
Improving Flavonoid Accumulation of Bioreactor-Cultured Adventitious Roots in Oplopanax elatus Using Yeast Extract

Abstract
1. Introduction
2. Results
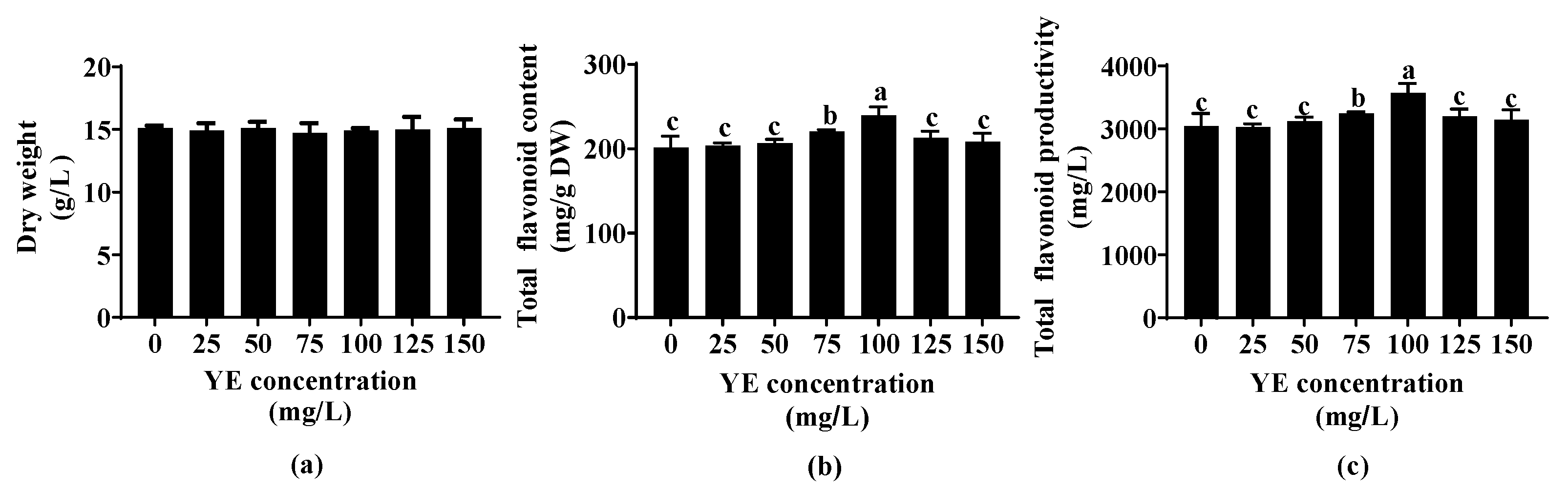
2.1. Effect of YE Concentration on Flavonoid Accumulation of Fed-Batch Bioreactor-Cultured ARs
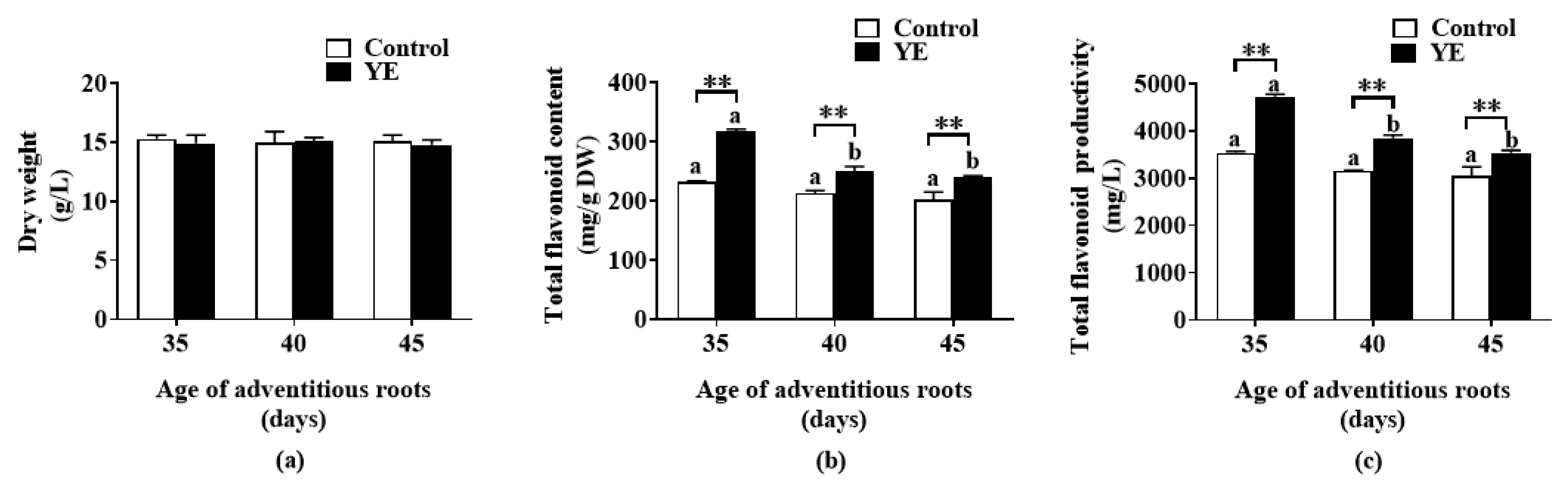
2.2. Effect of YE on Flavonoid Accumulation of Fed-Batch Bioreactor-Cultured ARs at Different Ages
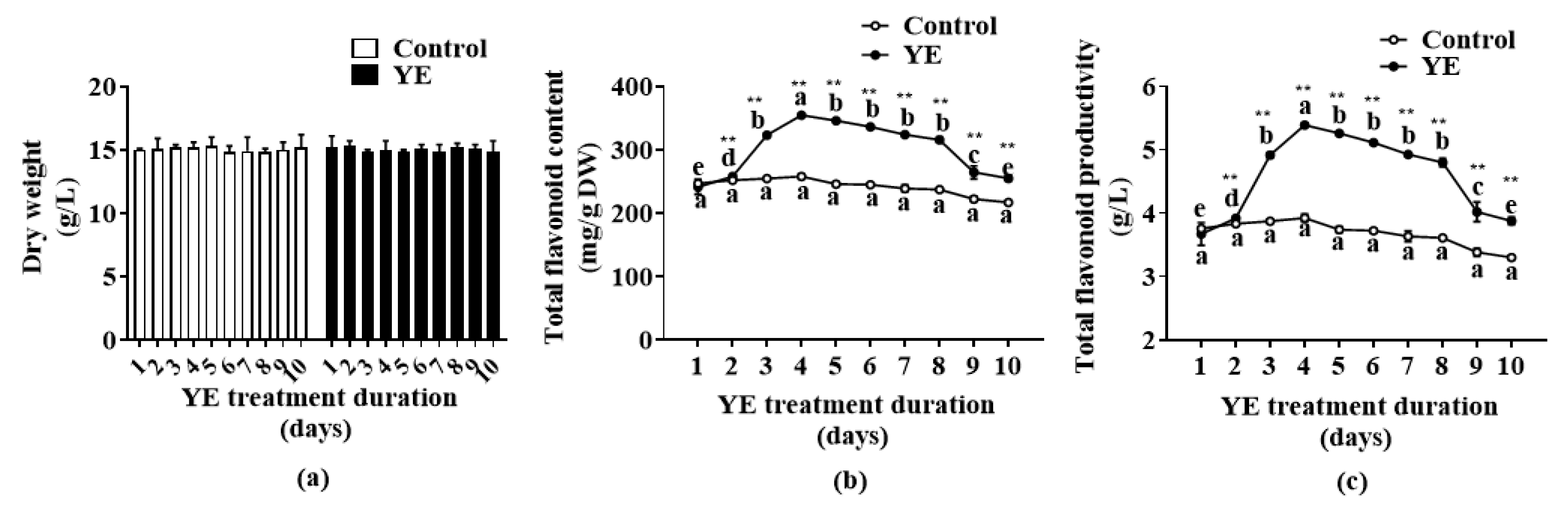
2.3. Effect of YE Treatment Duration on Flavonoid Accumulation of Fed-Batch Bioreactor-Cultured ARs
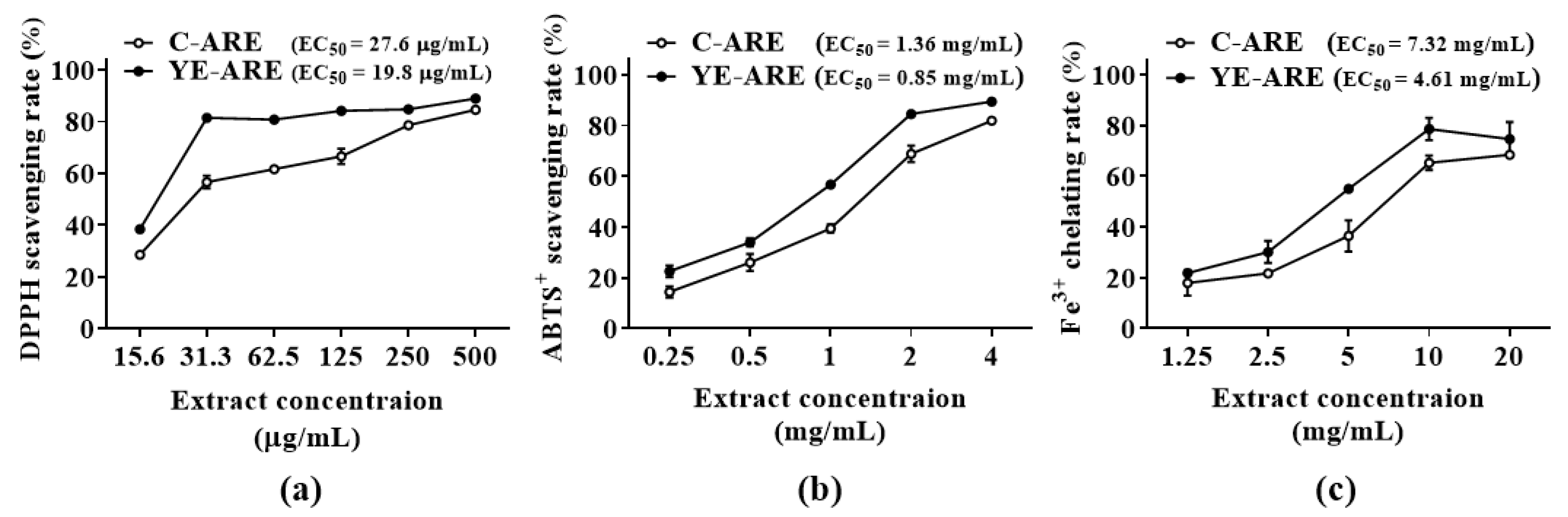
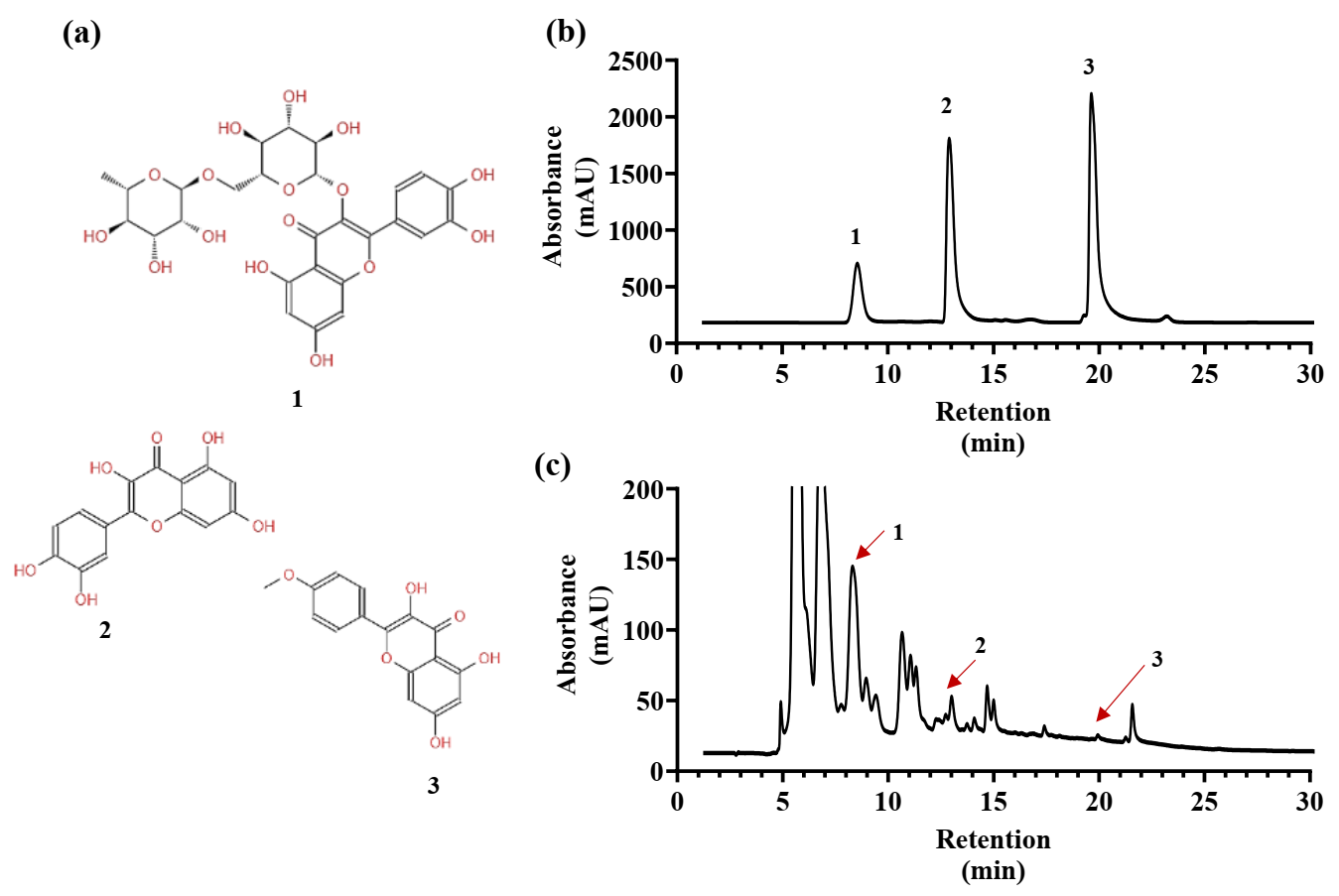
2.4. Comparison of Flavonoid Contents and Antioxidant Activities of YE-Treated and YE-Untreated ARs in Fed-Batch Bioreactor Culture System
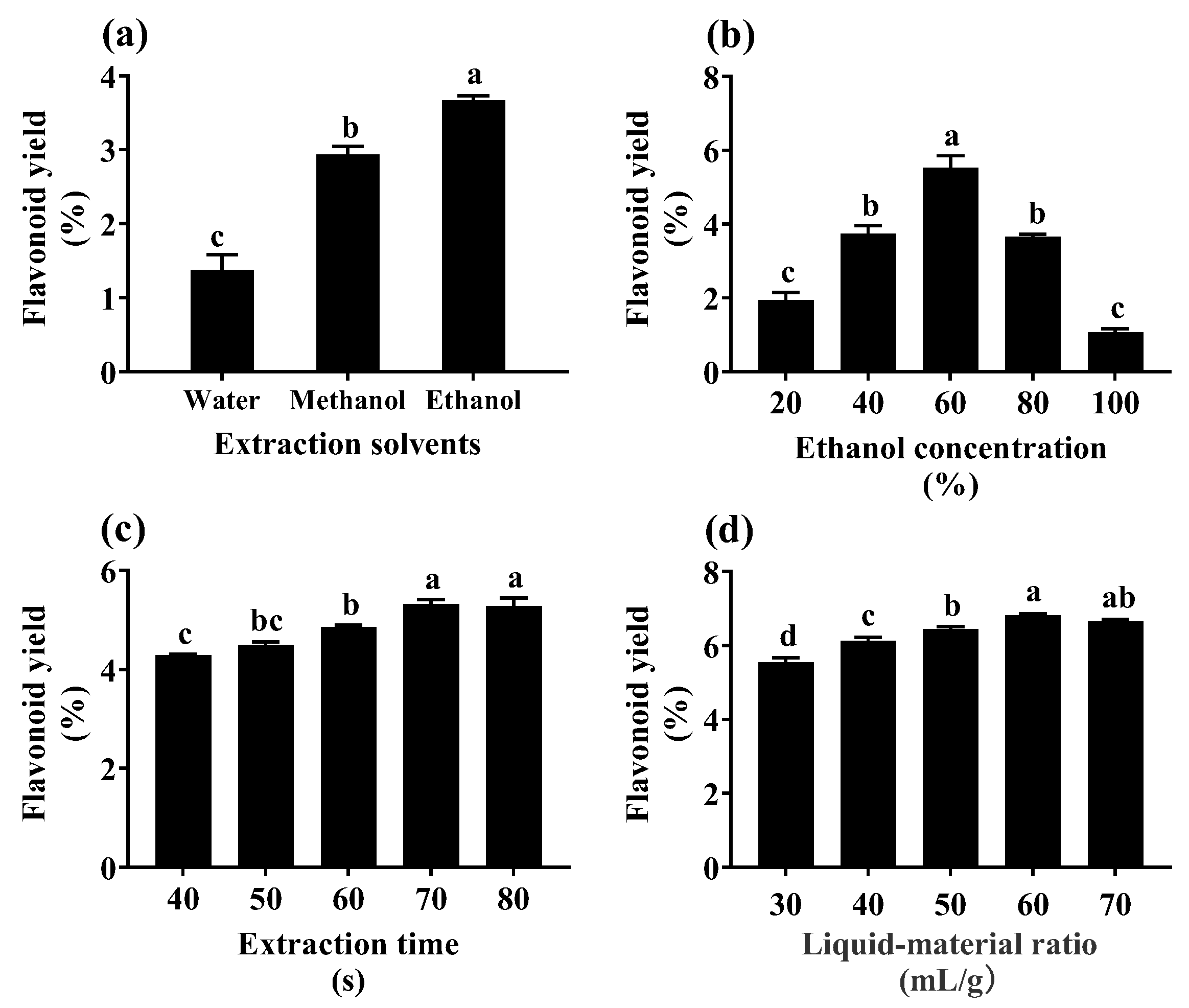
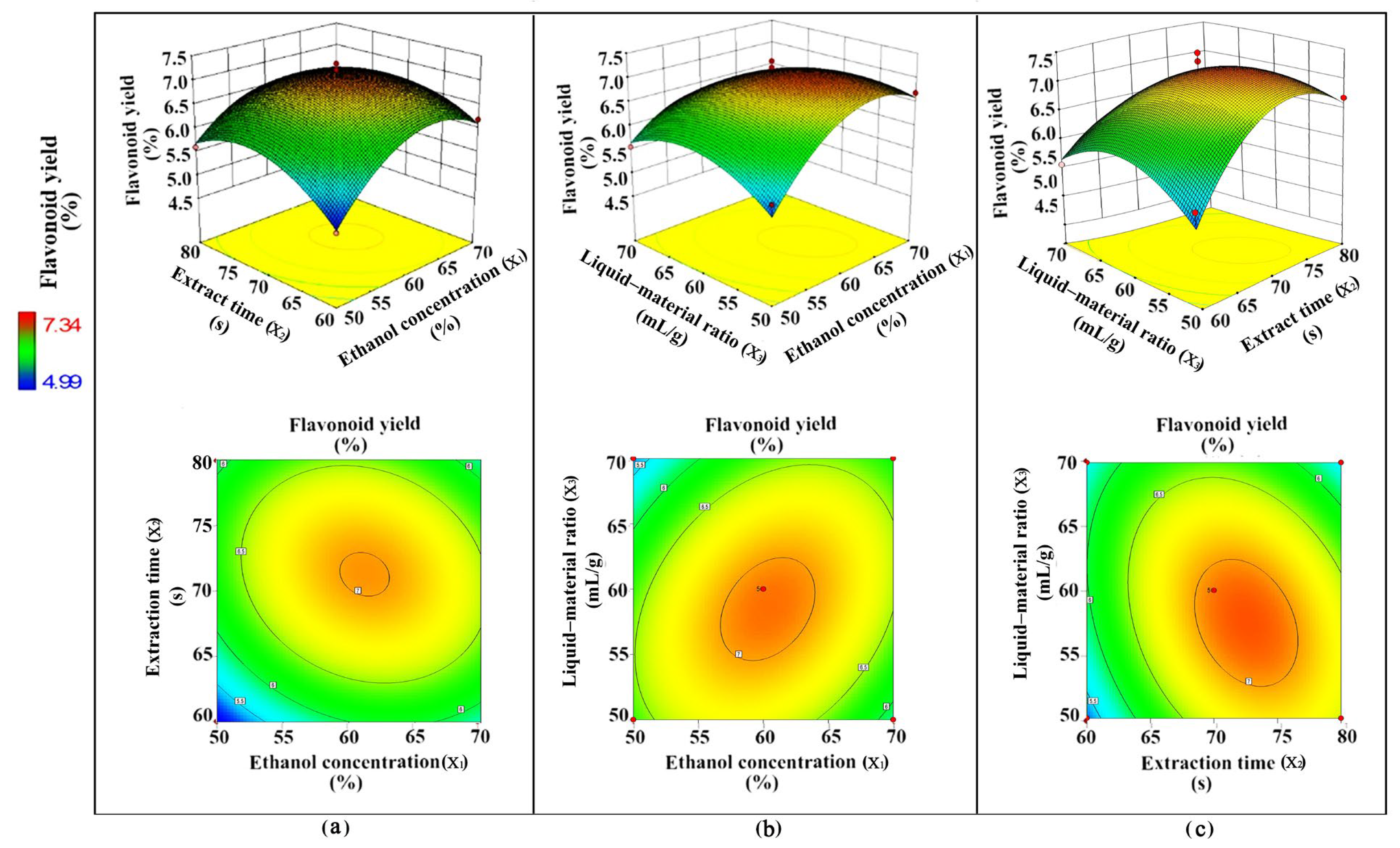
2.5. Flash Extraction Process Optimization of Flavonoids from YE-Treated Fed-Batch Bioreactor-Cultured ARs
3. Discussion
3.1. Optimization of YE Elicitation
3.2. Flavonoid Extraction Using Flash Extraction
4. Materials and Methods
4.1. Plant Materials and YE Solution Preparation
4.2. Fed-Batch AR Culture
4.3. YE Elicitation Experiment Design
4.4. Comparison of Flavonoid Contents and Antioxidant Activities between YE-Treated and YE-Untreated ARs in Fed-Batch Bioreactor Culture
4.5. Optimization of Extraction Process
4.6. Determination of AR Dry Weight
4.7. Determination of Flavonoid Content
4.8. Determination of Antioxidant Activities
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shikov, A.N.; Pozharitskaya, O.N.; Makarov, V.G.; Yang, W.Z.; Guo, D.A. Oplopanax elatus (Nakai) Nakai: Chemistry, traditional use and pharmacology. Chin. J. Nat. Med. 2014, 12, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.L.; Jin, M.Y.; Piao, X.C.; Yin, C.R.; Lian, M.L. Fed-batch culture of Oplopanax elatus adventitious roots: Feeding medium selection through comprehensive evaluation using an analytic hierarchy process. Biochem. Eng. J. 2021, 167, 107927. [Google Scholar] [CrossRef]
- Han, L.; Piao, X.C.; Jiang, Y.J.; Jiang, X.L.; Yin, C.R.; Lian, M.L. A high production of flavonoids and anthraquinones via adventitious root culture of Oplopanax elatus and evaluating antioxidant activity. Plant Cell Tiss. Organ Cult. 2019, 137, 173–179. [Google Scholar] [CrossRef]
- Jiang, X.L.; Piao, X.C.; Gao, R.; Jin, M.Y.; Jin, X.H.; Lian, M.L. Improvement of bioactive compound accumulation in adventitious root cultures of an endangered plant species, Oplopanax elatus. Acta Physiol. Plant. 2017, 39, 226. [Google Scholar] [CrossRef]
- Jiang, Y.J.; Piao, X.C.; Liu, J.S.; Lian, Z.X.; Kim, M.J.; Lian, M.L. Bioactive compound production by adventitious root culture of Oplopanax elatus in balloon-type airlift bioreactor systems and bioactivity property. Plant Cell Tiss. Organ Cult. 2015, 123, 413–425. [Google Scholar] [CrossRef]
- Jin, M.Y.; Piao, X.C.; Wu, X.H.; Fan, M.Z.; Li, X.F.; Yin, C.R.; Lian, M.L. Oplopanax elatus adventitious root production through fed-batch culture and their anti-bacterial effects. Plant Cell Tiss. Organ Cult. 2020, 140, 447–457. [Google Scholar] [CrossRef]
- Lee, K.J.; Park, Y.; Kim, J.Y.; Jeong, T.K.; Yun, K.S.; Paek, K.; Park, S.Y. Production of biomass and bioactive compounds from adventitious root cultures of Polygonum multiflorum using air-lift bioreactors. J. Plant Biotechnol. 2015, 42, 34–42. [Google Scholar] [CrossRef]
- Linh, N.T.N.; Cuong, L.K.; Tam, H.T.; Tung, H.T.; Luan, V.Q.; Hien, V.T.; Loc, N.H.; Nhut, D.T. Improvement of bioactive saponin accumulation in adventitious root cultures of Panax vietnamensis via culture periods and elicitation. Plant Cell Tiss. Organ Cult. 2019, 137, 101–113. [Google Scholar] [CrossRef]
- Wu, C.H.; Tang, J.; Jin, Z.X.; Wang, M.; Liu, Z.Q.; Huang, T.; Lian, M.L. Optimizing co-culture conditions of adventitious roots of Echinacea pallida and Echinacea purpurea in air-lift bioreactor systems. Biochem. Eng. J. 2018, 132, 206–216. [Google Scholar] [CrossRef]
- Jin, M.Y.; Hao, Y.J.; Zhang, K.X.; Yin, C.R.; Jiang, J.; Piao, X.C.; Lian, M.L. Fed-batch culture of Oplopanax elatus adventitious roots: Establishment of a complete culture system. Biochem. Eng. J. 2023, 194, 108898. [Google Scholar] [CrossRef]
- Choi, S.; Son, S.; Yun, S.; Kwon, O.; Seon, J.; Paek, K. Pilot-scale culture of adventitious roots of ginseng in a bioreactor system. Plant Cell Tiss. Organ Cult. 2000, 62, 187–193. [Google Scholar] [CrossRef]
- Cui, H.Y.; Baque, M.A.; Lee, E.J.; Paek, K. Scale-up of adventitious root cultures of Echinacea angustifolia in a pilot-scale bioreactor for the production of biomass and caffeic acid derivatives. Plant Biotechnol. Rep. 2013, 7, 297–308. [Google Scholar] [CrossRef]
- Ho, T.T.; Lee, K.J.; Lee, J.D.; Bhushan, S.; Paek, K.; Park, S.Y. Adventitious root culture of Polygonum multiflorum for phenolic compounds and its pilot-scale production in 500 L-tank. Plant Cell Tiss. Organ Cult. 2017, 130, 167–181. [Google Scholar] [CrossRef]
- An, X.L.; Yu, Y.; Fan, M.Z.; Wu, X.H.; Li, X.F.; Piao, X.C.; Lian, M.L. A fungal mycelium elicitor efficiently improved ginsenoside synthesis during adventitious root culture of Panax ginseng. J. Plant Biochem. Biotechnol. 2022, 31, 657–664. [Google Scholar] [CrossRef]
- Ho, T.T.; Lee, J.D.; Jeong, C.S.; Paek, K.; Park, S.Y. Improvement of biosynthesis and accumulation of bioactive compounds by elicitation in adventitious root cultures of Polygonum multiflorum. Appl. Microbiol. Biotechnol. 2018, 102, 199–209. [Google Scholar] [CrossRef]
- Le, K.C.; Im, W.T.; Paek, K.; Park, S.Y. Biotic elicitation of ginsenoside metabolism of mutant adventitious root culture in Panax ginseng. Appl. Microbiol. Biotechnol. 2018, 102, 1687–1697. [Google Scholar] [CrossRef]
- Huang, L.; Xiao, W.J.; Yang, G.; Mo, G.; Lin, S.F.; Wu, Z.G.; Guo, L.P. Mechanism exploration on synthesis of secondary metabolites in Sorbus aucuparia cell cultures treated with yeast extract. Chin. Trad. Herb. Drugs. 2014, 39, 2019–2023. [Google Scholar]
- Goncharuk, E.A.; Saibel, O.L.; Zaitsev, G.P.; Zagoskina, N.V. The elicitor effect of yeast extract on the accumulation of phenolic compounds in Linum grandiflorum cells Cultured in vitro and their antiradical activity. Biol Bull. 2022, 49, 620–628. [Google Scholar] [CrossRef]
- Maqsood, M.; Abdul, M. Yeast extract elicitation increases vinblastine and vincristine yield in protoplast derived tissues and plantlets in Catharanthus roseus. Rev. Bras. Farmacogn. 2017, 27, 549–556. [Google Scholar] [CrossRef]
- Liu, Y.; Yuan, K.; Ji, C. New method of extraction on the chemical components of Chinese medicinal plants-extracting method by smashing of plant tissue (EMS). Henan. Sci. 1993, 11, 265–268. [Google Scholar]
- Latif, A.; Maqbool, A.; Zhou, R.Z.; Arsalan, M.; Sun, K.; Si, Y.B. Optimized degradation of bisphenol A by immobilized laccase from Trametes versicolor using Box-Behnken design (BBD) and artificial neural network (ANN). J. Environ. Chem. Eng. 2022, 10, 107331. [Google Scholar] [CrossRef]
- Hao, Y.J.; Zhang, K.X.; Jin, M.Y.; Piao, X.C.; Lian, M.L.; Jiang, J. Improving fed-batch culture efficiency of Rhodiola sachalinensis cells and optimizing flash extraction process of polysaccharides from the cultured cells by BBD-RSM. Ind. Crop. Prod. 2023, 196, 116513. [Google Scholar] [CrossRef]
- Lee, E.J.; Park, S.Y.; Paek, K. Enhancement strategies of bioactive compound production in adventitious root cultures of Eleutherococcus koreanum Nakai subjected to methyl jasmonate and salicylic acid elicitation through airlift bioreactors. Plant Cell Tiss. Organ Cult. 2014, 120, 1–10. [Google Scholar] [CrossRef]
- Li, B.; Wang, B.; Li, H.; Peng, L.; Ru, M.; Liang, Z.; Yan, X.; Zhu, Y. Establishment of Salvia castanea Diels f. tomentosa Stib. hairy root cultures and the promotion of tanshinone accumulation and gene expression with Ag⁺, methyl jasmonate, and yeast extract elicitation. Protoplasma 2016, 253, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Kim, S.B.; Jo, Y.; Ahn, J.; Turk, A.; Kim, D.; Chang, B.; Kim, S.Y.; Jeong, C.S.; Hwang, B.Y.; et al. Curcubinoyl-conjugated flavonoids from methyl jasmonate-treated wild ginseng adventitious root cultures. Sci. Rep. 2021, 11, 12212. [Google Scholar] [CrossRef]
- Wang, J.A.; Gao, W.Y.; Zhang, J.; Huang, T.; Wen, T.T.; Huang, L.Q. Combination effect of lactoalbumin hydrolysate and methyl jasmonate on ginsenoside and polysaccharide production in Panax quinquefolium L. cells cultures. Acta Physiol. Plant. 2011, 33, 861–866. [Google Scholar] [CrossRef]
- Farjaminezhad, R.; Garoosi, G. Improvement and prediction of secondary metabolites production under yeast extract elicitation of Azadirachta indica cell suspension culture using response surface methodology. AMB Express 2021, 11, 1–16. [Google Scholar] [CrossRef]
- Loc, N.H.; Giang, N.T. Effects of elicitors on the enhancement of asiaticoside biosynthesis in cell cultures of centella (Centella asiatica L. Urban). Chem. Pap. 2012, 66, 642–648. [Google Scholar] [CrossRef]
- Vijayalakshmi, U.; Shourie, A. Yeast extract-mediated elicitation of anti-cancerous compounds licoisoflavone B, licochalcone A, and liquirtigenin in callus cultures of Glycyrrhiza glabra. Biotechnologia 2019, 100, 441–451. [Google Scholar] [CrossRef]
- Mahood, H.E.; Sarropoulou, V.; Tzatzani, T.T. Effect of explant type (leaf, stem) and 2,4-D concentration on callus induction: Influence of elicitor type (biotic, abiotic), elicitor concentration and elicitation time on biomass growth rate and costunolide biosynthesis in gazania (Gazania rigens) cell suspension cultures. Bioresour. Bioprocess. 2022, 9, 100. [Google Scholar]
- Abdul, M.N.A.; Kumar, I.S.; Nadarajah, K. Elicitor and receptor molecules: Orchestrators of plant defense and immunity. Int. J. Mol. Sci. 2020, 21, 963. [Google Scholar] [CrossRef] [PubMed]
- Kochan, E.; Szymczyk, P.; Kuźma, Ł.; Lipert, A.; Szymańska, G.S. Yeast extract stimulates ginsenoside production in hairy root cultures of American ginseng cultivated in shake flasks and nutrient sprinkle bioreactors. Molecules 2017, 22, 880–895. [Google Scholar] [CrossRef]
- Hao, Y.J.; An, X.L.; Sun, H.D.; Piao, X.C.; Gao, R.; Lian, M.L. Ginsenoside synthesis of adventitious roots in Panax ginseng is promoted by fungal suspension homogenate of Alternaria panax and regulated by several signaling molecules. Ind. Crop. Prod. 2020, 150, 112414. [Google Scholar] [CrossRef]
- Chen, H.; Chen, F. Effects of yeast elicitor on the growth and secondary metabolism of a high-tanshinone-producing line of the ti transformed Salvia miltiorrhiza cells in suspension culture. Process Biochem. 2000, 35, 837–840. [Google Scholar] [CrossRef]
- Yu, S.; Wu, X.H.; Wang, M.; Liu, L.L.; Ye, W.Q.; Jin, M.Y.; Piao, X.C.; Lian, M.L. Optimizing elicitation strategy of salicylic acid for flavonoid and phenolic production of fed-batch cultured Oplopanax elatus adventitious roots. J. Biotechnol. 2023, 368, 1–11. [Google Scholar] [CrossRef]
- Kan, L.N.; Wang, L.; Ding, Q.Z.; Wu, Y.W.; Ouyang, J. Flash extraction and physicochemical characterization of oil from Elaeagnus mollis Diels seeds. J. Oleo Sci. 2017, 66, 345–352. [Google Scholar] [CrossRef] [PubMed]
- Qin, D.Y.; Xi, J. Flash extraction: An ultra-rapid technique for acquiring bioactive compounds from plant materials. Trends Food Sci. Tech. 2021, 112, 581–591. [Google Scholar] [CrossRef]
- Xu, L.; Xu, J.; Shi, G.H.; Xiao, S.N.; Dai, R.K.; Wu, S.; Sun, B.S.; Zhang, X.S.; Zhao, Y.Q. Optimization of flash extraction, separation of ginsenosides, identification by HPLC-FT-ICR-MS and determination of rare ginsenosides in mountain cultivated ginseng. RSC Adv. 2020, 10, 44050–44057. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, C.; Huang, J.Q.; Xu, X.Q. Optimization of flash extraction of total polyphenols from leaves of Eriobotrya japonica lind by response surface methodology. Guangzhou Chem. Ind. 2019, 47, 112–115. [Google Scholar]
- Li, Y.; Zhao, T.M.; Huang, L.R.; Zou, T.; Wu, X.L. Response surface optimized extraction of flavonoids from Crotalaria ferruginea and antioxidant activities. Food Res. Dev. 2019, 40, 79–84. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth bioassays with tobacco tissues cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Zhang, W.; Piao, X.C.; Li, J.R.; Jin, Y.H.; Lian, M.L. Optimized culture medium for the production of flavonoids from Orostachys cartilaginea V.N. Boriss. callus cultures. In Vitro Cell. Dev. Biol.-Plant 2017, 53, 1–11. [Google Scholar] [CrossRef]
- Fu, R.; Zhang, Y.; Guo, Y.; Chen, F. Antioxidant and tyrosinase inhibition activities of the ethanol-insoluble fraction of water extract of Sapium sebiferum (L.) Roxb. leaves. S. Afr. J. Bot. 2014, 93, 98–104. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Total Flavonoids (mg/g DW) | Rutin (mg/g DW) | Quercetin (μg/g DW) | Kaempferide (μg/g DW) |
|---|---|---|---|---|
| control | 357.7 ± 38.9 | 2.4 ± 0.1 | 234.5 ± 10.3 | 117.4 ± 7.5 |
| YE | 582.2 ± 11.7 * | 2.9 ± 0.1 * | 266.6 ± 11.7 | 161.9 ± 9.7 * |
| Independent Variables | Coded Level | ||
|---|---|---|---|
| −1 | 0 | +1 | |
| ethanol concentration (X1) | 50% | 60% | 70% |
| extraction time (X2) | 60 s | 70 s | 80 s |
| liquid–material ratio (X3) | 50 mL/g | 60 mL/g | 70 mL/g |
| Groups | Codes (Levels) | Flavonoid Yield (%) | |||
|---|---|---|---|---|---|
| Ethanol Concentration (X1) | Extraction Time (X2) | Liquid−Material Ratio (X3) | Actual Value | Predicted Value | |
| 1 | 0 (60%) | −1 (60 s) | 1 (70 mL/g) | 5.56 ± 0.12 | 5.66 |
| 2 a | 0 (60%) | 0 (70 s) | 0 (60 mL/g) | 7.00 ± 0.03 | 7.06 |
| 3 | −1 (50%) | −1 (60 s) | 0 (60 mL/g) | 4.99 ± 0.08 | 5.04 |
| 4 | −1 (50%) | 0 (70 s) | 1 (70 mL/g) | 5.45 ± 0.11 | 5.30 |
| 5 a | 0 (60%) | 0 (70 s) | 0 (60 mL/g) | 6.92 ± 0.07 | 7.06 |
| 6 | 1(70%) | 0 (70 s) | 1 (70 mL/g) | 6.39 ± 0.16 | 6.22 |
| 7 | 0 (60%) | 1 (80 s) | −1 (50 mL/g) | 6.69 ± 0.01 | 6.60 |
| 8 | −1 (50%) | 1 (80 s) | 0 (60 mL/g) | 6.19 ± 0.05 | 6.10 |
| 9 | −1 (50%) | 0 (70 s) | −1 (50 mL/g) | 6.18 ± 0.15 | 6.36 |
| 10 | 1(70%) | 1 (80 s) | 0 (60 mL/g) | 5.90 ± 0.06 | 5.84 |
| 11 | 0 (60%) | 0 (70 s) | 0 (60 mL/g) | 6.82 ± 0.04 | 7.06 |
| 12 | 0 (60%) | −1 (60 s) | −1 (50 mL/g) | 5.50 ± 0.11 | 5.26 |
| 13 | 0 (60%) | 1 (80 s) | 1 (70 mL/g) | 5.29 ± 0.12 | 5.52 |
| 14 | 1 (70%) | −1 (60 s) | 0 (60 mL/g) | 5.60 ± 0.12 | 5.70 |
| 15 | 1 (70%) | 0 (70 s) | −1 (50 mL/g) | 5.69 ± 0.07 | 5.84 |
| 16 a | 0 (60%) | 0 (70 s) | 0 (60 mL/g) | 7.34 ± 0.10 | 7.06 |
| 17 a | 0 (60%) | 0 (70 s) | 0 (60 mL/g) | 7.21 ± 0.01 | 7.06 |
| Variables | Sun of Squares | Degree of Freedom | Mean Square | F | p |
|---|---|---|---|---|---|
| model | 8.14 | 9 | 0.90 | 14.37 | 0.0010 ** |
| X1 | 0.73 | 1 | 0.73 | 11.64 | 0.0113 * |
| X2 | 0.074 | 1 | 0.074 | 1.18 | 0.3137 |
| X3 | 0.23 | 1 | 0.23 | 3.73 | 0.0948 |
| X1X2 | 0.20 | 1 | 0.20 | 3.22 | 0.1159 |
| X1X3 | 0.53 | 1 | 0.53 | 8.47 | 0.0226 * |
| X2X3 | 0.51 | 1 | 0.51 | 8.13 | 0.0247 * |
| X12 | 2.55 | 1 | 2.55 | 40.49 | 0.0004 ** |
| X22 | 1.57 | 1 | 1.57 | 24.93 | 0.0016 ** |
| X32 | 1.14 | 1 | 1.14 | 18.12 | 0.0038 ** |
| residual | 0.44 | 7 | 0.063 | - | - |
| lack of fit | 0.26 | 3 | 0.086 | 1.90 | 0.2712 |
| pure error | 0.18 | 4 | 0.045 | - | - |
| cor total | 8.58 | 16 | - | - | - |
| R2 | 0.9487 |
| Experiment Repetition | Flavonoid Yield (%) | Average Flavonoid Yield (%) | RSD (%) |
|---|---|---|---|
| 1st | 7.04 | 7.08 | 1.58 |
| 2nd | 7.14 | ||
| 3th | 7.06 |
| Experiments | Solvent Type | Solvent Concentration (%) | Extraction Time (s) | Liquid−Material Ratio (mL/g) |
|---|---|---|---|---|
| 1 | Water, ethanol, methanol | 80 | 60 | 40 |
| 2 | Ethanol | 40, 50, 60, 70, 80 | 60 | 40 |
| 3 | Ethanol | 80 | 40, 50, 60, 70, 80 | 40 |
| 4 | Ethanol | 80 | 70 | 30, 40, 50, 60, 70 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, M.-Y.; Wang, M.; Wu, X.-H.; Fan, M.-Z.; Li, H.-X.; Guo, Y.-Q.; Jiang, J.; Yin, C.-R.; Lian, M.-L. Improving Flavonoid Accumulation of Bioreactor-Cultured Adventitious Roots in Oplopanax elatus Using Yeast Extract. Plants 2023, 12, 2174. https://doi.org/10.3390/plants12112174
Jin M-Y, Wang M, Wu X-H, Fan M-Z, Li H-X, Guo Y-Q, Jiang J, Yin C-R, Lian M-L. Improving Flavonoid Accumulation of Bioreactor-Cultured Adventitious Roots in Oplopanax elatus Using Yeast Extract. Plants. 2023; 12(11):2174. https://doi.org/10.3390/plants12112174
Chicago/Turabian StyleJin, Mei-Yu, Miao Wang, Xiao-Han Wu, Ming-Zhi Fan, Han-Xi Li, Yu-Qing Guo, Jun Jiang, Cheng-Ri Yin, and Mei-Lan Lian. 2023. "Improving Flavonoid Accumulation of Bioreactor-Cultured Adventitious Roots in Oplopanax elatus Using Yeast Extract" Plants 12, no. 11: 2174. https://doi.org/10.3390/plants12112174
APA StyleJin, M.-Y., Wang, M., Wu, X.-H., Fan, M.-Z., Li, H.-X., Guo, Y.-Q., Jiang, J., Yin, C.-R., & Lian, M.-L. (2023). Improving Flavonoid Accumulation of Bioreactor-Cultured Adventitious Roots in Oplopanax elatus Using Yeast Extract. Plants, 12(11), 2174. https://doi.org/10.3390/plants12112174