1. Introduction
Psittacanthus calyculatus is commonly known as Mexican mistletoe; it classified as a hemiparasite plant since it synthesizes its own chlorophyll, although it depends on its hosts to germinate and feed by taking vital substances from them through a modified root called the haustorium [
1,
2].
The seeds are dispersed by frugivorous birds, which deposit them on the branches of their hosts, to which they adhere by means of a mucilaginous substance called viscin [
3].
P. calyculatus is the most widely distributed species within the phyla Loranthaceae, with a wide distribution from Baja California, Sonora and Tamaulipas from Mexico to northern Argentina; it also has a wide range of hosts, such as trees, shrubs and cacti, both gymnosperms and angiosperms, among which are
Quercus spp. (oak),
Acacia spp.,
Juglans spp. (walnut),
Ficus spp.,
Populus spp.,
Salix spp. (willow),
Prosopis spp. (mesquite),
Prunus spp. (peach, capulin) and
Persea americana [
1,
3,
4]. Due to its great affinity with forest and fruit trees, Mexican mistletoe is considered a weed, so there is interest in developing sustainable management strategies, since existing ones (e.g., pruning, herbicides) are neither sufficient nor effective [
4,
5,
6,
7].
P. calyculatus has been used as a medicinal plant in traditional Mexican medicine, applied topically or by means of infusions to treat ailments such as fevers, infertility or diabetes, among other conditions [
8,
9,
10]. Recent studies in the area of pharmacology have reported that its foliage has coadjuvant effects in the treatment of arterial hypertension, since the substances present in its foliage act as vasodilators, lower cholesterol and blood sugar levels; they also display a cytotoxic effect against cancer cells [
11,
12,
13,
14].
Studies on other genera or species of the Loranthaceae family have shown the therapeutic potential of the different tissues of parasitic species [
15,
16,
17]. Serrano et al. [
15] reported the content of phenolic compounds and antioxidant capacity of the colored fruit of
Cladocolea loniceroides, a species endemic to Mexico.
Regarding the other species of the genus
Psittacanthus, which are endemic to other countries, the antimicrobial activity of
P. linearis fruits, as well as the phytochemical content and antioxidant capacity of the leaves of
P. plagiofilus and
P. cucullaris have been reported, using different extractive solvents such as ethanol and water [
18,
19,
20]
Therefore, the objective of this study was to determine the phenolic content and biological activities of the ripe fruits of P. calyculatus, since there are no studies demonstrating its potential benefits whose knowledge could contribute to the proposal of sustainable strategies for its control, as well as to the diversification of sources of secondary metabolites as the active ingredients of drugs and food supplements, among other things.
2. Results and Discussion
2.1. UV–Visible Spectrophotometry Analysis
2.1.1. Total Phenol Quantification
A significant effect of the host
P. laevigata and the extractive solvent on the TPC was observed (
Table 1). Thus, the highest content of total phenols in fruits was obtained when the host was
P. laevigata and the solvent used was acidified ethanol, being between 80.3% and 108% higher than the rest of the extracts evaluated.
There is a study on other parasitic species,
P. plagiophyllus, in which an ethanol–water combination was used in different proportions as extractive solvent to achieve a higher yield [
18]. This is the first phytochemical study of
P. calyculatus that performs a phytochemical analysis of the fruit. Serrano et al. [
15], evaluated the phenol and flavonoid content of
C. loniceroides (Loranthaceae family), and the authors reported higher values of fruit total polyphenol content (189.5 mg EAG/g DW) than those obtained in this study (71.396 mg EAG/g DW).
2.1.2. Total Flavonoids Quantification
In contrast to the total phenol content, the TFC of the host–host interaction and extractive solvent showed a significant effect on the flavonoid content of these extracts. On the other hand, the highest TFC was shown via the extracts with acidified ethanol when the host was
Q. deserticola, with a higher content (14.232 EQ mg/g DW) being between 25% and 29.5% higher than the rest of the extracts evaluated (
Table 1).
Studies of parasitic species of the Loranthaceae family have been carried out on the foliage, identifying flavonoids such as rutin, quercetin and hesperidin, as well as the presence of alkaloids, anthraquinones and steroids [
12,
13,
18,
19].
The study performed by Serrano et al. [
15], mentioned above, evaluated the flavonoid content of
C. loniceroides fruits as well, and the authors found higher contents of these compounds in the pigmented fruit (36.45 mg Rutin E/g DW) than in other parts of the plant. The reported values were higher than those found in the present work, which could be due to the use of rutin as a standard instead of quercetin.
2.1.3. Total Anthocyanins Content
The highest TAC was observed in fruits from
P. calyculatus when the host was
Q. deserticola and the solvent used was ethanol, being between 219% and 196% higher than the rest of the extracts. In general, a higher recovery of metabolites was achieved using acidified ethanol (
Table 1).
This is the first study to date to report on the total anthocyanin content of
P. calyculatus fruits; existing studies address the phytochemistry of leaves or unpigmented fruits belonging to other families or species [
15,
18,
19,
21]. However, the properties of pigmented berries such as strawberries, blueberries, blackberries, grapes and other red fruits have been extensively studied for their neuroprotective, anti-inflammatory and antihyperglycemic effects in various applications [
22,
23,
24,
25,
26]. Therefore, it is relevant to investigate the phytochemistry and biological properties of
P. calyculatus fruit, since it could be a potential source of metabolites of interest, which in turn would promote the use and conservation of important forest resources [
2,
5,
27].
It is important to note that certain physical and chemical modifications that the parasite undergoes have been reported by several authors, suggesting that these are the result of the influence of various biotic or abiotic factors, and especially emphasizing that the amount and content of metabolites depends mostly on the host [
2,
5,
28,
29].
It seems that the parasitic organism is dependent on its host to acquire the nutrients necessary for its survival, triggering a chemical host response, which, in this case, added value to the P. calyculatus fruit when considering the host P. laevigata as a promoter of the accumulation of fruit phenolic compounds and Q. deserticola as a promoter of the accumulation of fruit flavonoids and anthocyanins.
2.1.4. Antioxidant Activity
A significant effect of the host, the extractive solvent and their interactions was found on the antioxidant capacity of ethanolic extracts of
P. calyculatus fruits determined via the DPPH
● method, while the antioxidative capacity determined using the ABTS
•+ method was significantly influenced through the host and extractive solvent (
Table 2).
In the DPPH● assay, the highest activity was shown via the acidified extracts of both hosts. The ABTS•+ assay showed a significantly higher antioxidant activity in the acidified extract of the host P. laevigata (141.6–104.7%) compared to the rest of the extracts.
The host species of P. calyculatus showed no significant effect on the antioxidant activity of the ethanolic extracts of its fruits, determined via the ABTS•+ method, while the extractive solvent significantly influenced this biological activity in the extracts measured via the DPPH● method.
No studies were found documenting the antioxidant activity of
P. calyculatus fruits. However, the antioxidant activity of leaves collected from different hosts (
Hedera helix,
Fraxinus uhdei and
P. laevigata) was reported by Zarza et al. [
11]. The authors observed a higher antioxidant activity in the aqueous extracts of leaves from specimens hosted by
P. laevigata (91.01% inhibition in the DPPH
● assay). In addition to detecting a greater diversity of phenolic compounds (e.g., (+)-catechin, gallic acid and p-coumaric acid).
The differences found in the activity of the different extracts could be due to the chemical changes promoted by biotic and abiotic stimuli, as discussed above.
2.1.5. MIC and MBC of Ethanolic Extracts of Pericarp of P. calyculatus
The lowest MIC and MBC values of the
P. calyculatus fruit pericarp (
Table 3) was shown via the acidified ethanol extracts for both hosts,
P. laevigata and
Q. deserticola. MIC and MBC were the same for all three enteropathogenic bacteria (
Salmonella, Escherichia and
Shigella). Antimicrobial studies on the genus
Psittacanthus exist; however, they have not evaluated different hosts. Bailladores et al. [
20] reported the antimicrobial activity of the pigmented fruit of the genus
P. linearis in relation to
S. aureus,
E. coli and
Pseudomonas aeruginosa, showing that only the ethanolic extract of the fruit was more active against the bacteria
S. aureus, being more sensitive to its metabolites content (polyphenols with a concentration of 300 µg/mL), while the ethanolic extracts of the
P. calyculatus fruit evaluated in the present study showed the same inhibitory and bactericidal activity against
S. choleraesuis ATCC 10708,
E. coli ATCC 12792, and
S. flexneri ATCC 12022, observing that these bacteria were more sensitive to the total content of anthocyanins, followed by the content of total phenols, based on the correlation analysis (shown in
Section 2.3).
There are no reports on the antimicrobial activity of
P. calyculatus fruits; however, the leaf extracts of the
P. linearis and
P. cucullaris species have been evaluated against Gram-negative and Gram-positive bacteria (
P. aeruginosa,
S. aureus and
E. coli), resulting in an inhibitory effect on these microorganisms [
19,
20].
2.1.6. Antihypertensive Activity
The highest inhibition of ACE was observed in the absolute ethanol extract of fruits from both hosts (
Table 4), significantly exceeding the acidified ethanol extracts of
P. laevigata and
Q. deserticola fruits (376% and 43.8%, respectively).
There is a patent reported by Cervantes et al. [
27] about the antihypertensive properties of aqueous extracts of
P. calyculatus leaves, which described the vasorelaxation tests performed on male guinea pigs of the Hartley strain, showing relaxation levels of 66–92% in the aortic rings. The results of this work also showed the antihypertensive potential of the
P. calyculatus fruit from the same host (
P. laevigata) through ACE inhibition (92.59%).
There is another study on the vasomotor responses of the aortic rings of roots, using the ethanol–water extraction of leaves of the mistletoe
P. calyculatus, collected on
Quercus candicans, in which the authors concluded that
P. calyculatus had an effect on the endothelium, inducing the synthesis and release of nitric oxide at concentrations of 800 μg/mL [
30].
There are several studies on the medicinal properties of other African parasitic plants, such as
Loranthus micranthus and
Loranthus bengwensis, which suggest that their ACE inhibition effect is host-dependent [
31,
32,
33,
34,
35].
2.2. HPTLC
2.2.1. Detection of C3G in Ethanolic Extracts of Pericarp of P. calyculatus
Regarding the detection and quantification of bioactive metabolites, C3G, whose chemical structure provides a dark coloration of purple-black tones to grapes, blackberries and blueberries, among other pigmented fruits, was detected via HPTLC and TLC [
36,
37,
38].
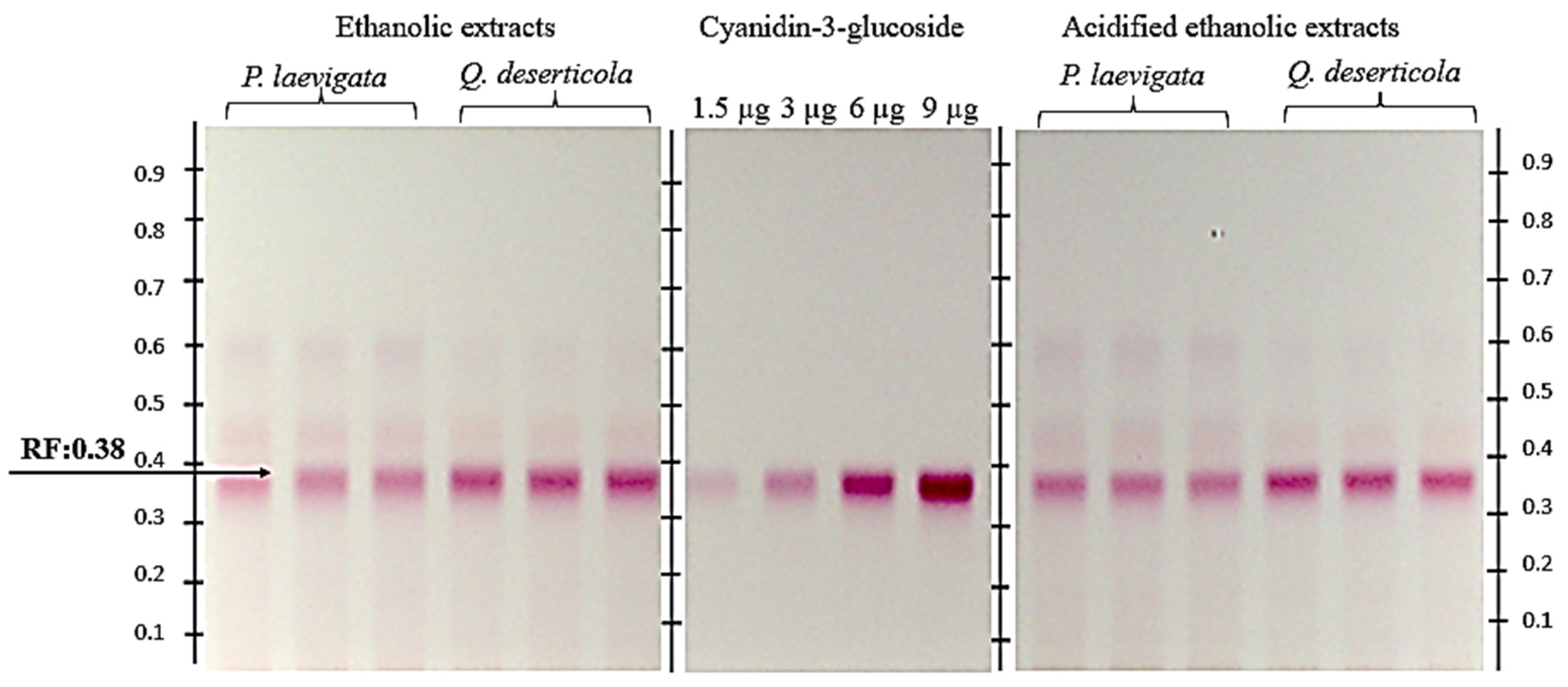
Figure 1 shows the chromatogram of
P. calyculatus fruit pericarp extracts: absolute ethanol, acidified ethanol and the C3G standard. A retention factor (Rf) value of 0.38 was obtained, and a coloration similar to that of the anthocyanin standard was used.
Regarding the quantification of C3G (
Table 5), a higher recovery of the metabolite was obtained with the acidified ethanol extracts of pericarp of
P. calyculatus fruits from both hosts (
P. laevigata and
Q. deserticola). The acidified extracts of fruits from specimens collected from the host
Q. deserticola were almost double the concentration of C3G (306.682 mg C3GE/g DW) of the rest of the extracts.
There are studies of plants leaves of other genera and species belonging to the Loranthaceae family, such as
Plicosepalus curviflorus,
Plicosepalus acacia,
Helicanthus elastica and
Scurrula atropurpurea, among others, in which quercetin, catechin, flavonol gallate, alkaloids, anthra-quinones, terpenoids, tannins and quinones have been identified via HPTLC and TLC [
39,
40,
41,
42,
43]. However, this is the first chromatographic study of the fruit of
P. calyculatus, since the chromatographic reports found on this species concern the leaves, generally using the HPLC technique [
13].
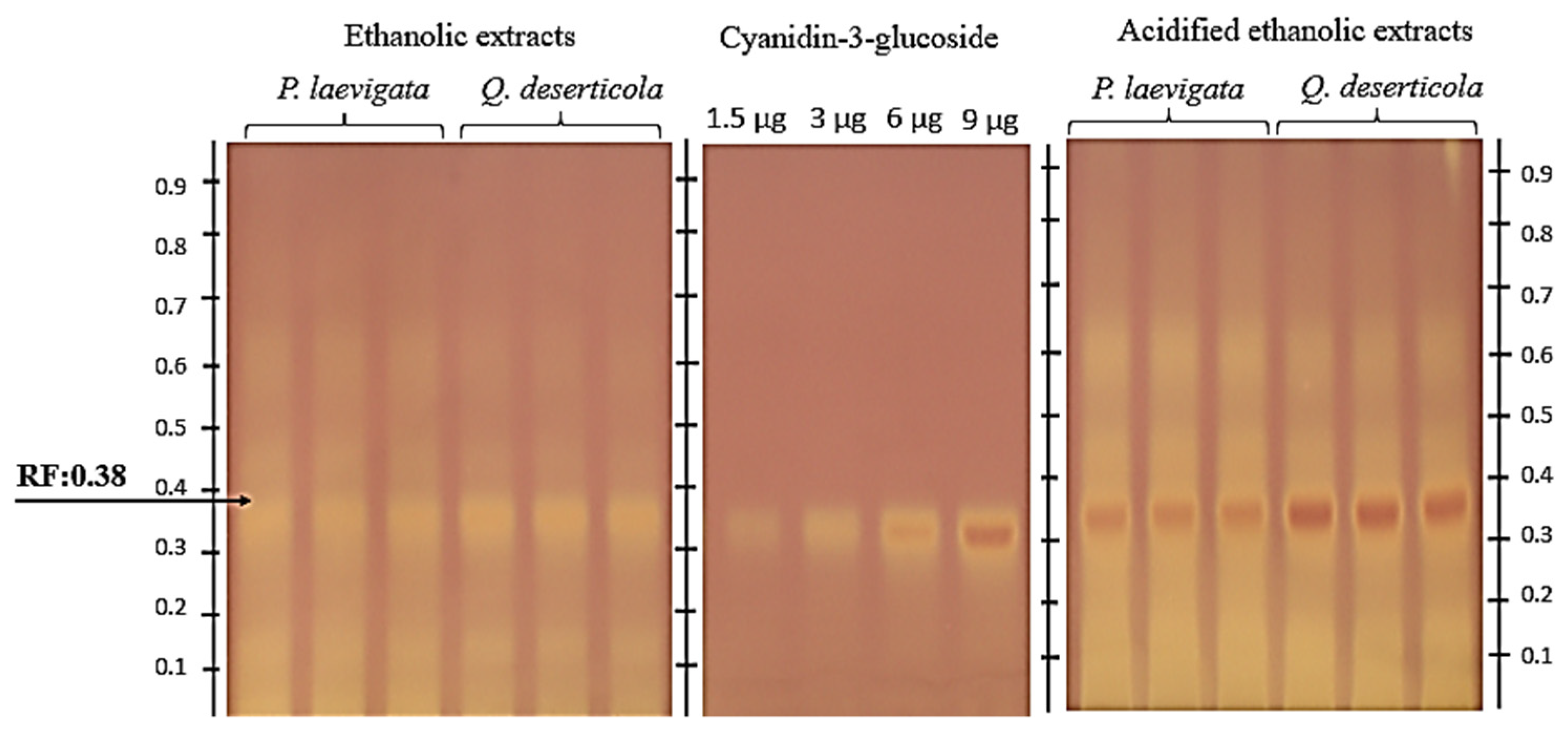
2.2.2. HPTLC–DPPH• Antioxidant Assay
As for the detection via HPTLC–DPPH of compounds with antioxidative potential (
Figure 2), bands coinciding with the retention factor of the C3G (Rf = 0.380) were found.
Similar results were obtained in a study on the phytotherapeutic potential of berries determined via their antioxidant capacity (blue-berried honeysuckles), in which C3G showed a strong signal upon derivatization with DPPH
• [
36]. It is worth mentioning that C3G has been identified in pigmented fruits [
44,
45].
2.3. Pearson’s Correlation of the Analyses Evaluated
Pearson’s correlation (
Table 6), together with the two-way statistical analyses, suggested that both the extractant solvent and the host have a significant effect on the recovery of metabolites from
P. calyculatus fruit correlating with biological activities, such as antioxidant, antimicrobial and antihypertensive activities. MIC and MBC showed a desirable high negative correlation with TAC, TPC and C3G. In the same way, ACE obtained a high negative correlation with TPC and TAC, in addition to showing a high positive correlation with MIC and MBC. Antioxidant activity identified using the ABTS
●+ method was highly correlated with TPC, while via the DPPH
● method it was highly correlated with TFC and C3G content.
3. Materials and Methods
HPLC-grade methanol, hydrochloric acid (HCl), chlorogenic acid, gallic acid, quercetin, Trolox, anthocyanin standards in the form of chloride salts, thiazolyl blue tetrazolium blue (MTT), benzenesulfonyl chloride (BSC), hippuryl-L-histidyl-L-leucine (HHL) and angiotensin converting enzymes (ACEs) were purchased from Sigma-Aldrich® (St. Louis, MO, USA). Silica gel plates 60 F254 for HPTLC (20 × 10 cm, Art. 1.05729.0001) were supplied by Merck® (Darmstadst, Germany). For mobile phases, toluene, ethyl acetate, acetic acid and formic acid supplied by Sigma-Aldrich® (St. Louis, MO, USA) were used. Broth and Mueller–Hinton agar were purchased from BD Bioxon (State of Mexico, Mexico). Certified strains of Escherichia coli ATCC 12792, Salmonella choleraesuis ATCC 10708 and Shigella flexneri ATCC 12022 were used.
3.1. Plant Samples
The mature fruits of P. calyculatus were collected on Q. deserticola and P. laevigata hosts in the localities of the La Arena municipality of Marcos Castellanos, Michoacán (19°59′25″ N, 102°53′57″ W) and Jiquilpan de Juárez, Michoacán (20°1′36″ N, 102°41′24″ W), in the autumn (October 2020). The plants were identified by M.S. Ignacio García Ruíz (collection number 9971 and 9974). Specimens were deposited in the CIIDIR—Michoacán Herbarium (CIMI).
3.2. Extract Preparation
The fruits were rinsed with distilled water, the pericarp was separated, lyophilized (Lyophilizer, LABCON, Kansas City, MO, USA) and stored at −20 °C until use. The pericarp of fruits from each host, i.e., oak and mesquite, was pulverized with a pestle and mortar. Extracts were prepared with absolute ethanol and ethanol acidified with 0.01% HCl. Samples of 1 g were extracted in 75 mL of extractive solvent via three washes. The extracts were sonicated at 55 ± 5 Hz (UL-TRAsonik, DENSTPLY, NEYTECH, Yucaipa, CA, USA) for 30 min at room temperature and left in agitation. The extracts were vacuum filtered through a 0.45 µm cellulose filter and stored at −20 °C.
3.3. Total Phenols Quantification
The quantification of total phenols (TPC) in the extracts was based on a calibration curve, as reported by Spinardi et al. [
46]. The calibration curve A
760 = 0.6032 [gallic acid] + 0.0404,
R2 = 0.9929, was obtained from 10 concentrations (0.25–2.5 mg/mL) of gallic acid. The absorbance was measured at 760 nm using a spectrophotometer (PowerWave HT, Biotek, Biotek Instruments, Winooski, VT, USA). The total content was expressed in milligram gallic acid equivalents per gram of dry weight (mg GAE/g DW). The assay was performed in triplicate.
3.4. Total Flavonoids Quantification
Total flavonoid content (TFC) was determined using the technique reported by Woisky and Salatino [
47] with some modifications, using a standard curve for quercetin, (A
425 = 1.1113 [quercetin] − 0.0009,
R2 = 0.9965) obtained from 10 concentrations (0.1–1 mg/mL). Absorbance was measured at 425 nm in the spectrophotometer (PowerWave HT, Biotek, Biotek Instruments, Winooski, VT, USA). The total content was expressed as milligram quercetin equivalents per gram of dry weight (mg QE/g DW). The assay was performed in triplicate.
3.5. Total Anthocyanin Quantification
The total anthocyanin content (TAC) was determined using the technique described by Abdel and Hucl [
48]. The absorbance was measured at 535 nm with the spectrophotometer (PowerWave HT, Biotek, Biotek Instruments, Winooski, VT, USA). Total anthocyanin content was expressed as a cyanidin-3-glucoside equivalent (mg C3GE/g DW) and calculated using the following formula:
3.6. High Performance Thin Layer Chromatography (HPTLC)
3.6.1. General Conditioning
The identification and quantification of secondary metabolites was performed using the techniques reported by Cretu et al. [
49]. Silica gel plates 60 F 254 of 10 × 20 cm (Merck
®, (Darmstadst, Germany) were used. Plates were activated by heating (TLC Plate Heater 3, CAMAG, Muttenz, Switzerland) at 100 °C for 3 min. After cooling to room temperature (22 °C ± 2), samples were applied (1 g/75 mL) with a sample applicator (ATS 4, CAMAG, Muttenz, Switzerland), and the plate was developed with 10 mL of solvent system in the automated development chamber (ADC 2, CAMAG, Muttenz, Switzerland) at 47% RH (relative humidity) (in equilibrium with a saturated solution of potassium thiocyanate KSCN) for 10 min. The plates were derivatized with a 1% natural products (NP) methanolic solution (2-aminoethyl diphenyl borate, Sigma-Aldrich
®, St. Louis, MO, USA) reagent and derivatization was performed in the immersion device (Chromatogram Immersion Device, CAMAG, Muttenz, Switzerland) at an immersion speed of 5 cm/s with an immersion time of 1 s. After derivatization, the plate was dried for 3 min at 100 °C (TLC Plate Heater 3, CAMAG, Muttenz, Switzerland). Images of each plate were documented using a TLC Visualizer (CAMAG, Muttenz, Switzerland) under visible light, at UV 254 nm and UV 366 nm.
Results were processed via the VisionCATS version 1.4.7.2018 software (CAMAG, Muttenz, Switzerland).
3.6.2. Identification and Quantification of Cyanidin-3-Glycoside
For the identification and determination of anthocyanins the techniques reported by Cretu et al. [
38] were applied. The analysis was performed in triplicate; silica gel plates 60 F 254 of 10 × 20 cm (Merck
®, Darmstadt, Germany) were used, the plates were conditioned (preheating at 100° C for 3 min) and then a sample applicator (ATS 4, CAMAG, Muttenz, Switzerland) was used. The C3G standard was loaded at different volumes 1. 5, 3, 6, 9 and 12 µL in order to obtain a calibration curve (y = −1.946 × 10
−15x
2 + 5.207
× 10
−8x − 3.155 × 10
−2,
R2 = 0.9999). The standard was prepared at a concentration of 1 mg/mL of methanol. The absolute and acidified ethanolic fruit pericarp extracts were prepared at a concentration of 1 g/75 mL, a volume of 2.5 µL was applied to samples extracted with absolute ethanol and a volume of 4 µL was applied to samples extracted with ethanol acidified. The bands were adjusted and applied, with a position of Y: 8 mm and X: 16.8 mm, with a length of 7 mm and a distance between lanes of 10.4 mm. A total of 2.0 µL of the extracts and standards was applied at a constant application rate (100 nL/s). Plate development was carried out in the automated development chamber (ADC 2, CAMAG, Muttenz, Switzerland) at 47% RH (in equilibrium with a saturated solution of potassium thiocyanate KSCN) for 10 min, the plate was developed with 10 mL of a solvent system comprising ethyl acetate:acetic acid:formic acid:water (10:1.1:1:1:2.3). The developed plate was visualized, and a digitized record was obtained under white light and visible light at 254 nm and 366 nm on the visualizer (TLC Visualizer, CAMAG, Muttenz, Switzerland). Subsequently, the plate was derivatized with the natural products (NP) reagent in the immersion device (Chromatogram Immersion Device, CAMAG, Muttenz, Switzerland) and dried in a plate heater (TLC Plate Heater 3, CAMAG, Muttenz, Switzerland) at 100 °C for 3 min.
3.6.3. Antioxidant Activity by HPTLC-DPPH●
Antioxidant activity was determined via the HPTLC-DPPH
● (2,2-difenil-1-picrilhidrazilo) method according to Orsini et al. [
50] with some modifications. Developed plates for the identification and quantification of anthocyanins as previously described were derivatized with DPPH
● reagent (0.2% in methanol) in the immersion device (Chromatogram Immersion Device, CAMAG, Muttenz, Switzerland) using a vertical velocity of 5 cm/s, for 3 s. The plates were dried at room temperature (22 ± 2 °C) in the dark for 30 min. The images obtained were documented using the TLC Visualizer (CAMAG, Muttenz, Switzerland) under visible light. The data obtained were processed with VisionCATS version 1.4.7.2018 software (CAMAG, Muttenz, Switzerland).
3.7. Biological Activities
3.7.1. Antioxidant Activity by ABTS•+ Assay
The radical-scavenging capacity of the methanolic extracts was determined using the ABTS
•+ method, as reported by Hosu et al. [
51]. The ABTS solution was prepared by dissolving 360 mg of ABTS
•+ (2,2′azinobis-(3-ethylbenzothiazdin-6-sulfonic acid) in 100 mL of deionized water, and 100 mL of ABTS solution was added to 100 mL of 2.45 mM potassium persulfate solution for the activation of ABTS
•+. The mixture reaction was conserved in the dark for 24 h. The absorbance was measured at 734 nm using (PowerWave HT, Biotek, Biotek Instruments, Winooski, VT, USA) and was adjusted to about 0.760 ± 0.001 through dilution with distilled water. A total of 280 µL of ABTS solution was added to 20 µL of methanolic extracts in a 96-well microplate, and the mixture was kept in the dark for 15 min. The absorbance of the sample and the blank were measured at 734 nm using the spectrophotometer (PowerWave HT, Biotek, Biotek Instruments, Winooski, VT, USA). The antioxidant capacity of the extracts was expressed as μmoles Trolox equivalent/g DW (μmol TE/g DW) using a calibration curve (μmol TE = −5.4371 [A
734nm] + 1.4357,
R2 = 0.9852) at five different concentrations (0–0.60 μmol Trolox). The assay was performed in triplicate.
3.7.2. Antioxidant Activity by DPPH● Assay
The antioxidant activity of the extracts was determined according to the DPPH
● method described by Marín et al. [
19] with some modifications. In 96-well microplates, 20 μL of the ethanolic and ethanolic acidified extracts diluted (1:4) and 200 μL of DPPH
● solution (150 mM, ethanolic solution) were added. The mixtures were incubated for 30 min in the dark. The absorbance was measured at 515 nm using the spectrophotometer (PowerWave HT, Biotek, Biotek Instruments, Winooski, VT, USA) and the antioxidative capacity of the extracts was expressed as the milligram equivalents of the antioxidant capacity of Trolox per gram dry weight (μM ET/g PS) using a calibration curve (μM ET = −2.8918 [A
515nm] + 1.4357,
R2 = 0.9849) at five concentrations (0–0.60 μmol Trolox).
3.7.3. Determination of Antihypertensive Activity via ACE Inhibition
The assay was performed based on the hydrolysis of hippuryl-L-histidyl-L-leucine (HHL) vua ACE to hippuric acid (HA) and histidyl-leucine (HL) as products, as reported by Lia et al. [
52], with some modifications. For each assay (sample, control and blank) according to Li et al. (2005), an 8 µL ACE inhibitor sample solution was prepared with 50 µL of 5 mM HHL in 100 mM sodium borate buffer (pH 8.3) containing 300 mM NaCl, which was previously incubated at 37 °C for 5 min. The reaction was initiated by the addition of 2 µL of ACE solution (100 mU/mL) and the mixture was incubated at 37 °C for 30 min. The reaction was halted by adding 40 µL of 1 M HCl, in conjunction with 30 µL of sodium borate. Subsequently, 96 µL of quinolone was added, followed by 32 µL of benzenesulfonyl chloride (BSC) and then incubation at 30 °C for 30 min in the dark. A buffer was then added to the mixture of 570 µL of absolute ethanol in each assay (sample, control and blank). Finally, followed by incubation at 30 °C for 30 min, absorbance was measured on a microplate reader (spectrophotometer, PowerWave HT, Biotek, Biotek Instruments, Winooski, VT, USA) at 492 nm. All measurements were performed in triplicate. A calibration curve versus HA concentration was obtained. Captopril (25 mg) was used as a control. The percentage calculation of ACE inhibitory activity was performed by applying the following equation:
where A = sample; B = control; and C = blank.
3.7.4. Determination of Antimicrobial Activity
The antimicrobial activity was determined according to that reported by Karuppasami et al. [
53] with some modifications. The MIC and MBC were determined using the standard broth microdilution method. Extracts with concentrations of 50, 37.5, 25, 18.75, 12.5, 6.54 and 2.35 mg/mL were placed in sterile polystyrene 96-well microplates containing 100 μL of Mueller–Hinton broth. A 24 h inoculum of
E. coli ATCC 12792,
S. choleraesuis ATCC 10708
S. flexneri ATCC 12022) was adjusted to 1 × 10
7 CFU/mL and 50 µL of each bacterial suspension were added. As controls, 200 μL of Mueller–Hinton broth and 20 μL of the inoculum adjusted to a density of 1 × 10
7 CFU/mL were used. The plates were incubated at 37 °C for 19 h and 20 μL of MTT tetrazolium salts were added before continuing the incubation for 45 min; finally, the MIC was recorded as the lowest concentration of the extracts without visible bacterial growth. For the determination of the MBC, as the lowest concentration that killed 100% of the bacteria, an aliquot of 50 μL from the wells without coloring was spread on plates with Mueller–Hinton agar medium and incubated at 37 °C for 24 h. The following controls were considered: positive control, broth + bacteria; negative control, broth only; and antibiotic control, broth + bacteria + antibiotic (30 µg ciprofloxacin). Three replicates of each of the dilutions and controls were performed. The tests were performed in triplicate on different dates.
3.8. Statistical Analysis
The results were reported with the mean ± standard deviation. A two-way analysis of variance (ANOVA) was performed, the separation of means was performed using Tukey’s test (p < 0.05) and Pearson’s correlation was also carried out. R Studio software version 4.0.3. was used.
4. Conclusions
The phytochemical and bioactive properties of the ripe fruits of Psittacanthus calyculatus were studied here for the first time. The cyanidin-3-glucoside was detected in ethanolic extracts (with absolute and acidified ethanol), which showed antihypertensive potential, antioxidant capacity and antibacterial activity (against S. choleraesuis, E. coli and S. flexneri). Moreover, it was observed that, by means of acidified ethanolic extraction, a greater recovery of metabolites was obtained from the fruit.
The results showed that the host was determinant for the accumulation of secondary metabolites. An effect of the hosts on the phenolic content of P. calyculatus fruits was found, the host P. laevigata favored the accumulation of phenolic compounds, while Q. deserticola promoted a higher content of flavonoids and anthocyanins. However, further research on the interaction of P. calyculatus and its hosts is needed to clarify this phenomenon.
Based on the results obtained, a range of potential benefits for ethnopharmacology, industry and agriculture, as well as other sectors, can be envisaged.
Author Contributions
Conceptualization, J.M.-T., M.V.A.-P., I.G.-R. and H.G.M.-V.; data curation, J.A.C.-D.; formal analysis, Z.O.-C., J.A.C.-D. and H.G.M.-V.; investigation, Z.O.-C.; methodology, Z.O.-C., J.G.C.-V., I.G.-R., J.O.B.-G. and H.G.M.-V.; project administration, H.G.M.-V.; resources, H.G.M.-V.; supervision, J.M.-T.; validation, Z.O.-C. and J.G.C.-V.; visualization, J.O.B.-G.; writing—original draft, Z.O.-C.; writing—review and editing, J.M.-T., M.V.A.-P. and H.G.M.-V. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Acknowledgments
The first author thanks the Consejo Nacional de Ciencia y Tecnología (CONACYT) in México for the financial support through the national scholarship 1007704. H.G.M.-V. and I.G.-R. appreciate the financial support from Instituto Politécnico Nacional through the projects SIP- 20210625 and SIP- 20210731, and the academic fellowships COFAA–IPN and EDI–IPN.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ornelas, J. Los Muérdagos Psittacanthus en México: Ecología, Evolución, Manejo y Conservación. Biodiversitas 2019, 146, 1689–1699. [Google Scholar]
- Pérez, J.I.J. Impacto del Muérdago (Psittacanthus calyculatus) en la Economía de las Familias Campesinas en Una Región del Subtrópico Mexicano. Perspect. Lat. 2016, 13, 141–156. [Google Scholar]
- Vázquez Collazo, I.; Geils, B.W. Psittacanthus in México; Chapter 2; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Vancuver, BC, Canada, 2002. [CrossRef]
- Vazquez Collazo, I.; Villa Rodríguez, A.; Madrigal Huendo, S. Los Muerdagos (Loranthaceae) En Michoacán; Sagarpa: Michoacán, Mexico, 2006; ISBN 9688006440. [Google Scholar]
- Cuevas, P.; Pérez, G.; Maldonado, Y.; González, A. Effects of Herbivory and Mistletoe Infection by Psittacanthus calyculatus on Nutritional Quality and Chemical Defense of Quercus deserticola along Mexican Forest Fragments. Plant Ecol. 2017, 218, 687–697. [Google Scholar] [CrossRef]
- Cantoral Herrera, M.; Patiñ Pineda, M.; Recillas Silva, I. Diagnóstico, Evaluación y Control de Plantas Parásitas de La Ciudad de México. In Proceedings of the Foro Nacional las Plantas Parásitas en México; Alvarado Rosales, D., Saavedra Romero, L., Eds.; Red Temática en Salud Forestal: Texcoco, México, 2017; p. 52. [Google Scholar]
- Gómez, M.; Sánchez, L.J.; Salazar, L.A. Anatomy of Mexican Species of the Genera Phoradendron and Psittacanthus, Endemic to the New World. Rev. Mex. Biodivers. 2011, 82, 1203–1218. [Google Scholar] [CrossRef]
- Esquivel, E.R.; Noriega-Cisneros, R.; Bello-González, M.A.; Saavedra-Molina, A.; Salgado-Garciglia, R. Plantas Utilizadas En La Medicina Tradicional Mexicana Con Propiedades Antidiabéticas y Antihipertensivas. Biológicas 2012, 14, 45–52. [Google Scholar]
- Hernández, T.; García-Bores, A.M.; Serrano, R.; Ávila, G.; Dávila, P.; Cervantes, H.; Peñalosa, I.; Flores-Ortiz, C.M.; Lira, R. Fitoquímica Y Actividades Biológicas De Plantas De Importancia En La Medicina Tradicional Del Valle De Tehuacán-Cuicatlán. Tip 2015, 18, 116–121. [Google Scholar] [CrossRef]
- Vázquez Rodríguez, E. Actividades Biológicas de Extractos de Plantas y de Sus Combinaciones. Ph.D. Thesis, Universidad Autónoma de Madrid, Instituto de Investigación en Ciencias de la Alimentación (CIAL), Madrid, Spain, 2016; p. 319. [Google Scholar]
- Zarza, L.O.; Bah, M.; Castañeda, R.; Mendoza, S.O. Relación Entre el Origen y la Capacidad Antihipertensiva de Especímenes de Psittacanthus calyculatus Recolectada Sobre Tres Diferentes Hospederos; Universidad Autónoma de Querétaro: Santiago de Querétaro, Mexico, 2009. [Google Scholar]
- Ibarra, C.; Rojas, A.; Mendoza, S.; Bah, M.; Gutiérrez, D.M.; Hernández-Sandoval, L.; Martínez, M. Vasoactive and Antioxidant Activities of Plants Used in Mexican Traditional Medicine for the Treatment of Cardiovascular Diseases. Pharm. Biol. 2010, 48, 732–739. [Google Scholar] [CrossRef]
- Moustapha, B.; Marina, G.A.D.; Raúl, F.O.; Raquel, C.M.; Mahinda, M. Chemical Constituents of the Mexican Mistletoe (Psittacanthus calyculatus). Molecules 2011, 16, 9397–9403. [Google Scholar] [CrossRef]
- Hernández Rodríguez, P.; Baquero, L.C.P.; Álvarez, M.F.R. Propiedades Químicas y Biológicas de Arbutus Unedo: Una Planta Con Potencial Medicinal. Rev. Cuba. Farm. 2015, 49, 144–155. [Google Scholar]
- Serrano, M.J.; Guerrero-Legarreta, I.; De la paz pérez-Olvera, C.; Soriano-Santos, J. Actividad Antioxidante y Efecto Citotóxico de Cladocolea loniceroides. Rev. Mex. Ing. Qum. 2011, 10, 161–170. [Google Scholar]
- dos Santos Gramma, L.S.; Marques, F.M.; Vittorazzi, C.; de Andrade, T.A.M.; Frade, M.A.C.; de Andrade, T.U.; Endringer, D.C.; Scherer, R.; Fronza, M. Struthanthus vulgaris Ointment Prevents an over Expression of Inflammatory Response and Accelerates the Cutaneous Wound Healing. J. Ethnopharmacol. 2016, 190, 319–327. [Google Scholar] [CrossRef]
- Leitão, F.; de Lima Moreira, D.; de Almeida, M.Z.; Guimarães Leitão, S. Secondary Metabolites from the Mistletoes Struthanthus marginatus and Struthanthus concinnus (Loranthaceae). Biochem. Syst. Ecol. 2013, 48, 215–218. [Google Scholar] [CrossRef]
- Bezerra, A.N.S.; Massing, L.T.; de Oliveira, R.B.; Mourão, R.H.V. Standardization and Anti-Inflammatory Activity of Aqueous Extract of Psittacanthus plagiophyllus Eichl. (Loranthaceae). J. Ethnopharmacol. 2017, 202, 234–240. [Google Scholar] [CrossRef]
- Marín, D.; Brango, J.; Galeano, P. Caracterización Química, Evaluación de la Actividad Antioxiodante y Antibacterial Del Extracto Crudo de Psittacanthus cucullaris. Momentos Cienc. 2013, 10, 2–10. [Google Scholar]
- Bailladores, J.P.; Delgado, G.E.; Wagner, M.L.; Rojas, C. In Vitro Tissue Culture, Preliminar Phytochemical Analysis, and Antibacterial Activity of Psittacanthus linearis (Killip) J.K. Macbride (Loranthaceae). Rev. Colomb. Biotecnol. 2019, 21, 22–35. [Google Scholar] [CrossRef]
- Ozpinar, H.; Ozpinar, N.; Eruygur, N. Effect of Viscum album L. Ssp. Austriacum (WİESP.) Vollman on Metronidazole Resistant and Sensitive Strains of Trichomonas vaginalis. South African J. Bot. 2019, 125, 81–85. [Google Scholar] [CrossRef]
- Subash, S.; Essa, M.M.; Al-Adawi, S.; Memon, M.A.; Manivasagam, T.; Akbar, M. Neuroprotective Effects of Berry Fruits on Neurodegenerative Diseases. Neural Regen. Res. 2014, 9, 1557–1566. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, M.; Shadnoush, M.; Khorshidian, N.; Mortazavian, A.M. Insights to Potential Antihypertensive Activity of Berry Fruits. Phyther. Res. 2021, 35, 846–863. [Google Scholar] [CrossRef] [PubMed]
- Karasawa, M.M.G.; Mohan, C. Fruits as Prospective Reserves of Bioactive Compounds: A Review. Nat. Products Bioprospect. 2018, 8, 335–346. [Google Scholar] [CrossRef]
- Raza, H.; Shehzad, M.A.; Baloach, A.; Ikram, R.M. Antioxidant and Antimicrobial Activity of Fruit Juices. Int. J. Agric. Sustain. Dev. 2019, 10, 9600. [Google Scholar] [CrossRef]
- Toledo De Oliveira, T.; Nagem, T.J.; Rocha Da Costa, M.; Marciano Da Costa, L.; Magalhães, N.M.; Stringheta, P.C.; Queiroga De Lima, E.; Kling De Moraes, G.H.; Da Silva Vieira, H. Propiedades Biológicas de Los Tintes Naturales. Ars Pharm. 2004, 45, 5–20. [Google Scholar]
- Cervantes, M.G.; Salazar, L.A.; Valle, J.R. Preparación de Psittacanthus calyculatus Con Propiedades Antihipertensivas, Métodos Para Su Producción y Uso de La Misma. Patent No. A61K36/00, March 2011. [Google Scholar]
- Luczkiewicz, M.; Cisowski, W.; Kaiser, P.; Ochocka, R.; Piotrowski, A. Comparative Analysis of Phenolic Acids in Mistletoe Plants from Various Hosts. Acta Pol. Pharm. Drug Res. 2001, 58, 373–379. [Google Scholar]
- Ogunmefun, O.T.; Fasola, T.R.; Saba, A.B.; Oridupa, O.A. The Ethnobotanical, Phytochemical and Mineral Analyses of Phragmanthera incana (Klotzsch), a Species of Mistletoe Growing on Three Plant Hosts in South-Western Nigeria. Int. J. Biomed. Sci. 2013, 9, 37–44. [Google Scholar]
- Rodríguez, M.E.; Pérez-Ordaz, L.; Serrato-Barajas, B.E.; Juárez-Oropeza, M.A.; Mascher, D.; Paredes-Carbajal, M.C. Endothelium-Dependent Effects of the Ethanolic Extract of the Mistletoe Psittacanthus calyculatus on the Vasomotor Responses of Rat Aortic Rings. J. Ethnopharmacol. 2003, 86, 213–218. [Google Scholar] [CrossRef]
- Obatomi, D.K.; Aina, V.O.; Temple, V.J. Efects of African Mistletoe Extract on Blood Pressure in Spontaneously Hypertensive Rats. Int. J. Pharmacogn. 1996, 34, 124–127. [Google Scholar] [CrossRef]
- Iwalokun, B.A.; Hodonu, S.A.; Nwoke, S.; Ojo, O.; Agomo, P.U. Evaluation of the Possible Mechanisms of Antihypertensive Activity of Loranthus micranthus: An African Mistletoe. Biochem. Res. Int. 2011. [Google Scholar] [CrossRef] [PubMed]
- Zorofchian, S.; Hajrezaei, M.; Abdul Kadir, H.; Zandi, K. Loranthus micranthus Linn.: Biological Activities and Phytochemistry. Evid.-Based Complement. Altern. Med. 2013. [Google Scholar] [CrossRef] [PubMed]
- Oboh, G.; Omojokun, O.S.; Ademiluyi, A.O. Drying Methods Alter Angiotensin-I Converting Enzyme Inhibitory Activity, Antioxidant Properties, and Phenolic Constituents of African Mistletoe (Loranthus Bengwensis L) Leaves. J. Evid.-Based Complement. Altern. Med. 2016, 21, 260–270. [Google Scholar] [CrossRef]
- Omojokun, O.S.; Oboh, G.; Ademiluyi, A.O. Effects of Drying on Cholinesterases and Angiotensin-I Converting Enzyme Inhibitory Potential and Phenolic Constituents of African Mistletoe (Loranthus Bengwensis L) Leaves from Kolanut Host Tree. J. Food Biochem. 2018, 42, e12510. [Google Scholar] [CrossRef]
- Kusznierewicz, B.; Piekarska, A.; Mrugalska, B.; Konieczka, P.; Namieśnik, J.; Bartoszek, A. Phenolic Composition and Antioxidant Properties of Polish Blue-Berried Honeysuckle Genotypes by HPLC-DAD-MS, HPLC Postcolumn Derivatization with ABTS or FC, and TLC with DPPH Visualization. J. Agric. Food Chem. 2012, 60, 1755–1763. [Google Scholar] [CrossRef]
- Forgacs, E.; Cserhati, T. Thin-Layer Chromatography of Natural Pigments: New Advances. J. Liq. Chromatogr. Relat. Technol. 2002, 25, 1521–1541. [Google Scholar] [CrossRef]
- Cretu, G.C.; Morlock, G.E. Analysis of Anthocyanins in Powdered Berry Extracts by Planar Chromatography Linked with Bioassay and Mass Spectrometry. Food Chem. 2014, 146, 104–112. [Google Scholar] [CrossRef]
- Bamane, F. Antioxidant Activities and Flavonoid Contents of Selected Plants Belonging to Family Loranthaceae. African J. Biotechnol. 2012, 11, 14380–14385. [Google Scholar] [CrossRef]
- Kumar, K.N.S.; Saraswathy, A.; Amerjothy, S. HPTLC Fingerprinting of Extracts of Mango Mistletoe Helicanthus elastica ( Desr.) Danser with Multiple Markers. J. Sci. Innov. Res. 2013, 2, 864–871. [Google Scholar]
- Mustarichie, R.; Runadi, D.; Ramdhani, D. The Antioxidant Activity and Phytochemical Screening of Ethanol Extract, Fractions of Water, Ethyl Acetate, and n-Hexane from Mistletoe Tea (Scurrula atropurpurea BL. Dans). Asian J. Pharm. Clin. Res. 2017, 10, 343–347. [Google Scholar] [CrossRef]
- Cyprian, T.A.; Sewuese, S.; Akacha, L.U. Preliminary Phytochemical Screening and Thin Layer Chromatography Analysis of Stem Bark Extracts of African Mistletoe Parasitic on Vitellaria paradoxa, Piliostigma thonningii and Combretum fragrans. Asian J. Appl. Chem. Res. 2019, 3, 1–6. [Google Scholar] [CrossRef]
- Orfali, R.; Perveen, S.; Siddiqui, N.A.; Alam, P.; Alhowiriny, T.A.; Al-Taweel, A.M.; Al-Yahya, S.; Ameen, F.; Majrashi, N.; Alluhayb, K.; et al. Pharmacological Evaluation of Secondary Metabolites and Their Simultaneous Determination in the Arabian Medicinal Plant Plicosepalus curviflorus Using HPTLC Validated Method. J. Anal. Methods Chem. 2019. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Li, Y.; Hou, D.X.; Wu, S. The Effects and Mechanisms of Cyanidin-3-Glucoside and Its Phenolic Metabolites in Maintaining Intestinal Integrity. Antioxidants 2019, 8, 479. [Google Scholar] [CrossRef]
- Košir, I.J.; Lapornik, B.; Andrenšek, S.; Wondra, A.G.; Vrhovšek, U.; Kidrič, J. Identification of Anthocyanins in Wines by Liquid Chromatography, Liquid Chromatography-Mass Spectrometry and Nuclear Magnetic Resonance. Anal. Chim. Acta 2004, 513, 277–282. [Google Scholar] [CrossRef]
- Spinardi, A.; Cola, G.; Gardana, C.S.; Mignani, I. Variation of Anthocyanin Content and Profile Throughout Fruit Development and Ripening of Highbush Blueberry Cultivars Grown at Two Different Altitudes. Front. Plant Sci. 2019, 10, 1045. [Google Scholar] [CrossRef]
- Woisky, R.G.; Salatino, A. Analysis of Propolis: Some Parameters and Procedures for Chemical Quality Control. J. Apic. Res. 1998, 37, 99–105. [Google Scholar] [CrossRef]
- Abdel-Aal, E.-S.M.; Hucl, P. A Rapid Method for Quantifying Total Anthocyanins in Blue Aleurone and Purple Pericarp Wheats. Cereal Chem. J. 1999, 76, 350–354. [Google Scholar] [CrossRef]
- Creţu, G.; Morlok, G.; Nechifor, G. Development of a Quantitative High Performance Thin Layer Cromatographic Method for Analysis of Delphinidin 3-Glucoside in Berry Extracts. UPB Sci. Bull. Ser. B Chem. Mater. Sci. 2013, 75, 69–78. [Google Scholar]
- Orsini, F.; Vovk, I.; Glavnik, V.; Jug, U.; Corradini, D. HPTLC, HPTLC-MS/MS and HPTLC-DPPH Methods for Analyses of Flavonoids and Their Antioxidant Activity in Cyclanthera pedata Leaves, Fruits and Dietary Supplement. J. Liq. Chromatogr. Relat. Technol. 2019, 42, 290–301. [Google Scholar] [CrossRef]
- Hosu, A.; Cimpoiu, C.; David, L.; Moldovan, B. Study of the Antioxidant Property Variation of Cornelian Cherry Fruits during Storage Using HPTLC and Spectrophotometric Assays. J. Anal. Methods Chem. 2016. [Google Scholar] [CrossRef] [PubMed]
- Li, G.H.; Liu, H.; Shi, Y.H.; Le, G.W. Direct Spectrophotometric Measurement of Angiotensin I-Converting Enzyme Inhibitory Activity for Screening Bioactive Peptides. J. Pharm. Biomed. Anal. 2005, 37, 219–224. [Google Scholar] [CrossRef]
- Karuppusamy, S.; Rajasekaran, K.M. High Throughput Antibacterial Screening of Plant Extracts by Resazurin Redox with Special Reference to Medicinal Plants of Western Ghats. Glob. J. Pharmacol. 2009, 3, 63–68. [Google Scholar]
Figure 1.
Chromatogram of pericarp samples of mature P. calyculatus fruit extracted with absolute ethanol from two different hosts, namely P. laevigata (Mesquite) and Q. deserticola (Oak). Standard C3G was applied at different volumes with four calibration points, i.e., 1.5, 3, 6 and 9 µL, equivalent to 1.5, 3, 6 and 9 µg; y = −1.946 × 10−15x2 + 5.207 × 10−8x − 3.155 × 10−2, R2 = 0.9999. In a sample application volume of 2 µL (26.66 DW /mL), in triplicate. Pericarp samples of P. calyculatus extracted with acidified ethanol with respect to two distinct hosts, P. laevigata (Mesquite) and Q. deserticola (Oak) (viewed under white light, derivatized with NP (Sigma-Aldrich®, St. Louis, MO, USA) under a solvent system of ethyl acetate:acetic acid:formic acid:water (10:1.1:1.1:2.3). Retention factor (Rf) = 0.38.
Figure 1.
Chromatogram of pericarp samples of mature P. calyculatus fruit extracted with absolute ethanol from two different hosts, namely P. laevigata (Mesquite) and Q. deserticola (Oak). Standard C3G was applied at different volumes with four calibration points, i.e., 1.5, 3, 6 and 9 µL, equivalent to 1.5, 3, 6 and 9 µg; y = −1.946 × 10−15x2 + 5.207 × 10−8x − 3.155 × 10−2, R2 = 0.9999. In a sample application volume of 2 µL (26.66 DW /mL), in triplicate. Pericarp samples of P. calyculatus extracted with acidified ethanol with respect to two distinct hosts, P. laevigata (Mesquite) and Q. deserticola (Oak) (viewed under white light, derivatized with NP (Sigma-Aldrich®, St. Louis, MO, USA) under a solvent system of ethyl acetate:acetic acid:formic acid:water (10:1.1:1.1:2.3). Retention factor (Rf) = 0.38.
Figure 2.
Chromatogram of pericarp samples of P. calyculatus extracted with absolute ethanol with respect to two different hosts P. laevigata (Mesquite) and Q. deserticola (Oak). Standard C3G was applied at different volumes with four calibration points, namely 1.5, 3, 6 and 9 µL, equivalent to 1.5, 3, 6 and 9 µg; y = −1.946 × 10−15x2 + 5.207 × 10−8x − 3.155 × 10−2, R2 = 0.9999. In a sample application, a volume of 2 µL (26.66 DW /mL), in triplicate, acidified the ethanol extracts of two distinct hosts, P. laevigata (Mesquite) and Q. deserticola (Oak), viewed under white light and a DPPH•-derived anthocyanin assay plate.
Figure 2.
Chromatogram of pericarp samples of P. calyculatus extracted with absolute ethanol with respect to two different hosts P. laevigata (Mesquite) and Q. deserticola (Oak). Standard C3G was applied at different volumes with four calibration points, namely 1.5, 3, 6 and 9 µL, equivalent to 1.5, 3, 6 and 9 µg; y = −1.946 × 10−15x2 + 5.207 × 10−8x − 3.155 × 10−2, R2 = 0.9999. In a sample application, a volume of 2 µL (26.66 DW /mL), in triplicate, acidified the ethanol extracts of two distinct hosts, P. laevigata (Mesquite) and Q. deserticola (Oak), viewed under white light and a DPPH•-derived anthocyanin assay plate.
Table 1.
Total phenolic, flavonoid and anthocyanin content of ethanolic extracts of P. calyculatus fruit pericarp from two different hosts.
Table 1.
Total phenolic, flavonoid and anthocyanin content of ethanolic extracts of P. calyculatus fruit pericarp from two different hosts.
| Host (H) | Extractive Solvent (ES) | Total Phenols (mg GAE/g DW) | Total Flavonoids (mg QE/g DW) | Total Anthocyanins (mg C3GE/g DW) |
|---|
| P. laevigata | Acidified ethanol
Ethanol | 71.396 ± 0.676 a
39.588 ± 0.394 d | 10.986 ± 0.486 b
12.438 ± 0.758 b | 2.209 ± 0.032 b
0.826 ± 0.003 c |
| Q. deserticola | Acidified ethanol
Ethanol | 64.457 ± 0.522 b
34.179 ± 0.356 c | 14.232 ± 0.772 a
11.374 ± 0.505 b | 2.431 ± 0.020 a
0.765 ± 0.034 c |
| | (ES) | * | * | * |
| (H) | * | * | * |
| Interaction (ES:H) | * | - | * |
Table 2.
Antioxidant capacity of ethanolic extracts of pericarp of P. calyculatus fruits from two different hosts.
Table 2.
Antioxidant capacity of ethanolic extracts of pericarp of P. calyculatus fruits from two different hosts.
| Host (H) | Extractive Solvent (ES) | DPPH• Method (mg TE/g DW) | ABTS•+ Method (mg TE/g DW) |
|---|
| P. laevigata | Acidified ethanol
Ethanol | 47.505 ± 0.247 ab
46.595 ± 0.137 bc | 214.810 ± 0.802 a
151.602 ± 0.401 c |
| Q. deserticola | Acidified ethanol
Ethanol | 48.197 ± 0.518 a
47.284 ± 0.299 c | 205.036 ± 0.201 b
120.922 ± 0.201 d |
| | (SE) | * | * |
| (H) | * | * |
| Interaction (SE:H) | * | - |
Table 3.
Antimicrobial activity of ethanolic extracts of P. calyculatus pericarp from two different hosts against S. choleraesuis, E. coli and S. flexneri.
Table 3.
Antimicrobial activity of ethanolic extracts of P. calyculatus pericarp from two different hosts against S. choleraesuis, E. coli and S. flexneri.
| Microorganisms | Host | Extract | MIC
(mg/mL) | MBC
(mg/mL) |
|---|
S. choleraesuis
ATCC 10708 | P. laevigata | Acidified ethanol
Ethanol | 6.25
12.5 | 12.5
16.6 |
| Q. deserticola | Acidified ethanol
Ethanol | 6.25
12.5 | 12.5
16.6 |
E. coli
ATCC 12792 | P. laevigata | Acidified ethanol
Ethanol | 6.25
12.5 | 12.5
16.6 |
| Q. deserticola | Acidified ethanol
Ethanol | 6.25
12.5 | 12.5
16.6 |
S. flexneri
ATCC 12022 | P. laevigata | Acidified ethanol
Ethanol | 6.25
12.5 | 12.5
16.6 |
| Q. deserticola | Acidified ethanol
Ethanol | 6.25
12.5 | 12.5
16.6 |
Table 4.
Antihypertensive activity of ethanolic extracts of the P. calyculatus fruit pericarp from two different hosts.
Table 4.
Antihypertensive activity of ethanolic extracts of the P. calyculatus fruit pericarp from two different hosts.
| Host (H) | Extractive Solvent (ES) | % of ACE Inhibition |
|---|
| P. laevigata | Acidified ethanol | 19.400 ± 3.0547 c |
| Ethanol | 92.592 ± 2.645 a |
| Q. deserticola | Acidified ethanol | 64.373 ± 1.527 b |
| Ethanol | 89.947 ± 2.645 a |
| | (ES) | * |
| (H) | * |
| Interaction (ES:H) | * |
Table 5.
Quantification of anthocyanins from pericarp of P. calyculatus from two different hosts by HPTLC.
Table 5.
Quantification of anthocyanins from pericarp of P. calyculatus from two different hosts by HPTLC.
| Host (H) | Extractive Solvent (ES) | C3GE mg/g DW |
|---|
| P. laevigata | Acidified ethanol | 201.846 ± 8.784 b |
| Ethanol | 118.384 ± 3.464 d |
| Q. deserticola | Acidified ethanol | 306.682 ± 11.804 a |
| Ethanol | 163.455 ± 3.854 c |
| | (ES) | * |
| (H) | * |
| Interaction (ES:H) | * |
Table 6.
Pearson’s correlation of biological activities and quantifications of metabolites in the ripe fruits of P. calyculatus.
Table 6.
Pearson’s correlation of biological activities and quantifications of metabolites in the ripe fruits of P. calyculatus.
| | TPC | TFC | TAC | C3G | DPPH● | ABTS●+ | MIC | MBC | ACE |
|---|
| TPC | 1 | 0.77 | 0.96 | 0.69 | 0.60 | 0.98 | −0.98 | −0.98 | −0.90 |
| TFC | | 1 | 0.83 | 0.89 | 0.88 | 0.68 | −0.79 | −0.79 | −0.43 |
| TAC | | | 1 | 0.85 | 0.75 | 0.95 | −0.99 | −0.99 | −0.78 |
| C3G | | | | 1 | 0.86 | 0.66 | −0.81 | −0.81 | −0.40 |
| DPPH● | | | | | 1 | 0.54 | −0.72 | −0.72 | −0.43 |
| ABTS●+ | | | | | | 1 | −0.96 | −0.96 | −0.84 |
| MIC | | | | | | | 1 | 1 | 0.84 |
| MBC | | | | | | | | 1 | 0.84 |
| ACE | | | | | | | | | 1 |
| Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).


 ,
, 


{kind=link}
{kind=link}