
Dietary Supplementation with Yerba Mate (Ilex paraguariensis) Infusion Increases IRS-1 and PI3K mRNA Levels and Enhances Insulin Sensitivity and Secretion in Rat Pancreatic Islets
 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results
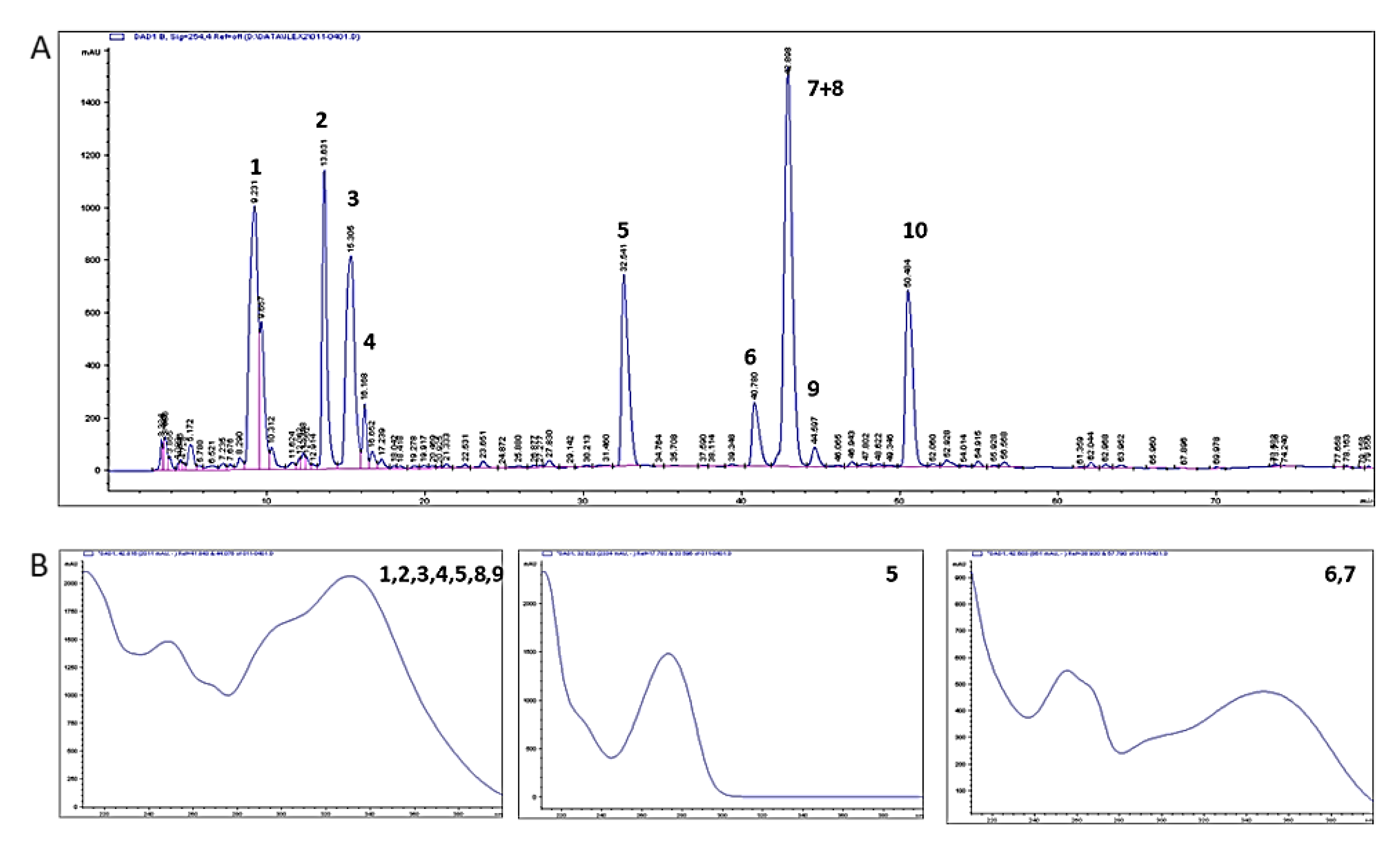
2.1. Composition of the Aqueous Extract of Yerba Mate
2.2. Body Weight, Food Intake, and Plasma Parameters
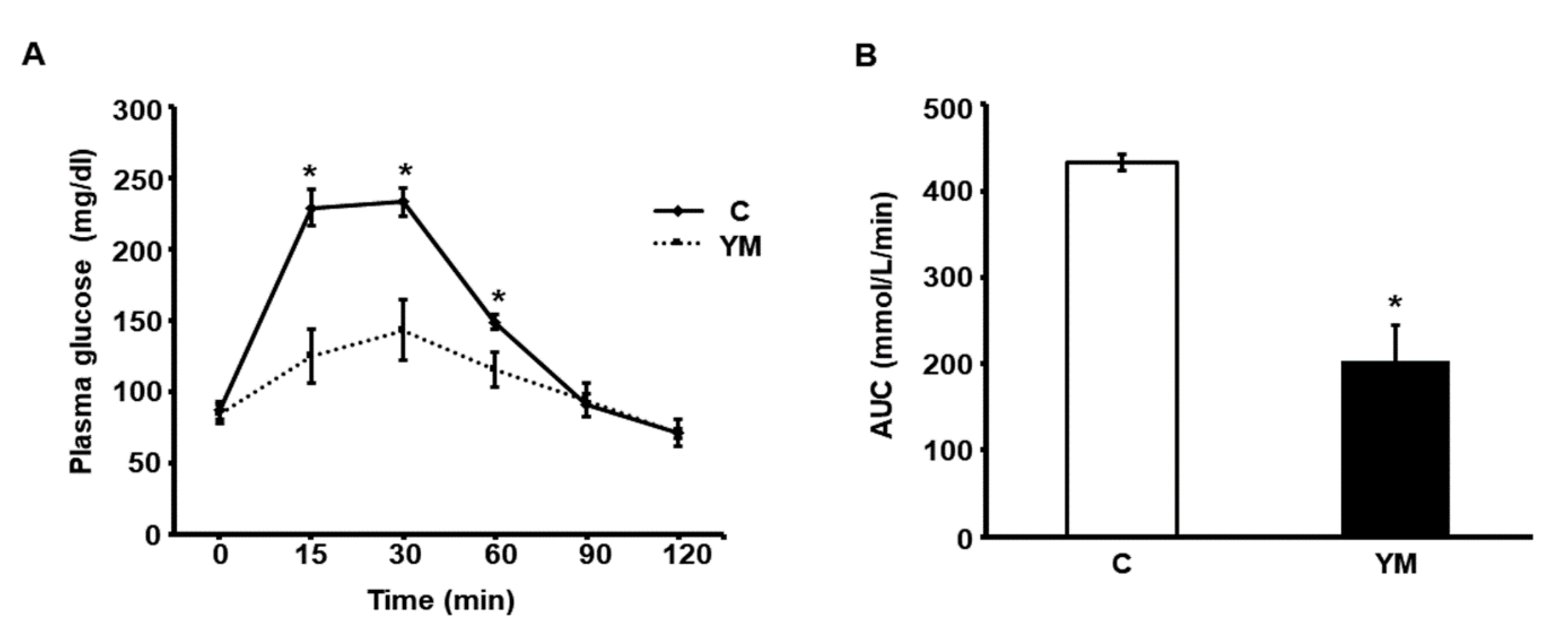
2.3. Glucose Tolerance Test
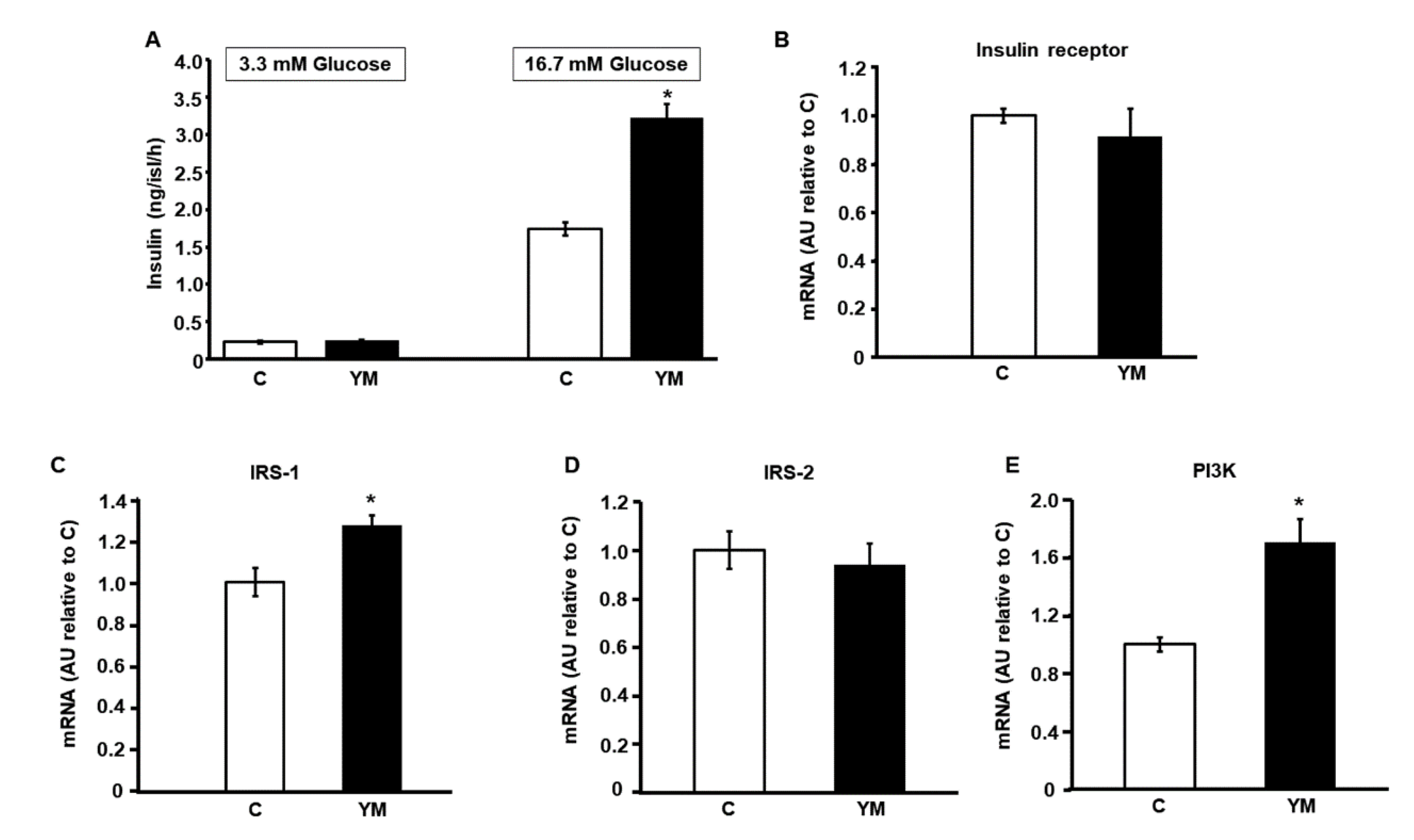
2.4. Glucose Stimulated Insulin Secretion
2.5. Islet mRNA Levels of Intracellular Signaling Proteins of Insulin Pathway
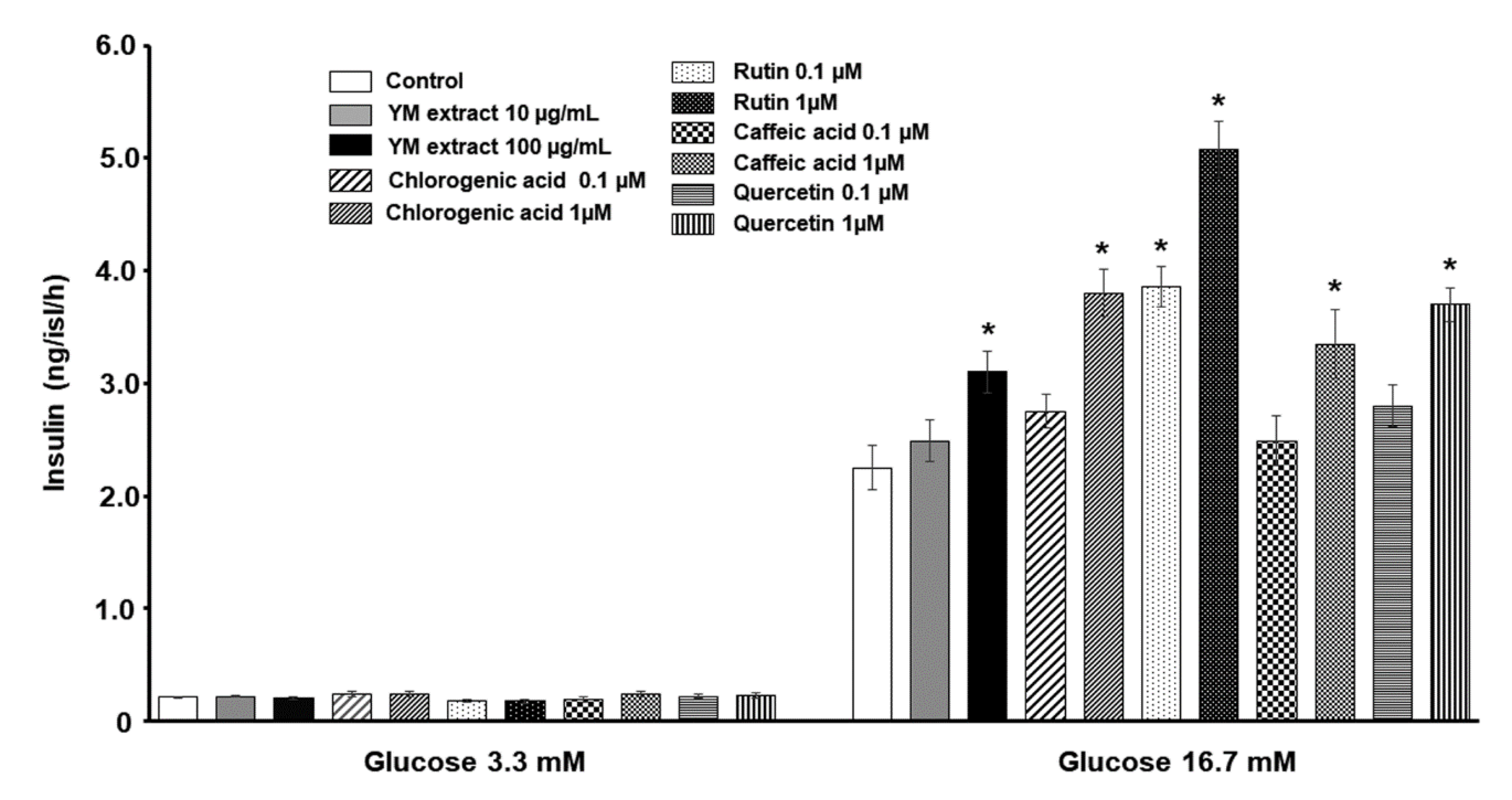
2.6. Direct Effect of Yerba Mate and Its Phenolic Components on Ex-Vivo Insulin Secretion of Untreated Pancreatic Islets
2.7. Gene Expression (mRNA Levels) of Inflammatory Mediators in Pancreatic Islets
3. Discussion
4. Materials and Methods
4.1. Chemicals and Drugs
4.2. Plant Material and Extraction
4.3. Experimental Animals
4.4. Chemical Analysis of Yerba Mate Aqueous Extract
4.5. Glucose Tolerance Test
4.6. Biochemical Analyses of Plasma
4.7. Glucose Stimulated Insulin Secretion
4.8. Gene Expression by Real-Time PCR (qPCR)
4.9. Statistical Data Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schinella, G.R.; Troiani, G.; Dávila, V.; De Buschiazzoand, P.M.; Tournier, H.A. Antioxidant effects of an aqueous extract of Ilex paraguariensis. Biochem. Biophys. Res. Commun. 2000, 269, 357–360. [Google Scholar] [CrossRef]
- Schinella, G.; Fantinelli, J.C.; Mosca, S.M. Cardioprotective effects of Ilex paraguariensis extract: Evidence for a nitric oxide-dependent mechanism. Clin. Nutr. 2005, 24, 360–366. [Google Scholar] [CrossRef]
- González Arbeláez, L.F.; Fantinelli, J.C.; Ciocci Pardo, A.; Caldiz, C.I.; Ríos, J.L.; Schinella, G.R.; Mosca, S.M. Effect of an Ilex paraguariensis (yerba mate) extract on infarct size in isolated rat hearts: The mechanisms involved. Food Funct. 2016, 7, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Schinella, G.; Neyret, E.; Console, G.; Tournier, H.; Prieto, J.M.; Rios, J.L.; Giner, R.M. An aqueous extract of Ilex paraguariensis reduces carrageenan-induced edema and inhibits the expression of cyclooxygenase-2 and inducible nitric oxide synthase in animal models of inflammation. Plant. Med. 2014, 80, 961–968. [Google Scholar] [CrossRef]
- Heckand, C.I.; De Mejia, E.G. Yerba Mate Tea (Ilex paraguariensis): A comprehensive review on chemistry, health implications, and technological considerations. J. Food Sci. 2007, 72, R138–R151. [Google Scholar]
- Bracesco, N.; Sanchez, A.G.; Contreras, V.; Meniniand, T.; Gugliucci, A. Recent advances on Ilex paraguariensis research: Minireview. J. Ethnopharmacol. 2011, 136, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Andújar, I.; Schinella, G.; Rios, J.L. Yerba Mate (Ilex paraguariensis): A psycho-stimulant plant with anti-oxidant properties. In Polyphenols: Food Sources, Bioactive Properties and Antioxidant Effects; Nova Science Publishers: New York, NY, USA, 2014; pp. 51–86. [Google Scholar]
- Choi, M.S.; Park, H.J.; Kim, S.R.; Kimand, D.Y.; Jung, U.J. Long-term dietary supplementation with yerba mate ameliorates diet-induced obesity and metabolic disorders in mice by regulating energy expenditure and lipid metabolism. J. Med. Food 2017, 20, 1168–1175. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.M.; Freitas, H.S.; Souza, M.F.F.; Açari, D.P.; Ribeiro, M.L.; Carvalhoand, P.O.; Bastos, D.H.M. Yerba mate (Ilex paraguariensis) aqueous extract decreases intestinal SGLT1 gene expression but does not affect other biochemical parameters in alloxan-diabetic Wistar rats. J. Agric. Food Chem. 2008, 56, 10527–10532. [Google Scholar] [CrossRef]
- Kang, Y.-R.; Lee, H.-Y.; Kim, J.-H.; Moon, D.-I.; Seo, M.-Y.; Park, S.-H.; Choi, K.-H.; Kim, C.-R.; Kim, S.-H.; Oh, J.-H.; et al. Anti-obesity and anti-diabetic effects of Yerba Mate (Ilex paraguariensis) in C57BL/6J mice fed a high-fat diet. Lab. Anim. Res. 2012, 28, 23–29. [Google Scholar] [CrossRef]
- Arçari, D.P.; Bartchewsky, W.; Dos Santos, T.W.; Oliveira, K.A.; Funck, A.; Pedrazzolli, J.; De Souza, M.F.; Saad, M.J.; Bastos, D.H.; Gambero, A.; et al. Antiobesity effects of yerba maté extract (Ilex paraguariensis) in high-fat diet-induced obese mice. Obesity 2009, 17, 2127–2133. [Google Scholar] [CrossRef]
- Hussein, G.M.; Matsuda, H.; Nakamura, S.; Akiyama, T.; Tamuraand, K.; Yoshikawa, M. Protective and ameliorative effects of maté (Ilex paraguariensis) on metabolic syndrome in TSOD mice. Phytomedicine 2011, 19, 88–97. [Google Scholar] [CrossRef] [PubMed]
- De Resende, P.E.; Kaiser, S.; Pittol, V.; Hoefel, A.L.; D’Agostini Silva, R.; Vieira Marques, C.; Kucharski, L.C.; Ortega, G.G. Influence of crude extract and bioactive fractions of Ilex paraguariensis A. St. Hil. (yerba mate) on the Wistar rat lipid metabolism. J. Funct. Foods 2015, 15, 440–451. [Google Scholar] [CrossRef]
- Keller, A.C.; Ma, J.; Kavalier, A.; He, K.; Brillantes, A.M.; Kennelly, E.J. Saponins from the traditional medicinal plant Momordica charantia stimulate insulin secretion in vitro. Phytomedicine 2011, 19, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Mu, S.; Chen, W.; Liu, S.; Cong, Y.; Liu, J.; Jia, N. Saponins of Momordica charantia increase insulin secretion in INS-1 pancreatic β-cells via the PI3K/Akt/FoxO1 signaling pathway. Endocrinol. Diabetes Nutr. 2021, 68, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Pereira, D.F.; Kappel, V.D.; Cazarolli, L.H.; Boligon, A.A.; Athayde, M.L.; Guesser, S.M.; Da Silva, E.L.; Silva, F.R. Influence of the traditional Brazilian drink Ilex paraguariensis tea on glucose homeostasis. Phytomedicine 2012, 19, 868–877. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.E.; Janson, J.; Bonner-Weir, S.; Ritzel, R.; Rizza, R.A.; Butler, P.C. Beta-cell deficit and increased beta-cell apoptosis in humans with type 2 diabetes. Diabetes 2003, 52, 102–110. [Google Scholar] [CrossRef]
- Khan, D.; Moffet, C.R.; Flatt, P.R.; Kelly, C. Role of islet peptides in beta cell regulation and type 2 diabetes therapy. Peptides 2018, 100, 212–218. [Google Scholar] [CrossRef]
- English, A.; Irwin, N. Nonclassical Islet Peptides: Pancreatic and Extrapancreatic Actions. Clin. Med. Insights Endocrinol. Diabetes 2019, 12, 1179551419888871. [Google Scholar] [CrossRef] [PubMed]
- Klein, G.A.; Stefanuto, A.; Boaventura, B.C.; De Morais, E.C.; Cavalcante Lda, S.; De Andrade, F.; Wazlawik, E.; Di Pietro, P.F.; Maraschinand, M.; Da Silva, E.L. Mate tea (Ilex paraguariensis) improves glycemic and lipid profiles of type 2 diabetes and pre-diabetes individuals: A pilot study. J. Am. Coll. Nutr. 2011, 30, 320–332. [Google Scholar] [CrossRef]
- Boaventura, B.C.B.; Di Pietro, P.F.; Stefanuto, A.; Klein, G.A.; de Morais, E.C.; de Andrade, F.; Wazlawikand, E.; da Silva, E.L. Association of mate tea (Ilex paraguariensis) intake and dietary intervention and effects on oxidative stress biomark ers of dyslipidemic subjects. Nutrition 2012, 28, 657–664. [Google Scholar] [CrossRef]
- Boaventura, B.C.B.; Di Pietro, P.F.; Klein, G.A.; Stefanuto, A.; de Morais, E.C.; de Andrade, F.; Wazlawik, E.; da Silva, E.L. Antioxidant potential of mate tea (Ilex paraguariensis) in type 2 diabetic mellitus and pre-diabetic individuals. J. Funct. Foods 2013, 5, 1057–1064. [Google Scholar] [CrossRef]
- Bravo, L.; Goya, L.; Lecumberri, L.E. LC/MS characterization of phenolic constituents of mate (Ilex paraguariensis, St. Hil.) and its antioxidant activity compared to commonly consumed beverages. Food Res. Int. 2007, 40, 393–405. [Google Scholar] [CrossRef]
- Borelli, M.I.; Francini, F.; Gagliardino, J.J. Autocrine regulation of glucose metabolism in pancreatic islets. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E111–E115. [Google Scholar] [CrossRef] [PubMed]
- Puangpraphant, S.; De Mejia, E.G. Saponins in yerba mate tea (Ilex paraguariensis A. St.-Hil) and quercetin synergistically inhibit iNOS and COX-2 in lipopolysaccharide-induced macrophages through NFkappaB pathways. J. Agric. Food Chem. 2009, 57, 8873–8883. [Google Scholar] [CrossRef]
- Zhang, S.; Kim, K.H. TNF-alpha inhibits glucose-induced insulin secretion in a pancreatic beta-cell line (INS-1). FEBS Lett. 2016, 377, 237–239. [Google Scholar] [CrossRef]
- Kaji, H. Adipose tissue-derived plasminogen activator inhibitor-1 function and regulation. Compr. Physiol. 2016, 6, 1873–1896. [Google Scholar]
- Samad, F.; UysaK, K.I.; Wiesbrock, S.M.; Pandey, M.; Hotamisligiand, G.S.; Loskutoff, D.J. Tumor necrosis factor alpha is a key component in the obesity-linked elevation of plasminogen activator inhibitor 1. Proc. Natl. Acad. Sci. USA 1999, 96, 6902–6907. [Google Scholar] [CrossRef]
- Borst, S.E. The role of TNF-α in insulin resistance. Endocrine 2004, 23, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Lanzetti, M.; Bezerra, F.S.; Romana-Souza, B.; Brando-Lima, A.C.; Koatz, V.L.; Portoand, L.C.; Valenca, S.S. Mate tea reduced acute lung inflammation in mice exposed to cigarette smoke. Nutrition 2008, 24, 375–381. [Google Scholar] [CrossRef]
- Pimentel, G.D.; Lira, F.S.; Rosa, J.C.; Caris, A.V.; Pinheiro, F.; Ribeiro, E.B.; Oller do Nascimento, C.M.; Oyama, L.M. Yerba mate extract (Ilex paraguariensis) attenuates both central and peripheral inflammatory effects of diet-induced obesity in rats. J. Nutr. Biochem. 2013, 24, 809–818. [Google Scholar] [CrossRef]
- Muñoz-Culla, M.; Sáenz-Cuesta, M.; Guereca-Barandiaran, M.J.; Ribeiroand, M.L.; Otaegui, D. Yerba mate (Ilex paraguariensis) inhibits lymphocyte activation in vitro. Food Funct. 2016, 7, 4556–4563. [Google Scholar] [CrossRef] [PubMed]
- Luz, A.B.G.; Da Silva, C.H.B.; Nascimento, M.V.P.S.; De Campos Facchin, B.M.; Baratto, B.; Fröde, T.S.; Reginatto, F.H.; Dalmarco, E.M. The anti-inflammatory effect of Ilex paraguariensis A. St. Hil (Mate) in a murine model of pleurisy. Int. Immunopharmacol. 2016, 36, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Bardy, G.; Virsolvy, A.; Quignard, J.F.; Ravier, M.A.; Bertrand, G.; Dalle, S.; Cros, G.; Magous, R.; Richard, S.; Oiry, C. Quercetin induces insulin secretion by direct activation of L-type calcium channels in pancreatic beta cells. Br. J. Pharmacol. 2013, 169, 1102–1113. [Google Scholar] [CrossRef]
- Youl, E.; Bardy, G.; Magous, R.; Cros, G.; Sejalon, F.; Virsolvy, A.; Richard, S.; Quignard, J.F.; Gross, R.; Petit, P.; et al. Quercetin potentiates insulin secretion and protects INS-1 pancreatic β-cells against oxidative damage via the ERK1/2 pathway. Br. J. Pharmacol. 2010, 161, 799–814. [Google Scholar] [CrossRef]
- Kappel, V.D.; Frederico, M.J.; Postal, B.G.; Mendes, C.P.; Cazarolli, L.H.; Silva, F.R. The role of calcium in intracellular pathways of rutin in rat pancreatic islets: Potential insulin secretagogue effect. Eur. J. Pharmacol. 2013, 702, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Oksbjerg, N.; Young, J.F.; Jeppesen, P.B. Caffeic acid, naringenin and quercetin enhance glucose-stimulated insulin secretion and glucose sensitivity in INS-1E cells. Diabetes Obes. Metab. 2014, 16, 602–612. [Google Scholar] [CrossRef]
- Mesquita, M.; Santos, E.; Kassuya, C.A.; Salvador, M.J. Chimarrão, terere and mate-tea in legitimate technology modes of preparation and consume: A comparative study of chemical composition, antioxidant, anti-inflammatory and anti-anxiety properties of the mostly consumed beverages of Ilex paraguariensis St. Hil. J. Ethnopharmacol. 2021, 279, 114401. [Google Scholar]
- Lacy, P.E.; Kostianovsky, M. Method for the Isolation of Intact Islets of Langerhans from the Rat Pancreas. Diabetes 1967, 6, 35–39. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasma Parameters | C | YM |
|---|---|---|
| Glucose (mg/dL) | 115.14 ± 2.35 | 108.62 ± 3.43 |
| Triglyceride (mg/dL) | 100 ± 7.5 | 87 ± 8.4 |
| Insulin (ng/mL) | 0.39 ± 0.03 | 0.44 ± 0.08 |
| FRAP (µmol Fe2+/L) | 255 ± 7.5 | 412 ± 17 * |
| TBARS (pmol/mg prot) | 598 ± 21 | 341 ± 33 * |
| HOMA-IR | 2.63 ± 0.27 | 2.77 ± 0.48 |
| HOMA-β | 11.59 ± 1.85 | 15.86 ± 6.77 |
| Gene | GeneBank | Sequences |
|---|---|---|
| Insulin receptor | NM_017071 | Fw 5′-ATATTGACCCGCCCCAGAGG-3′Rv 5′-TAGGTCCGGCGTTCATCAGA-3′ |
| IRS-1 | NM_012969 | Fw 5′-TGTGCCAAGCAACAAGAAAG-3′Rv 5′-ACGGTTTCAGAGCAGAGGAA-3′ |
| IRS-2 | NM_001168633.1 | Fw 5′-CTACCCACTGAGCCCAAGAG-3′Rv 5′-CCAGGGATGAAGCAGGACTA-3′ |
| PI3K | NM_053481 | Fw 5′-GGTTGTTGTTGCCCCAGAC-3′Rv 5′-GGTTGTTGTTGCCCCAGAC-3′ |
| TNF-α | NM_0.126675.3 | Fw 5′-GGCATGGATCTCAAAGACAACC-3′Rv 5′-CAAATCGGCTGACGGTGTG- 3′ |
| PAI-1 | NM_012620.1 | Fw 5′-CCACGGTGAAGCAGGTGGACT-3′Rv 5′-TGCTGGCCTCTAAGAAGGGG- 3′ |
| β-actin | NM_031144.3 | Fw 5′-AGAGGGAAATCGTGCGTGAC-3′Rv 5′-CGATAGTGATGACCTGACCGT-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maiztegui, B.; Villagarcía, H.G.; Román, C.L.; Flores, L.E.; Prieto, J.M.; Castro, M.C.; Massa, M.L.; Schinella, G.R.; Francini, F. Dietary Supplementation with Yerba Mate (Ilex paraguariensis) Infusion Increases IRS-1 and PI3K mRNA Levels and Enhances Insulin Sensitivity and Secretion in Rat Pancreatic Islets. Plants 2023, 12, 2620. https://doi.org/10.3390/plants12142620
Maiztegui B, Villagarcía HG, Román CL, Flores LE, Prieto JM, Castro MC, Massa ML, Schinella GR, Francini F. Dietary Supplementation with Yerba Mate (Ilex paraguariensis) Infusion Increases IRS-1 and PI3K mRNA Levels and Enhances Insulin Sensitivity and Secretion in Rat Pancreatic Islets. Plants. 2023; 12(14):2620. https://doi.org/10.3390/plants12142620
Chicago/Turabian StyleMaiztegui, Bárbara, Hernán Gonzalo Villagarcía, Carolina Lisi Román, Luis Emilio Flores, José María Prieto, María Cecilia Castro, María Laura Massa, Guillermo R. Schinella, and Flavio Francini. 2023. "Dietary Supplementation with Yerba Mate (Ilex paraguariensis) Infusion Increases IRS-1 and PI3K mRNA Levels and Enhances Insulin Sensitivity and Secretion in Rat Pancreatic Islets" Plants 12, no. 14: 2620. https://doi.org/10.3390/plants12142620
APA StyleMaiztegui, B., Villagarcía, H. G., Román, C. L., Flores, L. E., Prieto, J. M., Castro, M. C., Massa, M. L., Schinella, G. R., & Francini, F. (2023). Dietary Supplementation with Yerba Mate (Ilex paraguariensis) Infusion Increases IRS-1 and PI3K mRNA Levels and Enhances Insulin Sensitivity and Secretion in Rat Pancreatic Islets. Plants, 12(14), 2620. https://doi.org/10.3390/plants12142620