
Unlocking the Nexus between Leaf-Level Water Use Efficiency and Root Traits Together with Gas Exchange Measurements in Rice (Oryza sativa L.)

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Results
2.1. Genotype Performance and Significance of ANOVA for Gas Exchange Measurements and Root Traits
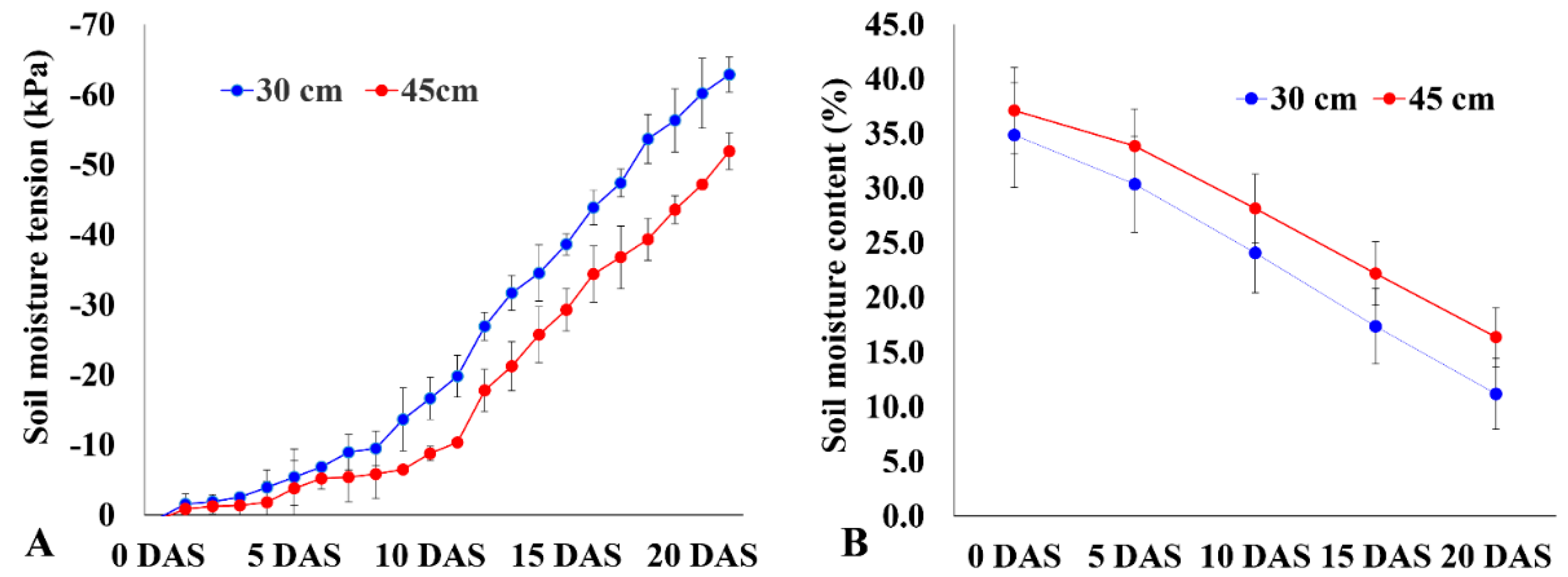
2.2. Variation in Instantaneous WUE (WUEi) and Biomass among Rice Accessions under Varying Moisture Levels
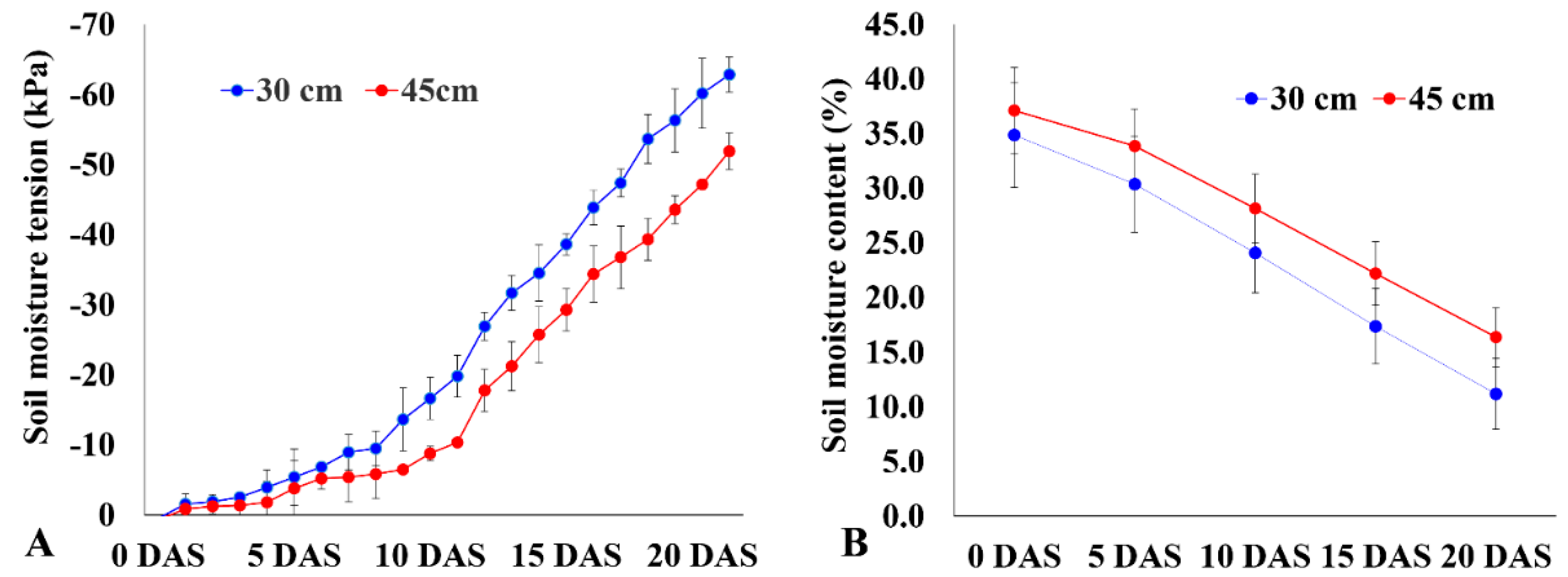
2.3. Differences in Root Traits among Genotypes under Varying Moisture Levels
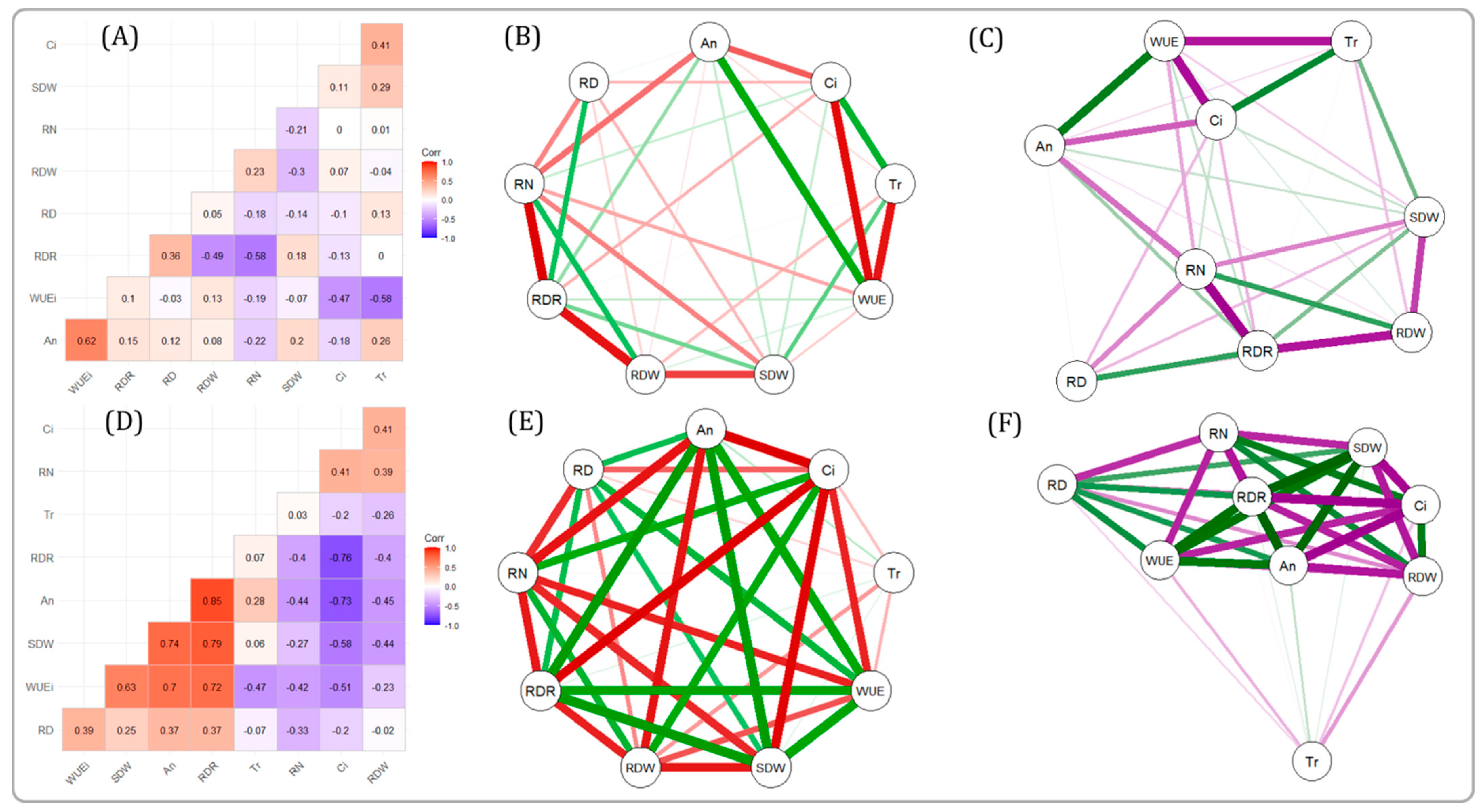
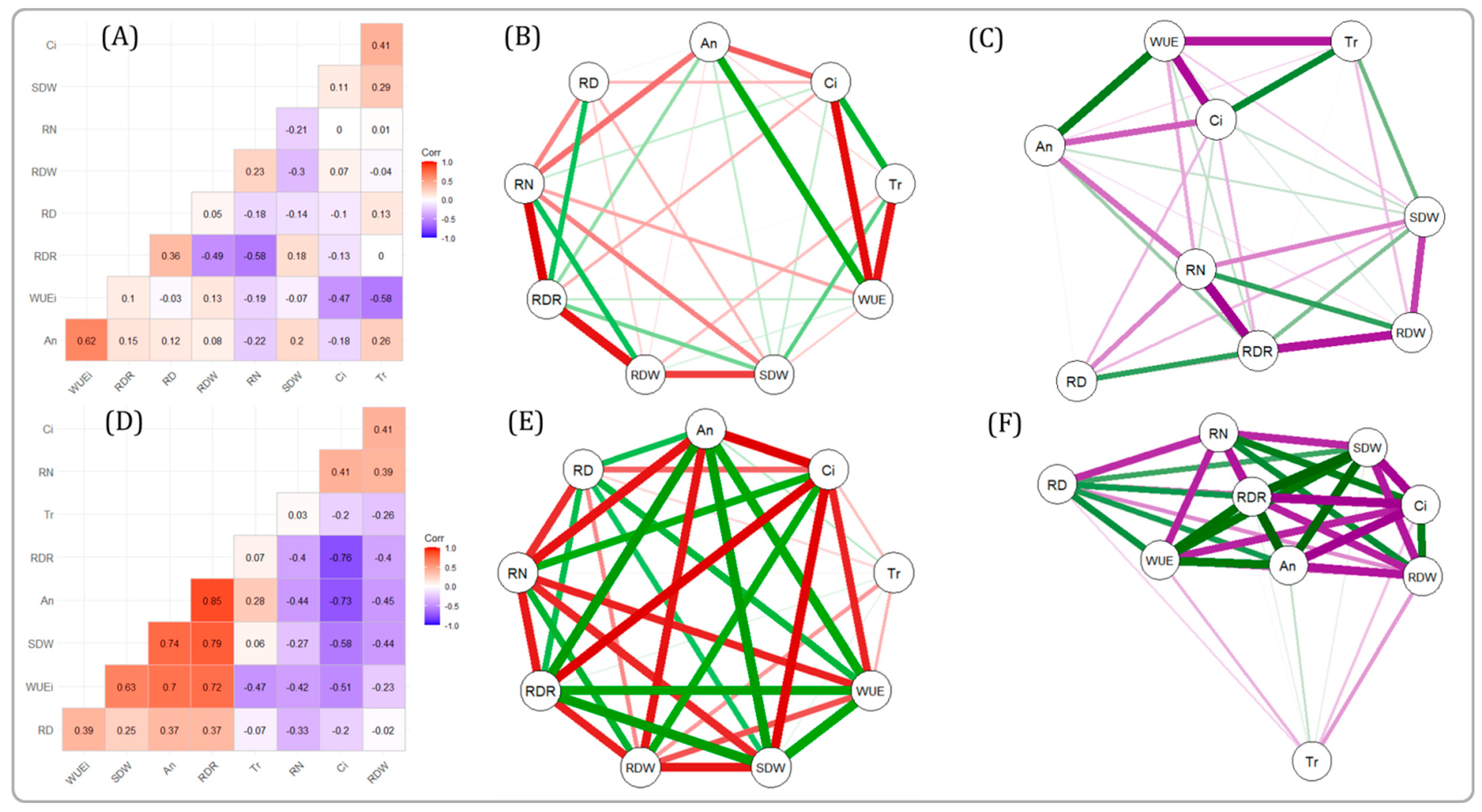
2.4. Correlation between Different Gas Exchange Traits and Root Traits
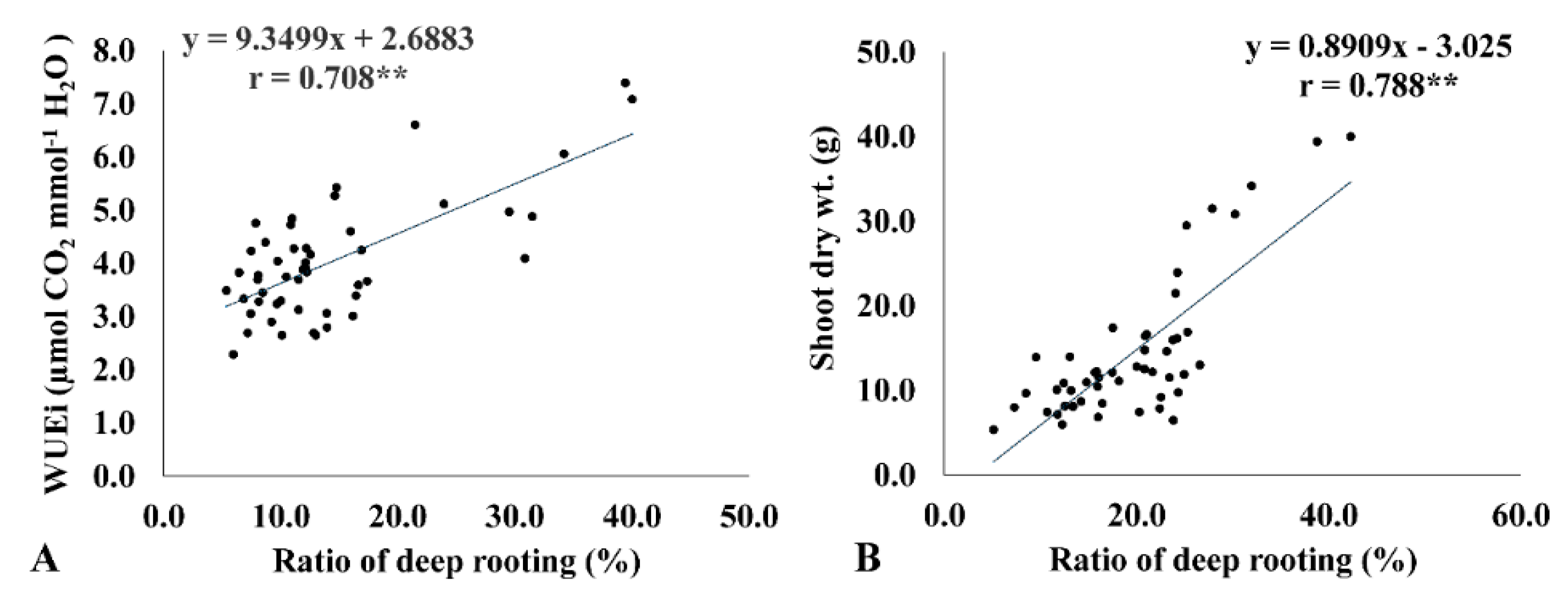
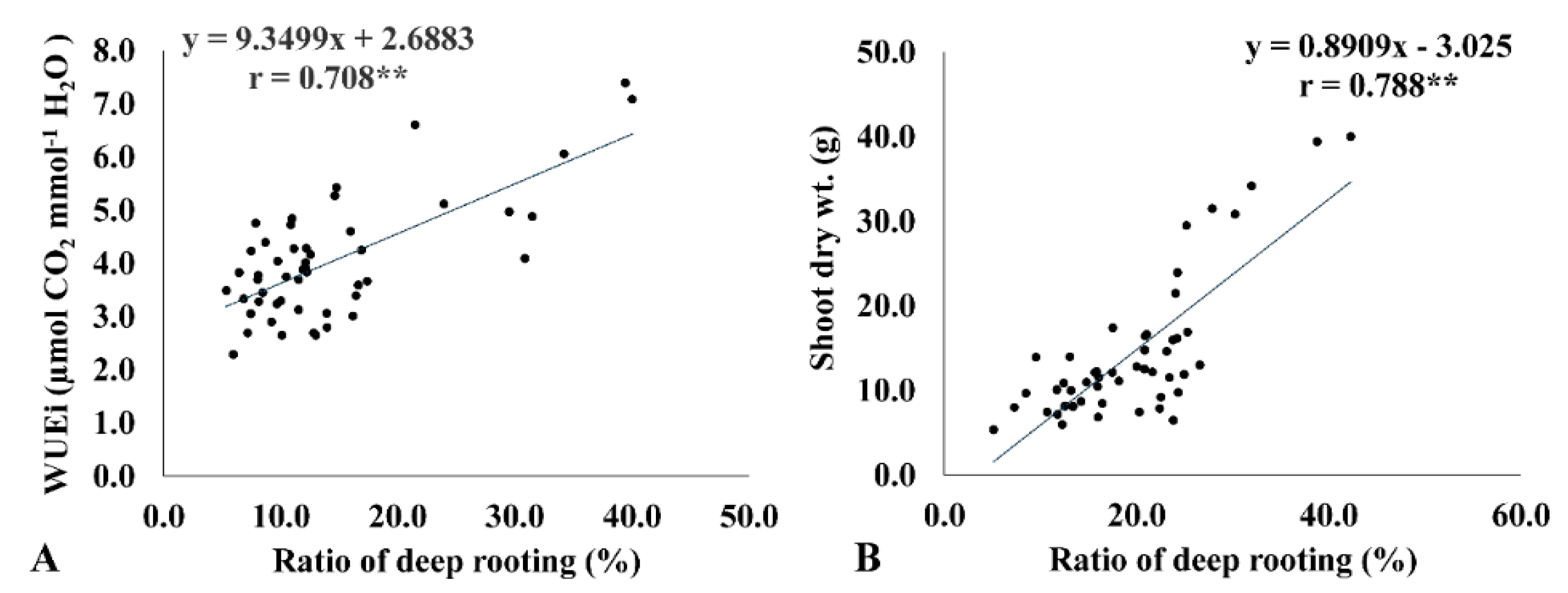
2.5. Instantaneous WUE (WUEi) Is Correlated with the Ratio of Deep Rooting and Biomass Accumulation
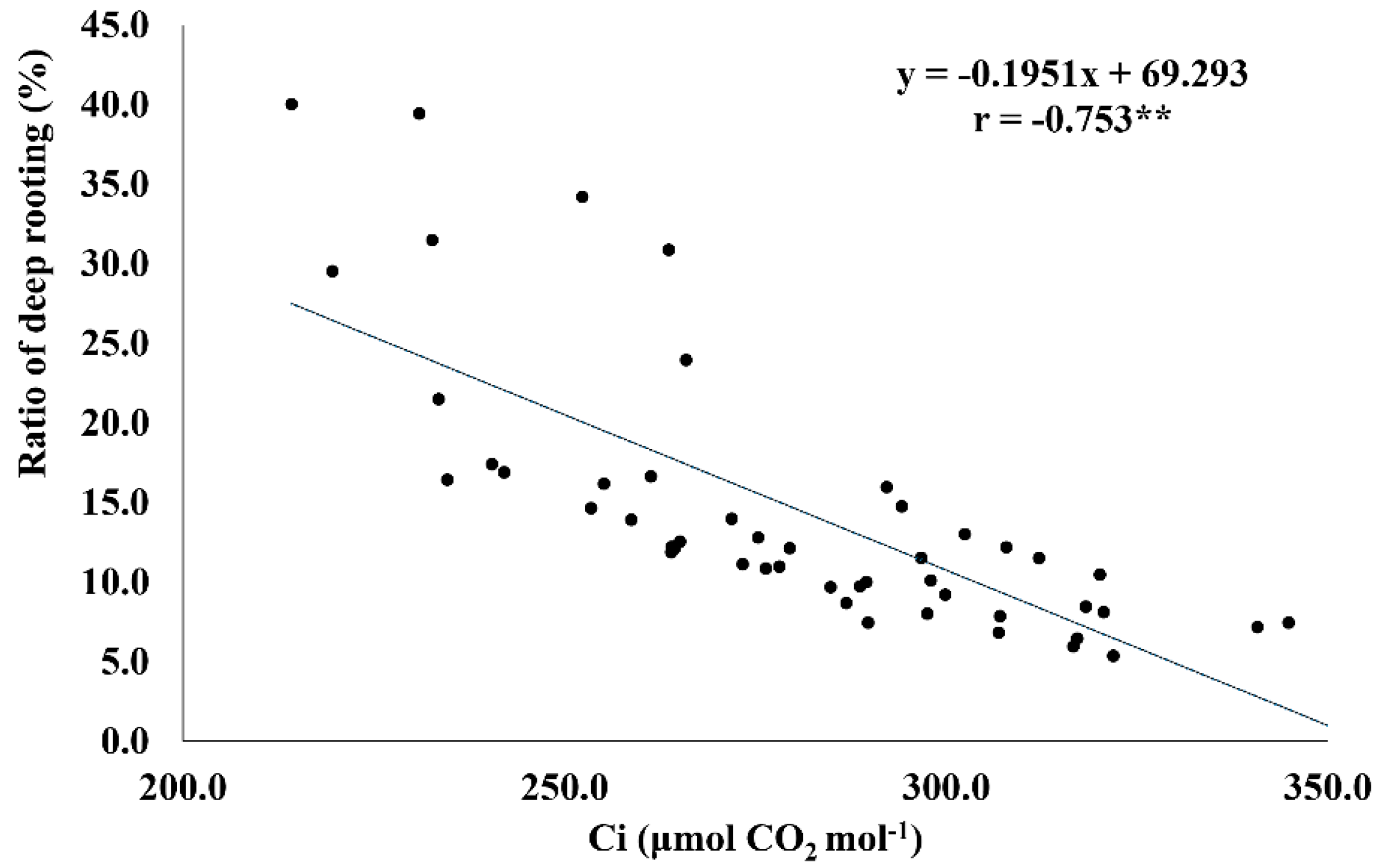
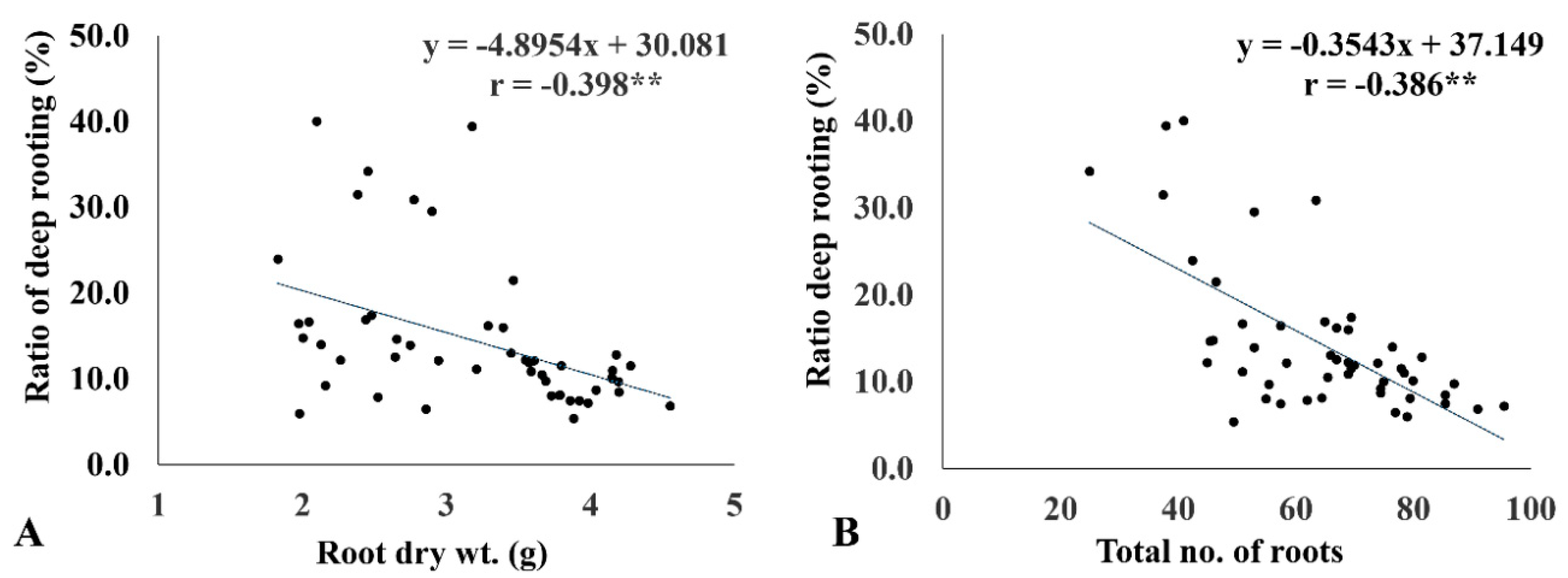
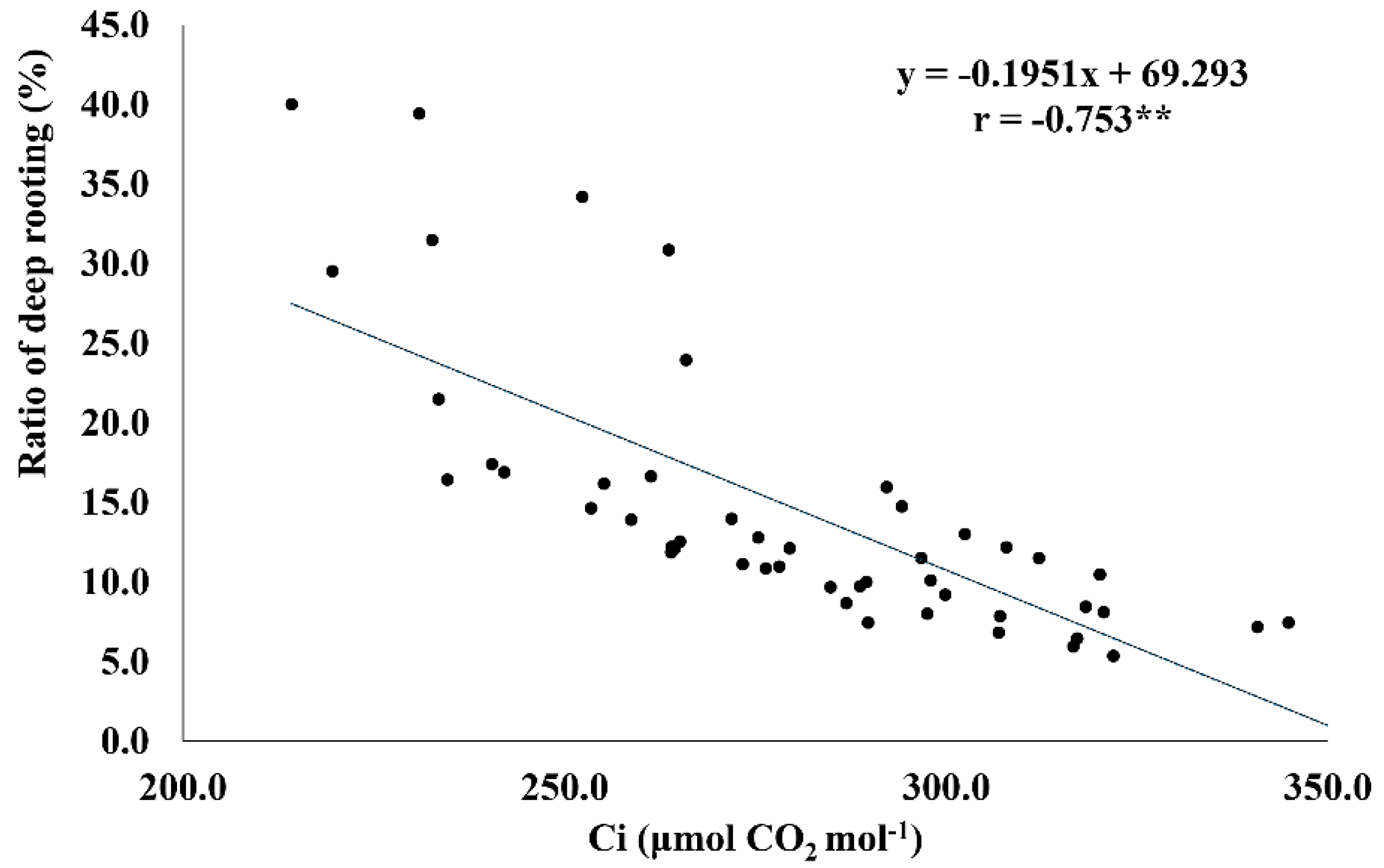
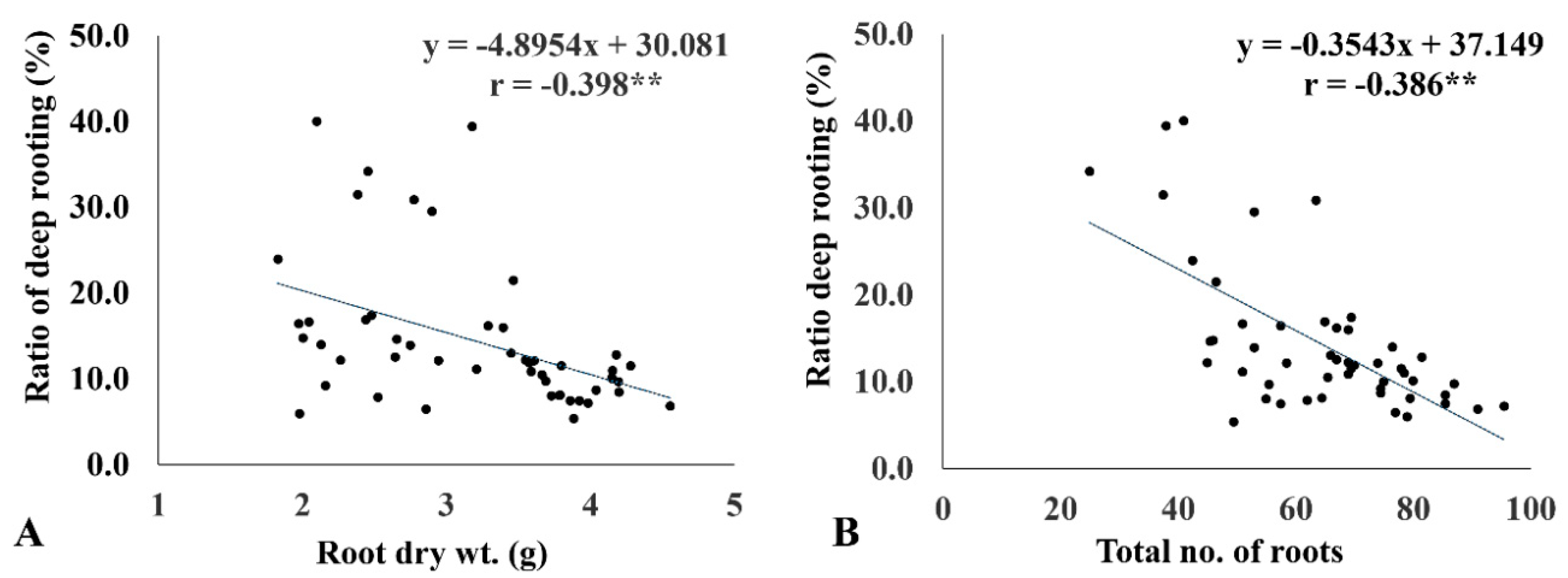
2.6. RDR Is Associated with Low Internal CO2 Concentration, Root Dry Weight, and Total Number of Roots
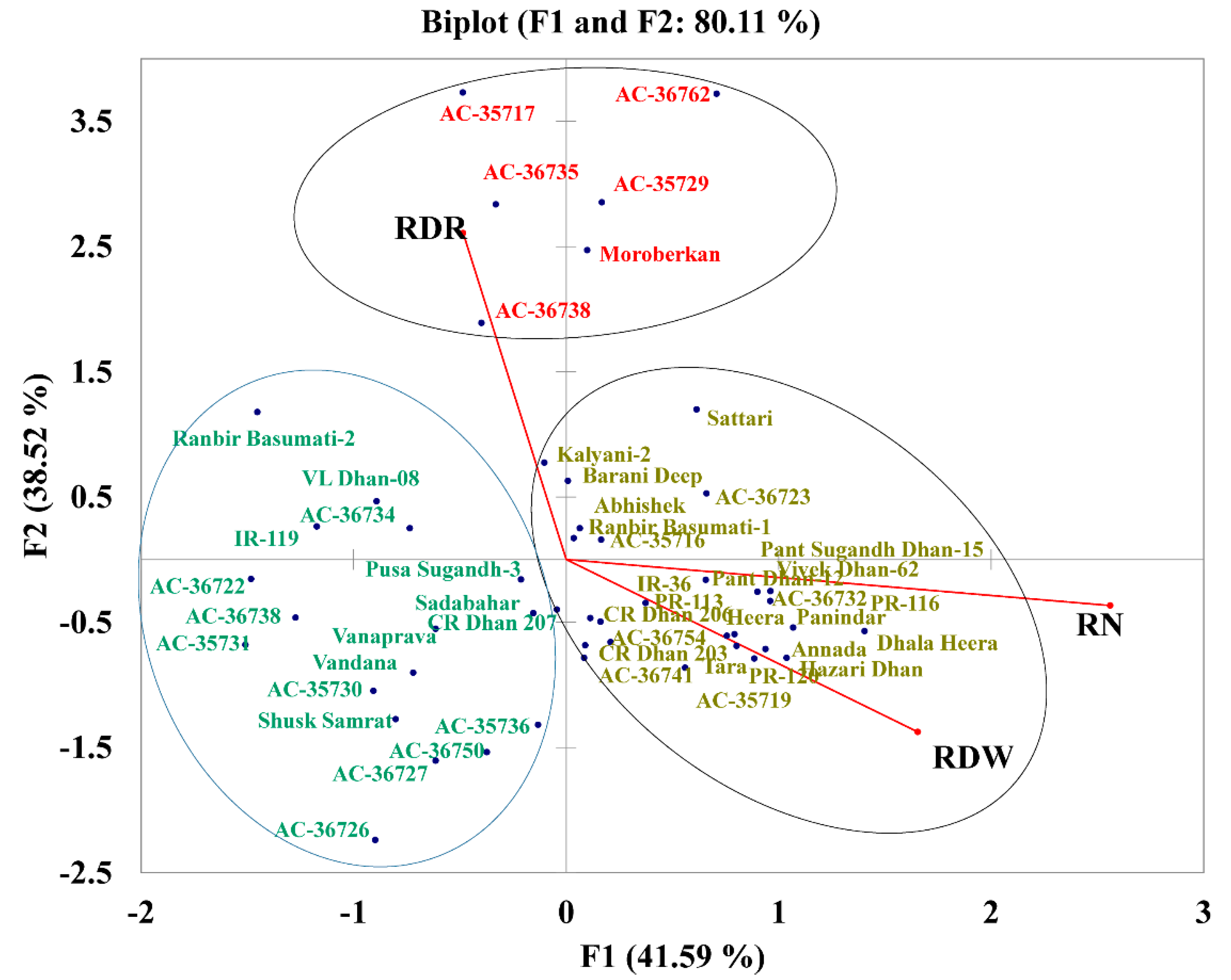
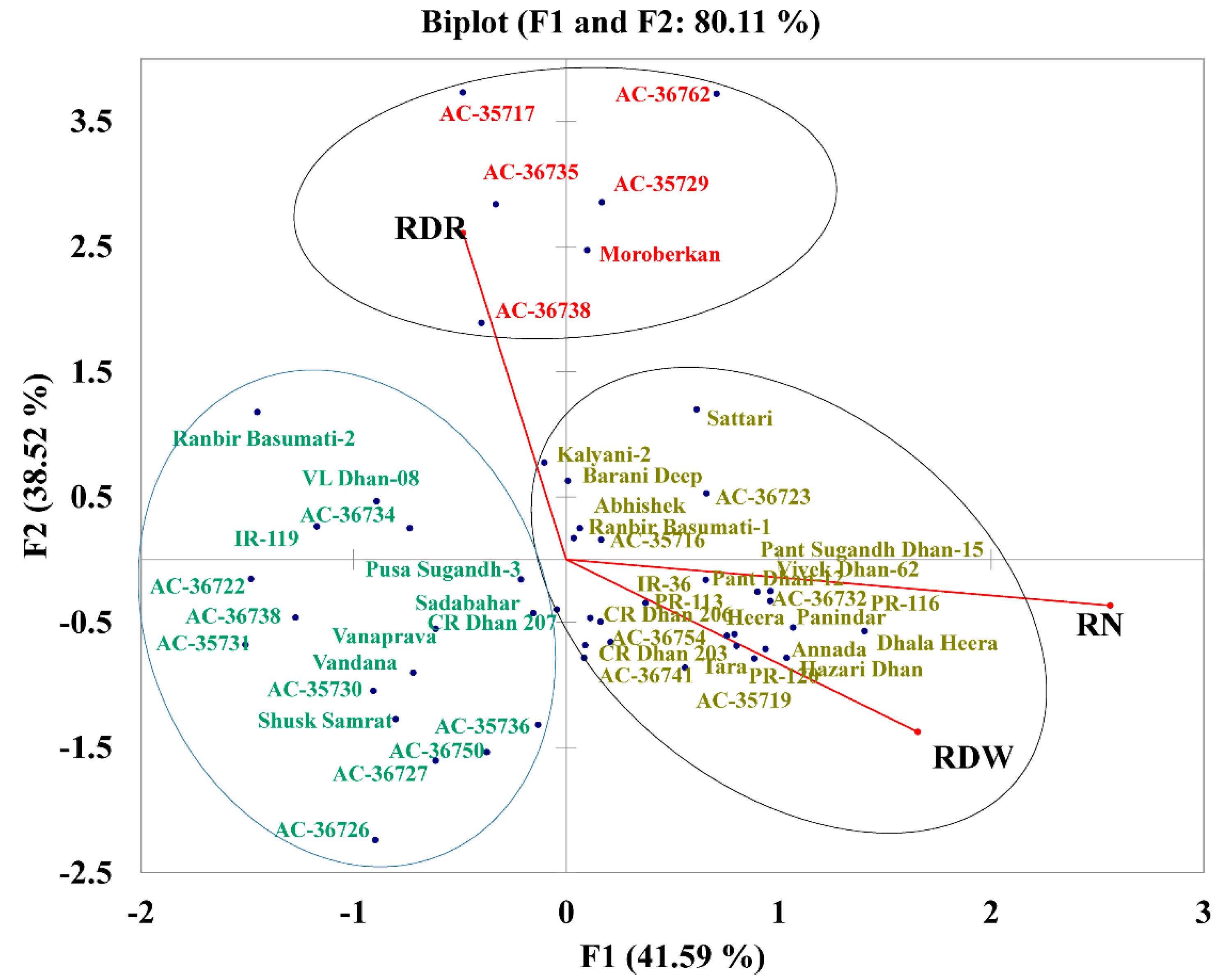
2.7. PCA with Root Traits Grouped the Genotypes into Distinct Categories Highlighting the Importance of RDR
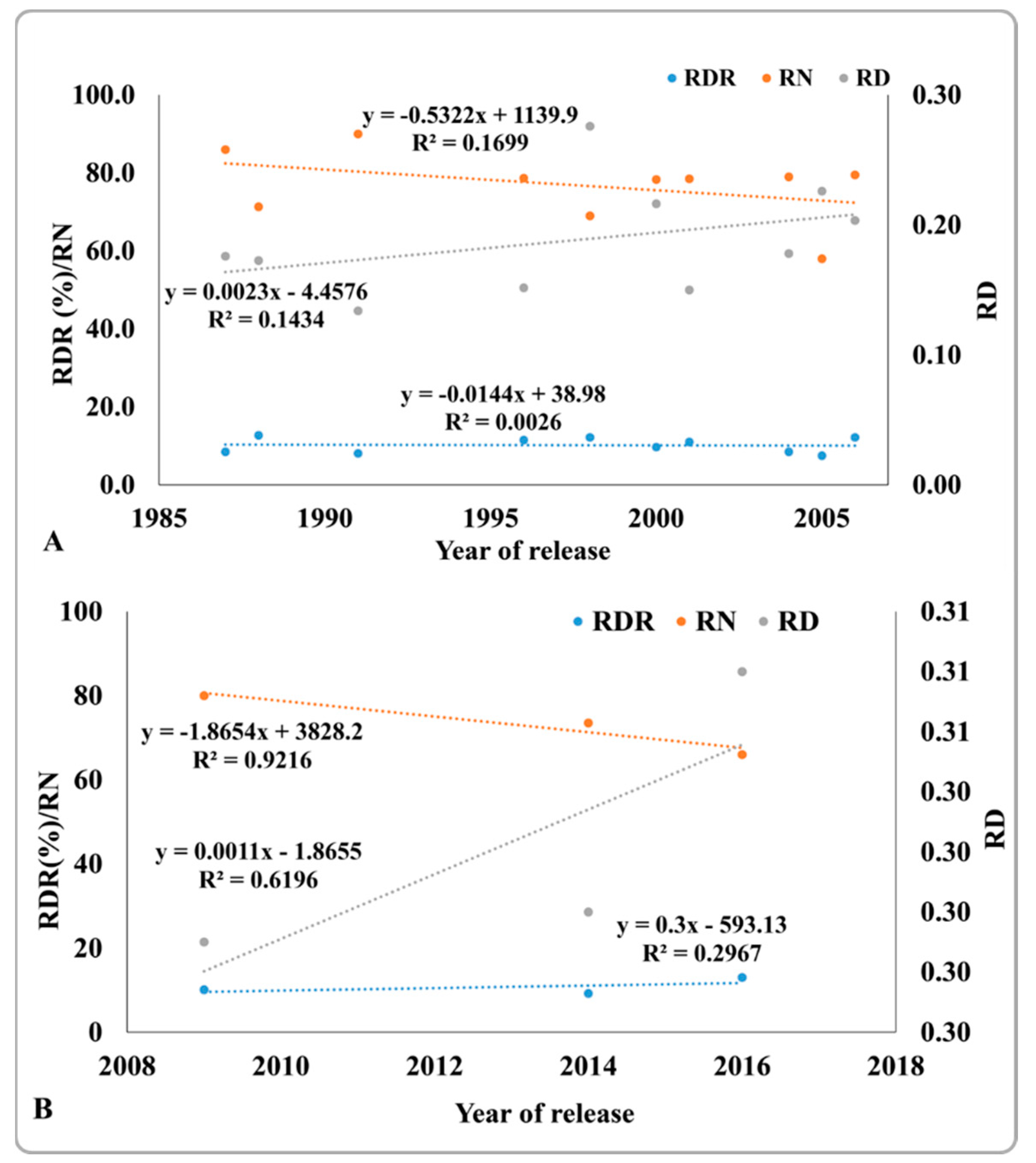
2.8. Changes in Root Traits in Improved Genotypes from 1987 to 2016
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.1.1. Root Trait Measurements
4.1.2. Gas Exchange Measurements
4.2. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Godfray, H.C.J.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. Food security: The challenge of feeding 9 billion people. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porter, J.R.; Xie, L.; Challinor, A.J.; Cochrane, K.; Howden, S.M.; Iqbal, M.M.; Lobell, D.B.; Travasso, M.I. Food security and food production systems. In Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Field, C.B., Barros, V.R., Dokken, D.J., Mach, K.J., Mastrandrea, M.D., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., Girma, B., Kissel, E.S., Levy, A.N., MacCracken, S., Mastrandrea, P.R., White, L.L., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; pp. 485–533. [Google Scholar]
- Vikram, P.; Swamy, B.P.M.; Dixit, S.; Singh, R.; Singh, B.P.; Miro, B.; Kohli, A.; Henry, A.; Singh, N.K.; Kumar, A. Drought susceptibility of modern rice varieties: An effect of linkage of drought tolerance with undesirable traits. Sci. Rep. 2015, 13, 14799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korres, N.E.; Norsworthy, J.K.; Burgos, N.R.; Oosterhuis, D.M. Temperature and drought impacts on rice production: An agronomic perspective regarding short- and long-term adaptation measures. Water Resour. Rural. Dev. 2017, 9, 12–27. [Google Scholar] [CrossRef]
- Bacom, M. Water Use Efficiency in Plant Biology, 1st ed.; John Wiley & Sons: Hoboken, NJ, USA, 2009; Available online: https://www.routledge.com/Water-Use-Efficiency-in-Plant-Biology/Bacon/p/book/9780849323546 (accessed on 13 May 2021).
- Elert, E. Rice by the numbers: A good grain. Nature 2014, 514, S50–S51. [Google Scholar] [CrossRef] [Green Version]
- Surendran, U.; Raja, P.; Jayakumar, M.; Subramoniam, S.R. Use of efficient water saving techniques for production of rice in india under climate change scenario: A critical review. J. Clean. Prod. 2021, 309, 127272. [Google Scholar] [CrossRef]
- Matsuda, S.; Takano, S.; Sato, M.; Furukawa, K.; Nagasawa, H.; Yoshikawa, S.; Kasuga, J.; Tokuji, Y.; Yazaki, K.; Nakazono, M.; et al. Rice stomatal closure requires guard cell plasma membrane ATP-binding cassette transporter RCN1/OsABCG5. Mol. Plant 2016, 9, 417–427. [Google Scholar] [CrossRef] [Green Version]
- Maria, J.; Parfitt, B.; Concenço, G.; Scivittaro, W.B.; Andres, A.; Trombetta Da Silva, J.; Alves, M.; Pinto, B. Soil and water management for sprinkler irrigated rice in southern Brazil. In Advances in International Rice Research; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Shareef, T.M.E.; Ma, Z.; Zhao, B.; Shareef, T.M.E.; Ma, Z.; Zhao, B. Essentials of drip irrigation system for saving water and nutrients to plant roots: As a guide for growers. J. Water Resour. Prot. 2019, 11, 1129–1145. [Google Scholar] [CrossRef] [Green Version]
- Patek, R.; Raha, P.; Laxmi, S.; Paul, A. Effect of antitranspirants on rice (Oryza sativa) grown under submerged and water stressed condition in an Inceptisol of Varanasi, Uttar Pradesh. J. Crop Weed 2019, 15, 100–106. [Google Scholar] [CrossRef]
- Abdallah, M.M.S.; El-Bassiouny, H.M.S.; AbouSeeda, M.A. Potential role of kaolin or potassium sulfate as anti-transpirant on improving physiological, biochemical aspects and yield of wheat plants under different watering regimes. Bull. Natl. Res. Cent. 2019, 43, 134. [Google Scholar] [CrossRef]
- Dharmappa, P.M.; Doddaraju, P.; Malagondanahalli, M.V.; Rangappa, R.B.; Mallikarjuna, N.M.; Rajendrareddy, S.H.; Ramanjinappa, R.; Mavinahalli, R.P.; Prasad, T.G.; Udayakumar, M.; et al. Introgression of root and water use efficiency traits enhances water productivity: An evidence for physiological breeding in rice (Oryza sativa L.). Rice 2019, 12, 14. [Google Scholar] [CrossRef]
- Caine, R.S.; Yin, X.; Sloan, J.; Harrison, E.L.; Mohammed, U.; Fulton, T.; Biswal, A.K.; Dionora, J.; Chater, C.C.; Coe, R.A.; et al. Rice with reduced stomatal density conserves water and has improved drought tolerance under future climate conditions. New Phytol. 2019, 221, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Urban, J.; Ingwers, M.W.; McGuire, M.A.; Teskey, R.O. Increase in leaf temperature opens stomata and decouples net photosynthesis from stomatal conductance in Pinus taeda and Populus deltoides x nigra. J. Exp. Bot. 2017, 68, 1757–1767. [Google Scholar] [CrossRef] [PubMed]
- Tombesi, S.; Nardini, A.; Frioni, T.; Soccolini, M.; Zadra, C.; Farinelli, D.; Poni, S.; Palliotti, A. Stomatal closure is induced by hydraulic signals and maintained by ABA in drought-stressed grapevine. Sci. Rep. 2015, 5, 12449. [Google Scholar] [CrossRef] [PubMed]
- Kholová, J.; Hash, C.T.; Kakkera, A.; Koová, M.; Vadez, V. Constitutive water-conserving mechanisms are correlated with the terminal drought tolerance of pearl millet [Pennisetum glaucum (L.) R. Br.]. J. Exp. Bot. 2010, 61, 369–377. [Google Scholar] [CrossRef] [Green Version]
- Ali, J.; Jewel, Z.A.; Mahender, A.; Anandan, A.; Hernandez, J.; Li, Z. Molecular genetics and breeding for nutrient use efficiency in rice. Int. J. Mol. Sci. 2018, 19, 1762. [Google Scholar] [CrossRef] [Green Version]
- Kitomi, Y.; Kanno, N.; Kawai, S.; Mizubayashi, T.; Fukuoka, S.; Uga, Y. QTLs underlying natural variation of root growth angle among rice cultivars with the same functional allele of deeper rooting 1. Rice 2015, 8, 16. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Xu, X.; Zhan, X.; Zhai, R.; Wu, W.; Shen, X.; Dai, G.; Cao, L.; Cheng, S. Identification of QRL7, a major quantitative trait locus associated with rice root length in hydroponic conditions. Breed. Sci. 2013, 63, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Obara, M.; Tamura, W.; Ebitani, T.; Yano, M.; Sato, T.; Yamaya, T. Fine-mapping of QRL6.1, a Major QTL for root length of rice seedlings grown under a wide range of NH4+ concentrations in hydroponic conditions. Theor. Appl. Genet. 2010, 121, 535–547. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Han, Y.; Liu, L.; Chen, Y.; Du, Y.; Zhang, J.; Sun, H.; Zhao, Q. QRT9, a quantitative trait locus controlling root thickness and root length in upland rice. J. Exp. Bot. 2015, 66, 2723–2732. [Google Scholar] [CrossRef] [Green Version]
- Catolos, M.; Sandhu, N.; Dixit, S.; Shamsudin, N.A.A.; Naredo, M.E.B.; McNally, K.L.; Henry, A.; Diaz, M.G.; Kumar, A. Genetic loci governing grain yield and root development under variable rice cultivation conditions. Front. Plant Sci. 2017, 8, 1763. [Google Scholar] [CrossRef] [Green Version]
- Dash, G.K.; Barik, M.; Debata, A.K.; Baig, M.J.; Swain, P. Identification of most important rice root morphological markers in response to contrasting moisture regimes under vegetative stage drought. Acta Physiol. Plant. 2017, 39, 8. [Google Scholar] [CrossRef]
- Lou, Q.; Chen, L.; Mei, H.; Wei, H.; Feng, F.; Wang, P.; Xia, H.; Li, T.; Luo, L. Quantitative trait locus mapping of deep rooting by linkage and association analysis in rice. J. Exp. Bot. 2015, 66, 4749–4757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uga, Y.; Kitomi, Y.; Yamamoto, E.; Kanno, N.; Kawai, S.; Mizubayashi, T.; Fukuoka, S. A QTL for Root growth angle on rice chromosome 7 is involved in the genetic pathway of deeper rooting 1. Rice 2015, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitomi, Y.; Nakao, E.; Kawai, S.; Kanno, N.; Ando, T.; Fukuoka, S.; Irie, K.; Uga, Y. Fine mapping of QUICK ROOTING 1 and 2, quantitative trait loci increasing root length in rice. G3 Genes Genomes Genet. 2018, 8, 727–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Guo, Z.; Lv, Y.; Cen, X.; Ding, X.; Wu, H.; Li, X.; Huang, J.; Xiong, L. Genetic control of the root system in rice under normal and drought stress conditions by genome-wide association study. PLoS Genet. 2017, 13, e1006889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corales, M.; Nguyen, N.T.A.; Abiko, T.; Mochizuki, T. Mapping quantitative trait loci for water uptake of rice under aerobic conditions. Plant Prod. Sci. 2020, 23, 436–451. [Google Scholar] [CrossRef]
- Sandhu, N.; Anitha Raman, K.; Torres, R.O.; Audebert, A.; Dardou, A.; Kumar, A.; Henry, A. Rice root architectural plasticity traits and genetic regions for adaptability to variable cultivation and stress conditions. Plant Physiol. 2016, 171, 2562–2576. [Google Scholar] [CrossRef] [Green Version]
- Uga, Y.; Okuno, K.; Yano, M. Dro1, a major QTL involved in deep rooting of rice under upland field conditions. J. Exp. Bot. 2011, 62, 2485–2494. [Google Scholar] [CrossRef] [Green Version]
- Qun, Z.; Ju, C.X.; Wang, Z.Q.; Zhang, H.; Liu, L.J.; Yang, J.C.; Zhang, J. Grain yield and water use efficiency of super rice under soil water deficit and alternate wetting and drying irrigation. J. Integr. Agric. 2017, 16, 1028–1043. [Google Scholar] [CrossRef]
- Thorup-Kristensen, K.; Halberg, N.; Nicolaisen, M.; Olesen, J.E.; Crews, T.E.; Hinsinger, P.; Kirkegaard, J.; Pierret, A.; Bodin Dresbøll, D. Digging deeper for agricultural resources, the value of deep rooting. Trends Plant Sci. 2020, 25, 406–417. [Google Scholar] [CrossRef] [Green Version]
- Gago, J.; Douthe, C.; Florez-Sarasa, I.; Escalona, J.M.; Galmes, J.; Fernie, A.R.; Flexas, J.; Medrano, H. Opportunities for improving leaf water use efficiency under climate change conditions. Plant Sci. 2014, 226, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Jagadish, S.V.K.; Murty, M.V.R.; Quick, W.P. Rice responses to rising temperatures—Challenges, perspectives and future directions. Plant Cell Environ. 2015, 38, 1686–1698. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, F.; Zhou, J.; Chen, F.; Wang, B.; Xie, X. Phytochrome B control of total leaf area and stomatal density affects drought tolerance in rice. Plant Mol. Biol. 2012, 78, 289–300. [Google Scholar] [CrossRef]
- Bertolino, L.T.; Caine, R.S.; Gray, J.E. Impact of stomatal density and morphology on water-use efficiency in a changing world. Front. Plant Sci. 2019, 10, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatfield, J.L.; Dold, C. Water-use efficiency: Advances and challenges in a changing climate. Front. Plant Sci. 2019, 10, 103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cal, A.J.; Sanciangco, M.; Rebolledo, M.C.; Luquet, D.; Torres, R.O.; McNally, K.L.; Henry, A. Leaf morphology, rather than plant water status, underlies genetic variation of rice leaf rolling under drought. Plant Cell Environ. 2019, 42, 1532–1544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siddiqui, M.N.; Léon, J.; Naz, A.A.; Ballvora, A. Genetics and genomics of root system variation in adaptation to drought stress in cereal crops. J. Exp. Bot. 2021, 72, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Ji, K.; Wang, Y.; Sun, W.; Lou, Q.; Mei, H.; Shen, S.; Chen, H. Drought-Responsive mechanisms in rice genotypes with contrasting drought tolerance during reproductive stage. J. Plant Physiol. 2012, 169, 336–344. [Google Scholar] [CrossRef]
- Lynch, J.P.; Chimungu, J.G.; Brown, K.M. Root anatomical phenes associated with water acquisition from drying soil: Targets for crop improvement. J. Exp. Bot. 2014, 65, 6155–6166. [Google Scholar] [CrossRef] [Green Version]
- Panda, S.; Majhi, P.K.; Anandan, A.; Mahender, A.; Veludandi, S.; Bastia, D.; Guttala, S.B.; Singh, S.K.; Saha, S.; Ali, J. Proofing direct-seeded rice with better root plasticity and architecture. Int. J. Mol. Sci. 2021, 22, 6058. [Google Scholar] [CrossRef]
- Mallikarjuna Swamy, B.P.; Kumar, A. Sustainable rice yield in water-short drought-prone environments: Conventional and molecular approaches. In Irrigation Systems and Practices in Challenging Environments; IntechOpen: London, UK, 2012. [Google Scholar] [CrossRef] [Green Version]
- Anandan, A.; Meher, J.; Sah, R.P.; Samantaray, S.; Parameswaran, C.; Panneerselvam, P.; Dash, S.K.; Swain, P.; Kartikeyan, P.; Annamalai, M.; et al. Enhancing input use efficiency in direct-seeded rice with classical and molecular breeding. In Rice Research for Enhancing Productivity, Profitability and Climate Resilience; Pathak, H., Nayak, A.K., Jena, M., Singh, O.N., Samal, P., Sharma, S.G., Eds.; ICAR-NRRI: Cuttack, Indian, 2018; pp. 73–89. ISBN 81-88409-04-09. [Google Scholar]
- Chen, X.; Zhang, J.; Chen, Y.; Li, Q.; Chen, F.; Yuan, L.; Mi, G. Changes in root size and distribution in relation to nitrogen accumulation during maize breeding in China. Plant Soil 2014, 374, 121–130. [Google Scholar] [CrossRef]
- Isaac, M.E.; Nimmo, V.; Gaudin, A.C.M.; Leptin, A.; Schmidt, J.E.; Kallenbach, C.M.; Martin, A.; Entz, M.; Carkner, M.; Rajcan, I.; et al. Crop domestication, root trait syndromes, and soil nutrient acquisition in organic agroecosystems: A systematic review. Front. Sustain. Food Syst. 2021, 5, 317. [Google Scholar] [CrossRef]
- Lynch, J.P. Root phenes that reduce the metabolic costs of soil exploration: Opportunities for 21st century agriculture. Plant Cell Environ. 2015, 38, 1775–1784. [Google Scholar] [CrossRef] [PubMed]
- Mathew, I.; Shimelis, H.; Mutema, M.; Clulow, A.; Zengeni, R.; Mbava, N.; Chaplot, V. Selection of wheat genotypes for biomass allocation to improve drought tolerance and carbon sequestration into soils. J. Agron. Crop Sci. 2019, 205, 385–400. [Google Scholar] [CrossRef]
- Lynch, J.P. Steep, cheap and deep: An ideotype to optimize water and n acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl. No. | Traits | Treatments | Improved Genotypes vs. Landraces | ||||
|---|---|---|---|---|---|---|---|
| WW | DS | ||||||
| df | p Value | df | p Value | df | p Value | ||
| 1 | Photosynthesis rate (An) | 97 | **** | 1 | ** | 1 | * |
| 2 | Transpiration rate (Tr) | 97 | **** | 1 | ** | 1 | * |
| 3 | Leaf-level water use efficiency (WUEi) | 97 | **** | 1 | ** | 1 | ** |
| 4 | Internal CO2 concentration (Ci) | 97 | **** | 1 | * | 1 | * |
| 5 | Shoot dry weight (SDW) | 97 | **** | 1 | * | 1 | * |
| 6 | Root dry weight (RDW) | 97 | **** | 1 | ** | 1 | ** |
| 7 | Ratio of deep rooting (RDR) | 97 | **** | 1 | * | 1 | * |
| 8 | Root number (RN) | 97 | **** | 1 | * | 1 | * |
| 9 | Root diameter (RD) | 97 | **** | 1 | * | 1 | * |
| Sl. No. | Genotypes | Type | SDW (g) | RDW (g) | RDR | RN | RD (mm) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WW | DS | PDR | WW | DS | PDR | WW | DS | WW | DS | WW | DS | |||
| 1 | CR Dhan 203 | IP | 40.6 | 23.9 | 41.2 | 3.89 | 2.86 | 26.5 | 10.3 | 6.5 | 84 | 77 | 0.425 | 0.325 |
| 2 | CR Dhan 206 | IP | 42.0 | 25.0 | 40.6 | 4.28 | 3.58 | 16.5 | 13.9 | 11.9 | 82 | 70 | 0.368 | 0.275 |
| 3 | CR Dhan 207 | IP | 43.7 | 26.6 | 39.1 | 3.89 | 3.45 | 11.3 | 16.4 | 13.0 | 79 | 66 | 0.425 | 0.308 |
| 4 | Vandana | IP | 45.0 | 22.5 | 50.1 | 3.12 | 2.53 | 19.1 | 12.4 | 7.9 | 81 | 62 | 0.345 | 0.186 |
| 5 | Shusk Samrat | IP | 44.4 | 20.3 | 54.2 | 3.54 | 2.93 | 17.4 | 13.2 | 7.5 | 73 | 58 | 0.369 | 0.226 |
| 6 | Barani Deep | IP | 37.3 | 13.1 | 64.9 | 1.93 | 2.13 | −10.4 | 17.5 | 14.0 | 94 | 77 | 0.347 | 0.197 |
| 7 | Pusa Sugandh-3 | IP | 53.3 | 22.6 | 57.6 | 3.72 | 2.16 | 41.9 | 14.3 | 9.2 | 78 | 75 | 0.345 | 0.166 |
| 8 | PR-113 | IP | 37.3 | 15.9 | 57.5 | 4.99 | 3.55 | 28.9 | 12.4 | 12.2 | 112 | 69 | 0.452 | 0.276 |
| 9 | Vivek Dhan-62 | IP | 49.2 | 20.0 | 59.3 | 4.23 | 4.18 | 1.2 | 13.5 | 12.8 | 96 | 82 | 0.389 | 0.134 |
| 10 | Pant Dhan-12 | IP | 53.7 | 16.0 | 70.2 | 3.66 | 2.56 | 30.2 | 15.1 | 6.8 | 75 | 91 | 0.412 | 0.158 |
| 11 | Annada | IP | 48.4 | 16.5 | 66.0 | 5.25 | 4.2 | 20.0 | 15.4 | 8.5 | 93 | 86 | 0.384 | 0.176 |
| 12 | Dhala Heera | IP | 31.5 | 16.1 | 48.9 | 5.30 | 4.28 | 19.3 | 15.6 | 11.5 | 89 | 78 | 0.389 | 0.149 |
| 13 | Heera | IP | 35.6 | 13.4 | 62.3 | 4.57 | 3.79 | 17.2 | 15.4 | 8.1 | 99 | 90 | 0.375 | 0.134 |
| 14 | Kalyani-2 | IP | 35.0 | 17.5 | 49.9 | 1.94 | 2.48 | −27.8 | 10.3 | 17.4 | 126 | 70 | 0.384 | 0.223 |
| 15 | Abhishek | IP | 28.1 | 9.5 | 66.0 | 3.13 | 2.75 | 12.1 | 13.1 | 13.9 | 111 | 73 | 0.365 | 0.205 |
| 16 | Pant Sugandh Dhan 15 | IP | 34.5 | 16.0 | 53.8 | 5.88 | 3.67 | 37.7 | 14.2 | 10.5 | 102 | 86 | 0.377 | 0.202 |
| 17 | PR-116 | IP | 34.8 | 11.8 | 66.1 | 5.52 | 3.99 | 27.8 | 12.6 | 7.2 | 114 | 96 | 0.374 | 0.266 |
| 18 | PR-120 | IP | 36.4 | 11.7 | 67.8 | 5.37 | 4.15 | 22.7 | 15.3 | 10.1 | 82 | 80 | 0.470 | 0.299 |
| 19 | Ranbir Basmati-1 | IP | 54.2 | 24.2 | 55.3 | 4.85 | 3.29 | 32.2 | 12.1 | 16.2 | 94 | 67 | 0.425 | 0.148 |
| 20 | Sattari | IP | 48.8 | 25.3 | 48.1 | 2.33 | 2.44 | −4.7 | 12.9 | 16.9 | 98 | 85 | 0.412 | 0.152 |
| 21 | Tara | IP | 30.9 | 14.3 | 53.8 | 5.44 | 4.04 | 25.7 | 10.2 | 8.7 | 127 | 85 | 0.421 | 0.149 |
| 22 | Vanaprava | IP | 45.4 | 15.7 | 65.5 | 3.74 | 2.95 | 21.3 | 11.9 | 12.1 | 100 | 59 | 0.423 | 0.146 |
| 23 | Hazari Dhan | IP | 38.1 | 14.8 | 61.2 | 6.19 | 4.16 | 32.9 | 15.8 | 11.0 | 72 | 79 | 0.418 | 0.181 |
| 24 | Sadabahar | IP | 33.6 | 12.3 | 63.4 | 4.25 | 1.98 | 53.4 | 15.1 | 6.0 | 95 | 79 | 0.471 | 0.175 |
| 25 | IR119 | IP | 47.5 | 21.1 | 55.6 | 1.82 | 2.05 | −12.4 | 10.4 | 16.6 | 108 | 51 | 0.402 | 0.184 |
| 26 | Panindar | IP | 47.2 | 24.3 | 48.4 | 3.92 | 3.69 | 5.9 | 13.6 | 9.7 | 82 | 87 | 0.389 | 0.256 |
| 27 | VL Dhan-08 | IP | 58.7 | 20.9 | 64.4 | 3.30 | 1.98 | 40.2 | 14.5 | 16.4 | 96 | 58 | 0.398 | 0.258 |
| 28 | IR36 | IP | 33.1 | 17.5 | 47.2 | 4.71 | 3.61 | 23.4 | 10.2 | 12.1 | 106 | 74 | 0.354 | 0.103 |
| 29 | Ranbir Basmati-2 | IP | 47.7 | 24.3 | 49.1 | 1.48 | 1.83 | −23.7 | 20.6 | 23.9 | 59 | 43 | 0.415 | 0.266 |
| 30 | Moroberkan | LR | 37.7 | 30.3 | 19.8 | 1.66 | 2.78 | −67.2 | 27.6 | 30.9 | 78 | 64 | 0.489 | 0.430 |
| 31 | AC 36722 | LR | 37.7 | 20.9 | 44.7 | 2.24 | 2.01 | 10.5 | 16.5 | 14.8 | 64 | 46 | 0.389 | 0.250 |
| 32 | AC 36723 | LR | 49.1 | 20.8 | 57.6 | 3.77 | 2.65 | 29.8 | 14.3 | 12.6 | 98 | 87 | 0.471 | 0.390 |
| 33 | AC 36732 | LR | 24.5 | 10.7 | 56.2 | 4.28 | 3.86 | 9.8 | 9.56 | 7.5 | 105 | 86 | 0.412 | 0.273 |
| 34 | AC 36741 | LR | 23.5 | 12.4 | 47.2 | 5.77 | 3.59 | 37.8 | 12.7 | 10.9 | 87 | 69 | 0.463 | 0.422 |
| 35 | AC 36726 | LR | 14.2 | 5.2 | 63.8 | 5.26 | 3.89 | 26.1 | 8.36 | 5.4 | 83 | 50 | 0.489 | 0.331 |
| 36 | AC 36735 | LR | 42.0 | 32.0 | 23.8 | 2.24 | 2.46 | −9.6 | 30.4 | 34.2 | 65 | 55 | 0.456 | 0.370 |
| 37 | AC 36754 | LR | 55.9 | 23.4 | 58.1 | 5.04 | 3.80 | 24.6 | 13.9 | 11.5 | 86 | 70 | 0.425 | 0.356 |
| 38 | AC 36750 | LR | 32.4 | 8.5 | 73.8 | 6.39 | 4.20 | 34.4 | 12.7 | 9.7 | 86 | 56 | 0.463 | 0.246 |
| 39 | AC 36734 | LR | 32.9 | 24.1 | 26.9 | 3.88 | 3.47 | 10.7 | 19.4 | 21.5 | 63 | 47 | 0.398 | 0.356 |
| 40 | AC 36727 | LR | 33.2 | 7.3 | 78.0 | 5.37 | 3.73 | 30.5 | 12.6 | 8.0 | 88 | 55 | 0.478 | 0.422 |
| 41 | AC 36762 | LR | 44.4 | 38.8 | 12.6 | 2.84 | 3.18 | −12.0 | 36.4 | 39.4 | 74 | 68 | 0.496 | 0.327 |
| 42 | AC 35716 | LR | 43.8 | 23.8 | 45.6 | 4.77 | 3.40 | 28.8 | 18.6 | 16.0 | 83 | 69 | 0.462 | 0.306 |
| 43 | AC 35717 | LR | 48.1 | 42.3 | 12.1 | 2.08 | 2.10 | −0.96 | 37.6 | 40.0 | 63 | 51 | 0.460 | 0.347 |
| 44 | AC 35719 | LR | 36.2 | 13.2 | 63.6 | 5.40 | 4.16 | 23.1 | 14.6 | 10.0 | 94 | 75 | 0.387 | 0.263 |
| 45 | AC 35729 | LR | 38.8 | 27.9 | 28.1 | 2.01 | 2.39 | −18.7 | 28.1 | 31.5 | 79 | 68 | 0.475 | 0.314 |
| 46 | AC 35730 | LR | 35.5 | 18.2 | 48.7 | 5.22 | 3.21 | 38.5 | 16.4 | 11.1 | 76 | 51 | 0.384 | 0.260 |
| 47 | AC 35731 | LR | 44.5 | 21.7 | 51.4 | 4.93 | 2.27 | 54.1 | 15.6 | 12.2 | 69 | 45 | 0.395 | 0.234 |
| 48 | AC 36738 | LR | 34.4 | 25.2 | 26.9 | 1.53 | 2.90 | −89.5 | 25.9 | 29.5 | 71 | 53 | 0.378 | 0.262 |
| 49 | AC 35736 | LR | 38.4 | 12.6 | 67.2 | 4.25 | 3.79 | 10.8 | 13.8 | 8.1 | 96 | 65 | 0.375 | 0.254 |
| CD 5% | 2.53 | 2.14 | 0.39 | 0.22 | 1.81 | 2.43 | 4.54 | 3.97 | 0.01 | 0.02 | ||||
| Sl. No. | Genotypes | Type | An | Ci | Tr | WUEi | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| WW | DS | WW | DS | WW | DS | WW | DS | |||
| 1 | CR Dhan 203 | IP | 22.8 | 11.3 | 244.3 | 317.2 | 7.86 | 2.94 | 2.90 | 3.83 |
| 2 | CR Dhan 206 | IP | 24.5 | 11.2 | 172.2 | 264.0 | 7.24 | 2.87 | 3.39 | 3.88 |
| 3 | CR Dhan 207 | IP | 24.7 | 10.9 | 252.0 | 302.5 | 8.25 | 4.12 | 2.99 | 2.65 |
| 4 | Vandana | IP | 26.6 | 12.6 | 134.5 | 307.1 | 7.22 | 2.66 | 3.68 | 4.75 |
| 5 | Shusk Samrat | IP | 22.1 | 10.7 | 155.1 | 289.8 | 6.75 | 3.51 | 3.27 | 3.05 |
| 6 | Barani Deep | IP | 25.2 | 11.3 | 232.3 | 271.9 | 9.23 | 4.03 | 2.73 | 2.80 |
| 7 | Pusa Sugandh-3 | IP | 27.7 | 12.9 | 244.3 | 299.9 | 8.95 | 4.44 | 3.10 | 2.90 |
| 8 | PR-113 | IP | 23.1 | 13.3 | 201.3 | 264.0 | 9.49 | 3.47 | 2.43 | 3.84 |
| 9 | Vivek Dhan-62 | IP | 19.2 | 9.4 | 241.1 | 275.4 | 9.50 | 3.48 | 2.02 | 2.69 |
| 10 | Pant Dhan-12 | IP | 22.0 | 12.5 | 247.7 | 306.9 | 10.21 | 3.74 | 2.15 | 3.33 |
| 11 | Annada | IP | 27.0 | 9.5 | 158.2 | 318.3 | 7.99 | 2.75 | 3.38 | 3.45 |
| 12 | Dhala Heera | IP | 17.9 | 8.5 | 200.1 | 312.2 | 6.62 | 2.72 | 2.70 | 3.13 |
| 13 | Heera | IP | 21.5 | 10.7 | 145.2 | 351.0 | 6.82 | 2.83 | 3.15 | 3.77 |
| 14 | Kalyani-2 | IP | 21.4 | 11.3 | 156.3 | 240.5 | 7.32 | 3.08 | 2.92 | 3.67 |
| 15 | Abhishek | IP | 20.0 | 11.1 | 213.5 | 258.7 | 7.17 | 3.62 | 2.78 | 3.06 |
| 16 | Pant Sugandh Dhan 15 | IP | 17.1 | 8.4 | 198.9 | 320.2 | 6.24 | 2.24 | 2.74 | 3.75 |
| 17 | PR-116 | IP | 23.1 | 9.0 | 204.9 | 340.8 | 7.77 | 3.36 | 2.98 | 2.69 |
| 18 | PR-120 | IP | 24.9 | 11.3 | 226.4 | 298.0 | 8.53 | 4.26 | 2.92 | 2.65 |
| 19 | Ranbir Basmati-1 | IP | 26.6 | 12.6 | 207.8 | 255.2 | 8.17 | 4.19 | 3.26 | 3.01 |
| 20 | Sattari | IP | 19.7 | 12.8 | 117.7 | 242.1 | 6.77 | 3.01 | 2.91 | 4.25 |
| 21 | Tara | IP | 27.9 | 11.1 | 139.1 | 286.9 | 7.67 | 2.52 | 3.64 | 4.39 |
| 22 | Vanaprava | IP | 20.2 | 10.5 | 161.7 | 279.5 | 8.52 | 2.62 | 2.37 | 4.01 |
| 23 | Hazari Dhan | IP | 19.8 | 11.2 | 207.0 | 278.1 | 7.47 | 2.32 | 2.65 | 4.84 |
| 24 | Sadabahar | IP | 21.9 | 8.6 | 130.7 | 316.7 | 7.20 | 3.76 | 3.04 | 2.29 |
| 25 | IR119 | IP | 18.7 | 10.4 | 208.3 | 261.3 | 6.70 | 2.89 | 2.79 | 3.59 |
| 26 | Panindar | IP | 26.3 | 13.4 | 175.9 | 288.7 | 7.64 | 3.31 | 3.44 | 4.04 |
| 27 | VL Dhan-08 | IP | 26.1 | 15.1 | 214.8 | 234.6 | 9.44 | 4.43 | 2.76 | 3.40 |
| 28 | IR36 | IP | 19.0 | 11.6 | 203.4 | 264.4 | 6.11 | 2.97 | 3.12 | 3.89 |
| 29 | Ranbir Basmati-2 | IP | 26.3 | 18.3 | 152.2 | 265.9 | 8.15 | 3.57 | 3.23 | 5.12 |
| 30 | Moroberkan | LR | 27.0 | 17.5 | 204.2 | 263.6 | 8.44 | 4.26 | 3.20 | 4.10 |
| 31 | AC 36722 | LR | 21.4 | 12.9 | 213.8 | 294.2 | 8.53 | 2.38 | 2.50 | 5.42 |
| 32 | AC 36723 | LR | 20.6 | 11.0 | 172.2 | 265.1 | 9.27 | 2.63 | 2.23 | 4.17 |
| 33 | AC 36732 | LR | 20.1 | 6.7 | 233.9 | 344.9 | 9.66 | 1.58 | 2.08 | 4.23 |
| 34 | AC 36741 | LR | 25.3 | 10.5 | 173.6 | 276.4 | 7.76 | 2.22 | 3.26 | 4.73 |
| 35 | AC 36726 | LR | 22.4 | 8.4 | 181.4 | 321.9 | 6.96 | 2.40 | 3.22 | 3.49 |
| 36 | AC 36735 | LR | 19.1 | 16.3 | 232.3 | 252.3 | 6.73 | 2.69 | 2.85 | 6.06 |
| 37 | AC 36754 | LR | 20.9 | 9.6 | 227.8 | 296.7 | 7.95 | 2.59 | 2.63 | 3.70 |
| 38 | AC 36750 | LR | 28.5 | 10.5 | 179.7 | 284.9 | 8.96 | 3.23 | 3.18 | 3.24 |
| 39 | AC 36734 | LR | 22.3 | 16.8 | 199.3 | 233.5 | 6.82 | 2.54 | 3.27 | 6.60 |
| 40 | AC 36727 | LR | 21.9 | 12.9 | 142.5 | 297.5 | 7.36 | 3.50 | 2.98 | 3.69 |
| 41 | AC 36762 | LR | 25.5 | 19.4 | 191.0 | 230.9 | 8.37 | 2.62 | 3.04 | 7.39 |
| 42 | AC 35716 | LR | 26.5 | 11.1 | 121.8 | 292.2 | 7.62 | 2.42 | 3.47 | 4.60 |
| 43 | AC 35717 | LR | 27.2 | 21.2 | 134.5 | 214.2 | 7.79 | 3.00 | 3.49 | 7.09 |
| 44 | AC 35719 | LR | 23.8 | 10.4 | 217.9 | 289.6 | 10.16 | 3.16 | 2.35 | 3.30 |
| 45 | AC 35729 | LR | 21.3 | 17.5 | 151.6 | 232.6 | 7.32 | 3.59 | 2.91 | 4.88 |
| 46 | AC 35730 | LR | 26.3 | 12.3 | 155.6 | 273.4 | 5.79 | 2.87 | 4.54 | 4.28 |
| 47 | AC 35731 | LR | 26.8 | 9.6 | 148.7 | 307.9 | 8.14 | 2.23 | 3.29 | 4.29 |
| 48 | AC 36738 | LR | 19.4 | 16.8 | 122.9 | 219.6 | 8.24 | 3.38 | 2.35 | 4.97 |
| 49 | AC 35736 | LR | 24.6 | 11.2 | 140.8 | 320.7 | 7.75 | 3.41 | 3.17 | 3.28 |
| CD 5% | 0.88 | 0.88 | 11.01 | 9.30 | 0.30 | 0.19 | 0.13 | 0.32 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gobu, R.; Dash, G.K.; Lal, J.P.; Swain, P.; Mahender, A.; Anandan, A.; Ali, J. Unlocking the Nexus between Leaf-Level Water Use Efficiency and Root Traits Together with Gas Exchange Measurements in Rice (Oryza sativa L.). Plants 2022, 11, 1270. https://doi.org/10.3390/plants11091270
Gobu R, Dash GK, Lal JP, Swain P, Mahender A, Anandan A, Ali J. Unlocking the Nexus between Leaf-Level Water Use Efficiency and Root Traits Together with Gas Exchange Measurements in Rice (Oryza sativa L.). Plants. 2022; 11(9):1270. https://doi.org/10.3390/plants11091270
Chicago/Turabian StyleGobu, Ramasamy, Goutam Kumar Dash, Jai Prakash Lal, Padmini Swain, Anumalla Mahender, Annamalai Anandan, and Jauhar Ali. 2022. "Unlocking the Nexus between Leaf-Level Water Use Efficiency and Root Traits Together with Gas Exchange Measurements in Rice (Oryza sativa L.)" Plants 11, no. 9: 1270. https://doi.org/10.3390/plants11091270
APA StyleGobu, R., Dash, G. K., Lal, J. P., Swain, P., Mahender, A., Anandan, A., & Ali, J. (2022). Unlocking the Nexus between Leaf-Level Water Use Efficiency and Root Traits Together with Gas Exchange Measurements in Rice (Oryza sativa L.). Plants, 11(9), 1270. https://doi.org/10.3390/plants11091270