

SNAC3 Transcription Factor Enhances Arsenic Stress Tolerance and Grain Yield in Rice (Oryza sativa L.) through Regulating Physio-Biochemical Mechanisms, Stress-Responsive Genes, and Cryptochrome 1b

 , , ,
, , ,  ,
, 
Abstract
1. Introduction
2. Results and Discussion
2.1. Generation and Molecular Characterization of Transgenic Rice Lines
2.2. SNAC3 Overexpression Enhances While SNAC3 Suppression Reduces Survival Rate, Leaf Relative Water Content, and Growth Characteristics of Rice under Arsenic Stress
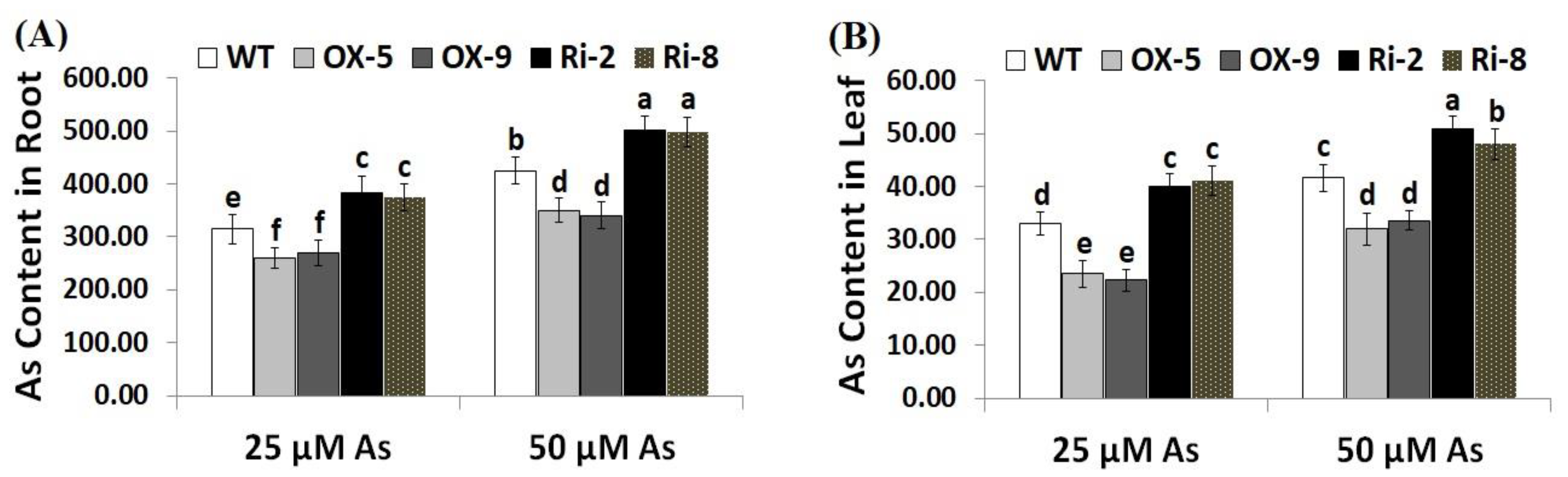
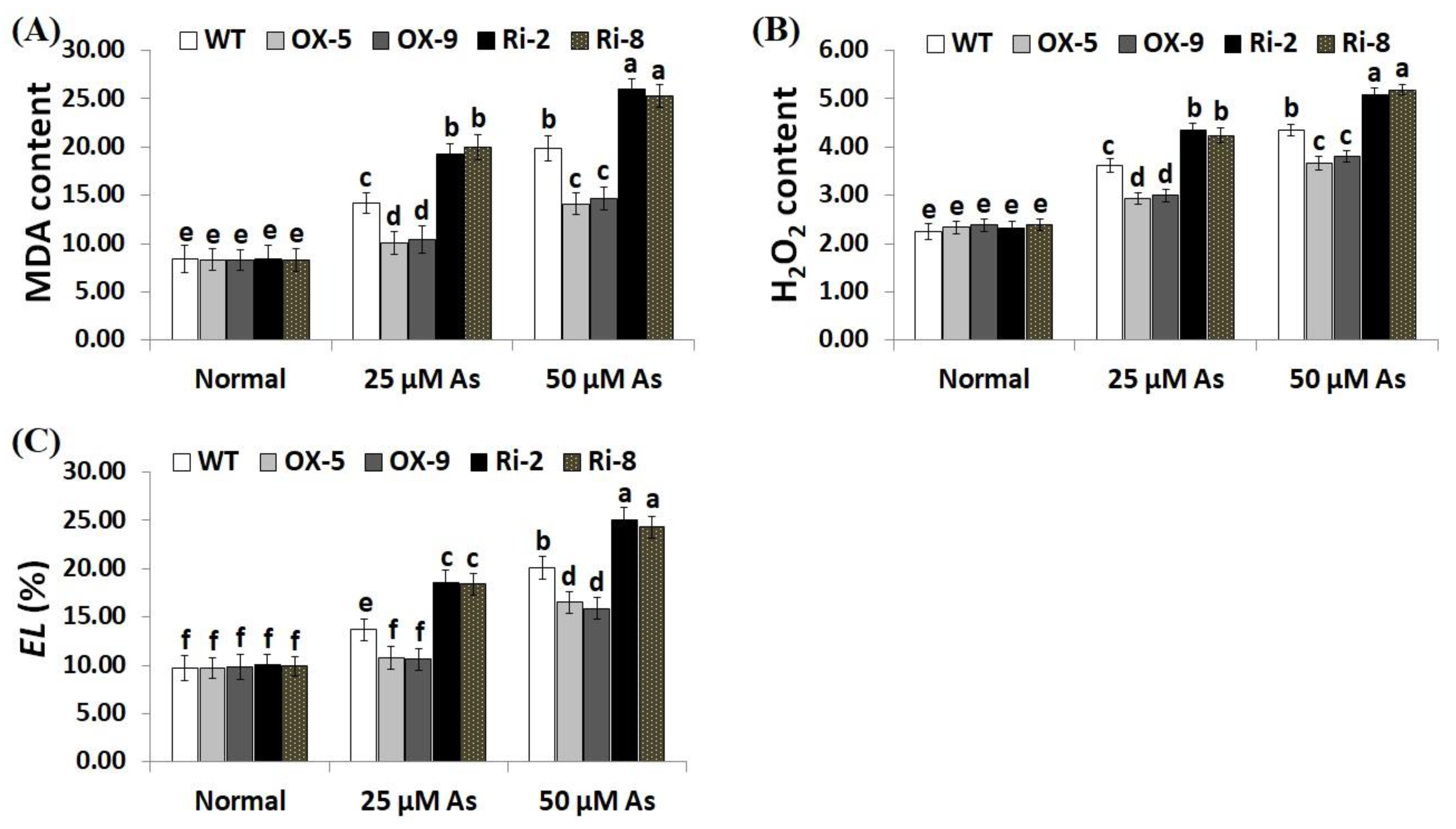
2.3. Overexpression of SNAC3 Reduces While Suppression of SNAC3 Increases Arsentic Uptake and Oxidative Stress Biomarkers in Rice under Arsenic Stress
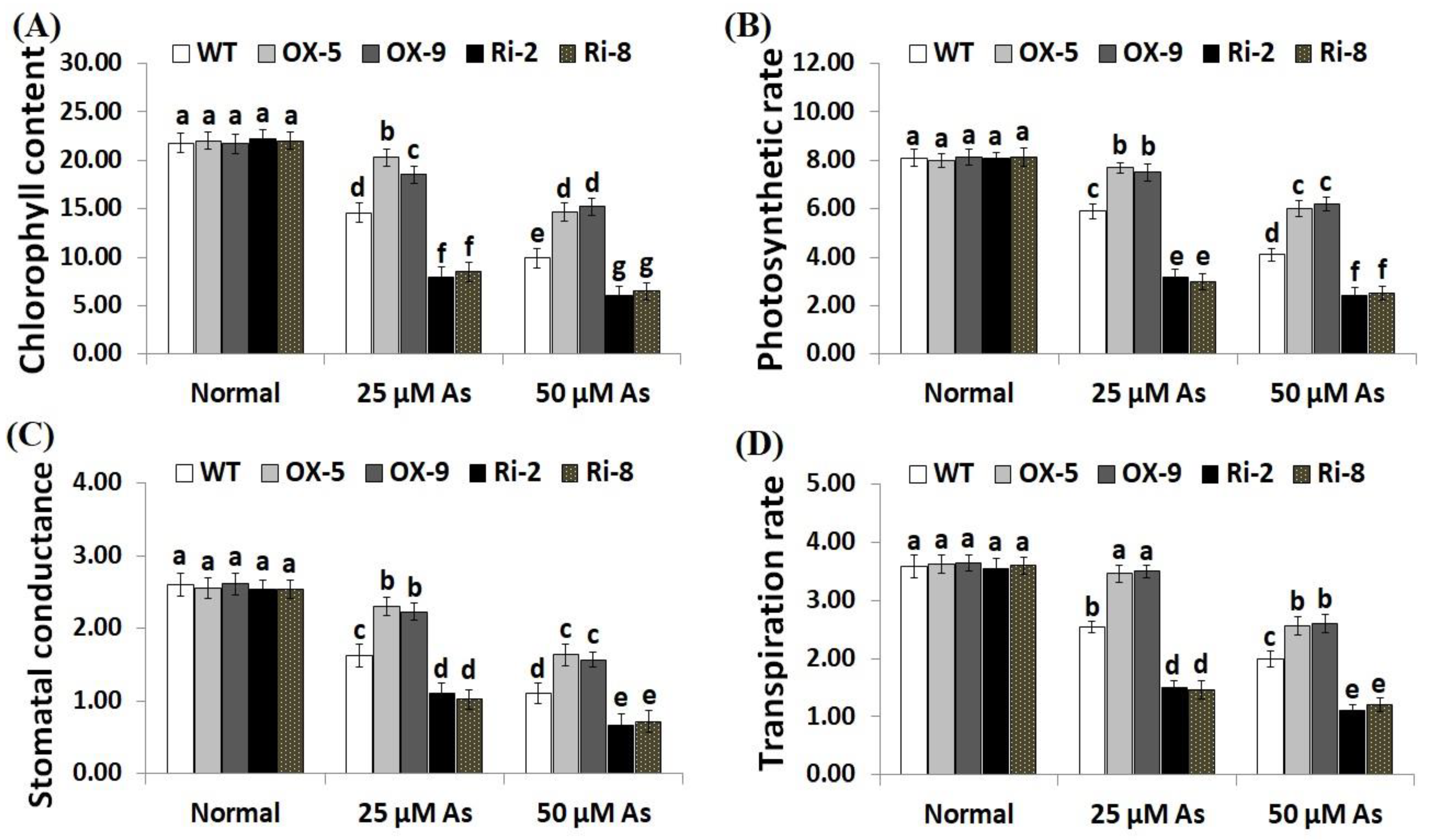
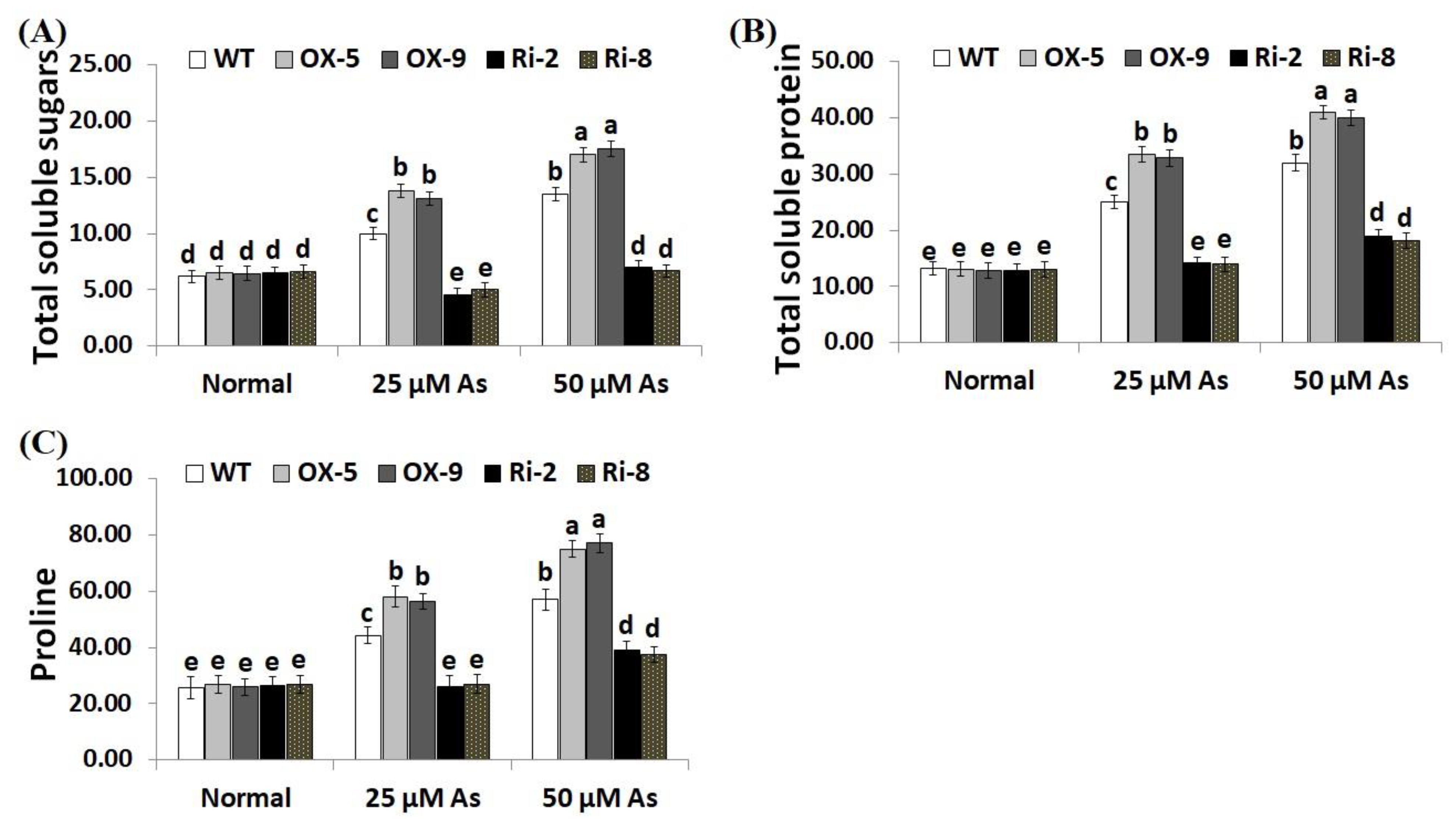
2.4. Overexpression of SNAC3 Enhances While Suppression of SNAC3 Reduces Chlorophyll Content, Gas-Exchange Parameters, and Osmolyte Accumulation in Rice under Arsenic Stress
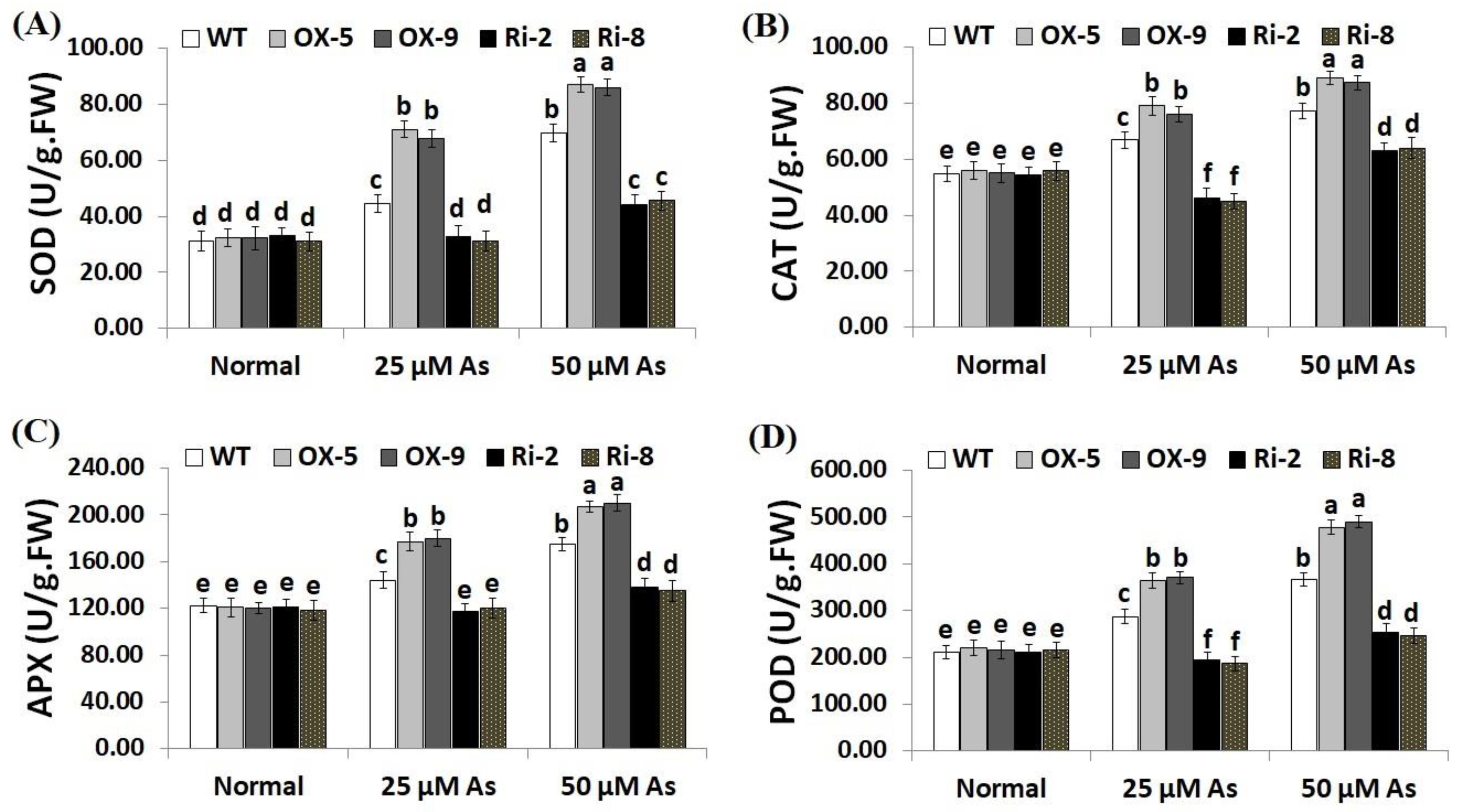
2.5. Overexpression of SNAC3 Induces While Suppression of SNAC3 Reduces Antioxidant Enzyme Activities in Rice under Arsenic Stress
2.6. SNAC3 Overexpression and Suppression Modulate Abiotic Stress-Associated Genes Expression in Rice under Arsenic Stress
2.7. Overexpression of SNAC3 Improves While Suppression of SNAC3 Reduces Rice Grain Yield and Yield Components under Arsenic Stress
3. Materials and Methods
3.1. Plant Material and Growth Circumstances
3.2. Construction of Plasmids and Transformation of Rice Plants
3.3. Molecular Characterization of Transformed Rice Lines
3.4. Arsenic Stress Treatments
3.5. Estimating the Morphological Characteristics and Relative Water Content
3.6. Estimating Arsenic Content in Plant Roots and Leaves
3.7. Measurement of Oxidative Stress Biomarkers
3.8. Estimation of Total Chlorophyll, Gas-Exchange Parameters, Soluble Protein, Sugars, Proline, Phenolics, and Flavonoids
3.9. Antioxidant Enzyme Assays
3.10. Analysis of Stress-Tolerant Genes Expression
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yuan, X.; Wang, H.; Cai, J.; Bi, Y.; Li, D.; Song, F. Rice NAC transcription factor ONAC066 functions as a positive regulator of drought and oxidative stress response. BMC Plant Biol. 2019, 19, 278. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.; Thounaojam, T.C.; Upadhyaya, H. Arsenic stress in Rice (Oryza sativa) and its amelioration approaches. Plant Stress 2022, 4, 100076. [Google Scholar] [CrossRef]
- Singh, V.; Singh, N.; Rai, S.N.; Kumar, A.; Singh, A.K.; Singh, M.P.; Sahoo, A.; Shekhar, S.; Vamanu, E.; Mishra, V. Heavy Metal Contamination in the Aquatic Ecosystem: Toxicity and Its Remediation Using Eco-Friendly Approaches. Toxics 2023, 11, 147. [Google Scholar] [CrossRef]
- Thijs, S.; Sillen, W.; Weyens, N.; Vangronsveld, J. Phytoremediation: State-of-the-art and a key role for the plant microbiome in future trends and research prospects. Int. J. Phytoremediat. 2017, 19, 23–38. [Google Scholar] [CrossRef]
- Noor, I.; Sohail, H.; Sun, J.; Nawaz, M.A.; Li, G.; Hasanuzzaman, M.; Liu, J. Heavy metal and metalloid toxicity in horticultural plants: Tolerance mechanism and remediation strategies. Chemosphere 2022, 303, 135196. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Al-Ghamdi, A.A.; Ali, H.M.; Alayafi, A.A.; Witczak, J.; Ahmad, M. Analysis of Genetic Variation and Enhancement of Salt Tolerance in French Pea (Pisum Sativum L.). Int. J. Mol. Sci. 2018, 19, 2433. [Google Scholar] [CrossRef] [PubMed]
- Zulfiqar, U.; Ayub, A.; Hussain, S.; Waraich, E.; El-Esawi, M.; Ishfaq, M.; Ahmad, M.; Ali, N.; Maqsood, M. Cadmium Toxicity in Plants: Recent Progress on Morpho-physiological Effects and Remediation Strategies. J. Soil Sci. Plant Nutr. 2022, 22, 212–269. [Google Scholar] [CrossRef]
- Castilhos, G.; Lazzarotto, F.; Spagnolo-Fonini, L.; Bodanese-Zanettini, M.H.; Margis-Pinheiro, M. Possible roles of basic helix-loop-helix transcription factors in adaptation to drought. Plant Sci. 2014, 223, 1–7. [Google Scholar] [CrossRef]
- Shao, H.B.; Wang, H.Y.; Tang, X.L. NAC transcription factors in plant multiple abiotic stress responses: Progress and prospects. Front Plant Sci. 2015, 6, 902. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Al-Ghamdi, A.A.; Ali, H.M.; Ahmad, M. Overexpression of AtWRKY30 Transcription Factor Enhances Heat and Drought Stress Tolerance in Wheat (Triticum aestivum L.). Genes 2019, 10, 163. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Alayafi, A.A. Overexpression of StDREB2 Transcription Factor Enhances Drought Stress Tolerance in Cotton (Gossypium barbadense L.). Genes 2019, 10, 142. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, X.; Xie, X.; Liu, L.; Fu, J.; Wang, Q. Maize transcription factor ZmNAC2 enhances osmotic stress tolerance in transgenic Arabidopsis. J. Plant Physiol. 2023, 282, 153948. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Zhang, H.; Li, H.; Lian, L.; Wei, Y.; Lin, Y.; Wang, L.; He, W.; Cai, Q.; Xie, H.; et al. IPA1 improves drought tolerance by activating SNAC1 in rice. BMC Plant Biol. 2023, 23, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Pan, L.; Yang, Z.; Su, M.; Xu, J.; Jiang, X.; Yin, X.; Wang, T.; Wan, F.; Chi, X. A MYB-related transcription factor from peanut, AhMYB30, improves freezing and salt stress tolerance in transgenic Arabidopsis through both DREB/CBF and ABA-signaling pathways. Front. Plant Sci. 2023, 14, 1136626. [Google Scholar] [CrossRef]
- Oliveira, P.N.d.; Matias, F.; Martínez-Andújar, C.; Martinez-Melgarejo, P.A.; Prudencio, Á.S.; Galeano, E.; Pérez-Alfocea, F.; Carrer, H. Overexpression of TgERF1, a Transcription Factor from Tectona grandis, Increases Tolerance to Drought and Salt Stress in Tobacco. Int. J. Mol. Sci. 2023, 24, 4149. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, C.; Wang, L.; Zhong, H.; Xu, X.; Cheng, Y.; Nian, H.; Liu, W.; Chen, P.; Zhang, A.; et al. GmABR1 encoding an ERF transcription factor enhances the tolerance to aluminum stress in Arabidopsis thaliana. Front. Plant Sci. 2023, 14, 1125245. [Google Scholar] [CrossRef]
- Uauy, C.; Distelfeld, A.; Fahima, T.; Blechl, A.; Dubcovsky, J.A. NAC gene regulating senescence improves grain protein, zinc, and iron content in wheat. Science 2006, 314, 1298–1301. [Google Scholar] [CrossRef]
- Nuruzzaman, M.; Sharoni, A.M.; Satoh, K.; Moumeni, A.; Venuprasad, R.; Serraj, R.; Kumar, A.; Leung, H.; Attia, K.; Kikuchi, S. Comprehensive gene expression analysis of the NAC gene family under normal growth conditions, hormone treatment, and drought stress conditions in rice using near-isogenic lines (NILs) generated from crossing Aday selection (drought tolerant) and IR64. Mol. Gen. Genom. 2012, 287, 389–410. [Google Scholar] [CrossRef]
- Nakashima, K.; Takasaki, H.; Mizoi, J.; Shinozaki, K.; Yamaguchi-Shinozaki, K. NAC transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta 2012, 1819, 97–103. [Google Scholar] [CrossRef]
- Fang, Y.; Liao, K.; Du, H.; Xu, Y.; Song, H.; Li, X.; Xiong, L. A stress-responsive NAC transcription factor SNAC3 confers heat and drought tolerance through modulation of reactive oxygen species in rice. J. Exp. Bot. 2015, 66, 6803–6817. [Google Scholar] [CrossRef]
- Tran, L.S.; Nakashima, K.; Sakuma, Y.; Simpson, S.D.; Fujita, Y.; Maruyama, K.; Fujita, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Isolation and functional analysis of Arabidopsis stress-inducible NAC transcription factors that bind to a drought-responsive cis-element in the early responsive to dehydration stress 1 promoter. Plant Cell 2004, 16, 2481–2498. [Google Scholar] [CrossRef] [PubMed]
- De Clercq, I.; Vermeirssen, V.; Van Aken, O.; Vandepoele, K.; Murcha, M.W.; Law, S.R.; Inzé, A.; Ng, S.; Ivanova, A.; Rombaut, D.; et al. The membrane-bound NAC transcription factor ANAC013 functions in mitochondrial retrograde regulation of the oxidative stress response in Arabidopsis. Plant Cell 2013, 25, 3472–3490. [Google Scholar] [CrossRef] [PubMed]
- Shahnejat-Bushehri, S.; Nobmann, B.; Devi Allu, A.; Balazadeh, S. JUB1 suppresses Pseudomonas syringae-induced defense responses through accumulation of DELLA proteins. Plant Signal. Behav. 2016, 11, e1181245. [Google Scholar] [CrossRef] [PubMed]
- Shahnejat-Bushehri, S.; Tarkowska, D.; Sakuraba, Y.; Balazadeh, S. Arabidopsis NAC transcription factor JUB1 regulates GA/BR metabolism and signalling. Nat. Plants 2016, 2, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.; Allu, A.D.; Garapati, P.; Siddiqui, H.; Dortay, H.; Zanor, M.I.; Asensi-Fabado, M.A.; Munne-Bosch, S.; Antonio, C.; Tohge, T.; et al. JUNGBRUNNEN1, a reactive oxygen species-responsive NAC transcription factor, regulates longevity in Arabidopsis. Plant Cell 2012, 24, 482–506. [Google Scholar] [CrossRef]
- Ebrahimian-Motlagh, S.; Ribone, P.A.; Thirumalaikumar, V.P.; Allu, A.D.; Chan, R.L.; Mueller-Roeber, B.; Balazadeh, S. JUNGBRUNNEN1 confers drought tolerance downstream of the HD-zip I transcription factor AtHB13. Front Plant Sci. 2017, 8, 2118. [Google Scholar] [CrossRef]
- Tak, H.; Negi, S.; Ganapathi, T.R. Banana NAC transcription factor MusaNAC042 is positively associated with drought and salinity tolerance. Protoplasma 2017, 254, 803–816. [Google Scholar] [CrossRef]
- Thirumalaikumar, V.P.; Devkar, V.; Mehterov, N.; Ali, S.; Ozgur, R.; Turkan, I.; Mueller-Roeber, B.; Balazadeh, S. NAC transcription factor JUNGBRUNNEN1 enhances drought tolerance in tomato. Plant Biotechnol. J. 2018, 16, 354–366. [Google Scholar] [CrossRef]
- Shahnejat-Bushehri, S.; Mueller-Roeber, B.; Balazadeh, S. Arabidopsis NAC transcription factor JUNGBRUNNEN1 affects thermomemory-associated genes and enhances heat stress tolerance in primed and unprimed conditions. Plant Signal Behav. 2012, 7, 1518–1521. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, H.-Y.; Chen, Y.-F. The transcription factor MYB40 is a central regulator in arsenic resistance in Arabidopsis. Plant Comm. 2021, 2, 100234. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.Z.; Chen, M.X.; Yu, L.J.; Xie, L.J.; Yuan, L.B.; Qi, H.; Xiao, M.; Guo, W.; Chen, Z.; Yi, K.; et al. OsARM1, an R2R3 MYB transcription factor, is involved in regulation of the response to arsenic stress in rice. Front. Plant Sci. 2017, 8, 1868. [Google Scholar] [CrossRef]
- Hu, H.H.; You, J.; Fang, Y.J.; Zhu, X.Y.; Qi, Z.Y.; Xiong, L.Z. Characterization of transcription factor gene SNAC2 conferring cold and salt tolerance in rice. Plant Mol. Biol. 2008, 67, 169–181. [Google Scholar] [CrossRef]
- Redillas, M.C.; Jeong, J.S.; Kim, Y.S.; Jung, H.; Bang, S.W.; Choi, Y.D.; Ha, S.H.; Reuzeau, C.; Kim, J.K. The overexpression of OsNAC9 alters the root architecture of rice plants enhancing drought resistance and grain yield under field conditions. Plant Biotechnol. J. 2012, 10, 792–805. [Google Scholar] [CrossRef]
- Lee, D.K.; Chung, P.J.; Jeong, J.S.; Jang, G.; Bang, S.W.; Jung, H.; Kim, Y.S.; Ha, S.H.; Choi, Y.D.; Kim, J.K. The rice OsNAC6 transcription factor orchestrates multiple molecular mechanisms involving root structural adaptions and nicotianamine biosynthesis for drought tolerance. Plant Biotechnol. J. 2017, 15, 754–764. [Google Scholar] [CrossRef]
- Huang, L.; Hong, Y.; Zhang, H.; Li, D.; Song, F. Rice NAC transcription factor ONAC095 plays opposite roles in drought and cold stress tolerance. BMC Plant Biol. 2016, 16, 203. [Google Scholar] [CrossRef]
- Jeong, J.S.; Kim, Y.S.; Redillas, M.C.; Jang, G.; Jung, H.; Bang, S.W.; Choi, Y.D.; Ha, S.H.; Reuzeau, C.; Kim, J.K. OsNAC5 overexpression enlarges root diameter in rice plants leading to enhanced drought tolerance and increased grain yield in the field. Plant Biotechnol. J. 2013, 11, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, Y.; Lv, B.; Li, J.; Luo, L.; Lu, S.; Zhang, X.; Ma, H.; Ming, F. The NAC family transcription factor OsNAP confers abiotic stress response through the ABA pathway. Plant Cell Physiol. 2014, 55, 604–619. [Google Scholar] [CrossRef]
- Tran, L.S.; Nishiyama, R.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Potential utilization of NAC transcription factors to enhance abiotic stress tolerance in plants by biotechnological approach. GM Crops 2010, 1, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Xiong, L. Genetic engineering and breeding of drought-resistant crops. Annu. Revi. Plant Biol. 2014, 65, 715–741. [Google Scholar] [CrossRef] [PubMed]
- Ishizaki, K.; Schauer, N.; Larson, T.R.; Graham, I.A.; Fernie, A.R.; Leaver, C.J. The mitochondrial electron transfer flavoprotein complex is essential for survival of Arabidopsis in extended darkness. Plant J. 2006, 47, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.S.; Kim, Y.S.; Baek, K.H.; Jung, H.; Ha, S.H.; Do Choi, Y.; Kim, M.; Reuzeau, C.; Kim, J.K. Root-specific expression of OsNAC10 improves drought tolerance and grain yield in rice under field drought conditions. Plant Physiol. 2010, 153, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Castrillo, G.; Sánchez-Bermejo, E.; de Lorenzo, L.; Crevillén, P.; Fraile-Escanciano, A.; Tc, M.; Mouriz, A.; Catarecha, P.; Sobrino-Plata, J.; Olsson, S.; et al. WRKY6 transcription factor restricts arsenate uptake and transposon activation in Arabidopsis. Plant Cell 2013, 25, 2944–2957. [Google Scholar] [CrossRef]
- Bajji, M.; Kinet, J.; Lutts, S. The use of the electrolyte leakage method for assessing cell membrane stability as a water stress tolerance test in durum wheat. Plant Growth Regul. 2002, 36, 61–70. [Google Scholar] [CrossRef]
- Ganguli, L.J. Lipid peroxidation-DNA damage by malondialdehyde. Mutat. Res. Fundam. Mol. Mech. 1999, 424, 83–95. [Google Scholar]
- Ben, K.R.; Abdelly, C.; Savouré, A. Proline, a multifunctional amino-acid involved in plant adaptation to environmental constraints. Biol. Aujourdhui 2012, 206, 291. [Google Scholar]
- Chaleff, R.S. Further characterization of picloram tolerant mutance of Nicotinana tabacum. Theor. Appl. Genet. 1980, 58, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Liu, P.; Zhu, J. Effect of magnesium (Mg) on contents of free proline, soluble sugar and protein in soybean leaves. J. Henan Agric. Sci. 2004, 6, 35–38. [Google Scholar]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alzahrani, S.M.; Ali, H.M.; Alayafi, A.A.; Ahmad, M. Serratia liquefaciens KM4 Improves Salt Stress Tolerance in Maize by Regulating Redox Potential, Ion Homeostasis, Leaf Gas Exchange and Stress-Related Gene Expression. Int. J. Mol. Sci. 2018, 19, 3310. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Alayafi, A.A. Overexpression of Rice Rab7 Gene Improves Drought and Heat Tolerance and Increases Grain Yield in Rice (Oryza sativa L.). Genes 2019, 10, 56. [Google Scholar] [CrossRef]
- Cai, W.; Liu, W.; Wang, W.S.; Fu, Z.W.; Han, T.T.; Lu, Y.T. Overexpression of rat neurons nitric oxide synthase in rice enhances drought and salt tolerance. PLoS ONE 2015, 10, e0131599. [Google Scholar] [CrossRef]
- Yu, J.; Lai, Y.; Wu, X.; Wu, G.; Guo, C. Overexpression of OsEm1 encoding a group I LEA protein confers enhanced drought tolerance in rice. Biochem. Biophys. Res. Commun. 2016, 478, 703–709. [Google Scholar] [CrossRef]
- Hakata, M.; Kuroda, M.; Ohsumi, A.; Hirose, T.; Nakamura, H.; Muramatsu, M.; Ichikawa, H.; Yamakawa, H. Overexpression of a rice TIFY gene increases grain size through enhanced accumulation of carbohydrates in the stem. Biosci. Biotechnol. Biochem. 2012, 76, 2129–2134. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.J.; Kim, Y.S.; Kwon, C.W.; Park, H.K.; Jeong, J.S.; Kim, J.K. Overexpression of the transcription factor AP37 in rice improves grain yield under drought conditions. Plant Physiol. 2009, 150, 1368–1379. [Google Scholar] [CrossRef] [PubMed]
- Luo, B.; Chen, J.; Zhu, L.; Liu, S.; Li, B.; Lu, H.; Ye, G.; Xu, G.; Fan, X. Overexpression of a high-affinity nitrate transporter OsNRT2.1 increases yield and manganese accumulation in rice under alternating wet and dry condition. Front. Plant Sci. 2018, 9, 1192. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Chen, W.; Dong, J.; Li, J.; Yang, F.; Wu, Z.; Zhou, H. Overexpression of OsmiR164b-resistant OsNAC2 improves plant architecture and grain yield in rice. J. Exp. Bot. 2018, 69, 1533–1543. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, D.; Arnon, D. Physiological aspects of availability of nutrients for plant growth. Soil Sci. 1941, 51, 431–444. [Google Scholar] [CrossRef]
- Lin, Y.J.; Zhang, Q. Optimising the tissue culture conditions for high efficiency transformation of indica rice. Plant Cell Rep. 2005, 23, 540–547. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Singh, A.P.; Dixit, G.; Mishra, S.; Dwivedi, S.; Tiwari, M.; Mallick, S.; Pandey, V.; Trivedi, P.K.; Chakrabarty, D.; Tripathi, R.D. Salicylic acid modulates arsenic toxicity by reducing its root to shoot translocation in rice (Oryza sativa L.). Front. Plant Sci. 2015, 6, 340. [Google Scholar] [CrossRef]
- Mousavi, S.R.; Niknejad, Y.; Fallah, H.; Tari, D.B. Methyl jasmonate alleviates arsenic toxicity in rice. Plant Cell Rep. 2020, 39, 1041–1060. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, S.; Dillenburg, L.C. Measurements of Leaf Relative Water Content in Araucaria angustifolia. R. Bras. Fisiol. Veg. 1999, 11, 69–75. [Google Scholar]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998, 135, 1–9. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Meth. Enzymol. 1987, 148, 350–382. [Google Scholar]
- Holá, D.; Benešová, M.; Honnerová, J.; Hnilička, F.; Rothová, O.; Kočová, M.; Hniličková, H. The evaluation of photosynthetic parameters in maize inbred lines subjected to water deficiency: Can these parameters be used for the prediction of performance of hybrid progeny? Photosynthetica 2010, 48, 545–558. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dey, P.M. Oligosaccharides. In Methods in Plant Biochemistry, Carbohydrates; Dey, P.M., Ed.; Academic Press: London, UK, 1990; Volume 2, pp. 189–218. [Google Scholar]
- Bates, L.; Waldren, P.P.; Teare, J.D. Rapid determination of free proline of water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Zieslin, N.; Ben-Zaken, R. Peroxidase activity and presence of phenolic substances in peduncles of rose flowers. Plant Physiol. Biochem. 1993, 31, 333–339. [Google Scholar]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Kono, Y. Generation of superoxide radical during autoxidation of hydroxylamine and an assay for superoxide dismutase. Arch. Biochem. Biophys. 1978, 186, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. Meth. Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Putter, J.; Becker, R. Methods of Enzymatic Analysis; Academic Press: New York, NY, USA, 1974; p. 685. [Google Scholar]
- Vighi, I.L.; Benitez, L.C.; Amaral, M.N.; Moraes, G.P.; Auler, P.A.; Rodrigues, G.S.; Deuner, S.; Maia, L.C.; Braga, E.J.B. Functional characterization of the antioxidant enzymes in rice plants exposed to salinity stress. Biol. Plant. 2017, 61, 1–11. [Google Scholar] [CrossRef]
- Hwang, O.J.; Back, K. Suppression of Rice Cryptochrome 1b Decreases Both Melatonin and Expression of Brassinosteroid Biosynthetic Genes Resulting in Salt Tolerance. Molecules 2021, 26, 1075. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment As (µM) | Rice Line | Plant Height (cm) | Root Fresh Weight (mg) | Root Dry Weight (mg) | Shoot Fresh Weight (mg) | Shoot Dry Weight (mg) |
|---|---|---|---|---|---|---|
| Normal (0) | WT | 28.2 ± 0.87 a | 8.9 ± 0.30 a | 1.5 ± 0.17 a | 24.8 ± 0.49 a | 4.9 ± 0.27 a |
| OX-5 | 28.4 ± 0.88 a | 9.0 ± 0.29 a | 1.7 ± 0.13 a | 25.1 ± 0.46 a | 5.1 ± 0.32 a | |
| OX-9 | 28.3 ± 0.85 a | 9.1 ± 0.34 a | 1.6 ± 0.14 a | 25.0 ± 0.45 a | 5.0 ± 0.27 a | |
| Ri-2 | 28.2 ± 0.84 a | 8.8 ± 0.31 a | 1.5 ± 0.15 a | 24.7 ± 0.48 a | 5.0 ± 0.28 a | |
| Ri-8 | 28.1 ± 0.81 a | 8.7 ± 0.30 a | 1.6 ± 0.14 a | 24.8 ± 0.47 a | 4.8 ± 0.26 a | |
| 25 | WT | 20.1 ± 0.91 e | 5.0 ± 0.33 b | 0.8 ± 0.13 c | 15.9 ± 0.48 d | 3.1 ± 0.29 c |
| OX-5 | 26.5 ± 0.77 b | 8.7 ± 0.28 a | 1.1 ± 0.13 b | 22.4 ± 0.61 b | 4.1 ± 0.32 b | |
| OX-9 | 26.2 ± 0.81 b | 8.6 ± 0.31 a | 1.1 ± 0.16 b | 22.1 ± 0.58 b | 4.0 ± 0.25 b | |
| Ri-2 | 17.2 ± 0.83 f | 3.2 ± 0.32 c | 0.5 ± 0.14 d | 11.9 ± 0.51 e | 2.3 ± 0.23 d | |
| Ri-8 | 17.3 ± 0.84 f | 3.3 ± 0.30 c | 0.5 ± 0.12 d | 11.8 ± 0.55 e | 2.4 ± 0.24 d | |
| 50 | WT | 16.1 ± 0.74 g | 3.5 ± 0.32 c | 0.6 ± 0.10 d | 12.1 ± 0.44 e | 2.5 ± 0.21 d |
| OX-5 | 23.9 ± 0.91 c | 4.8 ± 0.33 b | 0.8 ± 0.15 c | 19.0 ± 0.48 c | 3.4 ± 0.31 c | |
| OX-9 | 21.9 ± 0.88 d | 4.6 ± 0.29 b | 0.8 ± 0.14 c | 18.7 ± 0.52 c | 3.3 ± 0.29 c | |
| Ri-2 | 13.4 ± 0.83 h | 2.1 ± 0.29 d | 0.3 ± 0.15 e | 08.1 ± 0.48 f | 1.7 ± 0.27 e | |
| Ri-8 | 13.8 ± 0.81 h | 2.2 ± 0.27 d | 0.3 ± 0.14 e | 08.0 ± 0.52 f | 1.6 ± 0.30 e |
| Treatment As (µM) | Rice Line | Panicle Length (cm) | Number of Spikelets per Panicle | Total Number of Spikelets per Hill | Number of Filled Grains Per Hill | Filling Rate (%) | Total Grain Weight (g) |
|---|---|---|---|---|---|---|---|
| Normal (0) | WT | 18.0 ± 1.08 a | 90.0 ± 7.98 a | 1190.7 ± 144.2 a | 1047.1 ± 121.2 a | 88.1 ± 11.42 a | 17.9 ± 1.22 a |
| OX-5 | 18.1 ± 1.12 a | 91.0 ± 8.03 a | 1186.4 ± 138.3 a | 1049.4 ± 122.5 a | 88.5 ± 12.11 a | 18.0 ± 1.30 a | |
| OX-9 | 18.0 ± 1.16 a | 89.5 ± 9.13 a | 1182.5 ± 132.4 a | 1052.3 ± 124.2 a | 89.4 ± 10.55 a | 18.1 ± 1.41 a | |
| Ri-2 | 18.1 ± 1.15 a | 89.2 ± 9.11 a | 1184.7 ± 141.6 a | 1051.4 ± 121.6 a | 88.9 ± 10.73 a | 17.9 ± 1.32 a | |
| Ri-8 | 17.9 ± 1.14 a | 91.1 ± 9.11 a | 1193.1 ± 141.1 a | 1043.9 ± 131.3 a | 89.8 ± 11.39 a | 17.8 ± 1.21 a | |
| 25 | WT | 14.0 ± 1.21 c | 80.2 ± 8.11 b | 1095.4 ± 131.2 c | 799.2 ± 128.9 d | 70.2 ± 11.21 d | 14.1 ± 1.32 c |
| OX-5 | 17.9 ± 1.30 a | 89.3 ± 8.36 a | 1181.1 ± 140.7 a | 984.4 ± 135.1 b | 82.3 ± 11.35 b | 17.6 ± 1.28 a | |
| OX-9 | 17.8 ± 1.25 a | 88.4 ± 8.12 a | 1179.3 ± 134.8 a | 978.6 ± 133.7 b | 81.7 ± 10.29 b | 17.8 ± 1.26 a | |
| Ri-2 | 10.2 ± 1.13 e | 71.4 ± 7.88 c | 1017.5 ± 132.4 d | 711.5 ± 127.6 e | 62.1 ± 11.33 e | 11.2 ± 1.17 e | |
| Ri-8 | 10.4 ± 1.15 e | 73.6 ± 7.69 c | 1021.7 ± 126.4 d | 706.8 ± 124.2 e | 61.8 ± 11.45 e | 11.1 ± 1.13 e | |
| 50 | WT | 12.1 ± 1.13 d | 66.3 ± 8.10 d | 1021.5 ± 141.2 d | 621.8 ± 121.3 f | 54.7 ± 14.33 f | 12.7 ± 1.17 d |
| OX-5 | 16.1 ± 1.25 b | 79.3 ± 8.12 b | 1139.3 ± 127.6 b | 903.4 ± 122.4 c | 75.1 ± 14.38 c | 15.8 ± 1.21 b | |
| OX-9 | 15.9 ± 1.31 b | 80.1 ± 8.56 b | 1142.5 ± 131.2 b | 897.8 ± 113.5 c | 75.8 ± 12.47 c | 15.2 ± 1.32 b | |
| Ri-2 | 08.9 ± 1.32 f | 53.7 ± 8.51 e | 0944.8 ± 132.4 e | 535.3 ± 122.3 g | 43.4 ± 14.19 g | 10.1 ± 1.31 f | |
| Ri-8 | 09.1 ± 1.36 f | 54.1 ± 8.56 e | 0950.4 ± 132.2 e | 540.3 ± 125.8 g | 42.6 ± 14.21 g | 10.2 ± 1.34 f |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pooam, M.; El-Ballat, E.M.; Jourdan, N.; Ali, H.M.; Hano, C.; Ahmad, M.; El-Esawi, M.A. SNAC3 Transcription Factor Enhances Arsenic Stress Tolerance and Grain Yield in Rice (Oryza sativa L.) through Regulating Physio-Biochemical Mechanisms, Stress-Responsive Genes, and Cryptochrome 1b. Plants 2023, 12, 2731. https://doi.org/10.3390/plants12142731
Pooam M, El-Ballat EM, Jourdan N, Ali HM, Hano C, Ahmad M, El-Esawi MA. SNAC3 Transcription Factor Enhances Arsenic Stress Tolerance and Grain Yield in Rice (Oryza sativa L.) through Regulating Physio-Biochemical Mechanisms, Stress-Responsive Genes, and Cryptochrome 1b. Plants. 2023; 12(14):2731. https://doi.org/10.3390/plants12142731
Chicago/Turabian StylePooam, Marootpong, Enas M. El-Ballat, Nathalie Jourdan, Hayssam M. Ali, Christophe Hano, Margaret Ahmad, and Mohamed A. El-Esawi. 2023. "SNAC3 Transcription Factor Enhances Arsenic Stress Tolerance and Grain Yield in Rice (Oryza sativa L.) through Regulating Physio-Biochemical Mechanisms, Stress-Responsive Genes, and Cryptochrome 1b" Plants 12, no. 14: 2731. https://doi.org/10.3390/plants12142731
APA StylePooam, M., El-Ballat, E. M., Jourdan, N., Ali, H. M., Hano, C., Ahmad, M., & El-Esawi, M. A. (2023). SNAC3 Transcription Factor Enhances Arsenic Stress Tolerance and Grain Yield in Rice (Oryza sativa L.) through Regulating Physio-Biochemical Mechanisms, Stress-Responsive Genes, and Cryptochrome 1b. Plants, 12(14), 2731. https://doi.org/10.3390/plants12142731