
Plant Elicitor Peptide (Pep) Signaling and Pathogen Defense in Tomato


Abstract
1. Introduction
2. Biological Functions of Peps and Systemin
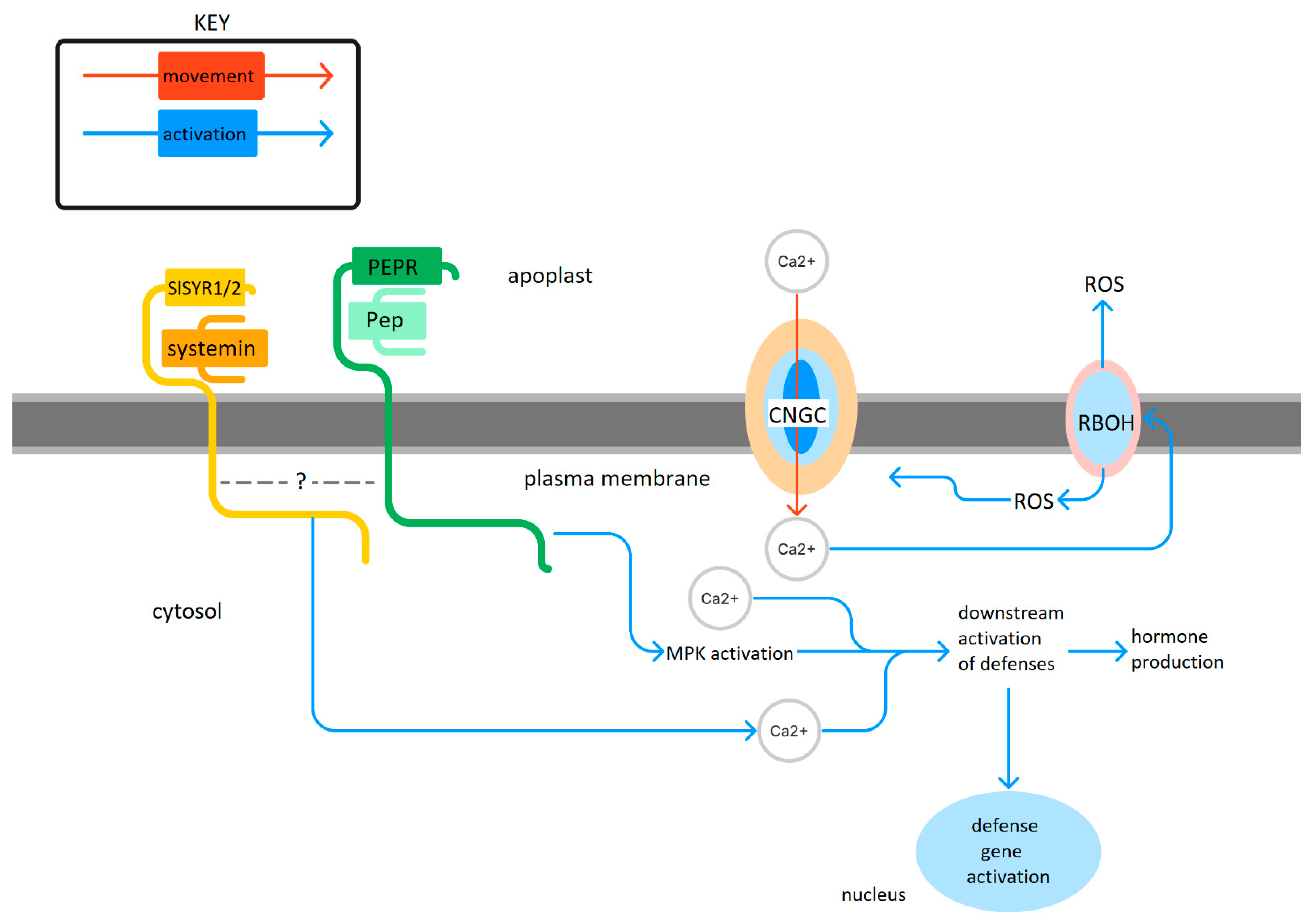
3. Local Responses to Peps
4. The Pep Receptor Identity Question
5. The Agricultural Context
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Badosa, E.; Planas, M.; Feliu, L.; Montesinos, L.; Bonaterra, A.; Montesinos, E. Synthetic Peptides against Plant Pathogenic Bacteria. Microorganisms 2022, 10, 1784. [Google Scholar] [CrossRef] [PubMed]
- Huot, B.; Yao, J.; Montgomery, B.L.; He, S.Y. Editor’s choice: Growth–Defense Tradeoffs in Plants: A Balancing Act to Optimize Fitness. Mol. Plant 2014, 7, 1267. [Google Scholar] [CrossRef] [PubMed]
- Figueroa-Macías, J.P.; García, Y.C.; Núñez, M.; Díaz, K.; Olea, A.F.; Espinoza, L. Plant growth-defense trade-offs: Molecular processes leading to physiological changes. Int. J. Mol. Sci. 2021, 22, 693. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.; Ougham, H.; Thomas, H.; Waaland, S. The Molecular Life of Plants; Wiley-Blackwell: Hoboken, NJ, USA, 2013. [Google Scholar]
- Ausubel, F.M. Are innate immune signaling pathways in plants and animals conserved? Nat. Immunol. 2005, 6, 973–979. [Google Scholar] [CrossRef]
- Maffei, M.E.; Arimura, G.I.; Mithöfer, A. Natural elicitors, effectors and modulators of plant responses. Nat. Prod. Rep. 2012, 29, 1288–1303. [Google Scholar] [CrossRef]
- Hogenhout, S.A.; Bos, J.I.B. Effector proteins that modulate plant–insect interactions. Curr. Opin. Plant Biol. 2011, 14, 422–428. [Google Scholar] [CrossRef]
- Basu, S.; Varsani, S.; Louis, J. Altering Plant Defenses: Herbivore-Associated Molecular Patterns and Effector Arsenal of Chewing Herbivores. MPMI 2017, 31, 13–21. [Google Scholar] [CrossRef]
- Ortiz-Morea, F.A.; Reyes-Bermudez, A.A. Bioactive molecules in plant defense: Signaling in growth and stress. In Bioactive Molecules in Plant Defense: Signaling in Growth and Stress; Jogaiah, S., Abdelrahman, M., Eds.; Springer: Cham, Switzerland, 2019. [Google Scholar] [CrossRef]
- Yang, X.; Deng, F.; Ramonell, K.M. Receptor-like kinases and receptor-like proteins: Keys to pathogen recognition and defense signaling in plant innate immunity. Front. Biol. 2012, 7, 155–166. [Google Scholar] [CrossRef]
- Zipfel, C. Early molecular events in PAMP-triggered immunity. Curr. Opin. Plant Biol. 2009, 12, 414–420. [Google Scholar] [CrossRef]
- Tanaka, K.; Heil, M. Damage-Associated Molecular Patterns (DAMPs ) in Plant Innate Immunity: Applying the Danger Model and Evolutionary Perspectives. Annu. Rev. Phytopathol. 2021, 59, 53–75. [Google Scholar] [CrossRef]
- Chen, Y.L.; Fan, K.T.; Hung, S.C.; Chen, Y.R. The role of peptides cleaved from protein precursors in eliciting plant stress reactions. New Phytol. 2020, 225, 2267–2282. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Huffaker, A. Endogenous peptide elicitors in higher plants. Curr. Opin. Plant Biol. 2011, 14, 351–357. [Google Scholar] [CrossRef]
- Klauser, D.; Desurmont, G.A.; Glauser, G.D.S.; Vallat, A.; Flury, P.; Boller, T.; Turlings, T.C.J.; Bartels, S. The Arabidopsis Pep-PEPR system is induced by herbivore feeding and contributes to JA-mediated plant defence against herbivory. J. Exp. Bot. 2015, 66, 5327–5336. [Google Scholar] [CrossRef]
- Huffaker, A.; Pearce, G.; Ryan, C.A. An endogenous peptide signal in Arabidopsis activates components of the innate immune response. Proc. Natl. Acad. Sci. USA 2006, 103, 10098–10103. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Huffaker, A.; Bryan, A.C.; Tax, F.E.; Ryan, C.A. PEPR2 is a second receptor for the Pep1 and Pep2 peptides and contributes to defense responses in Arabidopsis. Plant Cell 2010, 22, 508–522. [Google Scholar] [CrossRef]
- Ma, Y.; Zhao, Y.; Walker, R.K.; Berkowitz, G.A. Molecular Steps in the Immune Signaling Pathway Evoked by Plant Elicitor Peptides: Ca2+-Dependent Protein Kinases, Nitric Oxide, and Reactive Oxygen Species Are Downstream from the Early Ca2+ Signal. Plant Physiol. 2013, 163, 1459–1471. [Google Scholar] [CrossRef]
- Wang, A.; Guo, J.; Wang, S.; Zhang, Y.; Lu, F.; Duan, J.; Liu, Z.; Ji, W. BoPEP4, a C-Terminally Encoded Plant Elicitor Peptide from Broccoli, Plays a Role in Salinity Stress Tolerance. Int. J. Mol. Sci. 2022, 23, 3090. [Google Scholar] [CrossRef] [PubMed]
- Huffaker, A.; Pearce, G.; Veyrat, N.; Erb, M.; Turlings, T.C.J.J.; Sartor, R.; Shen, Z.; Briggs, S.P.; Vaughan, M.M.; Alborn, H.T.; et al. Plant elicitor peptides are conserved signals regulating direct and indirect antiherbivore defense. Proc. Natl. Acad. Sci. USA 2013, 110, 5707–5712. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.W.; Huffaker, A.; Crippen, D.; Robbins, R.T.; Goggin, F.L. Plant elicitor peptides promote plant defences against nematodes in soybean. Mol. Plant Pathol. 2018, 19, 858–869. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Zhang, X.; Liu, J.; Tao, K.; Li, C.; Xiao, S.; Zhang, W.; Li, J.F. Plant elicitor peptide signalling confers rice resistance to piercing-sucking insect herbivores and pathogens. Plant Biotechnol. J. 2022, 20, 991–1005. [Google Scholar] [CrossRef]
- Ruiz, C.; Nadal, A.; Montesinos, E.; Pla, M. Novel Rosaceae plant elicitor peptides as sustainable tools to control Xanthomonas arboricola pv. pruni in Prunus spp. Mol. Plant Pathol. 2018, 19, 418–431. [Google Scholar] [CrossRef]
- Lori, M.; Van Verk, M.C.; Hander, T.; Schatowitz, H.; Klauser, D.; Flury, P.; Gehring, C.A.; Boller, T.; Bartels, S. Evolutionary divergence of the plant elicitor peptides (Peps) and their receptors: Interfamily incompatibility of perception but compatibility of downstream signalling. J. Exp. Bot. 2015, 66, 5315–5325. [Google Scholar] [CrossRef]
- Yang, R.; Liu, J.; Wang, Z.; Zhao, L.; Xue, T.; Meng, J.; Luan, Y. The SlWRKY6-SlPROPEP-SlPep module confers tomato resistance to Phytophthora infestans. Sci. Hortic. 2023, 318, 112117. [Google Scholar] [CrossRef]
- Wang, L.; Einig, E.; Almeida-Trapp, M.; Albert, M.; Fliegmann, J.; Mithöfer, A.; Kalbacher, H.; Felix, G. The systemin receptor SYR1 enhances resistance of tomato against herbivorous insects. Nat. Plants 2018, 4, 152–156. [Google Scholar] [CrossRef]
- Huffaker, A.; Dafoe, N.J.; Schmelz, E.A. ZmPep1, an ortholog of Arabidopsis elicitor peptide 1, regulates maize innate immunity and enhances disease resistance. Plant Physiol. 2011, 155, 1325–1338. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Pearce, G.; Ryan, C.A. The cell surface leucine-rich repeat receptor for AtPep1, an endogenous peptide elicitor in Arabidopsis, is functional in transgenic tobacco cells. Proc. Natl. Acad. Sci. USA 2006, 103, 10104–10109. [Google Scholar] [CrossRef]
- Tang, J.; Han, Z.; Sun, Y.; Zhang, H.; Gong, X.; Chai, J. Structural basis for recognition of an endogenous peptide by the plant receptor kinase PEPR1. Cell Res. 2015, 25, 110–120. [Google Scholar] [CrossRef]
- Pearce, G.; Strydom, D.; Johnson, S.; Ryan, C.A. A Polypeptide from Tomato Leaves Induces Wound-Inducible Proteinase Inhibitor Proteins. Science 1991, 253, 895–898. [Google Scholar] [CrossRef]
- Xu, S.; Liao, C.J.; Jaiswal, N.; Lee, S.; Yun, D.J.; Lee, S.Y.; Garvey, M.; Kaplan, I.; Mengiste, T. Tomato PEPR1 ORTHOLOG RECEPTOR-LIKE KINASE1 regulates responses to systemin, necrotrophic fungi, and insect herbivory. Plant Cell 2018, 30, 2214–2229. [Google Scholar] [CrossRef]
- Trivilin, A.P.; Hartke, S.; Moraes, M.G. Components of different signalling pathways regulated by a new orthologue of AtPROPEP1 in tomato following infection by pathogens. Plant Pathol. 2014, 63, 1110–1118. [Google Scholar] [CrossRef]
- Xin, X.-F.; He, S.Y. Pseudomonas syringae pv. tomato DC3000: A Model Pathogen for Probing Disease Susceptibility and Hormone Signaling in Plants. Annu. Rev. Phytopathol. 2013, 51, 473–498. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.L. The origin of a land flora. Nat. Plants 2022, 8, 1352–1369. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhang, H.; Lin, J. Systemin-mediated long-distance systemic defense responses. N. Phytologist. 2020, 226, 1573–1582. [Google Scholar] [CrossRef] [PubMed]
- Huffaker, A.; Ryan, C.A. Endogenous peptide defense signals in Arabidopsis differentially amplify signaling for the innate immune response. Proc. Natl. Acad. Sci. USA 2007, 104, 10732–10736. [Google Scholar] [CrossRef]
- Buonanno, M.; Coppola, M.; Di Lelio, I.; Molisso, D.; Leone, M.; Pennacchio, F.; Langella, E.; Rao, R.; Monti, S.M. Prosystemin, a prohormone that modulates plant defense barriers, is an intrinsically disordered protein. Protein Sci. 2018, 27, 620–632. [Google Scholar] [CrossRef]
- Huffaker, A. Plant elicitor peptides in induced defense against insects. Curr. Opin. Insect Sci. 2015, 9, 44–50. [Google Scholar] [CrossRef]
- Wari, D.; Kuramitsu, K.; Kavallieratos, N.G. Sap-sucking pests; they do matter. Insects 2021, 12, 10–13. [Google Scholar] [CrossRef]
- Ling, H.; Fu, X.; Huang, N.; Zhong, Z.; Su, W.; Lin, W.; Cui, H.; Que, Y. A sugarcane smut fungus effector simulates the host endogenous elicitor peptide to suppress plant immunity. N. Phytol. 2022, 233, 919–933. [Google Scholar] [CrossRef]
- Furumizu, C.; Aalen, R.B. Peptide signaling through leucine-rich repeat receptor kinases: Insight into land plant evolution. New Phytol. 2023, 238, 977–982. [Google Scholar] [CrossRef]
- Zhao, B.; Liu, Q.; Wang, B.; Yuan, F. Roles of Phytohormones and Their Signaling Pathways in Leaf Development and Stress Responses. J. Agric. Food Chem. 2021, 69, 3566–3584. [Google Scholar] [CrossRef]
- Li, N.; Han, X.; Feng, D.; Yuan, D.; Huang, L.J. Signaling crosstalk between salicylic acid and ethylene/Jasmonate in plant defense: Do we understand what they are whispering? Int. J. Mol. Sci. 2019, 20, 671. [Google Scholar] [CrossRef] [PubMed]
- Thomma, B.; Eggermont, K.; Penninckx, I.A.M.A.; Mauch-Mani, B.; Vogelsang, R.; Cammue, B.P.A.; Broekaert, W.F. Separate jasmonate-dependent and salicylate-dependent defense-response pathways in Arabidopsis are essential for resistance to distinct microbial pathogens. Proc. Natl. Acad. Sci. USA 1998, 95, 15107–15111. [Google Scholar] [CrossRef] [PubMed]
- Tintor, N.; Ross, A.; Kanehara, K.; Yamada, K.; Fan, L.; Kemmerling, B.; Nürnberger, T.; Tsuda, K.; Saijo, Y. Layered pattern receptor signaling via ethylene and endogenous elicitor peptides during Arabidopsis immunity to bacterial infection. Proc. Natl. Acad. Sci. USA 2013, 110, 6211–6216. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-Y.; Lin, Y.-M.; Chao, T.-C.; Wang, J.-F.; Liu, A.-C.; Ho, F.-I.; Cheng, C.-P. Virus-induced gene silencing reveals the involvement of ethylene-, salicylic acid- and mitogen-activated protein kinase-related defense pathways in the resistance of tomato to bacterial wilt. Physiol. Plant. 2009, 136, 324–335. [Google Scholar] [CrossRef]
- Nasir, F.; Tian, L.; Chang, C.; Li, X.; Gao, Y.; Tran, L.S.P.; Tian, C. Current understanding of pattern-triggered immunity and hormone-mediated defense in rice (Oryza sativa) in response to Magnaporthe oryzae infection. Semin. Cell Dev. Biol. 2018, 83, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Mishina, T.E.; Zeier, J. Pathogen-associated molecular pattern recognition rather than development of tissue necrosis contributes to bacterial induction of systemic acquired resistance in Arabidopsis. Plant J. 2007, 50, 500–513. [Google Scholar] [CrossRef]
- Kawano, T.; Bouteau, F. Crosstalk between Intracellular and Extracellular Salicylic Acid Signaling Events Leading to Long-Distance Spread of Signals. Plant Cell Rep. 2013, 32, 1125–1138. [Google Scholar] [CrossRef]
- Saand, M.A.; Xu, Y.; Li, W.; Wang, J.; Cai, X. Cyclic nucleotide gated channel gene family in tomato: Genome-wide identification and functional analyses in disease resistance. Front. Plant Sci. 2015, 6, 303. [Google Scholar] [CrossRef]
- Sandor, R.; Der, C.; Grosjean, K.; Anca, I.; Noirot, E.; Leborgne-Castel, N.; Lochman, J.; Simon-Plas, F.; Gerbeau-Pissot, P. Plasma membrane order and fluidity are diversely triggered by elicitors of plant defence. J. Exp. Bot. 2016, 67, 5173. [Google Scholar] [CrossRef]
- Newman, M.A.; Sundelin, T.; Nielsen, J.T.; Erbs, G. MAMP (microbe-associated molecular pattern) triggered immunity in plants. Front. Plant Sci. 2013, 4, 139. [Google Scholar] [CrossRef]
- Kimura, S.; Waszczak, C.; Hunter, K.; Wrzaczek, M. Bound by fate: The role of reactive oxygen species in receptor-like kinase signaling. Plant Cell 2017, 29, 638–654. [Google Scholar] [CrossRef]
- Lecourieux, D.; Ranjeva, R.; Pugin, A. Calcium in plant defence-signalling pathways. N. Phytol. 2006, 171, 249–269. [Google Scholar] [CrossRef]
- Moyen, C.; Hammond-Kosack, K.E.; Jones, J.; Knight, M.R.; Johannes, E. Systemin triggers an increase of cytoplasmic calcium in tomato mesophyll cells: Ca2+ mobilization from intra- and extracellular compartments. Plant Cell Environ. 1998, 21, 1101–1111. [Google Scholar] [CrossRef]
- Ryan, C.A. The systemin signaling pathway: Differential activation of plant defensive genes. Biochim. Et Biophys. Acta 2000, 1477, 112–121. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Y.; Bao, Q.; Wang, H.; Hou, S. Plant elicitor peptide 1 fortifies root cell walls and triggers a systemic root-to-shoot immune signaling in Arabidopsis. Plant Signal. Behav. 2022, 17. [Google Scholar] [CrossRef]
- Bergey, D.R.; Ryan, C. a Wound- and Systemin-Inducible Calmodulin Gene Expression in Tomato Leaves. Plant Mol. Biol. 1999, 40, 815–823. [Google Scholar] [CrossRef]
- Han, G.-Z. Origin and evolution of the plant immune system. New Phytol. 2019, 222, 70–83. [Google Scholar] [CrossRef]
- Liu, P.L.; Du, L.; Huang, Y.; Gao, S.M.; Yu, M. Origin and diversification of leucine-rich repeat receptor-like protein kinase (LRR-RLK) genes in plants. BMC Evol. Biol. 2017, 17, 47. [Google Scholar] [CrossRef]
- Afzal, A.J.; Wood, A.J.; Lightfoot, D.A. Plant receptor-like serine threonine kinases: Roles in signaling and plant defense. Mol. Plant-Microbe Interact. 2008, 21, 507–517. [Google Scholar] [CrossRef]
- Dunning, F.M.; Sun, W.; Jansen, K.L.; Helft, L.; Bent, A.F. Identification and mutational analysis of Arabidopsis FLS2 leucine-rich repeat domain residues that contribute to flagellin perception. Plant Cell 2007, 19, 3297–3313. [Google Scholar] [CrossRef]
- Rahman, H.; Wang, X.Y.; Xu, Y.P.; He, Y.H.; Cai, X.Z. Characterization of tomato protein kinases embedding guanylate cyclase catalytic center motif. Sci. Rep. 2020, 10, 4078. [Google Scholar] [CrossRef] [PubMed]
- Gong, P.; Zhang, J.; Li, H.; Yang, C.; Zhang, C.; Zhang, X.; Khurram, Z.; Zhang, Y.; Wang, T.; Fei, Z.; et al. Transcriptional profiles of drought-responsive genes in modulating transcription signal transduction, and biochemical pathways in tomato. J. Exp. Bot. 2010, 61, 3563–3575. [Google Scholar] [CrossRef] [PubMed]
- Malinowski, R.; Higgins, R.; Luo, Y.; Piper, L.; Nazir, A.; Bajwa, V.S.; Clouse, S.D.; Thompson, P.R.; Stratmann, J.W. The tomato brassinosteroid receptor BRI1 increases binding of systemin to tobacco plasma membranes, but is not involved in systemin signaling. Plant Mol. Biol. 2009, 70, 603–616. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Pozo, N.; Menda, N.; Edwards, J.D.; Saha, S.; Tecle, I.Y.; Strickler, S.R.; Bombarely, A.; Fisher-York, T.; Pujar, A.; Foerster, H.; et al. The Sol Genomics Network (SGN)—From genotype to phenotype to breeding. Nucleic Acids Res. 2015, 43, D1036–D1041. [Google Scholar] [CrossRef] [PubMed]
- Bowman, J.L.; Kohchi, T.; Yamato, K.T.; Jenkins, J.; Shu, S.; Ishizaki, K.; Yamaoka, S.; Nishihama, R.; Nakamura, Y.; Berger, F.; et al. Insights into Land Plant Evolution Garnered from the Marchantia Polymorpha Genome. Cell 2017, 171, 287–304.e15. [Google Scholar] [CrossRef]
- DeFalco, T.A.; Zipfel, C. Molecular mechanisms of early plant pattern-triggered immune signaling. Mol. Cell 2021, 81, 3449–3467. [Google Scholar] [CrossRef]
- Montoya, T.; Nomura, T.; Farrar, K.; Kaneta, T.; Yokota, T.; Bishop, G.J. Cloning the Tomato Curl3 Gene Highlights the Putative Dual Role of the Leucine-Rich Repeat Receptor Kinase TBRI1/SR160 in Plant Steroid Hormone and Peptide Hormone Signaling. Plant Cell 2002, 14, 3163–3176. [Google Scholar] [CrossRef]
- Jones, J. Harpin. Pestic. Outlook 2001, 12, 134–135. [Google Scholar] [CrossRef]
- Wei, Z.M.; Laby, R.J.; Zumoff, C.H.; Bauer, D.W.; He, S.Y.; Collmer, A.; Beer, S.V. Harpin, elicitor of the hypersensitive response produced by the plant pathogen Erwinia amylovora. Science 1992, 257, 85–88. [Google Scholar] [CrossRef]
- Sands, L.B.; Cheek, T.; Reynolds, J.; Ma, Y.; Berkowitz, G.A. Effects of Harpin and Flg22 on Growth Enhancement and Pathogen Defense in Cannabis sativa Seedlings. Plants 2022, 11, 1178. [Google Scholar] [CrossRef]
- Esquivel-cervantes, L.F.; Tlapal-bolaños, B.; Tovar-pedraza, J.M.; Pérez-hernández, O.; Leyva-mir, S.G.; Camacho-tapia, M. Efficacy of Biorational Products for Managing Diseases of Tomato in Greenhouse Production. Plants 2022, 11, 1638. [Google Scholar] [CrossRef]
- Obradovic, A.; Jones, J.B.; Momol, M.T.; Olson, S.M.; Jackson, L.E.; Balogh, B.; Guven, K.; Iriarte, F.B. Integration of biological control agents and systemic acquired resistance inducers against bacterial spot on tomato. Plant Dis. 2005, 89, 712–716. [Google Scholar] [CrossRef]
- Choi, M.S.; Kim, W.; Lee, C.; Oh, C.S. Harpins, multifunctional proteins secreted by gram-negative plant-pathogenic bacteria. Mol. Plant-Microbe Interact. 2013, 26, 1115–1122. [Google Scholar] [CrossRef]
- Montesinos, E.; Badosa, E.; Pla, M.; Montesinos, L.; Montesinos, A. Antimicrobial and Defense Elicitor Peptides as Biopesticides for Plant Disease Control. In Biocontrol of Plant Disease; Wiley Online Library: Hoboken, NJ, USA, 2022. [Google Scholar]
- Burketova, L.; Trda, L.; Ott, P.G.; Valentova, O. Bio-based resistance inducers for sustainable plant protection against pathogens. Biotechnol. Adv. 2015, 33, 994–1004. [Google Scholar] [CrossRef]
- Aprile, A.M.; Coppola, M.; Turrà, D.; Vitale, S.; Cascone, P.; Diretto, G.; Fiore, A.; Castaldi, V.; Romanelli, A.; Avitabile, C.; et al. Combination of the Systemin peptide with the beneficial fungus Trichoderma afroharzianum T22 improves plant defense responses against pests and diseases. J. Plant Interact. 2022, 17, 569–579. [Google Scholar] [CrossRef]


{kind=link}
| DAMP | Species | Receptor (Predicted) * | Demonstrated Biological Activity of Pep Signaling | References |
|---|---|---|---|---|
| AtPeps 1-8 | Arabidopsis | AtPEPR1, AtPEPR2 | anti-chewing insect herbivore; anti-hemibiotrophic pathogen; anti-necrotrophic pathogen; root development | [15,16,17,18] |
| BoPep4 | broccoli | unreported | salinity response | [19] |
| GmPep1, GmPep2, GmPep3 | soybean | GmPEPR1a, GmPEPR2a, GmPEPR2 | anti-chewing insect herbivore, anti-nematode | [20,21] |
| OsPep3 | rice | OsPEPR1a, OsPEPR1b | anti-bacterial, anti-fungal; anti-piercing-sucking insect herbivore | [22] |
| PpPep1 and 2 | peach | PpPEPR1a, PpPEPR1b | anti-bacterial necrotrophic pathogen | [23] |
| SlPep | tomato | SlPEPR, SlGC17 | anti-necrotrophic pathogen | [24,25] |
| SlSystemin | tomato (homologs in Solanae clade) | SlSYR1, SlSYR2 | anti-necrotrophic pathogen; anti-chewing insect herbivore | [26] |
| ZmPep1, ZmPep3 | maize | ZmPEPR1a, ZmPEPR2a | anti-chewing insect herbivore; anti-necrotrophic pathogen | [20,27] |
| Protein | Alternate Name | Biological Role | Reference | SolGenomics ID | NCBI Identifier |
|---|---|---|---|---|---|
| SlPEPR | SlPORK1 | proposed Pep receptor | [24] | Solyc03g123860 | XP_004235511.1 |
| SlPEPR | systemin signaling pathway component | [31] | |||
| INRPK1c | expression upregulated in drought-susceptible cultivars during drought stress | [64] | |||
| SlGC18 | proposed PEPR; guanylyl cyclase | [63] | |||
| SlGC17 | n/a | proposed PEPR; guanylyl cyclase | [63] | Solyc03g112580.2.1 | XP_004236236.1 |
| SlSYR1 | * | systemin receptor (biochemical evidence) | [26] | Solyc03g082470.2.1 | XP_004235118.1 |
| SlSYR2 | n/a | systemin receptor | [26] | Solyc03g082450.2.1 | XP_004235119.1 |
| SlBRI1 | SR160; cu3 receptor | proposed systemin receptor, later shown to be incorrect by Malinowski et al., 67 | [69] | Solyc04g051510.1.1 | NP_001296180.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zelman, A.K.; Berkowitz, G.A. Plant Elicitor Peptide (Pep) Signaling and Pathogen Defense in Tomato. Plants 2023, 12, 2856. https://doi.org/10.3390/plants12152856
Zelman AK, Berkowitz GA. Plant Elicitor Peptide (Pep) Signaling and Pathogen Defense in Tomato. Plants. 2023; 12(15):2856. https://doi.org/10.3390/plants12152856
Chicago/Turabian StyleZelman, Alice K., and Gerald Alan Berkowitz. 2023. "Plant Elicitor Peptide (Pep) Signaling and Pathogen Defense in Tomato" Plants 12, no. 15: 2856. https://doi.org/10.3390/plants12152856
APA StyleZelman, A. K., & Berkowitz, G. A. (2023). Plant Elicitor Peptide (Pep) Signaling and Pathogen Defense in Tomato. Plants, 12(15), 2856. https://doi.org/10.3390/plants12152856