2016, 85, 28–38. [Google Scholar]
- Fort, R.S.; Barnech, T.J.M.; Dourron, J.; Colazzo, M.; Aguirre-Crespo, F.J.; Duhagon, M.A.; Álvarez, G. Isolation and structural characterization of bioactive molecules on prostate cancer from Mayan traditional medicinal plants. Pharmaceuticals 2018, 11, 78. [Google Scholar] [CrossRef] [PubMed]
- Husain, A.; Indani, A.; Bhutada, P. Hypercholesterolemia effectively managed with homoeopathic medicine Gautteria gaumeri (Yumel): Results from a clinical study in academic clinical set up in north India. Int. J. Adv. Med. 2017, 4, 772. [Google Scholar] [CrossRef]
- Godoy-Rodríguez, T. Actividad Antifúngica de Plantas Nativas de la Península de Yucatán Para el Control de Fitopatógenos Poscosecha de Capsicum spp. Master’s Thesis, Centro de Investigación Científica de Yucatán, Mérida, Mexico, 2019. [Google Scholar]
- Jimenez-Arellanes, A.; Mata, R.; Lotina-Henssen, B.; Lang, A.L.A.; Ibarra, L.V. Phytogrowth-inhibitory compounds from Malmea depressa. J. Nat. Prod. 1996, 59, 202–204. [Google Scholar] [CrossRef]
- Windy, T.; Rahayu, I.; Timotius, K.H. Antimicrobial and antihelmintic activities of asarone rich herbal materials: A review. Int. J. Herb. Med. 2021, 9, 1–7. [Google Scholar]
- Begum, J.; Sohrab, H.; Yusuf, M.; Chowdury, J.U. In vitro antifungal activity of azaron isolated from the rhizome extract of Acorus calamus L. Pak. J. Biol. Sci. 2004, 7, 1376–1379. [Google Scholar] [CrossRef]
- Chen, Y.; Li, J.; Li, S.X.; Zhao, J.; Bernier, U.R.; Becnel, J.J.; Agramonte, N.M.; Duke, S.O.; Cantrell, C.L.; Wedge, D.E. Identification and characterization of biopesticides from Acorus tatarinowii and A. calamus. In Medicinal and Aromatic Crops: Production, Phytochemistry and Utilization; American Chemical Society Symposium Series; American Chemical Society: Washington, DC, USA, 2016; Volume 3, pp. 121–143. [Google Scholar] [CrossRef]
- Momin, R.A.; Nair, M.G. Pest-managing efficacy of trans-asarone isolated from Daucus carota L. seeds. J. Agric. Food Chem. 2002, 50, 4475–4478. [Google Scholar] [CrossRef]
- Perrett, S.; Whitfield, P.J. Anthelmintic and pesticidal activity of Acorus gramineus (Araceae) is associated with phenylpropanoid asarones. Phytother. Res. 1995, 9, 405–409. [Google Scholar] [CrossRef]
- Enríquez, R.G.; Chávez, M.A.; Jauregui, F. Propenylbenzenes from Guatteria gaumeri. Phytochemistry 1980, 19, 2024–2025. [Google Scholar] [CrossRef]
- Suriani, N.L.; Suprapta, D.N.; Nazir, N.; Darmadi, A.A.K.; Parwanayoni, N.M.S.; Sudatri, N.W.; Yamin, B.M. Inhibitory activity of Piper caninum leaf extract against Curvularia spotting disease on rice plants. Indian J. Agric. Res. 2020, 54, 411–419. [Google Scholar] [CrossRef]
- Suriani, N.L.; Suprapta, D.N.; Suarsana, I.N.; Reddy, M.S.; Reddy, M.S.; Gunawan, S.; Herlambang, S.; Resiani, N.M.D.; Pratiwi, E.; Sabullah, M.K.; et al. Piper caninum extract and Brevibacillus agri mixture suppresses rice leaf spot pathogen; Nigrospora oryzae and improves the production of red rice (Oryza sativa L). Front. Sustain. Food Syst. 2022, 6, 1080481. [Google Scholar] [CrossRef]
- Cruz, S.; Cáceres, A.; Álvarez, L.; Apel, M.; Henríquez, A. Chemical diversity of essential oils from 15 Piper species from Guatemala. Acta Hortic. 2012, 964, 39–46. [Google Scholar] [CrossRef]
- Abou-Jawdah, Y.; Sobh, H.; Salameh, A. Antimycotic activities of selected plant flora, growing wild in Lebanon, against phytopathogenic fungi. J. Agric. Food Chem. 2002, 50, 3208–3213. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi. Approved Standard M38-A; National Committee for Clinical Laboratory Standards: Wayne, PA, USA, 2002. [Google Scholar]
- Irkin, R.; Korukluoglu, M. Control of Aspergillus niger with garlic, onion and leek extracts. Afr. J. Biotechnol. 2007, 6, 384–387. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Plant Species | Concentration | Mycelial Growth Inhibition (%) | |||
|---|---|---|---|---|---|
| Curvularia lunata ITC26 | |||||
| L | S | R | C | ||
| Ethanol (µg/mL) | |||||
| Alvaradoa amorphoides | 2000 | 75 b | 0 c | 0 d | |
| Helicteres baruensis | 2000 | 75 b | 25 b | 0 d | |
| Licaria sp. | 2000 | 0 d | 0 c | 75 b | |
| Mosannona depressa | 2000 | 0 d | 100 a | 100 a | |
| Piper neesianum | 2000 | 100 a | 0 c | 75 b | |
| Aqueous (% w/v) | |||||
| Byrsonima bucidifolia | 3 | 25 c | 25 b | 25 c | |
| Morella cerifera | 3 | 25 c | 0 c | 25 c | |
| RPMI | Negative C | 0 b | |||
| blank | 0 b | ||||
| Prochloraz 0.11% | Positive C | 100 a | |||
| Solvent | Extract/ Fraction | Curvularia lunata | ||||
|---|---|---|---|---|---|---|
| Concentration (µg/mL) | ||||||
| 1000 | 500 | 250 | 125 | MIC | ||
| Ethanol | MDT | 100 a | 100 a | 100 a | 0 c | 250++ |
| Hexane | MDT-a | 50 b | 0 e | 0 g | 0 c | >1000 |
| Acetonitrile | MDT-b | 100 a | 75 c | 25 f | 0 c | 1000+ |
| Precipitate | MDT-c | 100 a | 50 d | 0 c | 0 c | 1000+ |
| Ethanol | MDR | 100 a | 100 a | 100 a | 0 c | 250++ |
| Hexane | MDR-a | 50 b | 25 d | 0 g | 0 c | >1000 |
| Acetonitrile | MDR-b | 100 a | 83 b | 50 e | 0 c | 1000+ |
| Precipitate | MDR-c | 100 a | 100 a | 83 c | 0 c | 500+ |
| Ethanol | PNH | 100 a | 100 a | 0 g | 0 c | 500+ |
| Hexane | PNH-a | 25 c | 25 d | 0 g | 0 c | >1000 |
| Acetonitrile | PNH-b | 100 a | 100 a | 91 b | 0 c | 500+ |
| Precipitate | PNH-c | 100 a | 0 e | 0 g | 0 c | 1000+ |
| CS | α-Asarone | ne | 100 a | 75 d | 25 b | 500+ |
| NC | 0 d | 0 e | 0 g | 0 c | ||
| PC | 100 a | 100 a | 100 a | 100 a | ||
| Source | Curvularia lunata | ||
|---|---|---|---|
| Extract/Fraction | IC50 (CI) | IC95 (CI) | |
| M. depressa | MDT | 188 (42–308) | 218 (81–342) |
| MDT-b | 388 (298–528) | 627 (449–1022) | |
| MDT-c | 388 (298–528) | 627 (449–1022) | |
| MDR | 188 (42–308) | 218 (81–342) | |
| MDR-c | 229 (210–450) | 265 (241–465) | |
| α-asarone | CS | 190 (130–271) | 325 (253–631) |
| P. neesianum | PNH | 378 (278–490) | 428 (350–600) |
| PNH-b | 222 (205–470) | 256 (205–404) | |
| Extract | Treatment | Concentration µg/mL | Severity (%) | Effectiveness (%) |
|---|---|---|---|---|
| MDT | T1 | 750 | 0 e | 100 a |
| T2 | 500 | 0 e | 100 a | |
| T3 | 250 | 3.4 cd | 93.2 bcd | |
| PNH | T4 | 750 | 0 e | 100 a |
| T5 | 500 | 1.5 d | 97 bc | |
| T6 | 250 | 8 cb | 84 c | |
| α-asarona | T7 | 500 | 0 e | 100 a |
| T8 | 250 | 2.9 d | 94.2 bc | |
| T9 | 125 | 8 cb | 84 c | |
| PC | T10 | 450 | 0 e | 100 a |
| NC | T11 | - | 100 a | 0 e |
| solvent | T12 | - | 0 e | 0 e |
| Family | Species | Site | Voucher | Plant Parts |
|---|---|---|---|---|
| Rubiaceae | Alseis yucatanensis Standl. | 1 | JLT-3179 | L |
| Simaroubaceae | Alvaradoa amorphoides Liebm. | 2 | GC-8236 | L, S, R |
| Annonaceae | Annona primigenia Standl. and Steyerm | 2 | GC-8057 | L, SB |
| Malvaceae | Bakeridesia notolophium (A. Gray) Hochr. | 3 | RD-s/n | L, S |
| Acanthaceae | Bravaisia berlandieriana (Nees) T.F.Daniel | 4 | GC-8168 | L, S, R |
| Malpighiaceae | Byrsonima bucidifolia Standl. | 2 | GC-8087 | L, S, R |
| Asteraceae | Calea jamaicensis (L.) L. | 2 | GC-8084 | WP |
| Apocynaceae | Cameraria latifolia L. | 2 | JLT-1165 | L, SB, R |
| Sapotaceae | Chrysophyllum mexicanum Brandegee ex Standl. | 2 | GC-8082 | L, S, R |
| Polygonaceae | Coccoloba sp. | 5 | GC-8258 | L, S |
| Euphorbiaceae | Croton arboreus Millsp. | 2 | JLT-1132 | L, S, R |
| Euphorbiaceae | Croton itzaeus Lundell | 2 | JLT-1138 | L, SB, RB |
| Euphorbiaceae | Croton sp. | 5 | GC-8262 | WP |
| Sapindaceae | Cupania sp. | 6 | GC-8009 | L, S |
| Ebenaceae | Diospyros sp. | 4 | GC-8147 | L |
| Erythroxylaceae | Erythroxylum confusum Britton | 2 | JLT-1143 | L, S, R |
| Erythroxylaceae | Erythroxylum rotundifolium Lunan | 2 | GC-8179 | L, S |
| Erythroxylaceae | Erythroxylum sp. | 4 | GC-8137 | L |
| Myrtaceae | Eugenia sp. | 4 | GC-8127 | L, S, R |
| Euphorbiaceae | Euphorbia armourii Millsp. | 1 | JLT-3182 | WP |
| Rubiaceae | Guettarda combsii Urb. | 2 | GC-8047 | L, SB, RB |
| Malvaceae | Helicteres baruensis Jacq. | 1 | GC-8127 | L, S, R |
| Malpighiaceae | Heteropterys laurifolia (L.) A. Juss. | 2 | GC-8035 | L, SB, R |
| Violaceae | Hybanthus yucatanensis Millsp. | 4 | GC-8158 | L, S |
| Convolvulaceae | Ipomoea clavata (G. Don) Ooststr. Ex J.F.Macbr. | 1 | JLT-3181 | WP |
| Rhamnaceae | Karwinskia humboldtiana (Willd. Ex Roem. and Schult.) Zucc. | 1 | JLT-3188 | L |
| Lauraceae | Licaria sp. | 2 | GC-8037 | L, SB, RB |
| Apocynaceae | Macroscepis diademata (Ker Gawl.) W.D. Stevens | 1 | JLT-3187 | L, SB |
| Malpighiaceae | Malpighia glabra L. | 4 | GC-8144 | L, S, R |
| Myricaceae | Morella cerifera (L.) Small. | 2 | JLT-1137 | L, S, RB |
| Annonaceae | Mosannona depressa (Ball.) Chatrou | 2 | GC-8085 | L, SB, RB |
| Primulaceae | Parathesis cubana (A. DC.) Molinet and M.Gómez | 2 | JLT-1133 | L, SB, RB |
| Sapindaceae | Paullinia sp. | 4 | GC-8106 | L, R |
| Piperaceae | Piper neesianum C.DC. | 2 | GC-8080 | L, S, R |
| Rubiaceae | Psychotria nervosa Sw. | 2 | GC-8086 | WP |
| Rubiaceae | Randia aculeata L. | 4 | GC-8156 | L, S, R |
| Sapindaceae | Serjania caracasana (Jacq.) Willd | 4 | GC-8114 | L, S, R |
| Simaroubaceae | Simarouba glauca DC. | 2 | GC-8081 | L, SB, RB |
| Apocynaceae | Stemmadenia donnell-smithii (Rose) Woodson | 2 | GC-8056 | L, SB |
| Passifloraceae | Turnera aromatica Arbo | 2 | GC-8081 | WP |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cruz-Cerino, P.; Cristóbal-Alejo, J.; Ruiz-Carrera, V.; Gamboa-Angulo, M. Plant Extracts from the Yucatan Peninsula in the In Vitro Control of Curvularia lunata and Antifungal Effect of Mosannona depressa and Piper neesianum Extracts on Postharvest Fruits of Habanero Pepper. Plants 2023, 12, 2908. https://doi.org/10.3390/plants12162908
Cruz-Cerino P, Cristóbal-Alejo J, Ruiz-Carrera V, Gamboa-Angulo M. Plant Extracts from the Yucatan Peninsula in the In Vitro Control of Curvularia lunata and Antifungal Effect of Mosannona depressa and Piper neesianum Extracts on Postharvest Fruits of Habanero Pepper. Plants. 2023; 12(16):2908. https://doi.org/10.3390/plants12162908
Chicago/Turabian StyleCruz-Cerino, Patricia, Jairo Cristóbal-Alejo, Violeta Ruiz-Carrera, and Marcela Gamboa-Angulo. 2023. "Plant Extracts from the Yucatan Peninsula in the In Vitro Control of Curvularia lunata and Antifungal Effect of Mosannona depressa and Piper neesianum Extracts on Postharvest Fruits of Habanero Pepper" Plants 12, no. 16: 2908. https://doi.org/10.3390/plants12162908
APA StyleCruz-Cerino, P., Cristóbal-Alejo, J., Ruiz-Carrera, V., & Gamboa-Angulo, M. (2023). Plant Extracts from the Yucatan Peninsula in the In Vitro Control of Curvularia lunata and Antifungal Effect of Mosannona depressa and Piper neesianum Extracts on Postharvest Fruits of Habanero Pepper. Plants, 12(16), 2908. https://doi.org/10.3390/plants12162908